When Three Isn’t a Crowd: A Digyny Concept for Treatment-Resistant, Near-Triploid Human Cancers


 , ,
, ,
Abstract
:“All basic traits inherent in cancer cells are displayed in gametes and vice versa”Janis-Olgerts Erenpreiss [1]
1. Introduction
1.1. Glossary
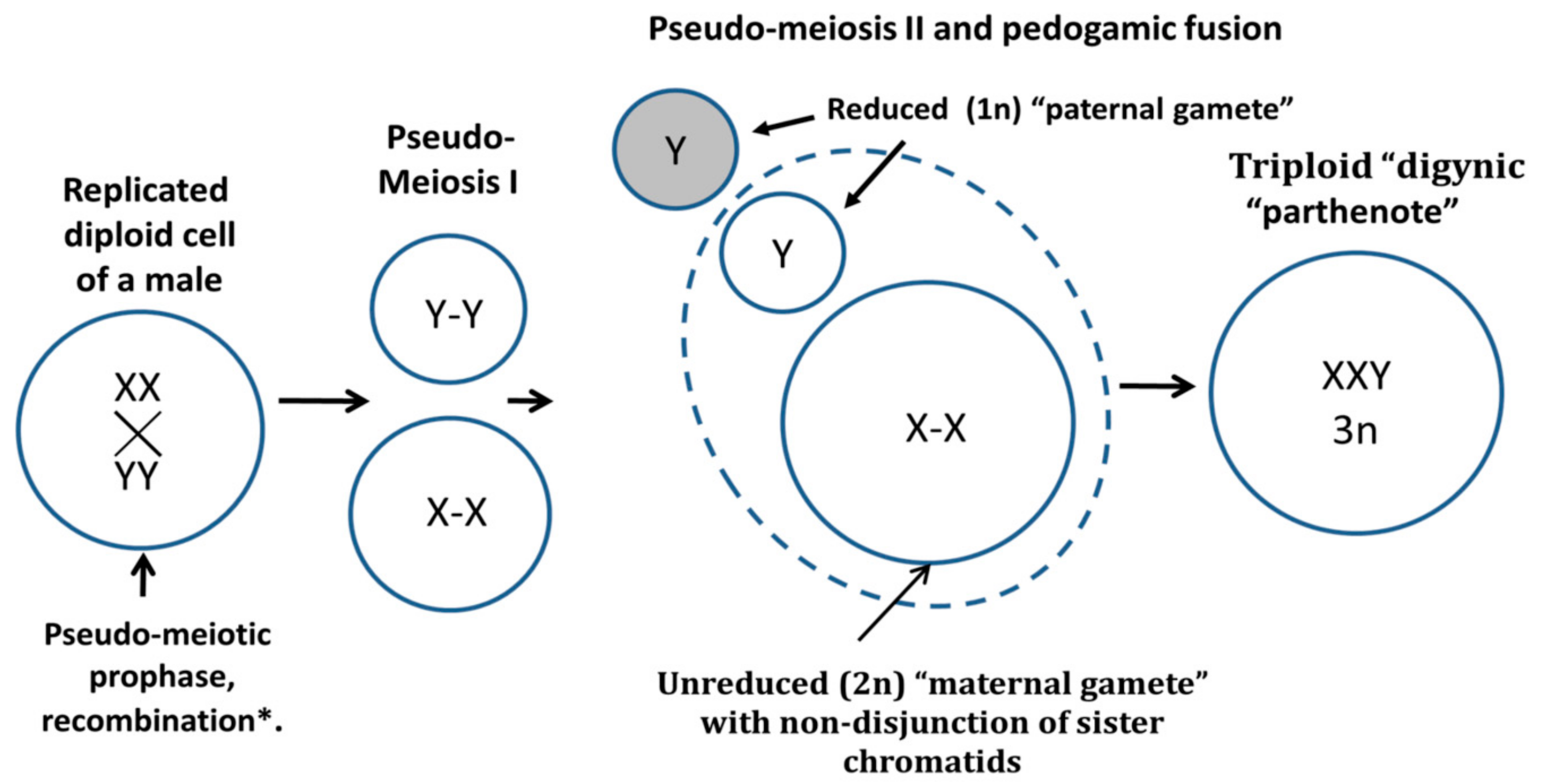
1.2. Digyny in Developmental Biology and Human Diploid-Triploid Mosaicism
1.3. Cancer Reprogramming To the Embryonality
1.4. Segregation of Haploid Genomes Is Coupled to Endoreduplication by Spindle Dysfunction
1.5. The Spindle Checkpoint Is Weak in the Preimplantation Embryo and Polyploidizing Tumor Cells
1.6. Digynic Zygotes in Human In Vitro Fertilization (IVF) Clinic
2. Materials and Methods
2.1. Patient Samples, Cell Lines, and Treatment
2.2. Fluorescent In Situ Hybridization (FISH)
2.3. DNA Image Cytometry
2.4. Immunofluorescence
2.5. In Silico Study of the Mitelman Database
3. Results
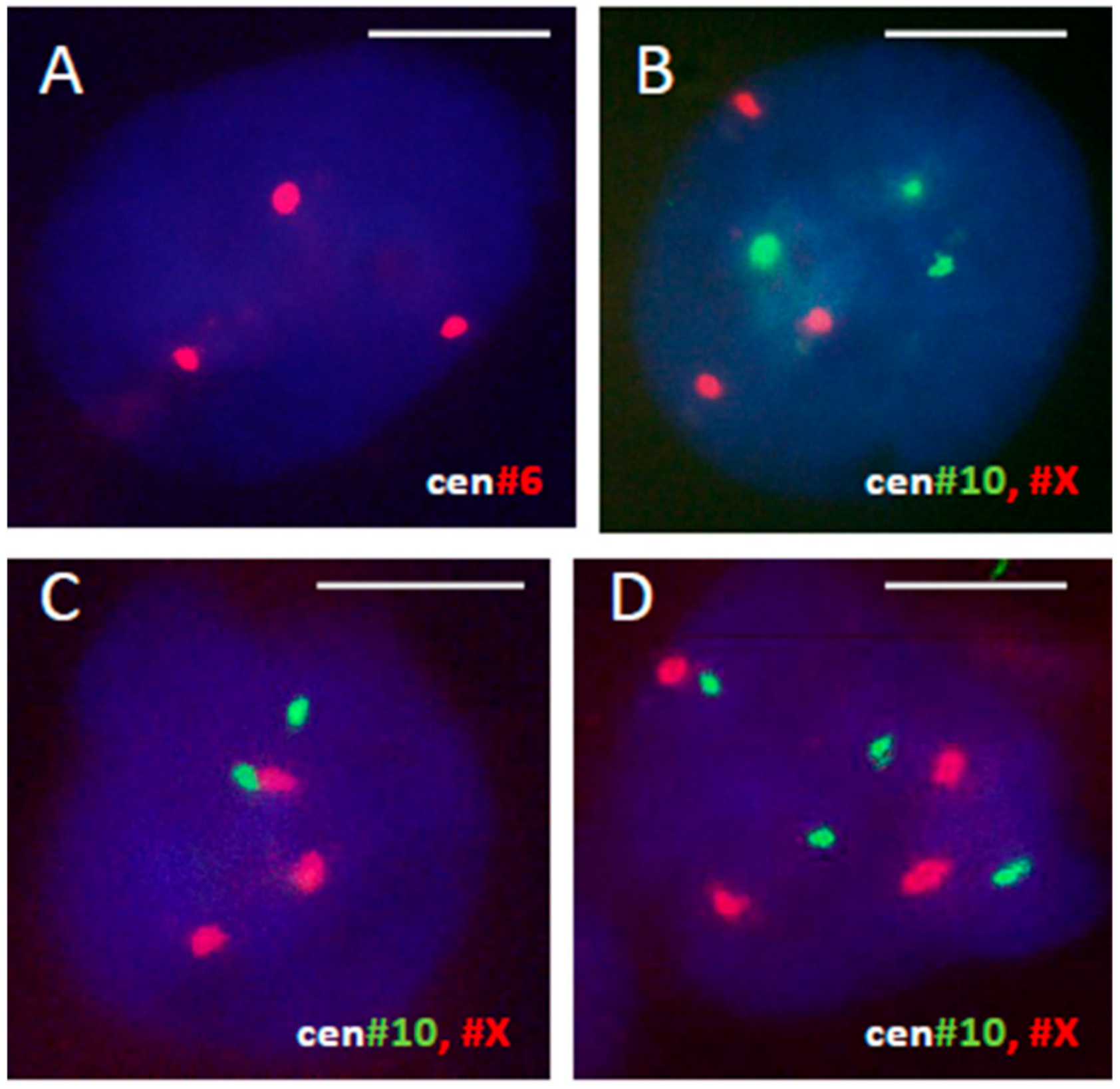
3.1. A Triploid Cell-Line May Coexist with Cycling Diploidy in Patient Tumors
3.1.1. Male Tumors (Renal Carcinoma)
3.1.2. Female Tumors (Breast Cancer)
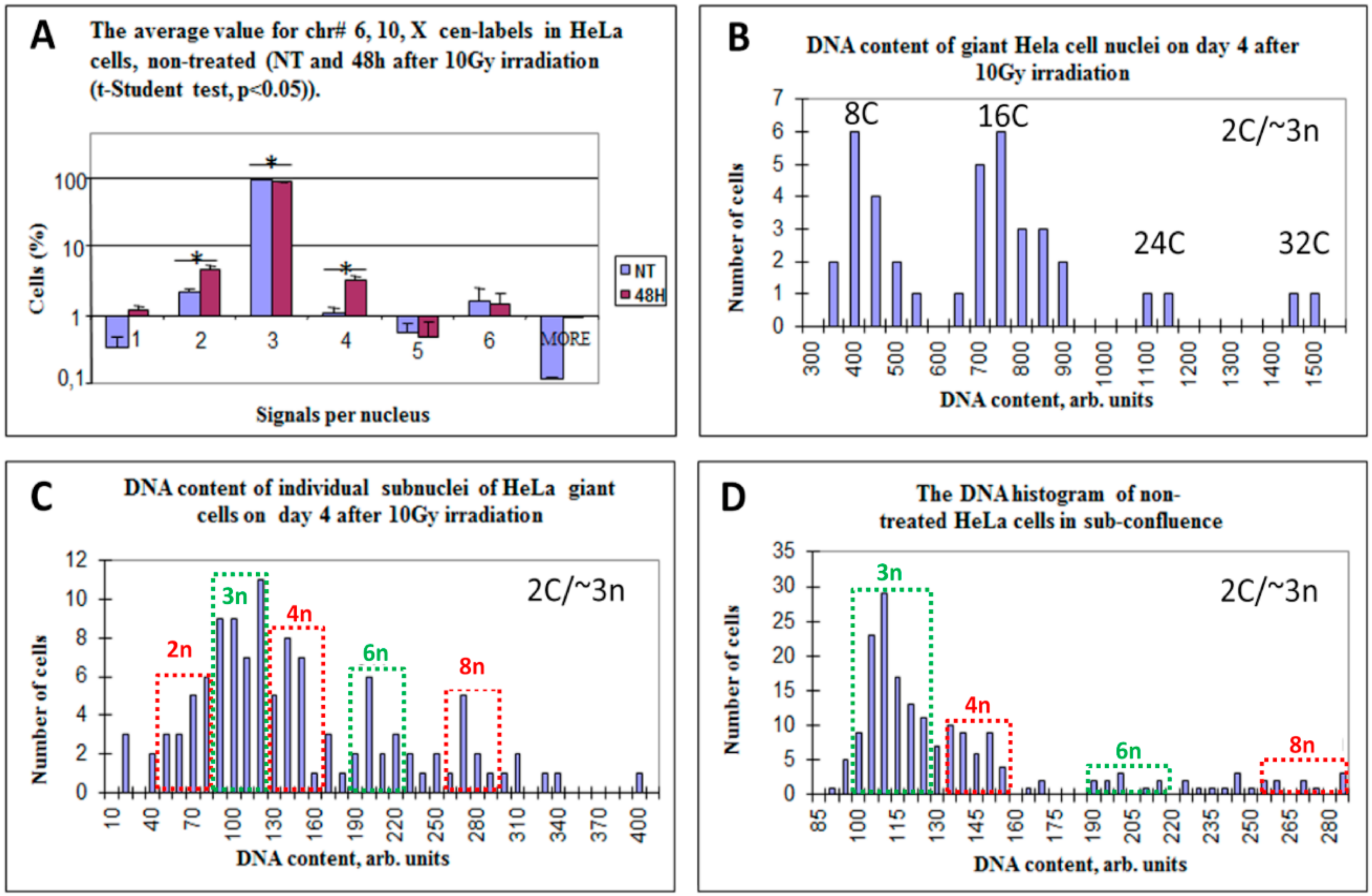
3.2. Diploid, Tetraploid, and Haploid Cell Nuclei Are Induced after Irradiation of Near-Triploid Cervical Carcinoma HeLa Cells
