When Unity Is Strength: The Strategies Used by Chlamydomonas to Survive Environmental Stresses



1
Institut de Biologie Physico-Chimique, UMR 8226, CNRS, Sorbonne Université, 75005 Paris, France
2
Faculty of Sciences, Doctoral School of Plant Sciences, Université Paris-Sud, Paris-Saclay, 91400 Orsay, France
*
Author to whom correspondence should be addressed.
Cells 2019, 8(11), 1307; https://doi.org/10.3390/cells8111307
Submission received: 13 September 2019
/
Revised: 18 October 2019
/
Accepted: 21 October 2019
/
Published: 23 October 2019
(This article belongs to the Special Issue Chlamydomonas Cell Biology)
Abstract
:The unicellular green alga Chlamydomonas reinhardtii is a valuable model system to study a wide spectrum of scientific fields, including responses to environmental conditions. Most studies are performed under optimal growth conditions or under mild stress. However, when environmental conditions become harsher, the behavior of this unicellular alga is less well known. In this review we will show that despite being a unicellular organism, Chlamydomonas can survive very severe environmental conditions. To do so, and depending on the intensity of the stress, the strategies used by Chlamydomonas can range from acclimation to the formation of multicellular structures, or involve programmed cell death.
1. Introduction
The photosynthetic green alga Chlamydomonas reinhardtii is the most prominent model organism in the green algae lineage for both basic research and biotechnological applications. One of the current challenges for a better understanding of the functioning of this unicellular model organism is to understand how it adapts to hostile environmental conditions. While the different stress response strategies of multicellular photosynthetic organisms are now well known (e.g., programmed cell death (PCD), hypersensitive reaction, and autophagy) [1], much less is known for unicellular organisms like Chlamydomonas. Nevertheless, several reports have described the response to environmental stresses of Chlamydomonas, showing how this alga is able to adapt and trigger specific responses according to the intensity of the stress it faces. For instance, in Chlamydomonas, like in other photosynthetic organisms, environmental stresses most often result in the disturbance of photosynthesis and the production of reactive oxygen species (ROS) [2,3,4,5,6]. The first line of defense aims at containing the occurrence of this oxidative stress by decreasing ROS production and increasing ROS degradation using multiple strategies, such as non-photochemical quenching [7], alternative electron transport pathways [7,8], or the activation of antioxidant enzymes and pathways [6]. If these strategies are not sufficient to limit the impact of environmental stresses on the cell, Chlamydomonas will trigger alternate processes. The first strategy is acclimation which consists of building stronger defenses to protect the cell against future stress episodes. If acclimation does not provide sufficient protection to the cell, the socialization processes will be triggered, which ranges from the formation of multicellular structures to the outbreak of PCD. In all cases, interaction between Chlamydomonas cells will allow survival under severe stress conditions. In this review, we will describe the different strategies implemented by Chlamydomonas to survive environmental stresses.
2. Coping Strategies for Moderate Stress
2.1. Acclimation: Building Defenses to Protect Cells from Future Stresses
Acclimation is regarded as an environmentally-induced, short-term response leading to an improved tolerance to subsequent stresses [9]. In Chlamydomonas, acclimated cells are known to survive stress that would kill non-acclimated cells. Acclimation has been shown to be induced by several stresses, including ROS [10,11], UV-B [12], salt [13], and ionic stress [14]. Interestingly ROS, such as hydrogen peroxide (H2O2) or superoxide (1O2), induce specific acclimation pathways, and no cross acclimation between different ROS is observed [9,10]. Similarly, in plants, specific signaling pathways are associated with different ROS [15]. In Chlamydomonas, singlet oxygen acclimation is mediated by the key regulator SAK1 (singlet oxygen acclimation knocked-out 1) and the sak1 mutant is unable to acclimate to 1O2 [10]. SAK1 is a basic leucine zipper (bZIP) transcription factor that regulates the expression of important genes involved in acclimation, including glutathione peroxidase—GPXH, and glutathione S-transferase—GSTS1 [10]. SOR1 (Singlet Oxygen Resistant 1) is also a bZIP transcription factor, and the corresponding mutant, which is more resistant to 1O2, has higher levels of GPXH and GSTS1 transcripts [16].
It seems that at least some of the mechanisms controlling acclimation in Chlamydomonas are conserved in plants. For example UV-B acclimation in plants is mainly controlled by UVR8 (UV resistance locus 8) that interacts with COP1 (constitutively photomorphogenic 1) to induce a signaling pathway involving the bZIP transcription factors HY5 (elongated hypocotyl 5) and HYH (HY5 homolog) [17]. In Chlamydomonas, the roles of UVR8 and COP1 appear to be similar, suggesting a very early evolution of UV-B acclimation in photosynthetic organisms [12].
Acclimation is triggered in conditions of moderate stress that do not kill cells. While in the face of harsher stresses, Chlamydomonas will be able to enable a range of alternate strategies involving interaction between multiple cells and/or programmed cell death (Figure 1).
2.2. Protection by Degradation
There are two main degradative pathways to recycle cellular components in eukaryotic cells: the ubiquitin-proteasome system (UPS) and autophagy.
UPS is a degradation pathway that allows eukaryotic cells to control the abundance of numerous cellular proteins [18,19]. Ubiquitinated proteins are targeted for degradation by the proteasome, a multiproteic complex. In Chlamydomonas, UPS was found to be activated by stresses such as chloroplast damage [20], cold stress [21], copper [22], arsenate [23], metal nanoparticles [24], or selenite [25]. Selenite stress is associated with ROS production, which can inhibit UPS at a high concentration [25], as previously reported in mammals [26]. This could indicate that in the face of harsh stress conditions, UPS is not sufficient and more dramatic responses have to be triggered. In plants, UPS plays a prominent role in stress responses by degrading specific transcription factors [27,28]. It would be interesting to determine if such mechanisms are conserved in Chlamydomonas.
Autophagy is a catabolic membrane-trafficking process where cytosolic components, including proteins and membranes, are enclosed selectively or non-selectively within a double membrane vesicle, termed the autophagosome, and delivered to the vacuole or lysosome for degradation by resident hydrolases [29,30]. Autophagy usually occurs at a low, basal level under optimal growth and is activated upon stress in order to maintain cellular homeostasis. It is a pro-survival mechanism under stress conditions allowing cells to eliminate damaged cellular elements and recycle them into essential components [31]. Selective forms of autophagy exist in plants, such as chlorophagy [32] or mitophagy [33] but have not yet been reported in Chlamydomonas. Nevertheless, in Chlamydomonas, non-selective autophagy, i.e., macroautophagy (hereafter referred to as ‘autophagy’), which degrades cytosolic components, has been extensively studied during the last decade [34,35]. Autophagy is well conserved among eukaryotes and is mediated by AuTophaGy-related (ATG) genes [36]. The main role of ATG proteins is the formation and regulation of autophagosomes where damaged components will be imported. In Chlamydomonas, autophagy is strongly induced under different stress conditions, including nitrogen or carbon starvation [37,38], endoplasmic reticulum stress (accumulation of unfolded proteins) [39], the impairment of starch biosynthesis [40], metal exposure [41], high-light, and carotenoid deficiency [42]. The study of ATG mutants shows that autophagy is essential for cell survival, chlorophyll content, and starch accumulation during nutrient starvation [34]. ROS appears to play a major role in the control of autophagy in all organisms, including plants and algae; they have been proposed to constitute a link between the perception of stress and autophagy induction [43]. The intracellular redox state was found to control autophagosome formation and the small disulfide oxidoreductase thioredoxin plays an important role through regulation of the activity of the Atg4 protease in both Saccharomyces cerevisiae [44] and Chlamydomonas [45]. In addition, oxidative stress triggered by endoplasmic reticulum (ER) stress plays a role in the induction of autophagy [39]. Therefore UPS and autophagy are induced under moderate stress conditions, probably through ROS signaling, to eliminate toxic molecules and recycle nutrients that will allow survival until a return to optimal conditions. However, under extreme stress conditions, alternate processes are required to allow cell survival (Figure 1).
3. Socialization Allows for Better Resistance to Severe Stress
3.1. Multicellular Structures: Stronger Together
When confronted with harsh environmental stress conditions, adaptation processes are not sufficient to survive and Chlamydomonas cells interact to form two types of multicellular structures: palmelloids and aggregates (Figure 2). The name “palmelloid” comes from the resemblance of this structure with the alga Palmella [46]. Palmelloids are composed of 4 to 16 cells surrounded by a cell wall and are the result of successive divisions induced by stress without degradation of the cell wall.
In Chlamydomonas, stress induced palmelloid formation was initially reported by Iwasa and Murakami in the presence of organic acids [47]. Since then, a few studies have described palmelloid formation in response to a diversity of stresses (Table 1). Palmelloids can be induced by biotic and abiotic stresses in Chlamydomonas, and palmelloid-like structures have been reported in other green algae, such as Scenedesmus or Chlorella, suggesting that this mechanisms of stress response may be conserved in green algae [48,49].
In their natural environment, microalgae, as primary producers, are surrounded by grazers able to ingest algal cells [63]. A common defensive strategy employed by algae is to form colonies or palmelloids to exceed the upper size limit for ingestion. The rotifer predator Brachionus calyciflorus is indeed able to induce palmelloidation of Chlamydomonas within a day [50]. Palmelloid formation has also been observed under abiotic stress conditions (Table 1). During palmelloidation induced by NaCl, Khona and colleagues showed that inside the palmelloid, a stress dependent exopolysaccharides matrix is generated, probably for cell protection [55]. Inhibition of the destruction of the wall surrounding palmelloids may participate in their stabilization and could be mediated by inactivation of specific metalloproteinases (MMP) [55]. When conditions are favorable again, palmelloids dissociate rapidly; in less than an hour, and cells start to divide again (Figure 2) [55]. In case of prolonged or excessive stress, the cells remain in palmelloid form but will eventually die, through a process that may involve programmed cell death [54,55]. Proteomic analysis of the medium surrounding palmelloids indicates that the abundance of several hydroxyproline-rich glycoprotein (HPRG), such as pherophorins and MMPs, increases during palmelloid dissociation [55], suggesting that the medium could play an important regulatory role in this process. Interestingly, cells in palmelloids were found to contain more autophagic vesicles, suggesting that both autophagy and palmelloidation processes could be initiated concomitantly or that cells first induce acclimation before engaging in the path of palmelloidation [35].
