Vitamin B6 and Its Role in Cell Metabolism and Physiology

Hellmann Lab, School of Biological Sciences, College of Liberal Arts and Sciences, Washington State University, Pullman, 99164-6234 WA, USA
*
Author to whom correspondence should be addressed.
Cells 2018, 7(7), 84; https://doi.org/10.3390/cells7070084
Submission received: 28 June 2018
/
Revised: 18 July 2018
/
Accepted: 20 July 2018
/
Published: 22 July 2018
(This article belongs to the Special Issue 2018 Select Papers by Cells’ Editorial Board Members)
Abstract
:Vitamin B6 is one of the most central molecules in cells of living organisms. It is a critical co-factor for a diverse range of biochemical reactions that regulate basic cellular metabolism, which impact overall physiology. In the last several years, major progress has been accomplished on various aspects of vitamin B6 biology. Consequently, this review goes beyond the classical role of vitamin B6 as a cofactor to highlight new structural and regulatory information that further defines how the vitamin is synthesized and controlled in the cell. We also discuss broader applications of the vitamin related to human health, pathogen resistance, and abiotic stress tolerance. Overall, the information assembled shall provide helpful insight on top of what is currently known about the vitamin, along with addressing currently open questions in the field to highlight possible approaches vitamin B6 research may take in the future.
1. Introduction
Vitamin B6 (vitB6), or pyridoxine, is a very important compound for general cellular metabolism [1]. Since its discovery in 1934 by György and colleagues [2], it has been implicated as a co-factor in more than 140 biochemical reactions in the cell [3]. Although most vitB6 co-catalyzed reactions are related to amino acid biosynthesis and catabolism, vitB6 also contributes to fatty acid biosynthesis, breakdown of certain storage compounds in animals and plants, as well as in the biosynthesis of plant hormones, neurotransmitters, and organelle-specific compounds such as chlorophyll [4,5,6,7,8,9,10]. In addition, vitB6 can quench reactive oxygen species (ROS) [11]. Because of its role in ROS scavenging and chlorophyll synthesis, vitB6 is beneficial for photosynthesis, and is discussed as a possible factor to alleviate abiotic and biotic stress [11].
The vitamin comprises a group of six chemically related compounds that all contain a pyridine ring as their core. They differ from each other in a variable group at the pyridine’s 4′ position, which can either be an amino methyl group (pyridoxamine (PM)), a hydroxyl methyl group (pyridoxine (PN)), or an aldehyde (pyridoxal (PL)) (Figure 1). Once the different derivatives are phosphorylated, they can function as co-factors, with pyridoxal 5′-phosphate (PLP) being the biologically active form of vitB6.
In the following review, we will briefly summarize the known biosynthetic pathways in eu- and prokaryotes, and then focus on transport, cellular roles, potential regulatory steps that affect its rate of synthesis, and its importance in human dietary and health issues.
For many years the only biosynthetic pathway known came from the prokaryote Escherichia coli. The pathway is comparably complex, but ultimately leads to the pyridoxine biosynthesis proteins A (PDXA; a 4-hydroxythreonine-4-phosphate dehydrogenase) and PDXJ (a pyridoxine 5′-phosphate synthase) that utilize deoxyxylose 5′-phosphate (DXP) and 4-phospohydroxy-L-threonine to synthesize pyridoxine 5′-phosphate (PNP). A flavin mononucleotide (FMN)-dependent PNP oxidase (PNPox) then converts PNP to PLP (Figure 2) [12,13,14]. An alternative pathway was only relatively recently discovered in fungi; plants; and some bacteria that uses a 3-carbon sugar (either glyceraldehyde 3′-phosphate or dihydroxyacetone phosphate), a pentose-phosphate (either ribose 5′-phosphate or ribulose 5′-phosphate), and glutamine to synthesize PLP [15,16]. Two critical enzymes called PDX1 and PDX2 form a larger PLP synthase complex to catalyze the formation of the vitamin [15,16,17,18]. This newer pathway is commonly referred to as the deoxyxylose 5′-phosphate (DXP)-independent pathway, to distinguish it from the biosynthetic pathway present in E. coli that depends on deoxyxylose 5′-phosphate as a precursor (Figure 2). Of note is that humans and most animals do not encode for any of the de novo enzymes, but rely solely on external supplies of the vitamin from their food. As such, vitB6 is an essential nutrient to humans.
The importance of the de novo pathway is emphasized by various studies in E. coli, Streptococcus pneumonia, Bacillus subtilis, yeast, and plants where its complete loss is lethal to the organism, but can be rescued by an exogenous supply of the vitamin [17,18,19,20,21,22,23,24]. Even a reduction in the biosynthetic efficiency causes severe developmental problems. For example, the plant Arabidopsis thaliana has two functional homologs of PDX1 involved in the de novo biosynthesis of vitB6 [16,18]. Both PDX1 proteins are expressed throughout the plant and appear to be mostly functionally redundant [18,19,25]. However, Arabidopsis null mutants affected in either one of the two PDX1 proteins show severe developmental defects, including stunted root growth, smaller rosette leaves, delayed flowering [17,18], and considerably larger cells, compared with wild-type [26]. Plants also developed larger seeds with increased protein, lipid, and carbohydrate contents [26].