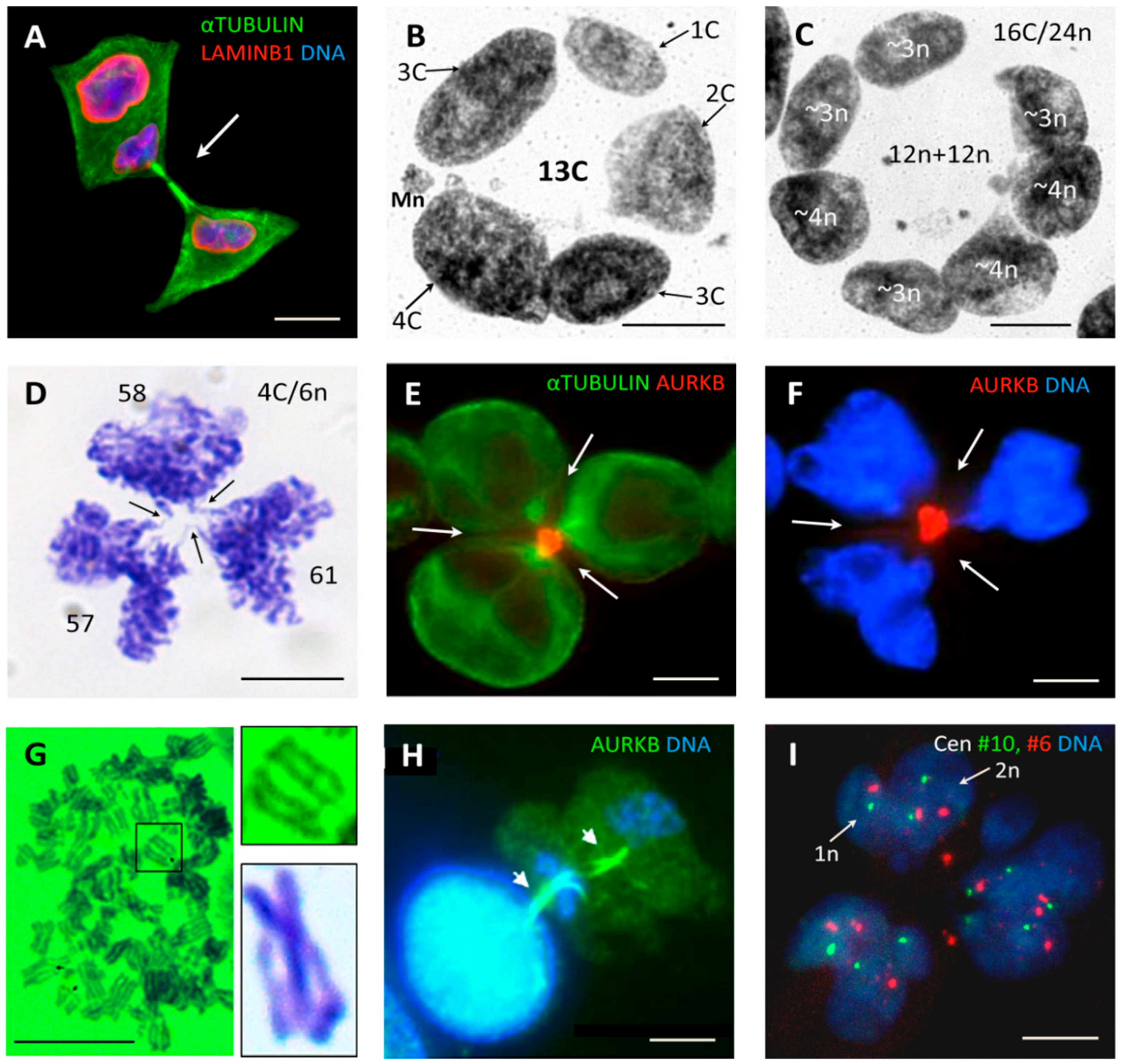
3.3. Multinucleated Giant Cells Contain DNA with Odd and Even Ploidy Numbers
3.4. Tripolar Mitosis of Endoreduplicated Cells May Convert Triploidy into Diploidy
3.5. Summary of Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Erenpreiss, J. Current Concepts of Malignant Growth. Part A: From a Normal Cell to Cancer; Zvaigzne: Riga, Latvia, 1993. [Google Scholar]
- Inhorn, S.L.; Therman, E.; Patau, K. Cytogenetic studies in spontaneous human abortion. Am. J. Clin. Pathol. 1964, 42, 528. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Sheltzer, J.M.; Amon, A. The aneuploidy paradox: Costs and benefits of an incorrect karyotype. Trends Genet. 2011, 27, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Holland, A.J.; Cleveland, D.W. Losing balance: The origin and impact of aneuploidy in cancer. EMBO Rep. 2012, 13, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Choi, N.; Kim, K.; Koo, H.-J.; Choi, J.; Kim, H.N. Chemoresistance of cancer cells: Requirements of tumor microenvironment-mimicking models in anti-cancer drug development. Theranostics 2018, 8, 5259–5275. