Protective Effect of Probiotic Bacteria and Estrogen in Preventing HIV-1-Mediated Impairment of Epithelial Barrier Integrity in Female Genital Tract


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Primary Genital Tract Epithelial Cells
2.2. Hormone and Contraceptive Treatment
2.3. Treatment of GECs with Lactobacilli and HIV-1 Exposure
2.4. Fluorescein Isothiocyanate (FITC)-Dextran Dye Assay for Cell Monolayer Permeability Assay
2.5. Cell Stress and Cytotoxicity Measured by Lactate Dehydrogenase Release
2.6. Magpix Multi-Analyte Assay for Measurement of Cytokines and Chemokines
2.7. Immunofluorescent Staining of Epithelial Cell Cultures
2.8. Statistical Analysis
3. Results
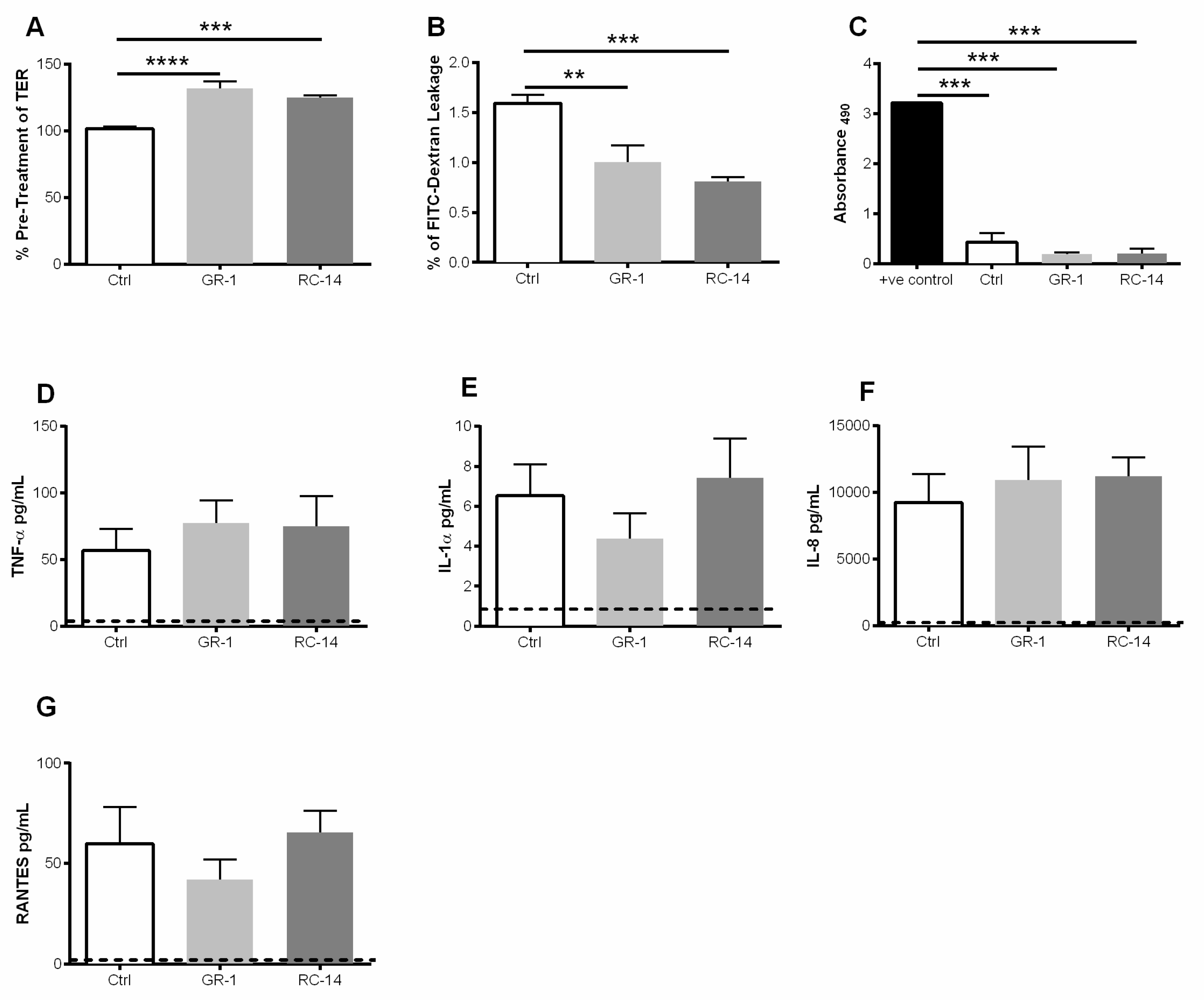
3.1. Exposure to Probiotic Strains of Lactobacilli Improves the Barrier Function of Primary Upper Genital Tract Epithelial Cells
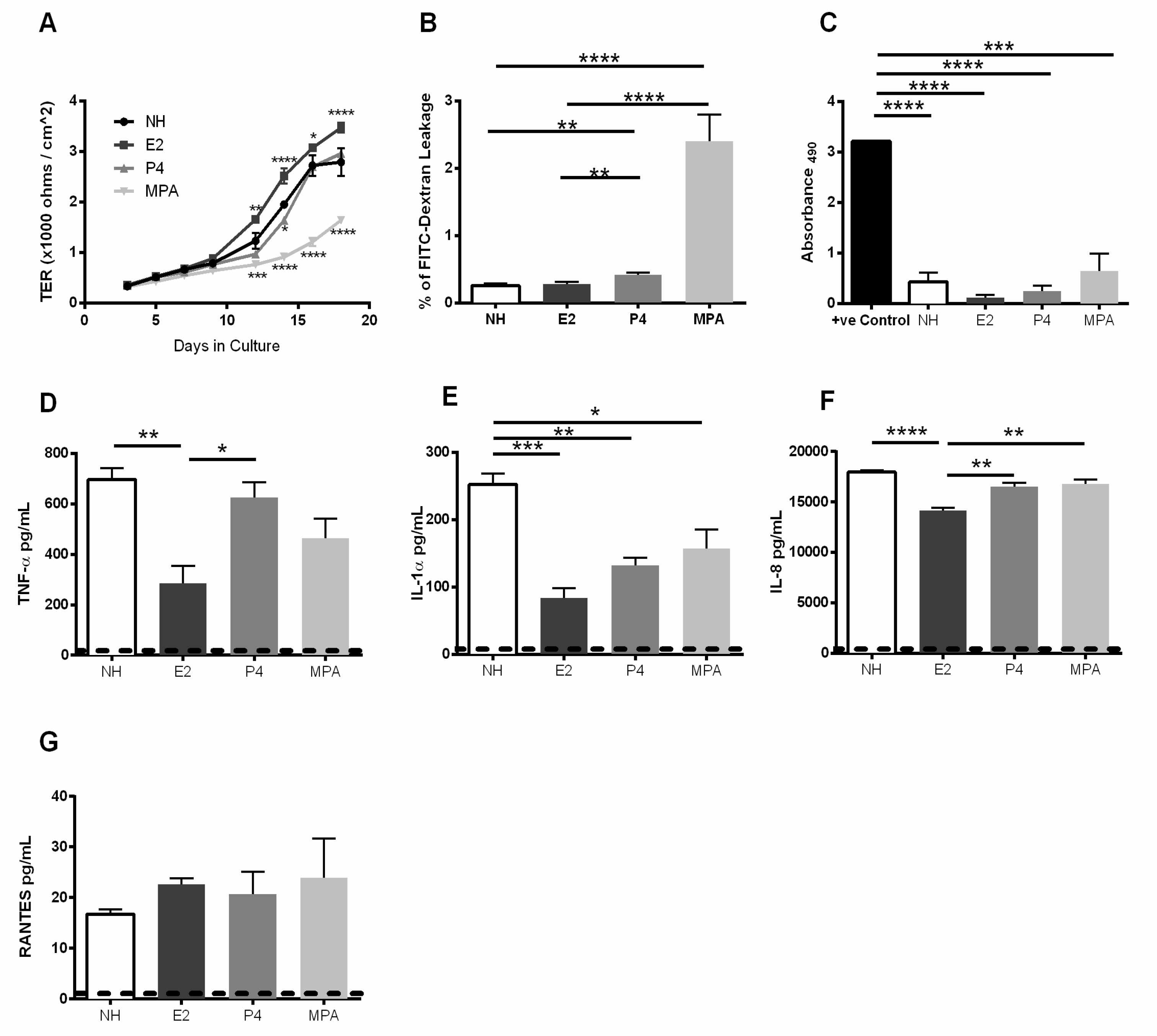
3.2. Estrogen Enhances Barrier Function and Modulates Proinflammatory Cytokine and Chemokine Response in Primary GECs
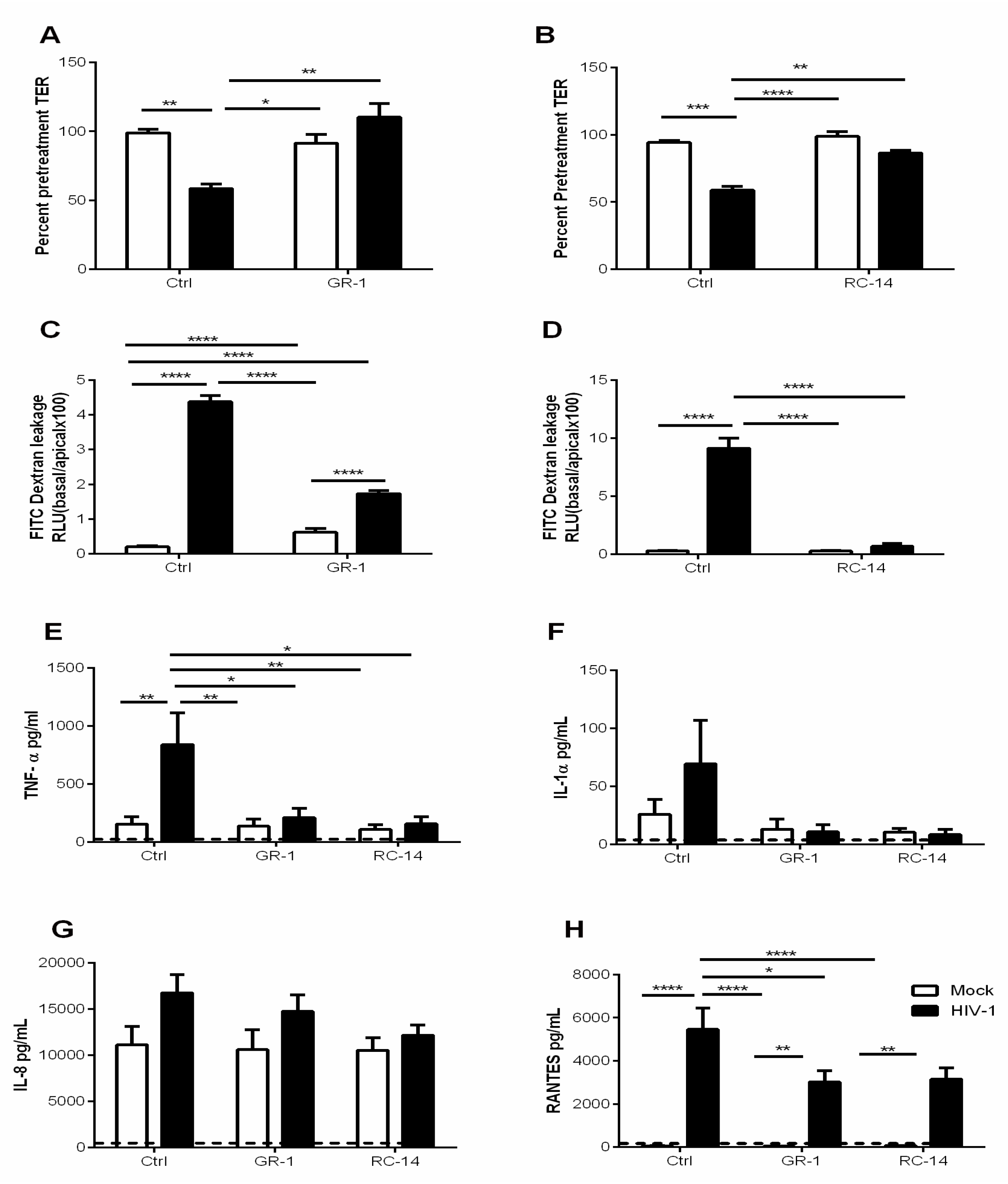
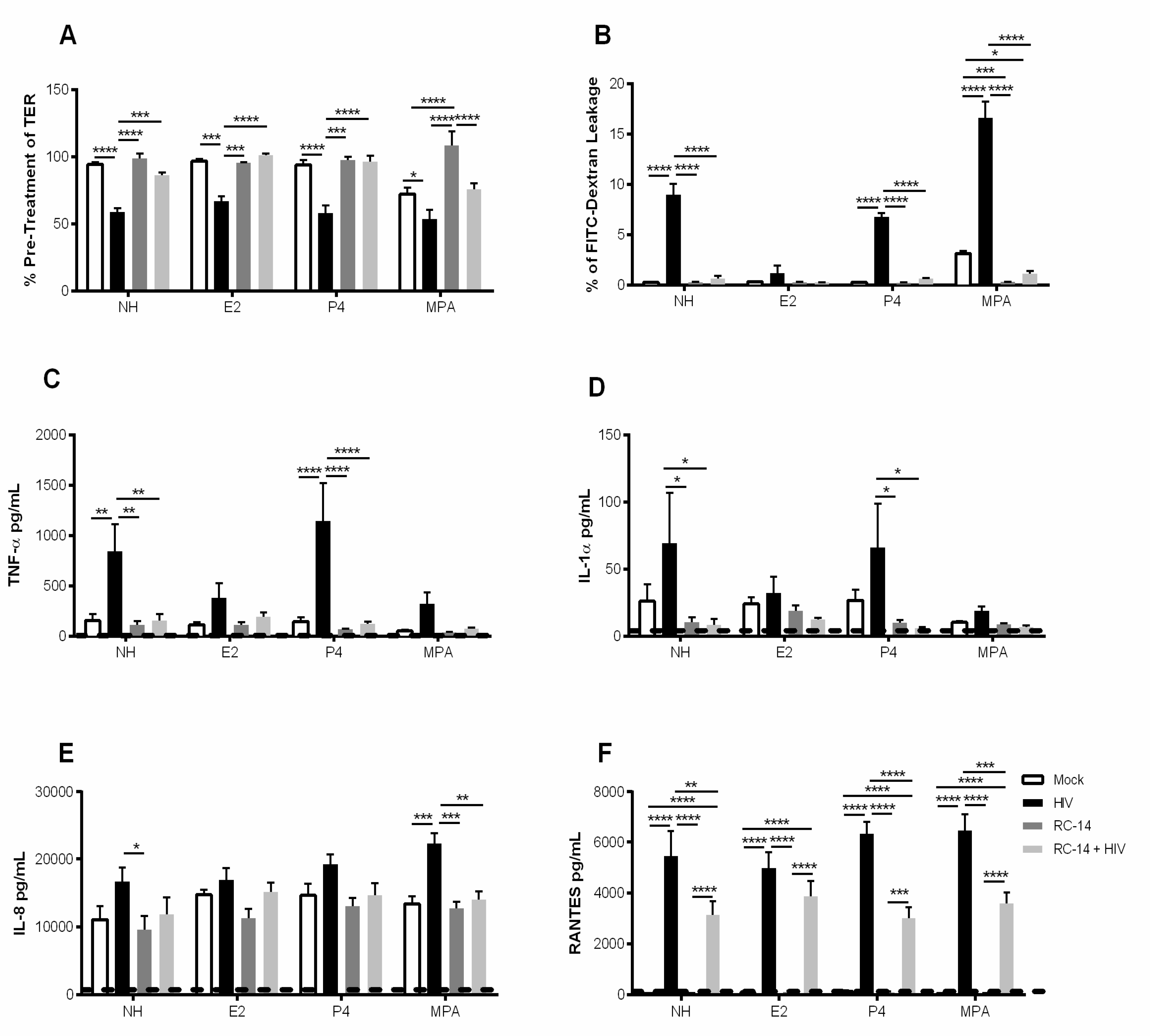
3.3. Pre-Exposure of GECs to Probiotic Lactobacilli Prevents HIV-1-Mediated Barrier Disruption and Downregulates Proinflammatory Cytokine Production by GECs
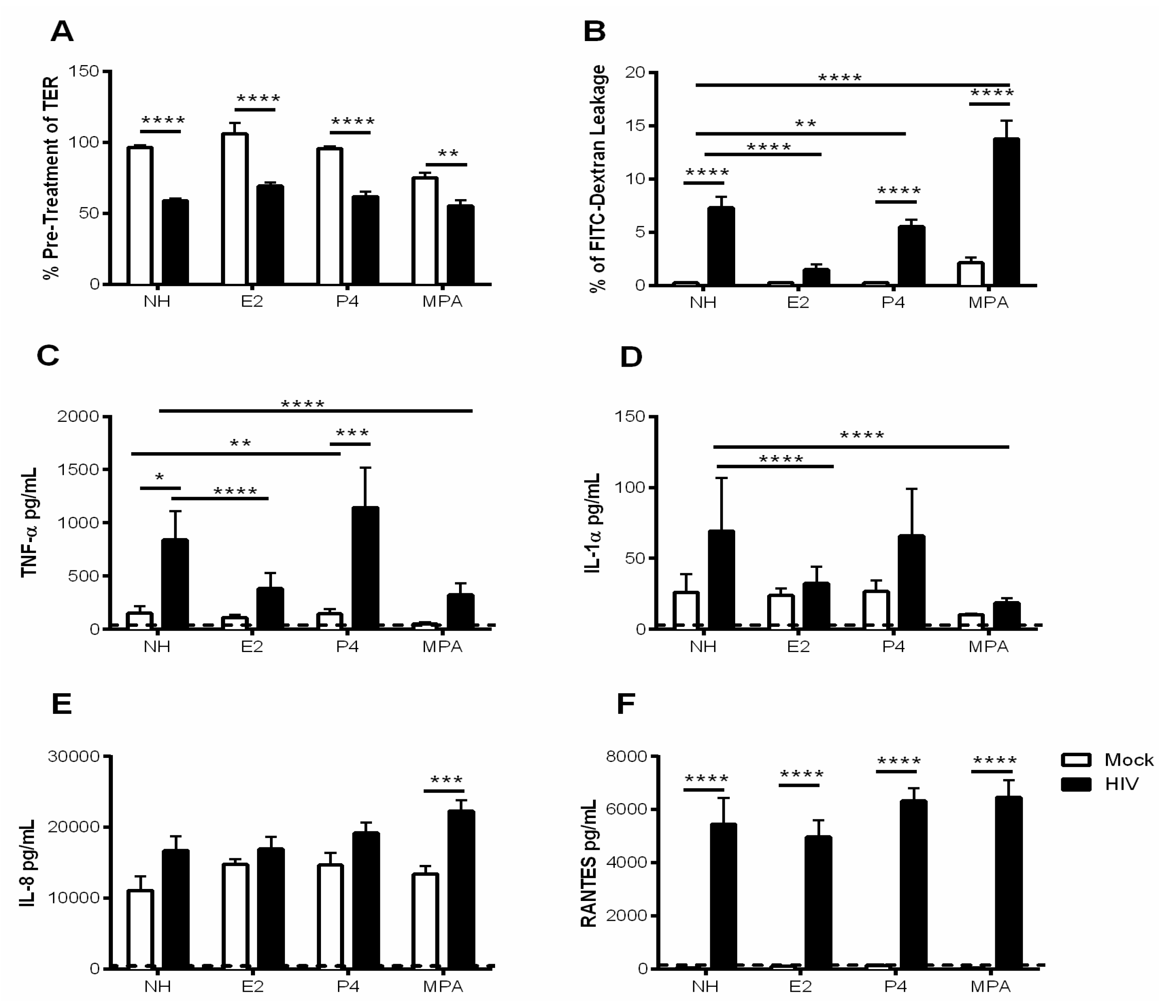
3.4. Estrogen Pre-Treatment Prevents Leakage and Decreases TNF-α Production but not RANTES Production Induced by HIV-1
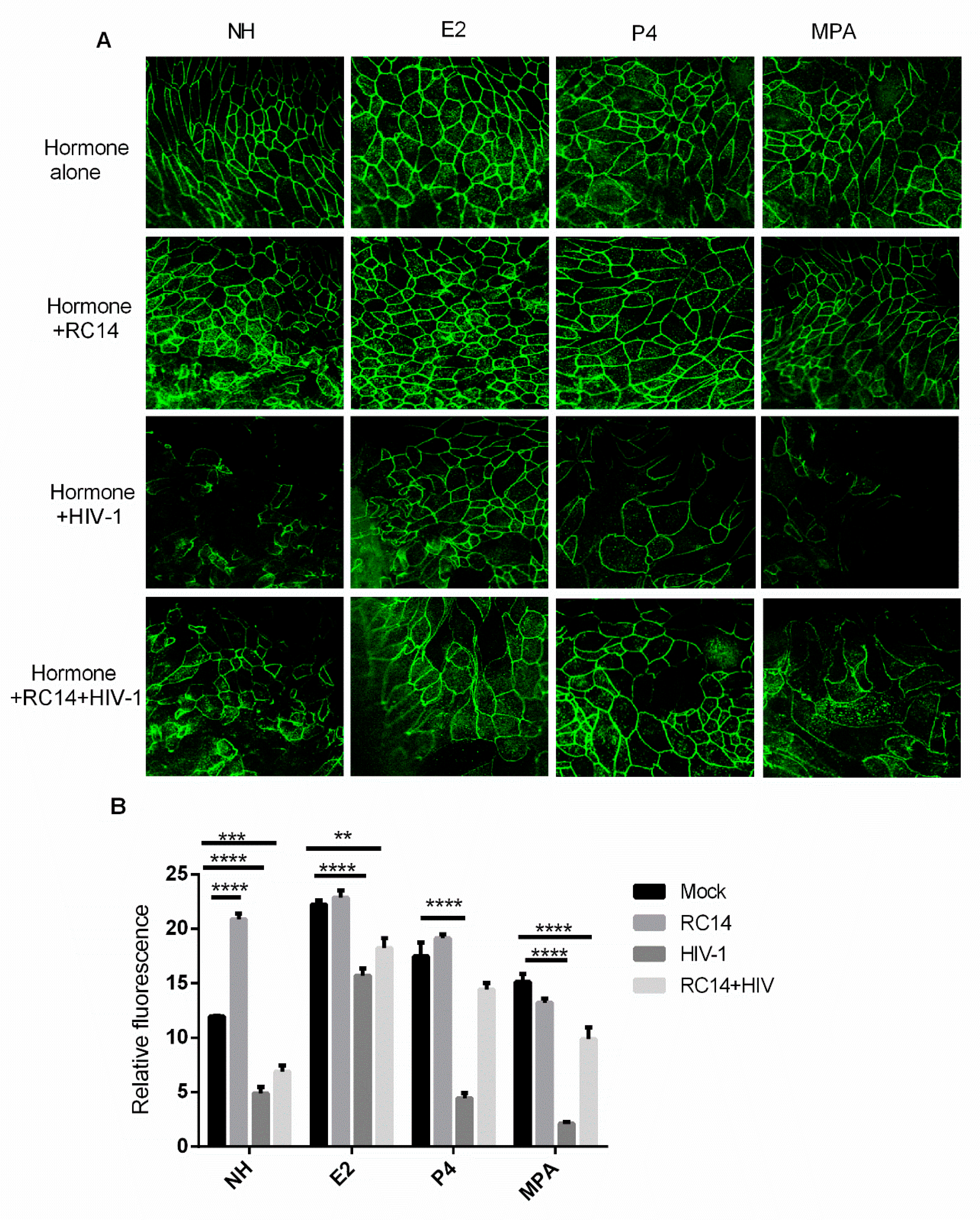
3.5. Probiotic Lactobacilli and Estrogen Prevent HIV-1-Mediated Epithelial Barrier Disruption
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNAIDS Gap Report 2014: Gender Based Inequalities. Available online: http://files.unaids.org/en/media/unaids/contentassets/documents/unaidspublication/2014/UNAIDS_Gap_report_en.pdf (accessed on 13 December 2016).