In more severe conditions, the size of palmelloids may be too limited to allow survival and Chlamydomonas forms larger multicellular structures called aggregates or flocculates (hereafter referred to as “aggregates”). Palmelloids and aggregates are distinct structures. Aggregates are formed by a few tens, to several thousand cells, held together in an extracellular matrix, whereas palmelloids are composed of 4 to 16 cells surrounded by a cell wall (Figure 2).
Aggregation can be induced by predators, such as Peranema trichophorum (Euglenoidea), forming multicellular structures containing up to 100,000 Chlamydomonas cells [60]. Aggregated cells can secrete a mucous extracellular matrix (ECM) that may improve resistance to digestion [50,64]. Interestingly, strains of Chlamydomonas with smaller cells that are more easily engulfed by Peranema but are also slow-swimming, were found to form more aggregates compared to large fast-swimming strains [60].
The formation of aggregates may result either from agglutination of cells or from consecutive cell divisions without cell separation. When aggregation was induced in a mixed population of Chlamydomonas strains by the Peranema predator, the aggregates were found to be composed of cells from the different strains [60]. In addition, the mixing of a stained population with an unstained one also resulted in the formation of mixed aggregates [60]. This suggests that Peranema induces aggregation by the agglutination of cells. By contrast, aggregates of Chlamydomonas induced by a rotifer predator were formed only in the presence of light, suggesting that active growth may be required and that the aggregates result from consecutive divisions [50]. Therefore, multiple mechanisms of aggregate formation may exist. Unraveling these mechanisms will require more extensive cellular and molecular studies under a wide range of stresses.
Abiotic stresses, such as acidic or basic pH, can also induce aggregation (Table 1). Chlamydomonas culture medium is usually adjusted to pH 7. Basic pH is an efficient inducer of aggregation, especially above 10, where cations such as Ca2+ and Mg2+ form precipitates which are suggested to trigger aggregation [61]. Aggregation at low and high pH is correlated with a raise of the zeta potential [61]. The zeta potential characterizes the surface charge of a particle [65]. When aggregation is induced in Chlamydomonas by cationic cassia, a biopolymer, a shift of zeta potential is also observed [66]. It is, therefore, conceivable that by modifying the charge of the surface of Chlamydomonas cells, their ability to interact with each other would be modified and that a rise of the cell surface zeta potential might be important for aggregation.
In several studies, the mechanisms of aggregation in Chlamydomonas have been confronted with other species capable of forming multicellular structures. The self-flocculating yeast Saccharomyces bayanus was shown to induce Chlamydomonas aggregation, indicating that similarities in the aggregation process could exist between these two unicellular organisms [67]. In Saccharomyces, flocculation is mainly controlled by flocculin (FLO) genes [68,69] for which no homologs are present in Chlamydomonas genome [70]. Nevertheless, the expression of yeast FLO5 in Chlamydomonas was shown to induce aggregation [71]. FLO5 is a lectin, a family of calcium dependent glycoproteins. Interestingly, concanavalin A, a plant lectin, is also able to induce aggregation in Chlamydomonas [67], maybe through binding to the flagella of vegetative cells [72,73]. The role of flagella seems to be important, since during aggregation induced by a predator, cell mobility was impaired although the flagella remained intact and active [74], and strains lacking flagella aggregated much less in response to pH stress [61]. Altogether these results suggest that, as in yeasts and plants, lectins could also play a role in the control of aggregation in Chlamydomonas, maybe in connection with flagella. Twenty genes containing a lectin-C domain have been identified in the Chlamydomonas genome [75] but their possible role in controlling aggregation remains to be explored.
Volvox carteri, a multicellular organism, belonging like Chlamydomonas to the order of Volvocales, uses an ECM mainly composed of HPRG to hold cells together. The heterologous expression of Volvox Algal-CAM (Cell Adhesion Molecule) is able to induce aggregation in Chlamydomonas [76]. Algal-CAM is a HPRG exhibiting an N-terminal extensin-like domain and two fasciclin1 (FAS1) domains. FAS1 is known to mediate cell-cell adhesion in many organisms, such as bacteria, fungi, plants, and algae [77,78]. The Chlamydomonas genome encodes five proteins showing identity with Algal-CAM, called FAS2, FAS3, FAS4, FAS7, and a Periostin-related protein (Cre17.g745097) [70], whose role in the aggregation process remains to be explored.
More than twenty-five examples of transitions from uni to multi-cellular organization have been reported in diverse phylogenetic groups [79]. Volvocales, with their colonial organisms like Volvox or Gonium and unicellular relatives like Chlamydomonas, are excellent models to study the evolutionary pathways leading from unicellularity to multicellularity [80,81]. Selection of Chlamydomonas sedimented cells (multicellular structures) through multiple generations allowed researchers to detect de novo multicellular forms [82]. Predation has also been used to create de novo multicellular Chlamydomonas lineages. Using selection in the presence of a ciliate predator (Paramecium tetraurelia), Chlamydomonas’ multicellular structures appeared after roughly 750 generations [83]. Since multicellular structures can be generated in response to specific stress conditions, it is conceivable that there is a link between stress responses, evolution, and the transition from unicellularity to multicellularity [81,84].
3.2. Programmed Cell Death: Sucide for the Good of the Community
Programmed cell death was identified in animals in 1972 on the basis of morphological criteria [85], and over the years, additional cellular hallmarks were found to distinguish PCD from necrosis, the accidental cell death. Mitochondria were found to play a crucial role in the control of PCD, and signals emanating from this organelle (particularly cytochrome C and apoptotic protease activating factor-1) lead to irreversible destruction of the cell [29]. This process starts with the activation of caspases, a family of proteases specifically induced during PCD. Caspases are cysteine proteases cleaving proteins after an aspartate residue, the specificity being conferred by the three amino acids preceding the aspartate [86]. The specific activities of the different caspases can be measured using artificial peptides as substrates. These peptides are often coupled to fluorophores, allowing detection of caspase activities by measuring changes in the fluorescence signal. Caspases can be inhibited using the same peptides coupled to aldehyde (CHO, reversible inhibition) or methylketone radicals (CMK, FMK: irreversible inhibition). During PCD, several caspases are activated for degradation of specific targets, resulting in DNA and nucleus fragmentation, dismantling of the cell, and the formation of apoptotic bodies [87]. The specific degradation of DNA during PCD is detectable in situ by the TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling) reaction. It can also be visualized through gel electrophoresis by the specific appearance of a DNA ladder of 180 base pairs, the size of a nucleosome, whereas DNA degradation during necrosis is random and results in a smear [88].
Although it adopts variable forms, PCD exists in all multicellular organisms, where it is required for homeostasis, various developmental processes, and resistance to biotic or abiotic stresses [89]. If for a multicellular organism, it is clear that the disappearance of certain cells by PCD can be beneficial to the whole individual (to eliminate cells infected by a pathogen or abnormal cells for example), for a unicellular organism like Chlamydomonas, it seems paradoxical that PCD could exist, as it would result in the destruction of the organism itself. However, a few examples of PCD have been described in microorganisms, including, bacteria, yeasts, and microalgae [90,91,92]. In recent years, several teams have attempted to identify specific criteria for PCD in Chlamydomonas. In particular, it has been shown that during death induced by different stresses, caspase-like activities are induced and specific inhibitors are able to block these activities but also cell death [93,94,95]. Like land plants, Chlamydomonas does not appear to encode canonical caspases; however, other types of proteases have been shown to be responsible for caspase-like activities in plants [96,97]. Chlamydomonas contains two metacaspase genes, MC1 and MC2 [70]. In animals and plants, metacaspases are arginine and lysine-specific proteases which are involved in PCD induction [98,99]. This role might be conserved in Chlamydomonas, since MC genes were found to be induced during oxidative stress [100]. Therefore, the exact function of Chlamydomonas MC1 and MC2 will have to be clarified, for example, using knockout mutants.
The specific degradation of DNA during PCD has also been described in Chlamydomonas in response to different stresses, and could be detected by TUNEL or by the detection of a DNA ladder by electrophoresis [93,94,95,101,102,103]. Other markers of PCD have also been detected in Chlamydomonas, such as phosphatidylserine membrane translocation by annexin V [102] or fragmentation of poly(ADP-ribose) polymerase (PARP) [95]. Altogether these results suggest that a PCD-like process exists in Chlamydomonas, while the very reason for its existence remains to be established. Several studies have attempted to address this question by inducing PCD using heat shock [103] or UV-C [102]. In both cases the principle was to induce PCD in a culture, then to get rid of the cells to recover only the pre-conditioned “PCD” culture medium where fresh cells are inoculated, grown, and treated with the same stress that induced PCD in the first part of the experiment. The authors found that the cells placed in the PCD medium were more resistant than control cells [102,103]. Thus, the most exposed cells would be destroyed by PCD and could release into the medium, molecules capable of helping remaining cells of the population to better resist to death. This suggests that PCD should be considered as a pro-survival mechanism at the level of the whole population (Figure 1). These results also indicate that the culture medium has a fundamental role and allows cells to communicate to better adapt to their environment. The identification of molecules involved in cell to cell communications and enabling increased resistance to stress would be a major milestone for understanding this peculiar type of survival mechanism.
4. Cellular Reinforcement Strategies
4.1. The Cell Wall: The Last Defender
The cell wall plays an important role in the mechanisms allowing the protection of Chlamydomonas from environmental stresses. It is mainly composed of HPRGs [104], whose nature is likely to change according to environmental conditions. Consistently, cell wall-less strains are less resistant to metals such as cadmium, cobalt, copper, and nickel [105,106,107]. The cell wall has been reported to be modified by environmental conditions, including high-CO2, the presence of singlet oxygen, sulfur depravation, or an acidic pH [9,57,108,109]. High-CO2 cells produce different HPRGs and MMPs [108]; under sulfur starvation the profile of extracellular proteins is strongly modified [109,110]; and incubation at pH 3.4 induces significant thickening of the cell wall [57].