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.T.; Pandis, N.; Heim, S. Tumours of the Breast. In Cancer Cytogenetics, 3rd ed.; Heim, S., Mitelman, F., Eds.; Willey: Hoboken, NJ, USA, 1995; pp. 493–516. [Google Scholar]
- Swanton, C.; Nicke, B.; Schuett, M.; Eklund, A.C.; Ng, C.; Li, Q.; Hardcastle, T.; Lee, A.; Roy, R.; East, P.; et al. Chromosomal instability determines taxane response. Proc. Natl. Acad. Sci. USA 2009, 106, 8671–8676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerashchenko, B.I.; Salmina, K.; Eglitis, J.; Huna, A.; Grjunberga, V.; Erenpreisa, J. Disentangling the aneuploidy and senescence paradoxes: A study of triploid breast cancers non-responsive to neoadjuvant therapy. Histochem. Cell Biol. 2016, 145, 497–508. [Google Scholar] [CrossRef]
- Vainshelbaum, N.M.; Zayakin, P.; Kleina, R.; Erenpreisa, J. Meta-analysis of cancer triploidy: Whole-genome rearrangements in male human tumours are characterised by XXY karyotypes. Genes 2019. accepted for publication. [Google Scholar]
- Mitelman, F.; Johansson, B.; Mertens, F. Mitelman Database of Chromosome Aberrations and Gene Fusions in Cancer. Available online: http://cgap.nci.nih.gov/Chromosomes/Mitelman (accessed on 1 January 2019).