- Hladik, F.; Hope, T.J. HIV infection of the genital mucosa in women. Curr. HIV/AIDS Rep. 2009, 6, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.V.; Kafka, J.K.; Ferreira, V.H.; Roth, K.; Kaushic, C. Innate and adaptive immune responses in male and female reproductive tracts in homeostasis and following HIV infection. Cell. Mol. Immunol. 2014, 11, 410–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, V.H.; Kafka, J.K.; Kaushic, C. Influence of Common Mucosal Co-Factors on HIV Infection in the Female Genital Tract. Am. J. Reprod. Immunol. 2014, 71, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.R.; Da Silva, M.V.; Cavellani, C.L.; Dos Reis, M.A.; Monteiro, M.L.G.D.R.; Teixeira, V.D.P.A.; Corrêa, R.R.M. Mucosal Immunity in the Female Genital Tract, HIV/AIDS. BioMed Res. Int. 2014, 2014, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaul, R.; Prodger, J.; Joag, V.; Shannon, B.; Yegorov, S.; Galiwango, R.; McKinnon, L. Inflammation and HIV Transmission in Sub-Saharan Africa. Curr. HIV/AIDS Rep. 2015, 12, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Passmore, J.-A.S.; Jaspan, H.B.; Masson, L. Genital inflammation, immune activation and risk of sexual HIV acquisition. Curr. Opin. HIV AIDS 2016, 11, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Wessels, J.M.; Felker, A.M.; Dupont, H.A.; Kaushic, C. The relationship between sex hormones, the vaginal microbiome and immunity in HIV-1 susceptibility in women. Dis. Model. Mech. 2018, 11, dmm035147. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.J.; Pudney, J.; Quayle, A.J. Immunological Microenvironments in the Human Vagina and Cervix: Mediators of Cellular Immunity Are Concentrated in the Cervical Transformation Zone1. Boil. Reprod. 2005, 73, 1253–1263. [Google Scholar] [Green Version]
- Haase, A.T. Targeting early infection to prevent HIV-1 mucosal transmission. Nature 2010, 464, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Nazli, A.; Chan, O.; Dobson-Belaire, W.N.; Ouellet, M.; Tremblay, M.J.; Gray-Owen, S.D.; Arsenault, A.L.; Kaushic, C. Exposure to HIV-1 Directly Impairs Mucosal Epithelial Barrier Integrity Allowing Microbial Translocation. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef]