In nutrient-limiting conditions, cell wall thickening can lead to Daphnia resistance in Chlamydomonas [111]. Indeed, Chlamydomonas is able to resist digestion in the gut of its predator Daphnia, while cell wall-less strains are digested [111,112,113]. This may be crucial for survival of the population under nutrient-limiting conditions where the cell cycle is arrested, and therefore, every single life matters for the population to survive. In open-water ecosystems, a Chlamydomonas population could remain at equilibrium thanks to this mechanism of resistance to digestion [111,114]. It has also been proposed that the passage through the gut allows Chlamydomonas to take up nutrients from Daphnia [64]. A complete cell wall does not seem to be crucial for aggregation, as cellwall-less strains are also able to generate aggregates [61,76,115]. The role of the cell wall in the stress response has also been highlighted in cysts, where a thicker cell wall helps the cell to resist to extreme conditions.
In Chlamydomonas, strains used for genetic or reverse genetic experiments are usually cell-wall-deficient because of their higher transformation efficiencies [116]. Therefore special attention should be paid to studies on stress resistance using such strains.
4.2. Cyst: Surviving Extreme Conditions
If, like Charles Darwin, you were walking in the snows of the Andes mountains, European Alps, Antarctica, or Alaska, you might encounter the strange phenomenon of red snow [117,118]. This pink–red coloration of the snow is due to extremophile microalgae living in these hostile environments. The red snow appears in the spring/summer melting snows, at the onset of high intensities of sun radiation [119,120]. Most of snow algae are members of the Volvocales order [117]. This is a striking example of the ability of some photosynthetic microorganisms to survive under very harsh conditions, such as very low temperature, drought, starvation, and high radiation [121,122]. For instance, Chlamydomonas nivalis turns red in response to environmental stress, where it becomes a cyst (also named “aplanospore” or “hypnospore”) [120,121]. In Haematococcus pluvialis, similar encystement has been shown to be triggered by stress conditions [123], including nutrient deprivation [124], high light [125], salinity [126,127], or high or low temperatures [128]. The red pigmentation is mainly due to the accumulation of astaxanthin [129] which is produced from β-carotenes through the xanthophyll cycle using the beta-carotene ketolase (BKT) [130,131]. The very large amount of cytoplasmic astaxanthin esters acts like a filter that absorbs the excess of light that could lead to photoinhibition and photooxidative damages [121,122,132,133]. A homologue of the BKT gene exists in Chlamydomonas, although its expression level and the amount of astaxanthin are very low [134,135]; therefore, the link between BKT, astaxanthin, and stress responses, remains to be investigated. A first step may have been taken by Perozeni and colleagues, who managed to produce astaxanthin in Chlamydomonas reinhardtii by overexpressing a re-designed version of BKT [134].
4.3. Zygospore: When Sex Comes to the Rescue
In environmental stress conditions, Chlamydomonas haploid cells can differentiate into gametes which can mate to form diploid zygotes (Figure 2). In Chlamydomonas, gametogenesis have been shown to be triggered by nitrogen starvation and light variations [136], but in related organisms such as Volvox, ROS can also induce sexual behavior [137], suggesting that a greater variety of stresses could induce this differentiation in Chlamydomonas. The zygote-specific wall is better adapted to stress, as it is thicker and reinforced by isodityrosine cross-linking of proteins [138], and transglutamination linking lysine to glutamine [139]. Additionally, its cell wall is composed of different HPRGs [140,141], some of which are only present in the zygote, such as ZSP1 and ZSP2 (zygotic serine-proline rich) [142,143]. Recently, a giant type I polyketide synthase (PKS1) was shown to be involved in zygospore maturation [143]. The PKS1 gene is strongly overexpressed within a few days after the mating. The pks1 mutant does not survive desiccation but is still able to germinate, suggesting that PKS1 is rather involved in stress resistance of the zygospore rather than in mating. PKS1 is mandatory for the formation of knob-like structures on the cell surface and contributes to the construction of the cell wall central layer [143,144]. This cell wall organization allows the cells to survive hostile environments, such as darkness, desiccation, starvation, and freezing [143,145,146,147]. For instance, zygospores can allow survival during the winter and can germinate when temperatures increase to release haploid cells [146]. Moreover zygospores can aggregate into clumps (Figure 2) that are extremely difficult to break up [142,144,148], and could provide additional resistance to extreme environmental stresses.
5. Conclusions
A better knowledge of the mechanisms controlling stress responses in Chlamydomonas is important both for basic research and biotechnological applications. Fundamental research may help in understanding how unicellular algae are able to withstand very unfavorable environmental conditions. We have seen specific socialization mechanisms implied, for which next to nothing is known, although this process may be central in the transition from uni to multicellularity. It would be interesting to understand which genes and signals are involved in controlling the dialogue between cells that results in multicellular structure formation but also PCD. A major goal would be to identify the molecules released into the medium by dying cells to help other cells survive unfavorable conditions. These molecules could be of great interest for many fields and applications. Understanding stress responses in Chlamydomonas may allow engineering microalgae for improved growth under harsh industrial conditions or high light intensities, and thereby increase the productivity and economic viability of large-scale cultures of microalgae. Cell aggregation could also prove to be a very interesting process to develop alternate and cheap methods for harvesting microalgae [149,150].
Author Contributions
All authors contributed to the writing of the review.
Funding
This research and the APC were funded by Centre national de la Recherche Scientifique, Sorbonne Université, Université Paris-Saclay and Agence Nationale de la Recherche LABEX DYNAMO ANR-LABX-011.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dickman, M.; Williams, B.; Li, Y.; de Figueiredo, P.; Wolpert, T. Reassessing apoptosis in plants. Nat. Plants 2017, 3, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Ferretti, U.; Sedlářová, M.; Pospíšil, P. Singlet oxygen production in Chlamydomonas reinhardtii under heat stress. Sci. Rep. 2016, 6, 20094. [Google Scholar] [CrossRef]
- Du, Z.-Y.; Lucker, B.F.; Zienkiewicz, K.; Miller, T.E.; Zienkiewicz, A.; Sears, B.B.; Kramer, D.M.; Benning, C. Galactoglycerolipid Lipase PGD1 Is Involved in Thylakoid Membrane Remodeling in Response to Adverse Environmental Conditions in Chlamydomonas. Plant Cell 2018, 30, 447–465. [Google Scholar] [CrossRef]
- Nagy, V.; Vidal-Meireles, A.; Podmaniczki, A.; Szentmihályi, K.; Rákhely, G.; Zsigmond, L.; Kovács, L.; Tóth, S.Z. The mechanism of photosystem-II inactivation during sulphur deprivation-induced H2 production in Chlamydomonas reinhardtii. Plant J. 2018, 94, 548–561. [Google Scholar] [CrossRef]
- Stoiber, T.L.; Shafer, M.M.; Armstrong, D.E. Induction of reactive oxygen species in Chlamydomonas reinhardtii in response to contrasting trace metal exposures. Environ. Toxicol. 2013, 28, 516–523. [Google Scholar] [CrossRef]