- Salmina, K.; Huna, A.; Kalejs, M.; Pjanova, D.; Scherthan, H.; Cragg, M.S.; Erenpreisa, J. The cancer aneuploidy paradox: In the light of evolution. Genes 2019, 10, 83. [Google Scholar] [CrossRef]
- Giaretti, W.; Venesio, T.; Sciutto, A.; Prevosto, C.; Geido, E.; Risio, M. Near-diploid and near-triploid human sporadic colorectal adenocarcinomas differ for KRAS2 and TP53 mutational status. Genes Chromosomes Cancer 2003, 37, 207–213. [Google Scholar] [CrossRef]
- Kondrashov, A.S. The asexual ploidy cycle and the origin of sex. Nature 1994, 370, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Yant, L.; Bomblies, K. Genome management and mismanagement—Cell-level opportunities and challenges of whole-genome duplication. Genes Dev. 2015, 29, 2405–2419. [Google Scholar] [CrossRef] [PubMed]
- Tsang, M.; Gantchev, J.; Netchiporouk, E.; Moreau, L.; Ghazawi, F.M.; Glassman, S.; Sasseville, D.; Litvinov, I.V. A study of meiomitosis and novel pathways of genomic instability in cutaneous T-cell lymphomas (CTCL). Oncotarget 2018, 9, 37647. [Google Scholar] [CrossRef] [PubMed]
- Collins, U.K. Collins English Dictionary; Collins: Glasgow, Scotland, 2014; ISBN 9780007522743. [Google Scholar]
- Stevenson, A. Oxford Dictionary of English; Oxford University Press: New York, NY, USA, 2010; ISBN 9780199571123. [Google Scholar]
- Schinkel, C.C.F.; Kirchheimer, B.; Dullinger, S.; Geelen, D.; De Storme, N.; Hörandl, E. Pathways to polyploidy: Indications of a female triploid bridge in the alpine species Ranunculus kuepferi (Ranunculaceae). Plant Syst. Evol. 2017, 303, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Comai, L. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Sundaram, M.; Guernsey, D.L.; Rajaraman, M.M.; Rajaraman, R. Neosis: A novel type of cell division in cancer. Cancer Biol. Ther. 2004, 3, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Rajaraman, R.; Rajaraman, M.M.; Rajaraman, S.R.; Guernsey, D.L. Neosis—A paradigm of self-renewal in cancer. Cell Biol. Int. 2005, 29, 1084–1097. [Google Scholar] [CrossRef]
- Harlan, J.R.; deWet, J.M.J.; On, Ö. Winge and a Prayer: The origins of polyploidy. Bot. Rev. 1975, 41, 361–390. [Google Scholar] [CrossRef]
- Otto, S.P. The evolutionary consequences of polyploidy. Cell 2007, 131, 452–462. [Google Scholar] [CrossRef]
- Pawlowitzki, I.H.; Cenani, A. Sporadic triploid cells in human blood and fibroblast cultures. Humangenetik 1967, 5, 65–69. [Google Scholar] [CrossRef]
- Austin, C.R. Anomalies of fertilization leading to triploidy. J. Cell. Comp. Physiol. 1960, 56 (Suppl. 1), 1–15. [Google Scholar] [CrossRef]
- Neaves, W.B.; Baumann, P. Unisexual reproduction among vertebrates. Trends Genet. 2011, 27, 81–88. [Google Scholar] [CrossRef]
- Daniel, A.; Wu, Z.; Darmanian, A.; Collins, F.; Jackson, J. Three different origins for apparent triploid/diploid mosaics. Prenat. Diagn. 2003, 23, 529–534. [Google Scholar] [CrossRef]
- Mason, A.S.; Pires, J.C. Unreduced gametes: Meiotic mishap or evolutionary mechanism? Trends Genet. 2015, 31, 5–10. [Google Scholar] [CrossRef]
- Roeder, G.S. Meiotic chromosomes: It takes two to tango. Genes Dev. 1997, 11, 2600–2621. [Google Scholar] [CrossRef]
- Zhou, L.; Gui, J. Natural and artificial polyploids in aquaculture. Aquac. Fish. 2017, 2, 103–111. [Google Scholar] [CrossRef]
- Sell, S.; Nicolini, A.; Ferrari, P.; Biava, P.M. Cancer: A problem of developmental biology; scientific evidence for reprogramming and differentiation therapy. Curr. Drug Targets 2016, 17, 1103–1110. [Google Scholar] [CrossRef]
- Yeom, Y.I.; Fuhrmann, G.; Ovitt, C.E.; Brehm, A.; Ohbo, K.; Gross, M.; Hübner, K.; Schöler, H.R. Germline regulatory element of Oct-4 specific for the totipotent cycle of embryonal cells. Development 1996, 122, 881–894. [Google Scholar]
- Salmina, K.; Jankevics, E.; Huna, A.; Perminov, D.; Radovica, I.; Klymenko, T.; Ivanov, A.; Jascenko, E.; Scherthan, H.; Cragg, M.; et al. Up-regulation of the embryonic self-renewal network through reversible polyploidy in irradiated p53-mutant tumour cells. Exp. Cell Res. 2010, 316, 2099–2112. [Google Scholar] [CrossRef]