- Nazli, A.; Kafka, J.K.; Ferreira, V.H.; Anipindi, V.; Mueller, K.; Osborne, B.J.; Dizzell, S.; Chauvin, S.; Mian, M.F.; Ouellet, M.; et al. HIV-1 gp120 Induces TLR2- and TLR4-Mediated Innate Immune Activation in Human Female Genital Epithelium. J. Immunol. 2013, 191, 4246–4258. [Google Scholar] [CrossRef] [PubMed]
- Wira, C.R.; Rodríguez-García, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Wira, C.R.; Fahey, J.V.; Ghosh, M.; Patel, M.V.; Hickey, D.K.; Ochiel, D.O. Sex Hormone Regulation of Innate Immunity in the Female Reproductive Tract: The Role of Epithelial Cells in Balancing Reproductive Potential with Protection against Sexually Transmitted Pathogens. Am. J. Reprod. Immunol. 2010, 63, 544–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushic, C.; Roth, K.L.; Anipindi, V.; Xiu, F. Increased prevalence of sexually transmitted viral infections in women: The role of female sex hormones in regulating susceptibility and immune responses. J. Reprod. Immunol. 2011, 88, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Wira, C.R.; Rodriguez-Garcia, M.; Shen, Z.; Patel, M.; Fahey, J.V. The Role of Sex Hormones and the Tissue Environment in Immune Protection Against HIV in the Female Reproductive Tract. Am. J. Reprod. Immunol. 2014, 72, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Family Planning Worldwide 2002 Data Sheet. Available online: https://www.prb.org/familyplanningworldwide2002datasheet (accessed on 13 December 2016).
- Huijbregts, R.P.H.; Helton, E.S.; Michel, K.G.; Sabbaj, S.; Richter, H.E.; Goepfert, P.A.; Hel, Z. Hormonal Contraception and HIV-1 Infection: Medroxyprogesterone Acetate Suppresses Innate and Adaptive Immune Mechanisms. Endocrinology 2013, 154, 1282–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, C.S.; Chen, P.-L.; Kwok, C.; Baeten, J.M.; Brown, J.; Crook, A.M.; Van Damme, L.; Delany-Moretlwe, S.; Francis, S.C.; Friedland, B.A.; et al. Hormonal Contraception and the Risk of HIV Acquisition: An Individual Participant Data Meta-analysis. PLoS Med. 2015, 12, e1001778. [Google Scholar] [CrossRef] [PubMed]
- Polis, C.B.; Phillips, S.J.; Curtis, K.M.; Westreich, D.J.; Steyn, P.S.; Raymond, E.; Hannaford, P.; Turner, A.N. Hormonal contraceptive methods and risk of HIV acquisition in women: A systematic review of epidemiological evidence. Contraception 2014, 90, 360–390. [Google Scholar] [CrossRef]