In addition to the two known de novo pathways, most, if not all, organisms have a salvage pathway that is able to convert and phosphorylate the different B6 vitamin derivatives into the catalytically active form, PLP. This is accomplished by the combined activities of at least three enzymes, an oxidase (PDXH), a pyridoxal reductase (PRL), and a kinase (PDXK) (Figure 3) [27,28,29,30]. At least two different kinases have been described that are either specific for PL (further referred to as PLK), or which recognize PN, PM, and PL as substrates (further referred to as PDXK), to generate the phosphorylated B6 vitamers [28,29,31]. The oxidase catalyzes the conversion of PNP and PMP to PLP [32,33], and the reductase forms PN from PL [34].
The salvage pathway is critical to make any non-phosphorylated B6 vitamers available to the cell as co-factors. This is generally relevant for PL, PM, or PN taken up with food and for their recycling in the cell. Consequently, it is essential for animals that depend on the pathway to interconvert the different B6 vitamers into PLP and to make them available as co-factors in enzymatic reactions. In addition, cellular phosphatases have been described that can de-phosphorylate PLP [35]. This may be needed to control the homeostasis of PLP in the cell, and can allow PL catabolism into 4-pyridoxic acid by an aldehyde oxidase [36].
In the liver of mammals, PLP becomes tightly bound to serum albumin as a Schiff base before being secreted into the circulatory blood system for delivery to the different tissues and organs [37,38]. Binding to albumin is discussed as a protective step against early de-phosphorylation of the vitamin. However, before uptake into the cell, PLP needs to be de-phosphorylated to PL by an extracellular, tissue-nonspecific alkaline phosphatase (ALP) [39,40,41]. As ALP only uses free PLP as a substrate, the uptake of PL is likely dependent on the rate PLP dissociates from albumin in the circulatory system.
The salvage pathway is without a doubt vital for animals. Just to provide one example, a genetic study in which the PDXH enzyme was knocked-out via RNAi in Drosophila led to flies that could not survive without additional supplementation of PLP [42]. Even PN was not able to compensate for the loss of the oxidase, corroborating that PLP is indispensable for cell functioning [42]. We are not aware of any described pdxK null mutants in animals (including human), but one can expect that such a mutant would also require a specific PLP-enriched diet to survive.
In comparison to animals, one would assume that organisms that have a de novo pathway do not require the salvage pathway for their survival. In fact, the few described examples show only mild developmental defects, but with a general tendency towards increased stress sensitivity in the affected organisms. For example, in the yeast Saccharomyces cerevisiae, loss of PDXH leads to increased sensitivity to oxidative stress caused by hydrogen peroxide [43]. Likewise, loss of either PDXK or PDXH also results in enhanced stress sensitive phenotypes in Arabidopsis plants [27,44]. Plant mutants affected in PDXH accumulate PNP and PMP with a decrease in PLP, which in turn may also limit PLP-dependent reactions [45]. Surprisingly, Arabidopsis pdxK mutants had nearly nine-fold elevated PLP levels compared with wild type [27]. Because this correlated with a strong up-regulation of the de novo pathway, it suggests that the de novo and the salvage pathways are tightly co-regulated [27]. Interestingly, pdxH mutants also showed mild aberrant developmental features, such as curled leaves or changed flowering time [45], but this was only observed in the absence of ammonium as the nitrogen source. The ammonium-dependency correlated with strongly reduced nitrate reductase (NR) activity in the cells of pdxH mutant plants. NR is required for the conversion of nitrate to ammonium, but may also be inhibited by PMP, as discussed by Colinas and co-workers [45]. Consequently, ammonium requirements in pdxH mutants may be the result of high PMP levels that negatively affect NR activity [45]. Of note is also that in fungi and plants, the salvage pathway might be critical for providing the organelles with a sufficient supply of PLP. While the de novo biosynthetic enzymes, as far as it has been described, are located in the cytoplasm, there is good evidence that salvage pathway enzyme are present in the cytosol and the organelles [43,44,46,47].
2. Complex Organization of PLP Synthases
The PDX1/PDX2 protein pair, which forms a PLP synthase, is widely distributed among archaea, bacteria, fungi, protists, and plants, and the corresponding complexes have been characterized in a variety of pro- and eukaryotes, including Geobacillus stearothermophilus (PdxS/PdxT) [48,49], B. subtilis (Pdx1/Pdx2 or PdxS/PdxT) [50,51], Thermotoga maritima (YaaD/YaaE) [52], S. cerevisiae [52,53], A. thaliana [54], and Plasmodium falciparum [55].
In all of these organisms, the PLP synthase is composed of 12 PDX1 units, and up to 12 PDX2 units. PDX1 proteins form two hexameric rings that interlock into a dodecamer. The hexamers align with each other in a way that a PDX1 protein on one hexamer interacts with two PDX1 molecules from the other hexamer [49,50,55]. This dodecameric double ring system is stable and does not require any specific substrate for PLP synthesis to retain its complex formation [50,55]. In contrast, PDX2s interact with the PDX1 ring only transiently, and this interaction requires the presence of glutamine [50,55]. After the amino group has been removed from glutamine by hydrolysis, the PDX2 proteins are more likely to leave the complex if no other glutamine is readily available for hydrolysis [55].
In plants, two recent studies have shown that PLP synthases contain a lysine swing or lysine relay mechanism that allows intermediate channeling of the substrate in the process of vitB6 biosynthesis [54,56]. Essentially, the lysine residues anchor the substrate to the PLP synthase and facilitate transfer of the substrate between the two active sites efficiently without extra domains or coenzymes [54]. These two sites are present in PDX1 proteins and are designated as P1 and P2 [56]. A lysine residue in the P1 site covalently binds to the pentose phosphate. The pentose phosphate is dephosphorylated, and together with water and ammonia from glutamine, form a chromophoric I320 intermediate with an absorbance maximum at 320 nm. This is shuttled to the P2 site, where it is condensed with a 3-carbon sugar to yield PLP [54,56].