- Erenpreisa, J.; Salmina, K.; Huna, A.; Jackson, T.R.; Vazquez-Martin, A.; Cragg, M.S. The “virgin birth”, polyploidy, and the origin of cancer. Oncoscience 2015, 2, 3–14. [Google Scholar] [CrossRef]
- Niu, N.; Mercado-Uribe, I.; Liu, J. Dedifferentiation into blastomere-like cancer stem cells via formation of polyploid giant cancer cells. Oncogene 2017, 36, 4887–4900. [Google Scholar] [CrossRef] [Green Version]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef]
- Shaffer, S.M.; Dunagin, M.C.; Torborg, S.R.; Torre, E.A.; Emert, B.; Krepler, C.; Beqiri, M.; Sproesser, K.; Brafford, P.A.; Xiao, M.; et al. Rare cell variability and drug-induced reprogramming as a mode of cancer drug resistance. Nature 2017, 546, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Gao, R.; Sei, E.; Brandt, R.; Hartman, J.; Hatschek, T.; Crosetto, N.; Foukakis, T.; Navin, N.E. Chemoresistance evolution in triple-negative breast cancer delineated by single-cell sequencing. Cell 2018, 173, 879–893. [Google Scholar] [CrossRef]
- Cohnheim, J. Vorlesungen uber Allgemeine Pathologie. Ein Handbuch fur Arzte und Studierende; Hirschwald: Berlin, Germany, 1880. [Google Scholar]
- Barry Pierce, G.; Johnson, L.D. Differentiation and cancer. In Vitro 1971, 7, 140–145. [Google Scholar] [CrossRef]
- Pierce, G.B. The cancer cell and its control by the embryo. Rous-Whipple Award lecture. Am. J. Pathol. 1983, 113, 117–124. [Google Scholar]
- Virchow, R.; Chance, F.; Goodsir, J.; Osborn, S.; King’s College London; Pathological Institute of Berlin; St. Thomas’s Hospital. Cellular Pathology as Based upon Physiological and Pathological Histology; Twenty Lectures Delivered in the Pathological Institute of Berlin during the Months of February, March, and April; John Churchill: London, UK, 1860. [Google Scholar]
- Ērenpreisa, J.; Dālmane, A.; Ērenpreiss, J. Jānis Oļģerts Ērenpreiss and his theory of carcinogenesis. Acta Med. Hist. Rigensia Riga 2000, 25, 281–285. [Google Scholar] [CrossRef]
- Vinnitsky, V. The development of a malignant tumor is due to a desperate asexual self-cloning process in which cancer stem cells develop the ability to mimic the genetic program of germline cells. Intrinsically Disord. Proteins 2014, 2, e29997. [Google Scholar] [CrossRef] [Green Version]
- Cofre, J.; Abdelhay, E. Cancer is to embryology as mutation is to genetics: Hypothesis of the cancer as embryological phenomenon. Sci. World J. 2017, 2017, 3578090. [Google Scholar] [CrossRef]
- Chen, J.; Niu, N.; Zhang, J.; Qi, L.; Shen, W.; Donkena, K.V.; Feng, Z.; Liu, J. Polyploid giant cancer cells (PGCCs): The evil roots of cancer. Curr. Cancer Drug Targets 2019, 19, 360–367. [Google Scholar] [CrossRef]
- Erenpreisa, J.; Cragg, M.S. Cancer: A matter of life cycle? Cell Biol. Int. 2007, 31, 1507–1510. [Google Scholar] [CrossRef]
- Buiķis, I.; Harju, L.; Freivalds, T. Origin of microcells in the human sarcoma cell line HT-1080. Anal. Cell. Pathol. 1999, 18, 73–85. [Google Scholar] [CrossRef]
- Zhang, S.; Mercado-Uribe, I.; Xing, Z.; Sun, B.; Kuang, J.; Liu, J. Generation of cancer stem-like cells through the formation of polyploid giant cancer cells. Oncogene 2014, 33, 116–128. [Google Scholar] [CrossRef]
- Kalejs, M.; Ivanov, A.; Plakhins, G.; Cragg, M.S.; Emzinsh, D.; Illidge, T.M.; Erenpreisa, J. Upregulation of meiosis-specific genes in lymphoma cell lines following genotoxic insult and induction of mitotic catastrophe. BMC Cancer 2006, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Erenpreisa, J.; Cragg, M.S.; Salmina, K.; Hausmann, M.; Scherthan, H. The role of meiotic cohesin REC8 in chromosome segregation in γ irradiation-induced endopolyploid tumour cells. Exp. Cell Res. 2009, 315, 2593–2603. [Google Scholar] [CrossRef]
- Ianzini, F.; Kosmacek, E.A.; Nelson, E.S.; Napoli, E.; Erenpreisa, J.; Kalejs, M.; Mackey, M.A. Activation of meiosis-specific genes is associated with depolyploidization of human tumor cells following radiation-induced mitotic catastrophe. Cancer Res. 2009, 69, 2296–2304. [Google Scholar] [CrossRef]
- Erenpreisa, J.; Kalejs, M.; Cragg, M.S. Mitotic catastrophe and endomitosis in tumour cells: An evolutionary key to a molecular solution. Cell Biol. Int. 2005, 29, 1012–1018. [Google Scholar] [CrossRef]