- Polis, C.B.; Curtis, K.M. Use of hormonal contraceptives and HIV acquisition in women: A systematic review of the epidemiological evidence. Lancet Infect. Dis. 2013, 13, 797–808. [Google Scholar] [CrossRef]
- Crook, A.M.; Ford, D.; Gafos, M.; Hayes, R.; Kamali, A.; Kapiga, S.; Nunn, A.; Chisembele, M.; Ramjee, G.; Rees, H.; et al. Injectable and oral contraceptives and risk of HIV acquisition in women: An analysis of data from the MDP301 trial. Hum. Reprod. 2014, 29, 1810–1817. [Google Scholar] [CrossRef]
- Ralph, L.J.; Gollub, E.L.; Jones, H.E. Hormonal contraceptive use and women’s risk of HIV acquisition: Priorities emerging from recent data. Curr. Opin. Obstet. Gynecol. 2015, 27, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Brind, J.; Condly, S.J.; Mosher, S.W.; Morse, A.R.; Kimball, J. Risk of HIV Infection in Depot-Medroxyprogesterone Acetate (DMPA) Users: A Systematic Review and Meta-analysis. Issues Law Med. 2015, 30, 129–139. [Google Scholar] [PubMed]
- Woods, M.W.; Zahoor, M.A.; Dizzell, S.; Verschoor, C.P.; Kaushic, C. Medroxyprogesterone acetate-treated human, primary endometrial epithelial cells reveal unique gene expression signature linked to innate immunity and HIV-1 susceptibility. Am. J. Reprod. Immunol. 2018, 79, e12781. [Google Scholar] [CrossRef] [PubMed]
- Kaushic, C.; Nazli, A.; Ferreira, V.H.; Kafka, J.K. Primary human epithelial cell culture system for studying interactions between female upper genital tract and sexually transmitted viruses, HSV-2 and HIV-1. Methods 2011, 55, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Fu, L.; Mauck, C.K.; Koenig, S.S.; Sakamoto, J.; Motsinger-Reif, A.A.; Doncel, G.F.; Zeichner, S.L. Twice-daily application of HIV microbicides alter the vaginal microbiota. mBio 2012, 3, e00370-12. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the upper genital tract by vaginal bacterial species in nonpregnant women. Am. J. Obstet. Gynecol. 2015, 212, 611.e1–611.e9. [Google Scholar] [CrossRef]