- Laloi, C.; Apel, K.; Danon, A. Reactive oxygen signalling: The latest news. Curr. Opin. Plant Biol. 2004, 7, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Jokel, M.; Johnson, X.; Peltier, G.; Aro, E.-M.; Allahverdiyeva, Y. Hunting the main player enabling Chlamydomonas reinhardtii growth under fluctuating light. Plant J. 2018, 94, 822–835. [Google Scholar] [CrossRef] [PubMed]
- Ledford, H.K.; Chin, B.L.; Niyogi, K.K. Acclimation to singlet oxygen stress in Chlamydomonas reinhardtii. Eukaryot. Cell 2007, 6, 919–930. [Google Scholar] [CrossRef]
- Wakao, S.; Chin, B.L.; Ledford, H.K.; Dent, R.M.; Casero, D.; Pellegrini, M.; Merchant, S.S.; Niyogi, K.K. Phosphoprotein SAK1 is a regulator of acclimation to singlet oxygen in Chlamydomonas reinhardtii. eLife 2014, 3, e02286. [Google Scholar] [CrossRef]
- Fischer, B.B.; Eggen, R.I.; Niyogi, K.K. Characterization of singlet oxygen-accumulating mutants isolated in a screen for altered oxidative stress response in Chlamydomonas reinhardtii. BMC Plant. Biol. 2010, 10, 279. [Google Scholar] [CrossRef] [PubMed]
- Tilbrook, K.; Dubois, M.; Crocco, C.D.; Yin, R.; Chappuis, R.; Allorent, G.; Schmid-Siegert, E.; Goldschmidt-Clermont, M.; Ulm, R. UV-B Perception and Acclimation in Chlamydomonas reinhardtii. Plant Cell 2016, 28, 966–983. [Google Scholar] [CrossRef] [PubMed]
- Meijer, H.J.G.; van Himbergen, J.A.J.; Musgrave, A.; Munnik, T. Acclimation to salt modifies the activation of several osmotic stress-activated lipid signalling pathways in Chlamydomonas. Phytochemistry 2017, 135, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Charles, E.D.; Muhamadali, H.; Goodacre, R.; Pittman, J.K. Biochemical signatures of acclimation by Chlamydomonas reinhardtii to different ionic stresses. Algal Res. 2019, 37, 83–91. [Google Scholar] [CrossRef]
- Laloi, C.; Stachowiak, M.; Pers-Kamczyc, E.; Warzych, E.; Murgia, I.; Apel, K. Cross-talk between singlet oxygen- and hydrogen peroxide-dependent signaling of stress responses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 672–677. [Google Scholar] [CrossRef]
- Fischer, B.B.; Ledford, H.K.; Wakao, S.; Huang, S.G.; Casero, D.; Pellegrini, M.; Merchant, S.S.; Koller, A.; Eggen, R.I.L.; Niyogi, K.K. SINGLET OXYGEN RESISTANT 1 links reactive electrophile signaling to singlet oxygen acclimation in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2012, 109, E1302–E1311. [Google Scholar] [CrossRef]
- Liang, T.; Yang, Y.; Liu, H. Signal transduction mediated by the plant UV-B photoreceptor UVR8. New Phytol. 2019, 221, 1247–1252. [Google Scholar] [CrossRef]
- Budenholzer, L.; Cheng, C.L.; Li, Y.; Hochstrasser, M. Proteasome Structure and Assembly. J. Mol. Biol. 2017, 429, 3500–3524. [Google Scholar] [CrossRef]
- Smalle, J.; Vierstra, R.D. The Ubiquitin 26s Proteasome Proteolytic Pathway. Annu. Rev. Plant. Biol. 2004, 55, 555–590. [Google Scholar] [CrossRef]
- Heredia-Martínez, L.G.; Andrés-Garrido, A.; Martínez-Force, E.; Pérez-Pérez, M.E.; Crespo, J.L. Chloroplast Damage Induced by the Inhibition of Fatty Acid Synthesis Triggers Autophagy in Chlamydomonas. Plant. Physiol. 2018, 178, 1112–1129. [Google Scholar] [CrossRef]
- Valledor, L.; Furuhashi, T.; Hanak, A.-M.; Weckwerth, W. Systemic Cold Stress Adaptation of Chlamydomonas reinhardtii. Mol. Cell. Proteomics 2013, 12, 2032–2047. [Google Scholar] [CrossRef] [PubMed]
- Jamers, A.; Van der Ven, K.; Moens, L.; Robbens, J.; Potters, G.; Guisez, Y.; Blust, R.; De Coen, W. Effect of copper exposure on gene expression profiles in Chlamydomonas reinhardtii based on microarray analysis. Aquat. Toxicol. 2006, 80, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Chamari, W.; Harry, K.; Belinda, W.; Robert, W. Differential Proteome Analysis of Chlamydomonas reinhardtii Response to Arsenic Exposure. Am. J. Plant. Sci. 2012, 3, 764. [Google Scholar]
- Simon, D.F.; Domingos, R.F.; Hauser, C.; Hutchins, C.M.; Zerges, W.; Wilkinson, K.J. Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. Appl. Environ. Microbiol. 2013, 79, 4774–4785. [Google Scholar] [CrossRef] [PubMed]
- Vallentine, P.; Hung, C.-Y.; Xie, J.; Van Hoewyk, D. The ubiquitin–proteasome pathway protects Chlamydomonas reinhardtii against selenite toxicity, but is impaired as reactive oxygen species accumulate. AoB Plants 2014, 6, plu062. [Google Scholar] [CrossRef] [PubMed]
- Shang, F.; Taylor, A. Ubiquitin-proteasome pathway and cellular responses to oxidative stress. Free Radic. Biol. Med. 2011, 51, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.L. Role of the Ubiquitin Proteasome System in Plant Response to Abiotic Stress. In International Review of Cell and Molecular Biology; Galluzzi, L., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 343, pp. 65–110. [Google Scholar]
- Lyzenga, W.J.; Stone, S.L. Abiotic stress tolerance mediated by protein ubiquitination. J. Exp. Bot. 2012, 63, 599–616. [Google Scholar] [CrossRef]
- Wang, C.; Youle, R.J. The Role of Mitochondria in Apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [Green Version]
- Nakatogawa, H.; Suzuki, K.; Kamada, Y.; Ohsumi, Y. Dynamics and diversity in autophagy mechanisms: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 2009, 10, 458–467. [Google Scholar] [CrossRef]
- Avin-Wittenberg, T. Autophagy and its role in plant abiotic stress management. Plant. Cell Environ. 2019, 42, 1045–1053. [Google Scholar] [CrossRef]
- Izumi, M.; Ishida, H.; Nakamura, S.; Hidema, J. Entire Photodamaged Chloroplasts Are Transported to the Central Vacuole by Autophagy. Plant. Cell 2017, 29, 377–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broda, M.; Millar, A.H.; Van Aken, O. Mitophagy: A Mechanism for Plant Growth and Survival. Trends Plant. Sci. 2018, 23, 434–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajikawa, M.; Yamauchi, M.; Shinkawa, H.; Tanaka, M.; Hatano, K.; Nishimura, Y.; Kato, M.; Fukuzawa, H. Isolation and Characterization of Chlamydomonas Autophagy-Related Mutants in Nutrient-Deficient Conditions. Plant. Cell Physiol. 2019, 60, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, M.E.; Couso, I.; Heredia-Martínez, L.G.; Crespo, J.L. Monitoring Autophagy in the Model Green Microalga Chlamydomonas reinhardtii. Cells 2017, 6, 36. [Google Scholar] [CrossRef]
- Wesselborg, S.; Stork, B. Autophagy signal transduction by ATG proteins: From hierarchies to networks. Cell. Mol. Life Sci. 2015, 72, 4721–4757. [Google Scholar] [CrossRef]
- Pérez-Pérez, M.E.; Florencio, F.J.; Crespo, J.L. Inhibition of Target of Rapamycin Signaling and Stress Activate Autophagy in Chlamydomonas reinhardtii. Plant. Physiol. 2010, 152, 1874–1888. [Google Scholar] [CrossRef]
- Goodenough, U.; Blaby, I.; Casero, D.; Gallaher, S.D.; Goodson, C.; Johnson, S.; Lee, J.-H.; Merchant, S.S.; Pellegrini, M.; Roth, R.; et al. The Path to Triacylglyceride Obesity in the sta6 Strain of Chlamydomonas reinhardtii. Eukaryot. Cell 2014, 13, 591–613. [Google Scholar] [CrossRef]
- Pérez-Martín, M.; Pérez-Pérez, M.E.; Lemaire, S.D.; Crespo, J.L. Oxidative Stress Contributes to Autophagy Induction in Response to Endoplasmic Reticulum Stress in Chlamydomonas reinhardtii. Plant. Physiol. 2014, 166, 997–1008. [Google Scholar] [CrossRef]
- Tran, Q.-G.; Cho, K.; Park, S.-B.; Kim, U.; Lee, Y.J.; Kim, H.-S. Impairment of starch biosynthesis results in elevated oxidative stress and autophagy activity in Chlamydomonas reinhardtii. Sci. Rep. 2019, 9, 9856. [Google Scholar] [CrossRef]
- Pérez-Martín, M.; Blaby-Haas, C.E.; Pérez-Pérez, M.E.; Andrés-Garrido, A.; Blaby, I.K.; Merchant, S.S.; Crespo, J.L. Activation of Autophagy by Metals in Chlamydomonas reinhardtii. Eukaryot. Cell 2015, 14, 964–973. [Google Scholar] [CrossRef]