- Gorgoulis, V.G.; Zacharatos, P.; Mariatos, G.; Liloglou, T.; Kokotas, S.; Kastrinakis, N.; Kotsinas, A.; Athanasiou, A.; Foukas, P.; Zoumpourlis, V.; et al. Deregulated expression of c-mos in non-small cell lung carcinomas: Relationship with p53 status, genomic instability, and tumor kinetics. Cancer Res. 2001, 61, 538–549. [Google Scholar]
- Klymiuk, I.; Kenner, L.; Adler, T.; Busch, D.H.; Boersma, A.; Irmler, M.; Fridrich, B.; Gailus-Durner, V.; Fuchs, H.; Leitner, N.; et al. In vivo functional requirement of the mouse Ifitm1 gene for germ cell development, interferon mediated immune response and somitogenesis. PLoS ONE 2012, 7, e44609. [Google Scholar] [CrossRef]
- Grell, K.G. Die Chromosomen von Aulacantha scolymantha Haeckel. Arch. Protistenkd 1953, 99, 1–54. [Google Scholar]
- Raĭkov, I.B. The Protozoan Nucleus, Morphology and Evolution; Springer: Wien, Austria, 1982. [Google Scholar]
- Huskins, C.L. Chromosome multiplication and reduction in somatic tissues; their possible relation to differentiation, reversion and sex. Nature 1948, 161, 80–83. [Google Scholar] [CrossRef]
- Walen, K. Normal human cells acquiring proliferative advantage to hyperplasia-like growth-morphology: Aberrant progeny cells associated with endopolyploid and haploid divisions. Cancer Clin. Oncol. 2013, 2. [Google Scholar] [CrossRef]
- Walen, K.H. Neoplastic-like cell changes of normal fibroblast cells associated with evolutionary conserved maternal and paternal genomic autonomous behavior (Gonomery). J. Cancer Ther. 2014, 5, 860–877. [Google Scholar] [CrossRef]
- Delhanty, J.D.; Handyside, A.H. The origin of genetic defects in the human and their detection in the preimplantation embryo. Hum. Reprod. Update 1995, 1, 201–215. [Google Scholar] [CrossRef]
- Zernicka-Goetz, M.; Huang, S. Stochasticity versus determinism in development: A false dichotomy? Nat. Rev. Genet. 2010, 11, 743–744. [Google Scholar] [CrossRef]
- Vanneste, E.; Voet, T.; Le Caignec, C.; Ampe, M.; Konings, P.; Melotte, C.; Debrock, S.; Amyere, M.; Vikkula, M.; Schuit, F.; et al. Chromosome instability is common in human cleavage-stage embryos. Nat. Med. 2009, 15, 577–583. [Google Scholar] [CrossRef]
- Hansis, C. Analysis of Oct-4 expression and ploidy in individual human blastomeres. Mol. Hum. Reprod. 2001, 7, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Wolpert, L.; Beddington, R.; Jessell, T.; Lawrence, P.; Meyerowitz, E.; Smith, J. Principles of Development, 2nd ed.; Oxford University Press: Oxford, UK, 2002; Volume 77, p. 327. [Google Scholar]
- Mayer, W.; Smith, A.; Fundele, R.; Haaf, T. Spatial separation of parental genomes in preimplantation mouse embryos. J. Cell Biol. 2000, 148, 629–634. [Google Scholar] [CrossRef]
- Illidge, T.; Cragg, M.; Fringe, B.; Olive, P.; Erenpreisa, J. Polyploid giant cells provide a survival mechanism for p53 mutant cells after DNA damage. Cell Biol. Int. 2000, 24, 621–633. [Google Scholar] [CrossRef]
- Erenpreisa, J.A.; Cragg, M.S.; Fringes, B.; Sharakhov, I.; Illidge, T.M. Release of mitotic descendants by giant cells from irradiated Burkitt’s lymphoma cell line. Cell Biol. Int. 2000, 24, 635–648. [Google Scholar] [CrossRef]
- Erenpreisa, J.; Kalejs, M.; Ianzini, F.; Kosmacek, E.A.; Mackey, M.A.; Emzinsh, D.; Cragg, M.S.; Ivanov, A.; Illidge, T.M. Segregation of genomes in polyploid tumour cells following mitotic catastrophe. Cell Biol. Int. 2005, 29, 1005–1011. [Google Scholar] [CrossRef]
- Erenpreisa, J.; Cragg, M.S.; Anisimov, A.P.; Illidge, T.M. Tumor cell embryonality and the ploidy number 32n: Is it a developmental checkpoint? Cell Cycle 2011, 10, 1873–1874. [Google Scholar] [CrossRef] [Green Version]
- Weihua, Z.; Lin, Q.; Ramoth, A.J.; Fan, D.; Fidler, I.J. Formation of solid tumors by a single multinucleated cancer cell. Cancer 2011, 117, 4092–4099. [Google Scholar] [CrossRef] [Green Version]
- Mirzayans, R.; Andrais, B.; Murray, D. Roles of polyploid/multinucleated giant cancer cells in metastasis and disease relapse following anticancer treatment. Cancers 2018, 10, 118. [Google Scholar] [CrossRef]