- Petrova, M.I.; Broek, M.V.D.; Balzarini, J.; Vanderleyden, J.; Lebeer, S. Vaginal microbiota and its role in HIV transmission and infection. FEMS Microbiol. Rev. 2013, 37, 762–792. [Google Scholar] [CrossRef] [Green Version]
- Verstraelen, H.; Vilchez-Vargas, R.; Desimpel, F.; Jauregui, R.; Vankeirsbilck, N.; Weyers, S.; Verhelst, R.; De Sutter, P.; Pieper, D.H.; Van De Wiele, T. Characterisation of the human uterine microbiome in non-pregnant women through deep sequencing of the V1-2 region of the 16S rRNA gene. PeerJ 2016, 4. [Google Scholar] [CrossRef]
- Doerflinger, S.Y.; Throop, A.L.; Herbst-Kralovetz, M.M. Bacteria in the Vaginal Microbiome Alter the Innate Immune Response and Barrier Properties of the Human Vaginal Epithelia in a Species-Specific Manner. J. Infect. Dis. 2014, 209, 1989–1999. [Google Scholar] [CrossRef] [Green Version]
- Gosmann, C.; Anahtar, M.N.; Handley, S.A.; Farcasanu, M.; Abu-Ali, G.; Bowman, B.A.; Padavattan, N.; Desai, C.; Droit, L.; Moodley, A.; et al. Lactobacillus-Deficient Cervicovaginal Bacterial Communities Are Associated with Increased HIV Acquisition in Young South African Women. Immunity 2017, 46, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Forney, L.J.; Ravel, J. The vaginal microbiome: Rethinking health and diseases. Annu. Rev. Microbiol. 2012, 66, 371–389. [Google Scholar] [CrossRef] [PubMed]
- Thwin, S.S.; Agnew, K.; Meier, A.; Stamm, W.E.; Eschenbach, D.A.; Patton, D.L.; Hooton, T.M.; Stapleton, A.E.; Winter, C. Influence of the Normal Menstrual Cycle on Vaginal Tissue, Discharge, and Microflora. Clin. Infect. Dis. 2000, 30, 901–907. [Google Scholar]
- Reid, G. The development of probiotics for women’s health. Can. J. Microbiol. 2017, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Benson, K.; Cramer, S.; Galla, H.-J. Impedance-based cell monitoring: barrier properties and beyond. Fluids Barriers CNS 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.H.; Dizzell, S.; Nazli, A.; Kafka, J.K.; Mueller, K.; Nguyen, P.V.; Tremblay, M.J.; Cochrane, A.; Kaushic, C. Medroxyprogesterone Acetate Regulates HIV-1 Uptake and Transcytosis but Not Replication in Primary Genital Epithelial Cells Resulting in Enhanced T-Cell Infection. J. Infect. Dis. 2015, 211, 1745–1756. [Google Scholar] [CrossRef]