The reaction process of the PLP synthase using the relay mechanism is exciting because, without additional co-enzymes or co-factors, it can perform such a variety of biochemical reactions ranging from isomerization to imine formation, ammonia addition, aldol-type condensation, cyclization, and aromatization [56]. Though both of these studies were conducted in Arabidopsis, this type of relay mechanism is likely conserved among PLP synthases from different organisms, considering the close homology and structural similarity of PDX proteins.
Whether the PLP synthase activity is regulated in response to cellular demands or other environmental or developmental factors is currently not well understood. Two interesting examples, however, come from plants that shed some light on possible regulatory mechanisms. First, A. thaliana, and most likely other plants as well, have a PDX1 protein (in Arabidopsis, this is named PDX1.2), which is not directly involved in the biosynthesis of vitB6 [57]. In recent years, the function of PDX1.2 has been explored, with data indicating that PDX1.2 may serve as a regulator of vitB6 biosynthesis under abiotic stress. PDX1.2 is able to interact with itself and the other PDX1 proteins, but it does not interact with PDX2 [18]. The protein is minimally expressed under normal growth conditions, but certain abiotic stressors, such as heat, strongly induce its expression [27,57,58,59], correlating with an increase in vitB6 biosynthesis [57]. This up-regulation of vitB6 biosynthesis under stress is missing in mutants with reduced PDX1.2 expression, making it overall likely that PDX1.2 acts as a positive regulator of PLP synthase activity in plants [57]. PDX1.2 may also have other functions independent of vitB6 because corresponding null mutants, in which the other PDX1 and 2 proteins are functional, develop an embryo lethal phenotype [58]. However, the precise reason for this developmental defect in these mutants still needs to be resolved.
The second example of a possible regulatory mechanism controlling PLP synthase activity is related to an acetolactate synthase (ALS), which has been found to interact with an Arabidopsis PDX1 protein in yeast-2-hybrid experiments [60]. ALS is the first enzyme in the branched-chain amino acid (BCAA) synthesis pathway, which leads to the production of leucine, isoleucine, and valine, for example. VitB6 is an essential co-factor for branched-chain amino acid transaminase (BCAT), the last step of BCAA synthesis [61]. Given that the results of the yeast-2-hybrid approach can be verified in planta, this finding could represent a novel connection between BCAA and vitB6 biosynthesis [60]. As BCAAs are not only synthesized in plants, but also in various other organisms including fungi, bacteria, and archaea, this potential regulatory interplay may have broader implications. Indeed, YggS, a PLP-binding protein that is widely distributed throughout bacteria, fungi, and eukarya, has been experimentally linked to BCAAs [62]. In an E. coli yggS knockout strain, excess PN led to a toxicity ring (a lack of cell growth on the media in a circle around the PN treatment), while supplementation of either PL or the BCAAs leucine and isoleucine suppressed the PN toxicity [62]. It is suggested that the suppression of PN toxicity by the two BCAAs bypasses the yggS knockout strain’s need for specific PLP-dependent enzymes, such as transaminase B, which catalyzes the last step in leucine and isoleucine synthesis [62].
It would be of interest to know whether PDX1 proteins undergo broader interactions with other PLP-dependent enzymes such as ALS. This may indicate a potential cellular feedback mechanism to either regulate vitB6 demand or for the production of certain amino acids.
3. Regulation of Salvage Pathway Genes
Regulation of the salvage pathway is critical to maintain PLP homeostasis, especially for organisms that lack the de novo biosynthetic pathway. In the following section, some of the known regulatory impacts that occur on the transcriptional and post-translational level are described.
An interesting example of transcriptional regulation comes from Salmonella typhimurium, a pathogenic Gram-negative bacterium that is mainly known for causing typhoid fever [63]. Here, PtsJ, a member of the MocR family of transcription factors, negatively regulates both PdxK and its own expression by binding to the respective promoters [63]. PtsJ contains a helix-turn-helix (HTH) motif for DNA binding at its N-terminal region, while its C-terminal region is related to the type I family of PLP-dependent enzymes, classically represented by aspartate aminotransferases [63,64]. Intriguingly, PtsJ can bind PLP, and this binding further enhances its repressor role in S. typhimurium, providing an elegant mechanism to control PdxK expression in a PLP-dependent manner [63]. Loss of PtsJ strongly up-regulates PdxK expression, but only mildly impacts PdxH [63]. In addition, PtsJ does not bind to the PdxH promoter, suggesting that other mechanisms are in place to control PdxH levels in S. typhimurium [63].
In Bombyx mori caterpillars, PLP levels are discussed to be critical for progression of the different larval stages, and recent research indicates that expression of PDXH and PDXK genes is under the control of the juvenile and molting hormones [65].
In Arabidopsis, one can observe that mutants lacking PDXH, PDXK, or PLR expression of the remaining salvage pathway genes is strongly up-regulated [27,34]. As loss of PDXK also results in higher PLP and total vitB6 levels, the up-regulation of PDXH in a pdxK mutant background is clearly not related to a vitB6 deficiency [34]. In addition, PDXK expression is repressed in roots upon salt stress treatment [44]. PDXH expression is widely inducible by light, heat, and different phytohormones, namely jasmonic acid, ethylene, and abscisic acid treatments. Similar to PDXK, salt treatment results in a down regulation of PDXH, as does exposure to drought [66].