- Pérez-Pérez, M.E.; Couso, I.; Crespo, J.L. Carotenoid deficiency triggers autophagy in the model green alga Chlamydomonas reinhardtii. Autophagy 2012, 8, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, M.E.; Lemaire, S.D.; Crespo, J.L. Reactive Oxygen Species and Autophagy in Plants and Algae. Plant. Physiol. 2012, 160, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Pérez, M.E.; Zaffagnini, M.; Marchand, C.H.; Crespo, J.L.; Lemaire, S.D. The yeast autophagy protease Atg4 is regulated by thioredoxin. Autophagy 2014, 10, 1953–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Pérez, M.E.; Lemaire, S.D.; Crespo, J.L. Control of Autophagy in Chlamydomonas Is Mediated through Redox-Dependent Inactivation of the ATG4 Protease. Plant. Physiol. 2016, 172, 2219–2234. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.H. The Chlamydomonas Sourcebook; Academic Press: San Diego, CA, USA, 1989; ISBN 978-0-12-326880-8. [Google Scholar]
- Iwasa, K.; Murakami, S. Palmelloid Formation of Chlamydomonas I. Palmelloid Induction by Organic Acids. Physiol. Plant. 1968, 21, 1224–1233. [Google Scholar] [CrossRef]
- Pančić, M.; Kiørboe, T. Phytoplankton defence mechanisms: Traits and trade-offs. Biol. Rev. 2018, 93, 1269–1303. [Google Scholar] [CrossRef]
- Fisher, R.M.; Bell, T.; West, S.A. Multicellular group formation in response to predators in the alga Chlorella vulgaris. J. Evol. Biol. 2016, 29, 551–559. [Google Scholar] [CrossRef]
- Lurling, M.; Beekman, W. Palmelloids formation in Chlamydomonas reinhardtii: Defence against rotifer predators? Ann. Limnol. Int. J. Limnol. 2006, 42, 65–72. [Google Scholar] [CrossRef]
- Iwasa, K.; Murakami, S. Palmelloid Formation of Chlamydomonas II. Mechanism of Palmelloid Formation by Organic Acids. Physiol. Plant. 1969, 22, 43–50. [Google Scholar] [CrossRef]
- Olsen, Y.; Knutsen, G.; Lien, T. Characteristics of Phosphorus Limitation in Chlamydomonas reinhardtii (chlorophyceae) and Its Palmelloids. J. Phycol. 1983, 19, 313–319. [Google Scholar] [CrossRef]
- Samadani, M.; Dewez, D. Cadmium accumulation and toxicity affect the extracytoplasmic polyphosphate level in Chlamydomonas reinhardtii. Ecotoxicol. Environ. Saf. 2018, 166, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Takouridis, S.J.; Tribe, D.E.; Gras, S.L.; Martin, G.J.O. The selective breeding of the freshwater microalga Chlamydomonas reinhardtii for growth in salinity. Bioresour. Technol. 2015, 184, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Khona, D.K.; Shirolikar, S.M.; Gawde, K.K.; Hom, E.; Deodhar, M.A.; D’Souza, J.S. Characterization of salt stress-induced palmelloids in the green alga, Chlamydomonas reinhardtii. Algal Res. 2016, 16, 434–448. [Google Scholar] [CrossRef]
- Neelam, S.; Subramanyam, R. Alteration of photochemistry and protein degradation of photosystem II from Chlamydomonas reinhardtii under high salt grown cells. J. Photochem. Photobiol. B 2013, 124, 63–70. [Google Scholar] [CrossRef]
- Visviki, I.; Santikul, D. The pH tolerance of Chlamydomonas applanata (Volvocales, Chlorophyta). Arch. Environ. Contam. Toxicol. 2000, 38, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Bray, D.F.; Wagenaar, E.B. Ultrastructure of Chlamydomonas eugametos palmelloids induced by chloroplatinic acid treatment. J. Bacteriol. 1975, 121, 338–343. [Google Scholar]
- Nakamura, K.; Sakon, M.; Hatanaka, M.K. Chemical Factors Affecting Palmelloid-Forming Activity of Chloroplatinic Acid on Chlamydomonas eugametos. Physiol. Plant. 1976, 36, 293–296. [Google Scholar] [CrossRef]
- Sathe, S.; Durand, P.M. Cellular aggregation in Chlamydomonas (Chlorophyceae) is chimaeric and depends on traits like cell size and motility. Eur. J. Phycol. 2016, 51, 129–138. [Google Scholar] [CrossRef]
- Fan, J.; Zheng, L.; Bai, Y.; Saroussi, S.; Grossman, A.R. Flocculation of Chlamydomonas reinhardtii with Different Phenotypic Traits by Metal Cations and High pH. Front. Plant. Sci. 2017, 8, 1997. [Google Scholar] [CrossRef]
- Goff, K.L.; Headley, J.V.; Lawrence, J.R.; Wilson, K.E. Assessment of the effects of oil sands naphthenic acids on the growth and morphology of Chlamydomonas reinhardtii using microscopic and spectromicroscopic techniques. Sci. Total Environ. 2013, 442, 116–122. [Google Scholar] [CrossRef]
- Day, J.G.; Gong, Y.; Hu, Q. Microzooplanktonic grazers—A potentially devastating threat to the commercial success of microalgal mass culture. Algal Res. 2017, 27, 356–365. [Google Scholar] [CrossRef]
- Porter, K.G. Viable gut passage of gelatinous green algae ingested by Daphnia. SIL Proc. 1922–2010 1975, 19, 2840–2850. [Google Scholar] [CrossRef]
- Wilson, W.W.; Wade, M.M.; Holman, S.C.; Champlin, F.R. Status of methods for assessing bacterial cell surface charge properties based on zeta potential measurements. J. Microbiol. Methods 2001, 43, 153–164. [Google Scholar] [CrossRef]
- Banerjee, C.; Ghosh, S.; Sen, G.; Mishra, S.; Shukla, P.; Bandopadhyay, R. Study of algal biomass harvesting through cationic cassia gum, a natural plant based biopolymer. Bioresour. Technol. 2014, 151, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Santos, E.; Vila, M.; de la Vega, M.; León, R.; Vigara, J. Study of bioflocculation induced by Saccharomyces bayanus var. uvarum and flocculating protein factors in microalgae. Algal Res. 2015, 8, 23–29. [Google Scholar]
- Ogata, T.; Izumikawa, M.; Kohno, K.; Shibata, K. Chromosomal location of Lg-FLO1 in bottom-fermenting yeast and the FLO5 locus of industrial yeast. J. Appl. Microbiol. 2008, 105, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.V. Flocculation in Saccharomyces cerevisiae: A review. J. Appl. Microbiol. 2011, 110, 1–18. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas Genome Reveals the Evolution of Key Animal and Plant Functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef]
- Díaz-Santos, E.; Vila, M.; Vigara, J.; León, R. A new approach to express transgenes in microalgae and its use to increase the flocculation ability of Chlamydomonas reinhardtii. J. Appl. Phycol. 2016, 28, 1611–1621. [Google Scholar] [CrossRef]
- Musgrave, A.; van der Steuyt, P.; Ero, L. Concanavalin A binding to Chlamydomonas eugametos flagellar proteins and its effect on sexual reproduction. Planta 1979, 147, 51–56. [Google Scholar] [CrossRef]
- Millikin, B.E.; Weiss, R.L. Localization of concanavalin A binding carbohydrate in Chlamydomonas flagella. J. Cell Sci. 1984, 68, 211–226. [Google Scholar] [PubMed]
- Boyd, M.; Rosenzweig, F.; Herron, M.D. Analysis of motility in multicellular Chlamydomonas reinhardtii evolved under predation. PLoS ONE 2018, 13, e0192184. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, G.L.; Miranda-Saavedra, D.; Barton, G.J. Genome Analysis of the Unicellular Green Alga Chlamydomonas reinhardtii Indicates an Ancient Evolutionary Origin for Key Pattern Recognition and Cell-Signaling Protein Families. Genetics 2008, 179, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Lowder, L.G.; Herbert, S.K. Heterologous expression of a Volvox cell adhesion molecule causes flocculation in Chlamydomonas reinhardtii. J. Appl. Phycol. 2015, 27, 721–731. [Google Scholar] [CrossRef]
- Seifert, G.J. Fascinating Fasciclins: A Surprisingly Widespread Family of Proteins that Mediate Interactions between the Cell Exterior and the Cell Surface. Int. J. Mol. Sci. 2018, 19, 1628. [Google Scholar] [CrossRef]
- Johnson, K.L.; Jones, B.J.; Bacic, A.; Schultz, C.J. The Fasciclin-Like Arabinogalactan Proteins of Arabidopsis. A Multigene Family of Putative Cell Adhesion Molecules. Plant. Physiol. 2003, 133, 1911–1925. [Google Scholar] [CrossRef] [Green Version]
- Grosberg, R.K.; Strathmann, R.R. The Evolution of Multicellularity: A Minor Major Transition? Annu. Rev. Ecol. Evol. Syst. 2007, 38, 621–654. [Google Scholar] [CrossRef] [Green Version]
- Prochnik, S.E.; Umen, J.; Nedelcu, A.M.; Hallmann, A.; Miller, S.M.; Nishii, I.; Ferris, P.; Kuo, A.; Mitros, T.; Fritz-Laylin, L.K.; et al. Genomic Analysis of Organismal Complexity in the Multicellular Green Alga Volvox carteri. Science 2010, 329, 223–226. [Google Scholar] [CrossRef]
- Olson, B.J. From brief encounters to lifelong unions. eLife 2013, 2, e01893. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Herron, M.D.; Howell, K.; Pentz, J.T.; Rosenzweig, F.; Travisano, M. Experimental evolution of an alternating uni- and multicellular life cycle in Chlamydomonas reinhardtii. Nat. Commun. 2013, 4, 2742. [Google Scholar] [CrossRef]