- Kimpton, J.; Emerman, M. Detection of Replication-competent and Pseudotyped Human Immunodficiency virus with a sensitive cell line on the basis of Activation of an Integrated Beta-Galactosidase Gene. J. Virol. 1992, 66, 2232–2239. [Google Scholar]
- Legrand, C.; Bour, J.M.; Jacob, C.; Capiaumont, J.; Martial, A.; Marc, A.; Wudtke, M.; Kretzmer, G.; Demangel, C.; Duval, D. Lactate dehydrogenase (LDH) activity of the cultured eukaryotic cells as marker of the number of dead cells in the medium [corrected]. J. Biotechnol. 1992, 25, 231–243. [Google Scholar] [CrossRef]
- Brotman, R.M.; Ravel, J.; Bavoil, P.M.; Gravitt, P.E.; Ghanem, K.G. Microbiome, sex hormones, and immune responses in the reproductive tract: Challenges for vaccine development against sexually transmitted infections. Vaccine 2014, 32, 1543–1552. [Google Scholar] [CrossRef]
- Saba, E.; Origoni, M.; Taccagni, G.; Ferrari, D.; Doglioni, C.; Nava, A.; Lisco, A.; Grivel, J.-C.; Margolis, L.; Poli, G. Productive HIV-1 Infection of Human Cervical Tissue Ex Vivo is Associated with the Secretory Phase of the Menstrual Cycle. Mucosal Immunol. 2013, 6, 1081–1090. [Google Scholar] [CrossRef]
- Calla, N.E.Q.; Miguel, R.D.V.; E Glick, M.; Kwiek, J.J.; Gabriel, J.M.; Cherpes, T.L. Exogenous oestrogen inhibits genital transmission of cell-associated HIV-1 in DMPA-treated humanized mice. J. Int. AIDS Soc. 2018, 21, e25063. [Google Scholar] [CrossRef]
- Maritz, M.F.; Ray, R.M.; Bick, A.J.; Tomasicchio, M.; Woodland, J.G.; Govender, Y.; Avenant, C.; Hapgood, J.P. Medroxyprogesterone acetate, unlike norethisterone, increases HIV-1 replication in human peripheral blood mononuclear cells and an indicator cell line, via mechanisms involving the glucocorticoid receptor, increased CD4/CD8 ratios and CCR5 levels. PLoS ONE 2018, 13, e0196043. [Google Scholar] [CrossRef]