An interesting example of transcriptional regulation comes from the fungal plant pathogen Rhizoctonia solani [67]. Here, a PLR gene was highly inducible by oxidative stress caused by paraquat and phenyl acetic acid treatments, and mildly responsive to hydrogen peroxide exposure [67]. Plants generate hydrogen peroxide as an early defense response upon fungal infections [68,69]. The fungal response pattern resembles that of classical reactive oxygen species (ROS) detoxification enzymes, such as catalase and glutathione-S-transferase [67]. Because de novo vitB6 genes were also up-regulated in the fungus, it is likely that this increase in vitB6 production contributes to quenching of ROS to protect the cell against their harmful impacts, further supporting the fungal effort to infect the plant tissue.
On the protein level, some factors have been described in E. coli and humans that regulate the activities of PDXK and PDXH. PDXK functions as a homodimer and requires mono- and bivalent metal ions such as K+ and Mg2+, respectively, as co-factors [29,70,71,72,73]. For the human PDXK, it has also been shown that the enzyme activity is highly stimulated by Na+, while Zn+ ions at a physiological pH of 7.3 had an inhibitory impact [70]. As K+ is the more prevalent ion under normal physiological conditions in the cell, it is likely that human PDXK acts in a Mg2+/K+ form [70]. Besides Zn+ ions, the activity of PDXK is also inhibited by MgATP. It is discussed that MgATP forms a ternary complex together with PLP at the active site, which reduces PDXK activity [30,74]. PDXH also functions as a homodimer and each subunit binds a single FMN as a co-factor [33,75]. The enzyme is inhibited by its product, PLP, as well as its substrate, PNP, while PMP does not affect its activity [76,77,78,79].
Because of the conserved nature of PDXK and PDXH proteins, one can expect that factors affecting their activities such as ions, PLP, or MgATP are likely very similar in most organisms.
4. Regulation of De Novo VitB6 Synthesis Genes
As indicated above, regulatory steps that directly affect the activity of de novo enzymes involved in vitB6 biosynthesis are not well defined. However, a variety of factors are known that control the expression of genes involved in the de novo pathways. These are mainly related to the overall growth situation of the respective organism, but in many cases, abiotic or biotic stress conditions also cause increased cellular gene expression levels of vitB6 biosynthetic genes. In the following section, we will provide a few examples of conditions regulating gene expression in the two pathways for pro- and eukaryotes.
In E. coli, expression of the PdxA gene is positively regulated when growth rates increase, and this up-regulation depends on Fis, a protein that can interact with and bend DNA, and is known to function as a transcriptional regulator [80,81,82]. Similarly, de novo vitB6 biosynthesis genes in B. subtilis (here called YaaD and E) and the yeast Schizosaccharomyces pombe have been reported to be positively regulated with increasing growth rates [23,83]. In B. subtilis, addition of PN and PL to the growth medium did not repress their expression, indicating that vitB6 is likely not causing a feedback regulatory loop on the transcriptional level [23].
In several prokaryotes that possess the DXP-dependent de novo biosynthesis pathway, such as Bacillus caucillus, Listeria monocytogenes, Corynebacterium glutamicum, and S. pneumoniae, it has been shown that expression of the genes encoding for the PLP synthase depends on PdxR [21,22,84,85]. PdxR is a MocR-related transcription factor that promotes expression of the de novo biosynthesis genes [86]. Like PtsJ, it has an HTH-DNA binding domain and a type I domain for PLP-dependent enzymes [86]. Its activity is down-regulated by binding to PLP, thereby providing, as described for S. typhimurium, a product-based feedback mechanism to control PLP biosynthesis on the transcriptional level in the cell [84].
Previous research has shown that vitB6 affects enzyme induction by steroid hormones [87]. These steroid hormone receptors belong to a superfamily of transcription factors that regulate physiological processes such as growth, development, reproduction, and behavior [88]. Work done by Cake et al. (1978) displayed that PLP could inhibit the binding to DNA-cellulose of the rat-liver glucocorticoid receptor, a receptor to which cortisol and other glucocorticoids bind [88,89]. This receptor was also found in humans and is similarly inhibited by PLP; however, it was also observed that estrogen and androgen receptors were also affected [88]. It is believed that vitB6 modulates the activities of different hormone steroid receptors on the transcriptional level [88]. For this binding to occur on nuclei, chromatin, or DNA, the receptor must undergo “activation” by a physiochemical alteration such as heat or ionic irregularities [89]. It has also been seen that enhanced levels of PLP can decrease the transcriptional response to these receptors, and deficiencies in vitB6 lead to an enhanced responsiveness to steroid hormones [87,88,89].
An interesting regulatory example comes from the bacterium Photorhabdus luminescens, which can switch between two lifestyles; a mutualistic (M) one, where it is in a symbiotic relationship with the nematode Heterorhabditis bacteriophora, and a parasitic (P) one, where it can infect other nematodes like Caenorhabditis elegans. Inversion of a single promoter controls whether P. luminescens assumes either the M or P state, which is followed by metabolic alterations and morphological changes, respectively [90]. As one can also observe different expression levels of P. luminescens’ pdxA and J genes in the M versus the P form (with a ratio of 2.1 for pdxA, and 0.49 for pdxJ, of M to P), the rate of vitB6 biosynthesis is discussed as a potential requirement for the bacterium to adopt the respective lifestyle [91].