- Rosenbusch, B.E. Mechanisms giving rise to triploid zygotes during assisted reproduction. Fertil. Steril. 2008, 90, 49–55. [Google Scholar] [CrossRef]
- Kalatova, B.; Jesenska, R.; Hlinka, D.; Dudas, M. Tripolar mitosis in human cells and embryos: Occurrence, pathophysiology and medical implications. Acta Histochem. 2015, 117, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Schwarz-Finsterle, J.; Scherthan, H.; Huna, A.; González, P.; Mueller, P.; Schmitt, E.; Erenpreisa, J.; Hausmann, M. Volume increase and spatial shifts of chromosome territories in nuclei of radiation-induced polyploidizing tumour cells. Mutat. Res. 2013, 756, 56–65. [Google Scholar] [CrossRef]
- Macville, M.; Schröck, E.; Padilla-Nash, H.; Keck, C.; Ghadimi, B.M.; Zimonjic, D.; Popescu, N.; Ried, T. Comprehensive and definitive molecular cytogenetic characterization of HeLa cells by spectral karyotyping. Cancer Res. 1999, 59, 141–150. [Google Scholar]
- Erenpreisa, J.; Ivanov, A.; Wheatley, S.P.; Kosmacek, E.A.; Ianzini, F.; Anisimov, A.P.; Mackey, M.; Davis, P.J.; Plakhins, G.; Illidge, T.M. Endopolyploidy in irradiated p53-deficient tumour cell lines: Persistence of cell division activity in giant cells expressing Aurora-B kinase. Cell Biol. Int. 2008, 32, 1044–1056. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Ghosh, I. Variation of stemline karyotype in a HeLa cell line. Z. Krebsforsch. Klin. Onkol. 1975, 84, 129–133. [Google Scholar] [CrossRef]
- Ianzini, F.; Mackey, M.A. Development of the large scale digital cell analysis system. Radiat. Prot. Dosim. 2002, 99, 289–293. [Google Scholar] [CrossRef]
- Erenpreisa, J.; Salmina, K.; Huna, A.; Kosmacek, E.A.; Cragg, M.S.; Ianzini, F.; Anisimov, A.P. Polyploid tumour cells elicit paradiploid progeny through depolyploidizing divisions and regulated autophagic degradation. Cell Biol. Int. 2011, 35, 687–695. [Google Scholar] [CrossRef]
- Kröger, C.; Afeyan, A.; Mraz, J.; Eaton, E.N.; Reinhardt, F.; Khodor, Y.L.; Thiru, P.; Bierie, B.; Ye, X.; Burge, C.B.; et al. Acquisition of a hybrid E/M state is essential for tumorigenicity of basal breast cancer cells. Proc. Natl. Acad. Sci. USA 2019, 116, 7353–7362. [Google Scholar] [CrossRef] [Green Version]
- Navin, N.E. The first five years of single-cell cancer genomics and beyond. Genome Res. 2015, 25, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Pera, F.; Schwarzacher, H.G. Die Verteilung der Chromosomen auf die Tochterzellkerne multipolarer Mitosen in euploiden Gewebekulturen von Microtus agrestis. Chromosoma 1969, 26, 337–354. [Google Scholar] [CrossRef]
- Palitti, F.; Rizzoni, M. Pattern of DNA segregation in multipolar anatelophases of different ploidy in euploid and aneuploid mammalian cells cultivated in vitro. Genetica 1972, 43, 130–147. [Google Scholar]
- Pera, F.; Rainer, B. Studies of multipolar mitoses in euploid tissue cultures. I. Somatic reduction to exactly haploid and triploid chromosome sets. Chromosoma (Berl.) 1973, 42, 71–86. [Google Scholar] [CrossRef]
- Rizzoni, M.; Palitti, F.; Perticone, P. Euploid segregation through multipolar mitosis in mammalian cell cultures. Identification of triploid, haploid, and segregating diploid cells in a diploid-euploid primary culture of rhesus kidney cells. Chromosoma 1974, 45, 151–162. [Google Scholar] [CrossRef]
- Pera, F.; Scholz, P. Polyploidization in vitro: Formation of a predominantly triploid cell population in an originally diploid tissue culture of Microtus agrestis. Hum. Genet. 1974, 21, 17–26. [Google Scholar] [CrossRef]
- Cho, H.; Noh, K.H.; Chung, J.-Y.; Takikita, M.; Chung, E.J.; Kim, B.W.; Hewitt, S.M.; Kim, T.W.; Kim, J.-H. Synaptonemal complex protein 3 is a prognostic marker in cervical cancer. PLoS ONE 2014, 9, e98712. [Google Scholar] [CrossRef]
- Vitale, I.; Senovilla, L.; Jemaà, M.; Michaud, M.; Galluzzi, L.; Kepp, O.; Nanty, L.; Criollo, A.; Rello-Varona, S.; Manic, G.; et al. Multipolar mitosis of tetraploid cells: Inhibition by p53 and dependency on Mos. EMBO J. 2010, 29, 1272–1284. [Google Scholar] [CrossRef]