- Masson, L.; Passmore, J.-A.S.; Liebenberg, L.J.; Werner, L.; Baxter, C.; Arnold, K.B.; Williamson, C.; Little, F.; Mansoor, L.E.; Naranbhai, V.; et al. Genital Inflammation and the Risk of HIV Acquisition in Women. Clin. Infect. Dis. 2015, 61, 260–269. [Google Scholar] [CrossRef]
- Herold, B.C.; Keller, M.J.; Shi, Q.; Hoover, D.R.; Carpenter, C.A.; Huber, A.; Parikh, U.M.; Agnew, K.J.; Minkoff, H.; Colie, C.; et al. Plasma and Mucosal HIV Viral Loads Are Associated with Genital Tract Inflammation In HIV-Infected Women. JAIDS J. Acquir. Immune Defic. Syndr. 2013, 63, 485–493. [Google Scholar] [CrossRef]
- Hunt, P.W.; Sinclair, E.; Rodriguez, B.; Shive, C.; Clagett, B.; Funderburg, N.; Robinson, J.; Huang, Y.; Epling, L.; Martin, J.N.; et al. Gut Epithelial Barrier Dysfunction and Innate Immune Activation Predict Mortality in Treated HIV Infection. J. Infect. Dis. 2014, 210, 1228–1238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles and applications of probiotic Lactobacillus strains. Appl. Microbiol. Biotechnol. 2018, 102, 8135–8143. [Google Scholar] [CrossRef]
- Kozakova, H.; Schwarzer, M.; Tuckova, L.; Srutkova, D.; Czarnowska, E.; Rosiak, I.; Hudcovic, T.; Schabussova, I.; Hermanova, P.; Zakostelska, Z.; et al. Colonization of Germ-Free Mice with a Mixture of Three Lactobacillus Strains Enhances the Integrity of Gut Mucosa and Ameliorates Allergic Sensitization. Cell. Mol. Immunol. 2016, 13, 251–262. [Google Scholar] [CrossRef]
- Valenti, P.; Rosa, L.; Capobianco, D.; Lepanto, M.S.; Schiavi, E.; Cutone, A.; Paesano, R.; Mastromarino, P. Role of Lactobacilli and Lactoferrin in the Mucosal Cervicovaginal Defense. Front. Immunol. 2018, 9, 376. [Google Scholar] [CrossRef]
- Masson, L.; Arnold, K.B.; Little, F.; Mlisana, K.; Lewis, D.A.; Mkhize, N.; Gamieldien, H.; Ngcapu, S.; Johnson, L.; Lauffenburger, D.A.; et al. Inflammatory cytokine biomarkers to identify women with asymptomatic sexually transmitted infections and bacterial vaginosis who are at high risk of HIV infection. Sex. Transm. Infect. 2016, 92, 186–193. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Guo, S.; Ye, D.; Ma, T.Y. TNF-α Modulation of Intestinal Epithelial Tight Junction Barrier Is Regulated by ERK1/2 Activation of Elk-1. Am. J. Pathol. 2013, 183, 1871–1884. [Google Scholar] [CrossRef]