A connection to abiotic stress can be found in R. solani, a plant pathogenic fungus, where transcript levels of PDX1 and PDX2 increase significantly when exposed to the superoxide generator paraquat [67]. Increased expression of PDX genes in response to abiotic stress, especially to conditions that increase ROS, has also been reported in other fungi, B. subtilis, and plants, and appears to be of a general nature [25,83,92,93,94,95,96]. These studies provide some evidence that PDXs are not only key proteins necessary for vitB6 biosynthesis, but are also vital factors for abiotic stress tolerance. This notion is corroborated, for example, by pdx1 Arabidopsis plant mutants that are hypersensitive to osmotic and oxidative stress [25]. In fact, Arabidopsis PDX genes are up-regulated by a variety of abiotic stress conditions such as high light, chilling, and drought [59]. The promoters of PDX1 genes further contain a wide set of cis-regulatory elements that are recognized by MYB, MYC, and WRKY transcription factor families, and that are often involved in gene regulation after stress response [97,98,99,100]. PDX1 promoters also contain abscisic acid-responsive elements (ABRE) and ethylene-responsive elements (ERE) [19]. Ethylene and abscisic acid are two classical stress-related phytohormones. In addition, a sugar response elements (SRE) was reported in PDX1 promoters [19], which is also of interest as one of the first plant PDX1 proteins identified came out of a screen for sugar-responsive mutants [18,101]. Although much of these data are based on in silico analysis, the variety of elements detectable in PDX promoters indicates that in plants and other organisms, PLP synthase activity is tightly intertwined with the general cellular metabolism and physiology of the cell.
5. Transport and Distribution of VitB6
For all organisms, it is crucial to have sufficient vitB6 available in the cell. For organisms that lack a de novo biosynthesis pathway, it is further essential that they have an efficient extracellular uptake system in place. However, there are very few known transporters located at the plasma membrane, and even fewer known transporters that are present at the membranes of intracellular compartments [102]. Surprisingly, transporters for vitB6 in eukaryotes have only been described in yeast and plants that already have a de novo pathwfay (Figure 4). In the yeasts S. cerevisiae and S. pombe, two transporters called Tpn1p and Bsu1, respectively, facilitate import of PN, PL, and PM across the plasma membrane [103,104]. Tpn1p belongs to the purine cytosine permease family, and its activity is up-regulated when vitB6 levels decrease in the yeast growth medium [104], although the mechanism of this regulation remains open. Bsu1 shows similarity to the MULTIDRUG RESISTANCE (MDR) family of transport proteins [103]. Of note is that it also facilitates transport of the antihypertensitive drug amiloride that inhibits Na+/K+ transporters [105]. Because amiloride is an effective drug to treat patients that suffer from low levels of K+ in the blood serum (hypokalemia) [105,106,107], it is discussed that an unknown ortholog of BsuI may facilitate vitB6 uptake in humans (Figure 4) [103,108].
Recently, members of the plant purine permease (PUP) family, who were originally identified as transporters for the phytohormone cytokinin, also showed the ability to transport PN, PL, and PM [109,110,111]. Work in A. thaliana on PUP1 showed that only non-phosphorylated vitB6 derivatives are transported, not the active PLP form [111]. Work in tobacco on the PUP member NICOTINE UPTAKE PERMEASE 1 (NUP1) yielded similar results [110]. Interestingly, NUP1 was originally described to facilitate transport of nicotine from the apoplastic space, and to affect alkaloid metabolism. As such, the vitB6 transport added a new and interesting facet to NUP1 function in tobacco. Overall, the precise meaning of plant purine permeases transporting B6 vitamers is not fully resolved. One discussed aspect is that they may allow long-distance distribution of non-phosphorylated B6 vitamers to supply vitB6 taken up by the roots from the soil to other organs [110,111]. Nevertheless, it is unclear to what extent such transport is indeed a critical requirement for plants, as the genes encoding for the DXP-independent pathway are expressed in all tissues [18]. In addition, as outlined above, these PUPs have broad substrate specificity, and are thus likely to be required in the cell for a much wider range of physiological processes than just vitB6 metabolism.
The majority of known vitB6 transporters from yeast and plants are likely located at the plasma membrane (Figure 4). This leaves the question open on how the organelles are supplied with the vitamin. Subcellular localization studies of PDX1 and PDX2 proteins in plants showed that the de novo synthesis via the DXP-independent route most likely takes place only in the cytosol [16]. This is to some degree surprising, as major steps in vitB6-dependent amino acid metabolism are present in mitochondria and chloroplasts [46,112]. It was found that in S. cerevisiae, the mitochondrial carrier protein Mtm1p is a high affinity importer that is responsible for transport of PLP across the mitochondrial membrane [113]. Interestingly, although Mtm1p is not an iron carrier, it plays a role in mitochondrial Fe–S cluster biosynthesis, and its deletion results in defects in iron homeostasis and heme biosynthesis, highlighting a link between PLP transport and mitochondrial iron homeostasis [113]. In plants, it remains ambiguous how chloroplasts are sufficiently supplied with the vitamin, especially because the end-product of the DXP-independent pathway is PLP, a charged compound that cannot passively diffuse across the membrane, and that is not transported by PUPs [111].
As mentioned above, in contrast to the de novo pathway, the salvage pathway in plants, and likely other eukaryotes, appears to take place in the cytosol, as well as in the organelles. At least for PDXH and PDXK from Arabidopsis, localization in the chloroplasts has been reported (Figure 4) [43,47]. If the non-phosphorylated forms of the vitamin can passively diffuse into the chloroplast, the presence of the salvage pathway in the organelle may assure availability of PLP for enzymatic reactions. However, it remains questionable whether a passive system would supply sufficient amounts to satisfy the vitB6 demand in the chloroplast.