- Stewenius, Y.; Gorunova, L.; Jonson, T.; Larsson, N.; Hoglund, M.; Mandahl, N.; Mertens, F.; Mitelman, F.; Gisselsson, D. Structural and numerical chromosome changes in colon cancer develop through telomere-mediated anaphase bridges, not through mitotic multipolarity. Proc. Natl. Acad. Sci. USA 2005, 102, 5541–5546. [Google Scholar] [CrossRef] [Green Version]
- Gisselsson, D.; Håkanson, U.; Stoller, P.; Marti, D.; Jin, Y.; Rosengren, A.H.; Stewénius, Y.; Kahl, F.; Panagopoulos, I. When the genome plays dice: Circumvention of the spindle assembly checkpoint and near-random chromosome segregation in multipolar cancer cell mitoses. PLoS ONE 2008, 3, e1871. [Google Scholar] [CrossRef]
- Gisselsson, D.; Jin, Y.; Lindgren, D.; Persson, J.; Gisselsson, L.; Hanks, S.; Sehic, D.; Mengelbier, L.H.; Øra, I.; Rahman, N.; et al. Generation of trisomies in cancer cells by multipolar mitosis and incomplete cytokinesis. Proc. Natl. Acad. Sci. USA 2010, 107, 20489–20493. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Yue, L.; Spradling, A.C. The Drosophila fusome, a germline-specific organelle, contains membrane skeletal proteins and functions in cyst formation. Development 1994, 120, 947–956. [Google Scholar]
- Ye, C.J.; Regan, S.; Liu, G.; Alemara, S.; Heng, H.H. Understanding aneuploidy in cancer through the lens of system inheritance, fuzzy inheritance and emergence of new genome systems. Mol. Cytogenet. 2018, 11, 31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
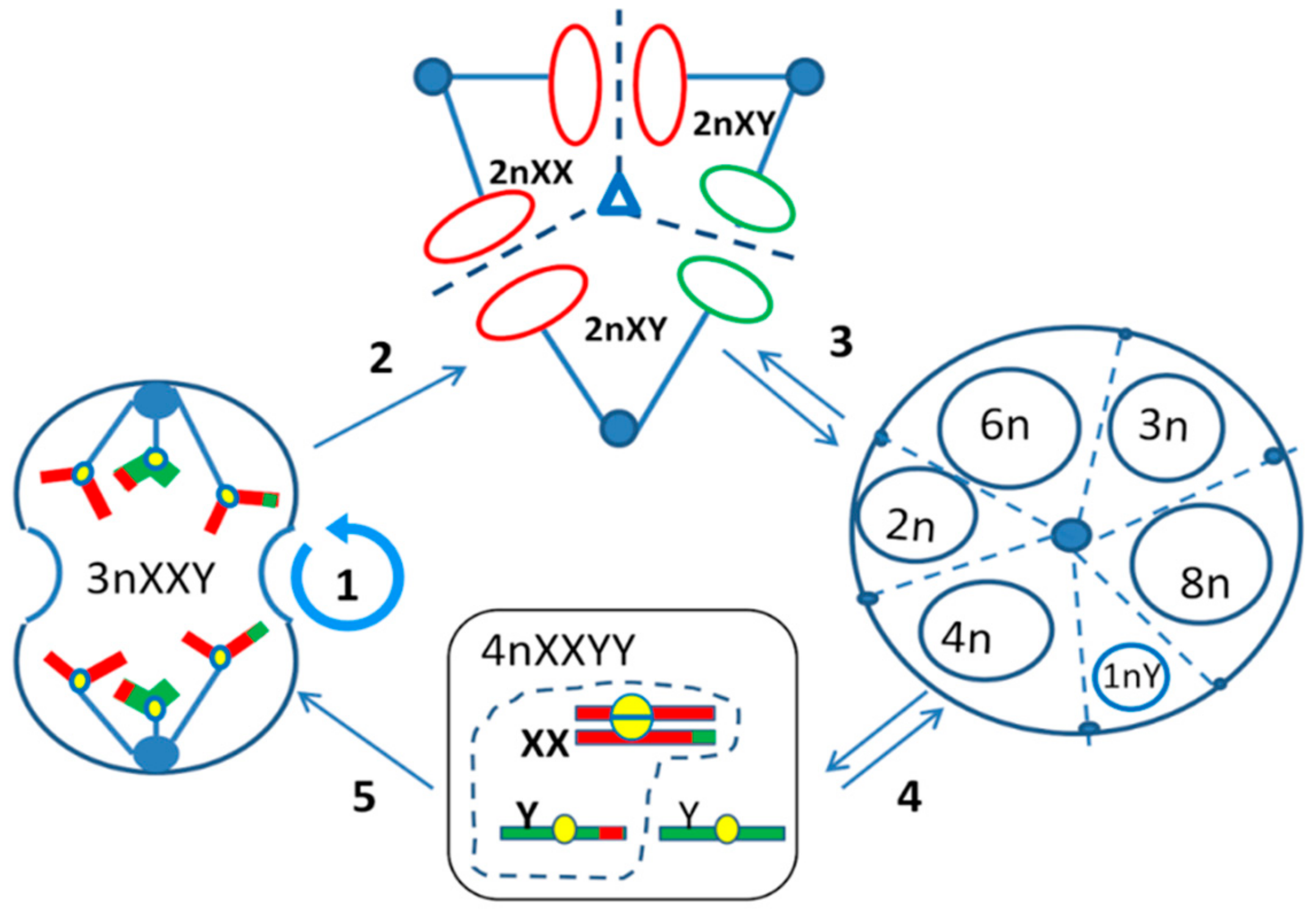{kind=link}
| Case | Chr1 | Sex1 | Chr2 | Sex2 | Chr3 | Sex3 | Chr4 | Sex4 |
|---|---|---|---|---|---|---|---|---|
| 1 | 70 | XXY | 46 | XY | 42 | XXY | 45 | XXY |
| 2 | 75 | XXY | 68 | XXY | 87 | XXXXYY | 48 | XY |
| 3 | 46 | XX, -Y | 45 | XX, -Y | 62 | XX, -Y | ||
| 4 | 74 | XXY | 46 | X, -Y | 46 | XY |
| Chromosome X | Chromosome 6 | Chromosome 10 | |||||
|---|---|---|---|---|---|---|---|
| Share of Cell Nuclei with a Certain Number of Centromere Signals | Number of Nuclei | % | Number of Nuclei | % | Number of Nuclei | % | |
| Diploid and tetraploid nuclei | NT | 19 | 3.1 | 17 | 3.1 | 23 | 3.4 |
| Triploid and other cell nuclei | NT | 586 | 96.9 | 523 | 96.9 | 649 | 96.6 |
| Diploid and tetraploid nuclei | 48h | 53 | 10.2 | 34 | 6.2 | 54 | 7.0 |
| Triploid and other cell nuclei | 48h | 466 | 89.8 | 518 | 93.8 | 714 | 93.0 |
| χ-squared test, p-value | 2.4·10−20 | 3.6·10−5 | 3.7·10−12 | ||||
| Malignant Tumor Type | Number of Karyotypes | XXYY (%) | X,−Y,+X (%) | YY,−X (or with Fragmented X) (%) |
|---|---|---|---|---|
| Seminoma | 78 | 12.82 | 2.56 | 0.00 |
| Osteosarcoma | 61 | 6.56 | 0 | 0.00 |
| Lung carcinoma | 237 | 2.95 | 0.84 | 0.00 |
| Gastric carcinoma | 74 | 0.00 | 4.05 | 0.00 |
| Head and neck squamous cell carcinoma | 191 | 3.14 | 0 | 0.00 |
| Colon adenocarcinoma | 98 | 3.06 | 3.06 | 0.00 |
| Transitional cell carcinoma | 104 | 0.96 | 0 | 0.00 |
| Chondrosarcoma | 85 | 5.88 | 0 | 0.00 |
| Malignant melanoma | 134 | 1.49 | 0.75 | 0.00 |
| Glioblastoma | 215 | 6.51 | 0 | 0.00 |
| Renal carcinoma | 577 | 1.39 | 0.87 | 0.17 |
| Mesothelioma | 72 | 5.56 | 0 | 0.00 |
| Rhabdomyosarcoma * | 92 | 30.43 | 3.26 | 1.09 |
| Ewing sarcoma | 228 | 3.51 | 0 | 0.00 |
| Liposarcoma | 147 | 3.40 | 0 | 0.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salmina, K.; Gerashchenko, B.I.; Hausmann, M.; Vainshelbaum, N.M.; Zayakin, P.; Erenpreiss, J.; Freivalds, T.; Cragg, M.S.; Erenpreisa, J. When Three Isn’t a Crowd: A Digyny Concept for Treatment-Resistant, Near-Triploid Human Cancers. Genes 2019, 10, 551. https://doi.org/10.3390/genes10070551
Salmina K, Gerashchenko BI, Hausmann M, Vainshelbaum NM, Zayakin P, Erenpreiss J, Freivalds T, Cragg MS, Erenpreisa J. When Three Isn’t a Crowd: A Digyny Concept for Treatment-Resistant, Near-Triploid Human Cancers. Genes. 2019; 10(7):551. https://doi.org/10.3390/genes10070551
Chicago/Turabian StyleSalmina, Kristine, Bogdan I. Gerashchenko, Michael Hausmann, Ninel M. Vainshelbaum, Pawel Zayakin, Juris Erenpreiss, Talivaldis Freivalds, Mark S. Cragg, and Jekaterina Erenpreisa. 2019. "When Three Isn’t a Crowd: A Digyny Concept for Treatment-Resistant, Near-Triploid Human Cancers" Genes 10, no. 7: 551. https://doi.org/10.3390/genes10070551