- Turner, J.R.; Buschmann, M.M.; Romero-Calvo, I.; Sailer, A.; Shen, L. The role of molecular remodeling in differential regulation of tight junction permeability. Semin. Cell Dev. Boil. 2014, 36, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [Green Version]
- Pelekanou, V.; Kampa, M.; Kiagiadaki, F.; Deli, A.; Theodoropoulos, P.; Agrogiannis, G.; Patsouris, E.; Tsapis, A.; Castanas, E.; Notas, G. Estrogen Anti-Inflammatory Activity on Human Monocytes is Mediated Through Cross-Talk Between Estrogen Receptor ERα36 and GPR30/GPER1. J. Leukoc. Biol. 2016, 99, 333–347. [Google Scholar] [CrossRef]
- Santos, R.S.; De Fatima, L.A.; Frank, A.P.; Carneiro, E.M.; Clegg, D.J. The effects of 17 alpha-estradiol to inhibit inflammation in vitro. Boil. Sex Differ. 2017, 8, 30. [Google Scholar] [CrossRef]
- Chandra, N.; Thurman, A.R.; Anderson, S.; Cunningham, T.D.; Yousefieh, N.; Mauck, C.; Doncel, G.F. Depot Medroxyprogesterone Acetate Increases Immune Cell Numbers and Activation Markers in Human Vaginal Mucosal Tissues. AIDS Res. Hum. Retroviruses 2013, 29, 592–601. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dizzell, S.; Nazli, A.; Reid, G.; Kaushic, C. Protective Effect of Probiotic Bacteria and Estrogen in Preventing HIV-1-Mediated Impairment of Epithelial Barrier Integrity in Female Genital Tract. Cells 2019, 8, 1120. https://doi.org/10.3390/cells8101120
Dizzell S, Nazli A, Reid G, Kaushic C. Protective Effect of Probiotic Bacteria and Estrogen in Preventing HIV-1-Mediated Impairment of Epithelial Barrier Integrity in Female Genital Tract. Cells. 2019; 8(10):1120. https://doi.org/10.3390/cells8101120
Chicago/Turabian StyleDizzell, Sara, Aisha Nazli, Gregor Reid, and Charu Kaushic. 2019. "Protective Effect of Probiotic Bacteria and Estrogen in Preventing HIV-1-Mediated Impairment of Epithelial Barrier Integrity in Female Genital Tract" Cells 8, no. 10: 1120. https://doi.org/10.3390/cells8101120