Alternatively, the organelles may have yet undescribed vitB6 transporters of their own that differ from the ones present at the plasma membrane. One possible example relevant for eukaryotes comes from the bacterial phylum firmicutes, where a modular built transporter, PdxT, of the ATP BINDING CASSETTE (ABC) family, can transport vitB6 (Figure 4) [114]. The transporter is coupled with a PdxK, likely to directly phosphorylate and activate B6 vitamers after their uptake into the cell [114].
In contrast to the biosynthetic pathways, knowledge explaining transport over long distances and within the cell is currently only poorly understood in eukaryotes and will require more thorough investigations to fully comprehend vitB6 metabolism in the cell.
6. Vitamin B6 and Its Involvement in Cellular Metabolism
As mentioned in the beginning, vitB6 is involved in more than 140 different metabolic reactions in the cell. To provide detailed information about all its functions would exceed the aim of this review. Therefore, in the following section, we will depict a few examples in which PLP has a very crucial role in cellular metabolism and physiology to provide a better appreciation of this vitamin’s role in the cell.
6.1. VitB6 Involvement in Protein Folding
PLP has also been reported to play a role as a chaperone in folding of PLP-enzymes [115]. For instance, cystalysin, a hemolytic protein from Treponema denticola, requires PLP for proper folding and stabilization of the protein [115,116]. The aspartate aminotransferase in E. coli requires PLP for proper stabilization in a native structure, and as the protein disassociates into a partially folded intermediate, PLP is reduced into a pyridoxyllysine (PPL) derivative to ‘lock’ the co-factor in place to further stabilize the protein [117]. Found in E. coli and B. subtilis as well, serine hydroxymethyltransferase (SHMT), which is part of the folate cycle, does not require PLP for folding, but is still required for proper functioning of the protein [115,118]. There is very little known about the function of vitB6 as a chaperone, thus more research is required, although perhaps the lack of research available is indicative of a very limited role.
6.2. VitB6 Involvement in Amino Acid Biosynthesis
Probably the most important function of active, phosphorylated vitB6 in the cell is related to the biosynthesis, as well as the degradation of amino acids. Here, the vitamin is often used for transamination reactions where, by forming an amino acid–PLP Schiff base intermediate, an amino group is transferred to a ketoacid to form new amino acids. In addition, vitB6 is also used for other types of reactions such as α-decarboxylations or racemizations [119].
The impact on amino acid metabolism is exemplified in plants overexpressing a PDX1 protein. Here, significantly increased levels of methionine, glycine, and proline were detected, while there was a decrease in not only γ-amino butyric acid and asparagine, but also in carbohydrates, such as glucose and sucrose [26,120]. In comparison, PDX2 overexpressing plants had decreased levels of β-alanine, arginine, and glutamine, however, there was an increase in proline levels [120]. In plants overexpressing both PDX1.1 and PDX2, a more general, strong increase in sugars and amino acids was detected, along with a decrease in organic acids [26], indicating that an even up-regulation of both PDX1 and PDX2 is a requirement to promote amino acid biosynthesis on a broader scale.
The function of PLP-dependent enzymes in amino acid metabolism also has significant indirect impacts on secondary metabolites that depend on amino acids as their precursors. For example, this is the case for some plant hormones (phytohormones) such as ethylene, auxin, or cytokinin [5,121,122,123]. Auxin biosynthesis requires a tryptophan aminotransferase that catalyzes the step to indole-3-pyruvate, an immediate precursor of the auxin indole-3-acetic acid from L-tryptophan and 2-oxoglutarate [122]. Ethylene biosynthesis depends on the PLP-dependent enzyme ACC (1-aminocyclopropane-1-carboxilic acid) synthase, which generates ACC from S-Adenosyl methionine [5,123], and cytokinin biosynthesis involves lysine decarboxylases, which are known to need PLP as a co-factor [123].
Consequently, it has been demonstrated for auxin and ethylene that homeostasis of these two phytohormones is affected in plant mutants with strongly reduced abilities to synthesize vitB6 [19]. Because auxin is critical for root development, these findings correspond to the shorter root growth seen in vitB6 deficient plants [19].
6.3. VitB6 and Degradation of Cellular Storage Compounds
Glycogen and starch are two major storage compounds in animals, fungi, bacteria, and plants. Glycogen is a branched polymer that mainly consists of α-1,4-glycosidic bonds with every tenth glucose moiety also forming α-1,6-glycosidic bonds. It is required in humans as an energy storage form in brain and muscle cells that can be quickly made available [124]. Starch is a major carbohydrate storage form in plastids, and is also built up by α-1,4- and α-1,6-glycosidic bonds [125]. Glycogen can be made accessible to the cell by glycogen or starch phosphorylase, a PLP-dependent enzyme that hydrolyzes the α-1,4-glycosidic bonds to generate glucose-1-phosphate [81,83,124,126]. In mammals, glycogen phosphorylase is found predominantly in the liver, muscle, and brain [127]. Inhibition of glycogen phosphorylase is discussed as a way to control the glycemic blood sugar levels in affected patients, and a possible mechanism to treat type 2 diabetes [128,129].