- Herron, M.D.; Borin, J.M.; Boswell, J.C.; Walker, J.; Chen, I.-C.K.; Knox, C.A.; Boyd, M.; Rosenzweig, F.; Ratcliff, W.C. De novo origins of multicellularity in response to predation. Sci. Rep. 2019, 9, 2328. [Google Scholar] [CrossRef]
- Boraas, M.E.; Seale, D.B.; Boxhorn, J.E. Phagotrophy by a flagellate selects for colonial prey: A possible origin of multicellularity. Evol. Ecol. 1998, 12, 153–164. [Google Scholar] [CrossRef]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef]
- Earnshaw, W.C.; Martins, L.M.; Kaufmann, S.H. Mammalian Caspases: Structure, Activation, Substrates, and Functions During Apoptosis. Annu. Rev. Biochem. 1999, 68, 383–424. [Google Scholar] [CrossRef]
- He, B.; Lu, N.; Zhou, Z. Cellular and Nuclear Degradation during Apoptosis. Curr. Opin. Cell Biol. 2009, 21, 900–912. [Google Scholar] [CrossRef]
- Zhivotosky, B.; Orrenius, S. Assessment of apoptosis and necrosis by DNA fragmentation and morphological criteria. Curr. Protoc. Cell Biol. 2001, 12, 18.3.1–18.3.23. [Google Scholar]
- Fuchs, Y.; Steller, H. Programmed Cell Death in Animal Development and Disease. Cell 2011, 147, 742–758. [Google Scholar] [CrossRef] [Green Version]
- Bayles, K.W. Bacterial programmed cell death: Making sense of a paradox. Nat. Rev. Microbiol. 2014, 12, 63–69. [Google Scholar] [CrossRef]
- Bidle, K.D. Programmed Cell Death in Unicellular Phytoplankton. Curr. Biol. 2016, 26, R594–R607. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Gutierrez, D.; Eisenberg, T.; Büttner, S.; Meisinger, C.; Kroemer, G.; Madeo, F. Apoptosis in yeast: Triggers, pathways, subroutines. Cell Death Differ. 2010, 17, 763–773. [Google Scholar] [CrossRef]
- Sirisha, V.L.; Sinha, M.; D’Souza, J.S. Menadione-induced caspase-dependent programmed cell death in the green chlorophyte Chlamydomonas reinhardtii. J. Phycol. 2014, 50, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Yordanova, Z.P.; Woltering, E.J.; Kapchina-Toteva, V.M.; Iakimova, E.T. Mastoparan-induced programmed cell death in the unicellular alga Chlamydomonas reinhardtii. Ann. Bot. 2013, 111, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Vavilala, S.L.; Gawde, K.K.; Sinha, M.; D’Souza, J.S. Programmed cell death is induced by hydrogen peroxide but not by excessive ionic stress of sodium chloride in the unicellular green alga Chlamydomonas reinhardtii. Eur. J. Phycol. 2015, 50, 422–438. [Google Scholar] [CrossRef]
- Ge, Y.; Cai, Y.-M.; Bonneau, L.; Rotari, V.; Danon, A.; McKenzie, E.A.; McLellan, H.; Mach, L.; Gallois, P. Inhibition of cathepsin B by caspase-3 inhibitors blocks programmed cell death in Arabidopsis. Cell Death Differ. 2016, 23, 1493–1501. [Google Scholar] [CrossRef]
- Hatsugai, N.; Yamada, K.; Goto-Yamada, S.; Hara-Nishimura, I. Vacuolar processing enzyme in plant programmed cell death. Front. Plant. Sci. 2015, 6, 234. [Google Scholar] [CrossRef]
- Coll, N.S.; Vercammen, D.; Smidler, A.; Clover, C.; Van Breusegem, F.; Dangl, J.L.; Epple, P. Arabidopsis type I metacaspases control cell death. Science 2010, 330, 1393–1397. [Google Scholar] [CrossRef]
- Tsiatsiani, L.; Van Breusegem, F.; Gallois, P.; Zavialov, A.; Lam, E.; Bozhkov, P.V. Metacaspases. Cell Death Differ. 2011, 18, 1279–1288. [Google Scholar] [CrossRef]
- Murik, O.; Elboher, A.; Kaplan, A. Dehydroascorbate: A possible surveillance molecule of oxidative stress and programmed cell death in the green alga Chlamydomonas reinhardtii. New Phytol. 2014, 202, 471–484. [Google Scholar] [CrossRef]
- Zuo, Z.; Zhu, Y.; Bai, Y.; Wang, Y. Acetic acid-induced programmed cell death and release of volatile organic compounds in Chlamydomonas reinhardtii. Plant. Physiol. Biochem. PPB 2012, 51, 175–184. [Google Scholar] [CrossRef]
- Moharikar, S.; D’Souza, J.S.; Kulkarni, A.B.; Rao, B.J. Apoptotic-Like Cell Death Pathway Is Induced in Unicellular Chlorophyte Chlamydomonas reinhardtii (chlorophyceae) Cells Following Uv Irradiation: Detection and Functional Analyses1. J. Phycol. 2006, 42, 423–433. [Google Scholar] [CrossRef]
- Durand, P.M.; Rashidi, A.; Michod, R.E. How an organism dies affects the fitness of its neighbors. Am. Nat. 2011, 177, 224–232. [Google Scholar] [CrossRef]
- Ferris, P.J.; Woessner, J.P.; Waffenschmidt, S.; Kilz, S.; Drees, J.; Goodenough, U.W. Glycosylated Polyproline II Rods with Kinks as a Structural Motif in Plant Hydroxyproline-Rich Glycoproteins. Biochemistry 2001, 40, 2978–2987. [Google Scholar] [CrossRef]
- Macfie, S.M.; Tarmohamed, Y.; Welbourn, P.M. Effects of cadmium, cobalt, copper, and nickel on growth of the green alga Chlamydomonas reinhardtii: The influences of the cell wall and pH. Arch. Environ. Contam. Toxicol. 1994, 27, 454–458. [Google Scholar] [CrossRef]
- Macfie, S.M.; Welbourn, P.M. The Cell Wall as a Barrier to Uptake of Metal Ions in the Unicellular Green Alga Chlamydomonas reinhardtii (Chlorophyceae). Arch. Environ. Contam. Toxicol. 2000, 39, 413–419. [Google Scholar] [CrossRef]
- Prasad, M.N.V.; Drej, K.; Skawińska, A.; Strałka, K. Toxicity of Cadmium and Copper in Chlamydomonas reinhardtii Wild-Type (WT 2137) and Cell Wall Deficient Mutant Strain (CW 15). Bull. Environ. Contam. Toxicol. 1998, 60, 306–311. [Google Scholar] [CrossRef]
- Baba, M.; Suzuki, I.; Shiraiwa, Y. Proteomic analysis of high-CO(2)-inducible extracellular proteins in the unicellular green alga, Chlamydomonas reinhardtii. Plant. Cell Physiol. 2011, 52, 1302–1314. [Google Scholar] [CrossRef]
- Takahashi, H.; Braby, C.E.; Grossman, A.R. Sulfur Economy and Cell Wall Biosynthesis during Sulfur Limitation of Chlamydomonas reinhardtii. Plant. Physiol. 2001, 127, 665–673. [Google Scholar] [CrossRef]
- Davies, J.P.; Yildiz, F.H.; Grossman, A. Sac1, a putative regulator that is critical for survival of Chlamydomonas reinhardtii during sulfur deprivation. EMBO J. 1996, 15, 2150–2159. [Google Scholar] [CrossRef]
- Van Donk, E.; LÜrling, M.; Hessen, D.O.; Lokhorst, G.M. Altered cell wall morphology in nutrient-deficient phytoplankton and its impact on grazers. Limnol. Oceanogr. 1997, 42, 357–364. [Google Scholar] [CrossRef] [Green Version]
- van Donk, E.; Hessen, D.O. Grazing resistance in nutrient-stressed phytoplankton. Oecologia 1993, 93, 508–511. [Google Scholar] [CrossRef]
- Lürling, M.; Van Donk, E. Life history consequences for Daphnia pulex feeding on nutrient-limited phytoplankton. Freshw. Biol. 1997, 38, 693–709. [Google Scholar] [CrossRef]
- Porter, K.G. Selective Grazing and Differential Digestion of Algae by Zooplankton. Nature 1973, 244, 179. [Google Scholar] [CrossRef]
- Scholz, M.; Hoshino, T.; Johnson, D.; Riley, M.R.; Cuello, J. Flocculation of wall-deficient cells of Chlamydomonas reinhardtii mutant cw15 by calcium and methanol. Biomass Bioenergy 2011, 35, 4835–4840. [Google Scholar] [CrossRef]
- Mini, P.; Demurtas, O.C.; Valentini, S.; Pallara, P.; Aprea, G.; Ferrante, P.; Giuliano, G. Agrobacterium-mediated and electroporation-mediated transformation of Chlamydomonas reinhardtii: A comparative study. BMC Biotechnol. 2018, 18, 11. [Google Scholar] [CrossRef]
- Segawa, T.; Matsuzaki, R.; Takeuchi, N.; Akiyoshi, A.; Navarro, F.; Sugiyama, S.; Yonezawa, T.; Mori, H. Bipolar dispersal of red-snow algae. Nat. Commun. 2018, 9, 3094. [Google Scholar] [CrossRef]
- Darwin, C. Journal of Researches into the Natural History and Geology of the Countries Visited during the Voyage of HMS Beagle Round the World, under the Command of Capt. Fitz Roy, R.N., 2nd ed.; Cambridge University Press: London, UK, 1860. [Google Scholar]
- Procházková, L.; Remias, D.; Holzinger, A.; Řezanka, T.; Nedbalová, L. Ecophysiological and morphological comparison of two populations of Chlainomonas sp. (Chlorophyta) causing red snow on ice-covered lakes in the High Tatras and Austrian Alps. Eur. J. Phycol. 2018, 53, 230–243. [Google Scholar] [CrossRef]
- Müller, T.; Bleiß, W.; Martin, C.-D.; Rogaschewski, S.; Fuhr, G. Snow algae from northwest Svalbard: Their identification, distribution, pigment and nutrient content. Polar Biol. 1998, 20, 14–32. [Google Scholar] [CrossRef]