6.4. VitB6 and Its Relevance in Tetrapyrrole Biosynthesis
Hemes, cobalamins, and chlorophylls are all tetrapyrroles required for cellular metabolism in animals, plants, and microorganisms, respectively. Heme is an iron-binding prosthetic group in metalloproteins, likely best known for its function in hemoglobins to bind oxygen for long distance transport in erythrocytes [130]. Cobalamin (vitB12) is a cobalt-binding molecule that is ubiquitously required in living organisms for fatty acid and amino acid metabolism, as well as DNA synthesis [131]. Chlorophyll is a magnesium-binding porphyrin that is essential for photosynthesis in plants, algae, and cyanobacteria [132,133]. Biosynthesis of all of these compounds depends on the activity of aminolevulinic acid synthase, a PLP-dependent enzyme that uses succinyl–CoA and glycine to generate δ-aminolevulinic acid, an immediate precursor in tetrapyrrole biosynthesis [134,135].
6.5. VitB6 and Its Role in Neurotransmitter Biosynthesis
Epidemiological surveys have found that a greater intake of foods that contain high amounts of vitB6 correlates with better mental health [136,137,138,139,140]. This may be related to the fact that vitB6-dependent enzymes are needed for the biosynthesis of at least three important neurotransmitters; epinephrine (also known as adrenaline), dopamine (dihydroxyphenethylamine), and serotonin [138] (Figure 5). Epinephrine is critical for acute stress responses (fight-or-flight response) in humans [141] and is classically used as a medication for anaphylaxis, a severe allergic reaction, or cardiac arrest [142,143]. Dopamine can be considered a ‘reward’ neurotransmitter that can regulate positive emotions but also motoric control, and it has been associated with Parkinson’s disease and schizophrenia [144,145,146,147,148]. Serotonin (or 5-hydroxytryptamine) is known as a neurotransmitter that helps patients with depression by promoting well-being and contentment [149,150,151]. In this context, it is of note that depression is predicted to be the number one cause of mental disorders by the World Health Organization by the year 2030. It has become a public health problem here in the United States [152], and multiple studies have shown that deficiencies in vitB6 negatively affect mental health and cognitive abilities [138,140,153,154].
The biosynthesis of epinephrine, serotonin, and dopamine is independently influenced by a universal methyl donor, S-adenosylmethionine (SAM), a critical intermediate in the methionine-cycle [140,155,156,157,158,159,160] (Figure 5). Consequently, several studies have shown that a deficiency in vitB6 correlates with a deficiency in folate and cobalamin [138,140,153,154].
Homocysteine is methylated to methionine by 5-methyltetrahydrofolate, which is needed for the production of purines and thymidylate. Methylation of homocyteine to methionone depends on two other important vitamins, cobalamin (vitB12), and folate (vitB9), where PLP-dependent enzymes such as SHMT, which converts tetrahydrofolate (THF) to 5, 10-methylene tetrahydrofolate, are involved (Figure 6). Elevated levels of homocysteine, otherwise known as hyperhomocysteinemia, are recognized as a cardiovascular risk factor; are a predictor of primary-cause vascular mortality; and have also been associated with mental retardation, seizures, depression, schizophrenia, and cognitive impairment [138,153,160,161]. Homocysteine can be converted in three different ways in the cell. One possibility is the conversion to cysteine via a transulfuration pathway by the PLP-dependent enzyme cystathionine β-lyase. Alternatively, it can be converted via the vitB12-dependent enzyme methionine synthase to methionine. A third option is the PLP-independent reaction to S-adenosyl homocysteine (SAH) by SAH hydrolase (Figure 6). If vitB6 levels are low in the cell, homocysteine is primarily converted to SAH because the transulfuration pathway is affected, as well as tetrapyrrole biosynthesis, which is needed to make vitB12. As a consequence, SAH can accumulate, which inhibits S-adenosyl methionine (SAM)-dependent methylation reactions (Figure 6) [162]. This will cause a decrease in the production of certain neurotransmitters by reducing the synthesis of tetrahydrobiopterin, an essential co-factor for the hydroxylation of phenylalanine and tryptophan that serve as precursors for epinephrine, serotonin, and dopamine [140,163]. It is believed that this accumulation of either homocysteine or SAH, in connection with vitB6 deficiency, is an instigating cause for depressive symptoms and other mental health issues, as well as neurotoxic effects [138,140,154]. This notion is supported by animal studies where supplementation of vitB6 led to higher levels of serotonin in the brain [140].
7. VitB6 Requirements in Human Nutrition
The sections above clearly emphasize the relevance of the vitamin for basic cellular metabolism in all organisms. It is, therefore, of interest to explore to what extent the vitamin is provided in our daily diet, and whether fortification with it may be recommendable. We have previously provided a review on vitB6 and human health [1], and will just provide a few current developments as an update.
The recommended dietary allowances (RDA) for vitB6 provided by the United States Food and Nutrition Board of the Institute of Medicine ranges between 1.3 mg (young adults) to 1.7 mg (adult males), and can reach up to 2 mg for lactating women. Taking these values into consideration, potato, for example, which is a staple food in many countries, represents an excellent nutritional source for this vitamin, as baked potatoes or potato chips contain up to 23% and 60% of the RDA values per 100 g product, respectively (Table 1). Many other products, such as bananas, nuts, and even meat or eggs, contain good to high amounts of the vitamin (Table 1). In addition, the vitamin is heat stable, and processing steps such as cooking or frying do not affect its content. Overall, in a healthy population that has access to a balanced diet, one can emphasize that vitB6 deficiency is uncommon. Although, certain age groups such as older adults (above age 45) have been reported to have significantly lower PLP plasma levels, and may benefit from a fortified vitB6 diet [165,166].