- Remias, D.; Lütz-Meindl, U.; Lütz, C. Photosynthesis, pigments and ultrastructure of the alpine snow alga Chlamydomonas nivalis. Eur. J. Phycol. 2005, 40, 259–268. [Google Scholar] [CrossRef]
- Gorton, H.L.; Vogelmann, T.C. Ultraviolet Radiation and the Snow Alga Chlamydomonas nivalis (Bauer) Wille. Photochem. Photobiol. 2003, 77, 608–615. [Google Scholar] [CrossRef]
- Boussiba, S. Carotenogenesis in the green alga Haematococcus pluvialis: Cellular physiology and stress response. Physiol. Plant. 2000, 108, 111–117. [Google Scholar] [CrossRef]
- Kakizono, T.; Kobayashi, M.; Nagai, S. Effect of carbon/nitrogen ratio on encystment accompanied with astaxanthin formation in a green alga, Haematococcus pluvialis. J. Ferment. Bioeng. 1992, 74, 403–405. [Google Scholar] [CrossRef]
- Li, Y.; Sommerfeld, M.; Chen, F.; Hu, Q. Effect of photon flux densities on regulation of carotenogenesis and cell viability of Haematococcus pluvialis (Chlorophyceae). J. Appl. Phycol. 2010, 22, 253–263. [Google Scholar] [CrossRef]
- Li, Y.; Sommerfeld, M.; Chen, F.; Hu, Q. Consumption of oxygen by astaxanthin biosynthesis: A protective mechanism against oxidative stress in Haematococcus pluvialis (Chlorophyceae). J. Plant. Physiol. 2008, 165, 1783–1797. [Google Scholar] [CrossRef]
- Sarada, R.; Tripathi, U.; Ravishankar, G.A. Influence of stress on astaxanthin production in Haematococcus pluvialis grown under different culture conditions. Process. Biochem. 2002, 37, 623–627. [Google Scholar] [CrossRef]
- Tjahjono, A.E.; Hayama, Y.; Kakizono, T.; Terada, Y.; Nishio, N.; Nagai, S. Hyper-accumulation of astaxanthin in a green alga Haematococcus pluvialis at elevated temperatures. Biotechnol. Lett. 1994, 16, 133–138. [Google Scholar] [CrossRef]
- Rezanka, T.; Nedbalová, L.; Sigler, K.; Cepák, V. Identification of astaxanthin diglucoside diesters from snow alga Chlamydomonas nivalis by liquid chromatography-atmospheric pressure chemical ionization mass spectrometry. Phytochemistry 2008, 69, 479–490. [Google Scholar] [CrossRef]
- Grossman, A.R.; Lohr, M.; Im, C.S. Chlamydomonas reinhardtii in the Landscape of Pigments. Annu. Rev. Genet. 2004, 38, 119–173. [Google Scholar] [CrossRef]
- Lemoine, Y.; Schoefs, B. Secondary ketocarotenoid astaxanthin biosynthesis in algae: A multifunctional response to stress. Photosynth. Res. 2010, 106, 155–177. [Google Scholar] [CrossRef]
- Bidigare, R.R.; Ondrusek, M.E.; Kennicutt, M.C.; Iturriaga, R.; Harvey, H.R.; Hoham, R.W.; Macko, S.A. Evidence for a photoprotective function for secondary carotenoids of snow algae. J. Phycol. 1993, 29, 427–434. [Google Scholar] [CrossRef]
- Scibilia, L.; Girolomoni, L.; Berteotti, S.; Alboresi, A.; Ballottari, M. Photosynthetic response to nitrogen starvation and high light in Haematococcus pluvialis. Algal Res. 2015, 12, 170–181. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Baier, T.; Zanoni, F.; Zoccatelli, G.; Lauersen, K.J.; Wobbe, L.; Ballottari, M. Turning a green alga red: Engineering astaxanthin biosynthesis by intragenic pseudogene revival in Chlamydomonas reinhardtii. bioRxiv 2019, 535989. [Google Scholar] [CrossRef]
- Strenkert, D.; Schmollinger, S.; Gallaher, S.D.; Salomé, P.A.; Purvine, S.O.; Nicora, C.D.; Mettler-Altmann, T.; Soubeyrand, E.; Weber, A.P.M.; Lipton, M.S.; et al. Multiomics resolution of molecular events during a day in the life of Chlamydomonas. Proc. Natl. Acad. Sci. USA 2019, 116, 2374–2383. [Google Scholar] [CrossRef]
- Zou, Y.; Wenzel, S.; Müller, N.; Prager, K.; Jung, E.-M.; Kothe, E.; Kottke, T.; Mittag, M. An Animal-Like Cryptochrome Controls the Chlamydomonas Sexual Cycle. Plant. Physiol. 2017, 174, 1334–1347. [Google Scholar] [CrossRef]
- Nedelcu, A.M.; Marcu, O.; Michod, R.E. Sex as a response to oxidative stress: A twofold increase in cellular reactive oxygen species activates sex genes. Proc. R. Soc. B Biol. Sci. 2004, 271, 1591–1596. [Google Scholar] [CrossRef]
- Waffenschmidt, S.; Woessner, J.P.; Beer, K.; Goodenough, U.W. Isodityrosine cross-linking mediates insolubilization of cell walls in Chlamydomonas. Plant. Cell 1993, 5, 809–820. [Google Scholar]
- Waffenschmidt, S.; Kusch, T.; Woessner, J.P. A Transglutaminase Immunologically Related to Tissue Transglutaminase Catalyzes Cross-Linking of Cell Wall Proteins in Chlamydomonas reinhardtii. Plant. Physiol. 1999, 121, 1003–1015. [Google Scholar] [CrossRef]
- Lopez, D.; Hamaji, T.; Kropat, J.; De Hoff, P.; Morselli, M.; Rubbi, L.; Fitz-Gibbon, S.; Gallaher, S.D.; Merchant, S.S.; Umen, J.; et al. Dynamic Changes in the Transcriptome and Methylome of Chlamydomonas reinhardtii throughout Its Life Cycle. Plant. Physiol. 2015, 169, 2730–2743. [Google Scholar] [CrossRef]
- Joo, S.; Nishimura, Y.; Cronmiller, E.; Hong, R.H.; Kariyawasam, T.; Wang, M.H.; Shao, N.C.; El Akkad, S.-E.-D.; Suzuki, T.; Higashiyama, T.; et al. Gene Regulatory Networks for the Haploid-to-Diploid Transition of Chlamydomonas reinhardtii. Plant. Physiol. 2017, 175, 314–332. [Google Scholar] [CrossRef]
- Suzuki, L.; Woessner, J.P.; Uchida, H.; Kuroiwa, H.; Yuasa, Y.; Waffenschmidt, S.; Goodenough, U.W.; Kuroiwa, T. A Zygote-Specific Protein with Hydroxyproline-Rich Glycoprotein Domains and Lectin-Like Domains Involved in the Assembly of the Cell Wall of Chlamydomonas reinhardtii (chlorophyta). J. Phycol. 2000, 36, 571–583. [Google Scholar] [CrossRef]
- Heimerl, N.; Hommel, E.; Westermann, M.; Meichsner, D.; Lohr, M.; Hertweck, C.; Grossman, A.R.; Mittag, M.; Sasso, S. A giant type I polyketide synthase participates in zygospore maturation in Chlamydomonas reinhardtii. Plant. J. 2018, 95, 268–281. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Electron microscopy of zygospore formation in Chlamydomonas reinhardii. Protoplasma 1976, 87, 297–315. [Google Scholar] [CrossRef]
- VanWinkle-Swift, K.P.; Rickoll, W.L. The Zygospore Wall of Chlamydomonas Monoica (chlorophyceae): Morphogenesis and Evidence for the Presence of Sporopollenin1. J. Phycol. 1997, 33, 655–665. [Google Scholar] [CrossRef]
- Suzuki, L.; Johnson, C. Photoperiodic control of germination in the unicell Chlamydomonas. Naturwissenschaften 2002, 89, 214–220. [Google Scholar] [CrossRef]
- Trainor, F.R.; Gladych, R. Survival of algae in a desiccated soil: A 35-year study. Phycologia 1995, 34, 191–192. [Google Scholar] [CrossRef]
- Suzuki, L.; Yuasa, Y.; Kuroiwa, T. Transcription and Translation involved in Pellicle Formation in the Chlamydomonas reinhardtii Zygote. Cytologia 1997, 62, 421–425. [Google Scholar] [CrossRef]
- Avnimelech, Y.; Troeger, B.W.; Reed, L.W. Mutual Flocculation of Algae and Clay: Evidence and Implications. Science 1982, 216, 63–65. [Google Scholar] [CrossRef]
- Salim, S.; Bosma, R.; Vermuë, M.H.; Wijffels, R.H. Harvesting of microalgae by bio-flocculation. J. Appl. Phycol. 2011, 23, 849–855. [Google Scholar] [CrossRef]
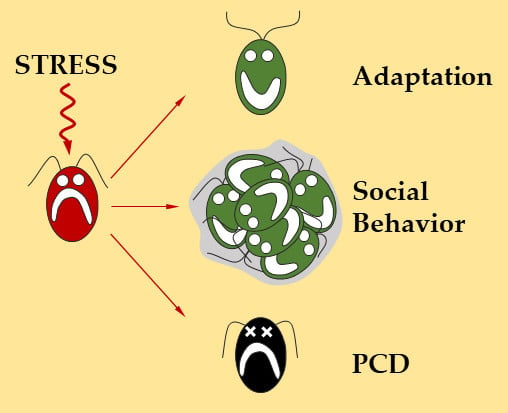
Figure 1.
The strategies used by Chlamydomonas to survive increasing stress intensities. Under optimal conditions Chlamydomonas can grow normally (green cell); under moderate stress conditions (light green cell), several acclimation processes can be triggered to allow the cells to better adapt and resist the occurrence of any additional stress of similar or greater magnitude. Under more intense stress conditions (orange cell), cells can form multicellular structures such as palmelloids or aggregates that can dissociate when environmental conditions improve. Under very harsh stress conditions (red cell), the most exposed cells will self-destruct and release in the medium, molecules that will allow other cells to survive, to avoid the disappearance of the entire population. In the case of a stress of an intensity such that the cell cannot overcome (skull), the cell is physically destroyed by its environment through necrosis.
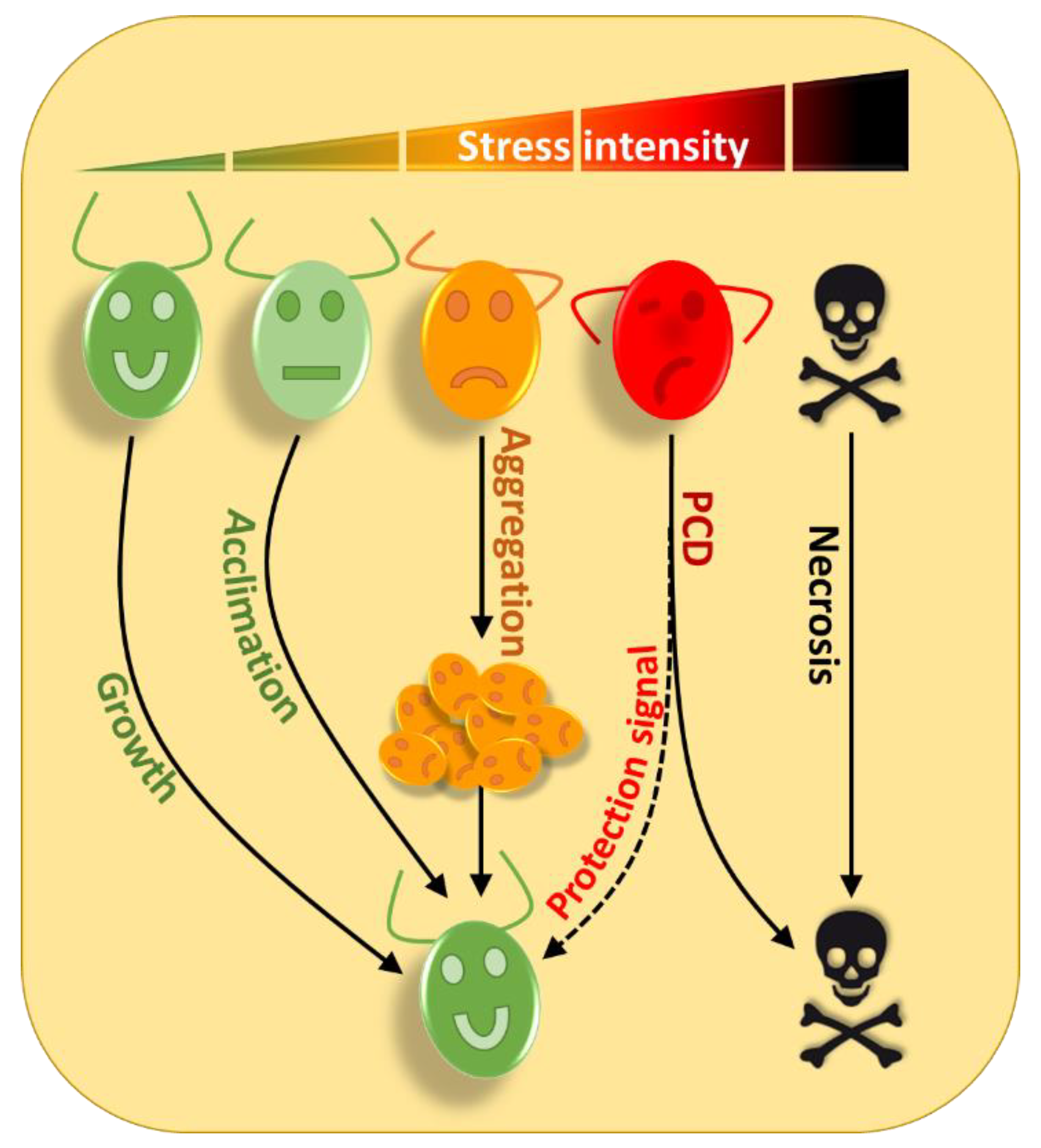
Figure 1.
The strategies used by Chlamydomonas to survive increasing stress intensities. Under optimal conditions Chlamydomonas can grow normally (green cell); under moderate stress conditions (light green cell), several acclimation processes can be triggered to allow the cells to better adapt and resist the occurrence of any additional stress of similar or greater magnitude. Under more intense stress conditions (orange cell), cells can form multicellular structures such as palmelloids or aggregates that can dissociate when environmental conditions improve. Under very harsh stress conditions (red cell), the most exposed cells will self-destruct and release in the medium, molecules that will allow other cells to survive, to avoid the disappearance of the entire population. In the case of a stress of an intensity such that the cell cannot overcome (skull), the cell is physically destroyed by its environment through necrosis.

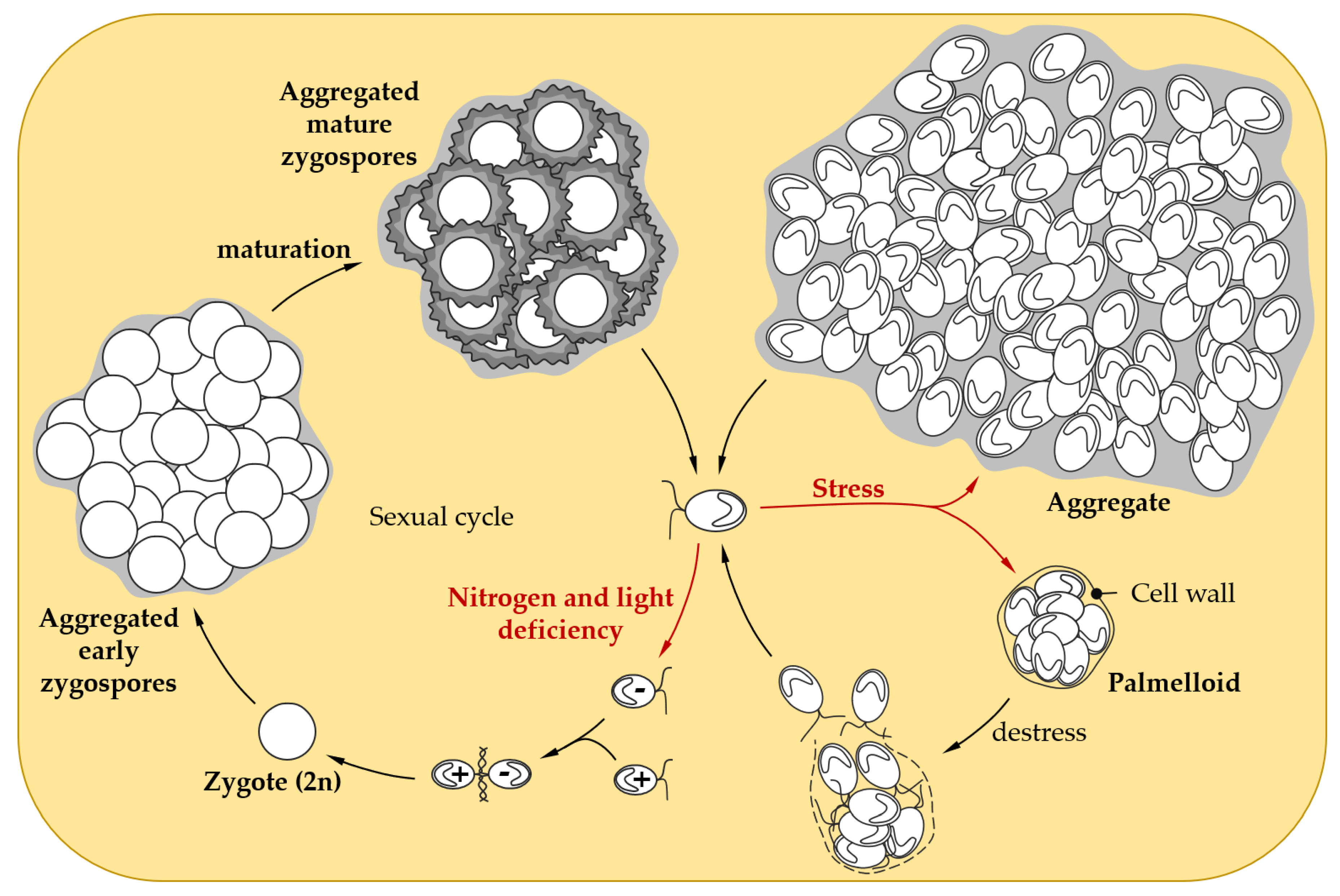
Figure 2.
Social behavior in response to stress in Chlamydomonas. When facing adverse conditions, Chlamydomonas vegetative cells can form palmelloids, a cluster of 4 to 16 cells surrounded by a cell wall resulting from the division of a single cell. As soon as the environmental conditions improve, the outer cell wall is hatched, to allow the liberation of daughter cells [55]. When confronted with harsher stress conditions, Chlamydomonas cells are able to form larger multicellular structures called aggregates that might confer stress tolerance [60,61]. Under conditions of non-optimal growth (e.g., nitrogen or light limitation), Chlamydomonas can induce gamete differentiation to enable sexual reproduction, which leads to zygote formation after the mating of gametes. Maturation of the zygospore confers resistance to environmental stresses, such as desiccation [64]. In liquid cultures, early and mature zygospores can form aggregates, that could further enhance resistance to stress [65,66,67].
Figure 2.
Social behavior in response to stress in Chlamydomonas. When facing adverse conditions, Chlamydomonas vegetative cells can form palmelloids, a cluster of 4 to 16 cells surrounded by a cell wall resulting from the division of a single cell. As soon as the environmental conditions improve, the outer cell wall is hatched, to allow the liberation of daughter cells [55]. When confronted with harsher stress conditions, Chlamydomonas cells are able to form larger multicellular structures called aggregates that might confer stress tolerance [60,61]. Under conditions of non-optimal growth (e.g., nitrogen or light limitation), Chlamydomonas can induce gamete differentiation to enable sexual reproduction, which leads to zygote formation after the mating of gametes. Maturation of the zygospore confers resistance to environmental stresses, such as desiccation [64]. In liquid cultures, early and mature zygospores can form aggregates, that could further enhance resistance to stress [65,66,67].


{kind=link}
{kind=link}
{kind=link}
Table 1.
Multicellular structure formation in response to stress in Chlamydomonas. In response to different types of biotic or abiotic stresses and depending on their intensity, Chlamydomonas can form palmelloids or aggregates, suggesting that socialization may be a conserved mechanism that help algal cells adapt to harsh environmental conditions.
Table 1.
Multicellular structure formation in response to stress in Chlamydomonas. In response to different types of biotic or abiotic stresses and depending on their intensity, Chlamydomonas can form palmelloids or aggregates, suggesting that socialization may be a conserved mechanism that help algal cells adapt to harsh environmental conditions.
| Behavior | Stress | Conditions | Reference |
|---|---|---|---|
| Palmelloids | Predator | Brachionus calyciflorus | [50] |
| Organic acids (succinate, fumarate, aspartate, glutamate, glycolate, citrate, phthalate) | 0.15–5% | [47] | |
| EDTA, GEDTA | 1.25 mM | [51] | |
| Calcium deficiency | <3.5 µM | ||
| Phosphorous deficiency | <1 µg/L | [52] | |
| Cadmium | 200–400 µM | [53] | |
| NaCl | 300–700 mM | [54] | |
| 100–150 mM | [55] | ||
| 50–150 mM | [56] | ||
| Acidic pH | pH 4.4 | [57] | |
| Chloroplatinic acid | 50 µM | [58,59] | |
| Aggregates | Predator | Brachionus calyciflorus | [50] |
| Peranema trichophorum | [60] | ||
| Acidic pH | pH 3.4 | [57] | |
| pH 2.5–pH 4 | [61] | ||
| Basic pH | pH 10–pH 13 | ||
| FeCl3, CaCl2, MgCl2 | 1–10 mM | ||
| Naphthenic acids | 100 mg/L | [62] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
de Carpentier, F.; Lemaire, S.D.; Danon, A. When Unity Is Strength: The Strategies Used by Chlamydomonas to Survive Environmental Stresses. Cells 2019, 8, 1307. https://doi.org/10.3390/cells8111307
AMA Style
de Carpentier F, Lemaire SD, Danon A. When Unity Is Strength: The Strategies Used by Chlamydomonas to Survive Environmental Stresses. Cells. 2019; 8(11):1307. https://doi.org/10.3390/cells8111307
Chicago/Turabian Stylede Carpentier, Félix, Stéphane D. Lemaire, and Antoine Danon. 2019. "When Unity Is Strength: The Strategies Used by Chlamydomonas to Survive Environmental Stresses" Cells 8, no. 11: 1307. https://doi.org/10.3390/cells8111307
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.