Deficiencies can be caused by high and continuous alcohol consumption and certain antiepileptic drugs, as well as severe renal diseases [167,168,169,170,171,172,173,174,175,176]. In addition, vitB6 deficiency can result from some malabsorption syndromes, such as Crohn’s disease, ulcerative colitis, or celiac disease, and certain genetic disorders such as homocystinuria may cause deficiencies in vitB6 [167,175,177,178,179].
If vitB6 deficiency occurs, it is often associated with dermatitis, microcytic anemia, or electroencephalographic abnormalities [167,175,180,181]. Sometimes, weakening of the immune function, convulsive seizures, and depression and confusion have also been reported [167,175,182,183,184,185,186].
In contrast, overdosage with the vitamin is hard to accomplish. The current daily tolerable upper intake level (UL) for adults (19 years and older) recommended by the Food and Nutrition Board of the Institute of Medicine in the United States is 100 mg [167]. Nevertheless, some case studies have been reported where high dosages of the vitamin caused neurological disorders [167,187,188,189]. It needs to be emphasized that often in these studies the daily levels exceeded the UL by far, and were provided over prolonged periods of time (for example 2000 to 6000 mg/day of pyridoxine for 2 to 40 months described by Schaumburg and co-workers [187]), making it not very likely that one would experience such symptoms under normal conditions [187,190,191,192].
8. VitB6 and the Potential to Develop New Medical Drugs
Knowledge of vitB6 and its related enzymes are used to treat diseases on three levels: supplementation of vitB6, targeting select enzymes in the vitB6 biosynthesis pathways, and targeting PLP-dependent enzymes. There are several reviews out that discuss this topic at length [164,193,194,195,196], so here we will only highlight a few interesting examples.
As vitB6 is important for neurotransmitter biosynthesis, one can anticipate that many neurological disorders such as Parkinson’s, Alzheimer’s, epilepsy, autism, and schizophrenia are affected by PLP availability [164]. In the case of pyridoxine-dependent epilepsy (PDE), recurrent neonatal or infantile seizures are alleviated by high doses of PLP [197]. There are currently no dosing recommendations, though the typical long-term doses of PLP prescribed do not exceed the UL [198]. PDE patients taking doses over 500 mg/day are closely monitored for adverse affects, such as sensory neuropathy [198]. PDE is caused by mutations in the ALDH7A1 (Antiquitin) gene in the lysine degradation pathway, which leads to the build-up of the lysine intermediate L-aminoadipate-semialdehyde (AASA) and its cyclic form, 1-piperideine-6-carboxylic acid (P6C) [199]. P6C can react with and thereby inactivate PLP. It is hypothesized that the subsequent depletion of PLP in the brain causes overexcitement and epileptic seizures due to reduced PLP-dependent biosynthesis of the inhibitory neurotransmitter γ-aminobutyric acid (GABA) [193]. Recently, a zebrafish model for PDE was created, by generation of an aldh7a1 knockout line, that will allow researchers to better study this disease [200]. In another example, dopamine deficiencies are commonly responsible for the primary symptoms of Parkinson’s disease [164]. Looking at neurons of the substantia nigra region in the brain of humans, one of the four differentially expressed genes found was PDXK [164,201]. It is discussed that if PDXK is up-regulated in dopaminergic neurons, which are the main source of dopamine in the midbrain, this might alleviate some symptoms of Parkinson’s disease, although this particular method has not yet been explored [201]. However, other authors have cast doubts on these results because the studies were conducted in isolated populations with the possibility of a high degree of inbreeding due to marrying within a local community, along with the lack of the same differential expression of the four genes, including the pyridoxal kinase locus, in these populations [201,202,203].
In the remaining paragraphs, we will explore the role of pathogens and vitB6, a field that has been rapidly emerging, and that may provide interesting opportunities to develop new strategies to prevent infections.
VitB6 has already been known for quite some time to play a role in pathogen–host interactions. Some early work comes from the phytopathogen Cercospora nicotianae, a fungus that uses a secretable photosensitizer that generates ROS in response to illumination. This production of ROS is needed to facilitate penetration of host plant tissue with the fungal hyphae [204,205]. Intriguingly, a screen for Cercospora mutants sensitive to their own toxin resulted in the identification of fungi that were affected in vitB6 biosynthesis [206,207,208]. This was actually a first discovery that vitB6 is a very potent antioxidant that can efficiently protect against increasing ROS in the cell [207]. It was also a first discovery that it plays a critical role in the host–pathogen interplay.
Plants on the other hand also appear to take advantage of vitB6, but rather as a way to reduce their infection risk by pathogens. This notion is supported by a recent example from Arabidopsis, where mutants affected in de novo and salvage pathway genes displayed significantly higher susceptibility to infection by the bacterium Pseudomonas syringae pv. tomato (Pst) and the fungus Botrytis cinerea [209]. However, it remains open whether increased susceptibility of the plants was caused by a generally reduced metabolic rate, and thus lowered cellular fitness, or whether vitB6 is critical to induce an effective cellular defense response against a pathogen.
Another interesting example stems from the previously mentioned bacterium P. luminescens that symbiotically associates with the entomopathogenic nematode H. bacteriophora [210]. In the absence of this symbiotic relationship, P. luminescens is infectious to insects and other nematodes. A genetic screen for virulent deficient P. luminescens mutants found that reduced pathogenicity can be caused by a mutation in PdxB, an erythronate-4-phosphate dehydrogenase, which is part of the DXP-dependent de novo pathway [211]. pdxB mutants show an overall poor growth, which can be restored by supplementing growth media with PLP, but also with non-phosphorylated vitB6, demonstrating that the bacterium has efficient vitB6 uptake systems and a functional salvage pathway [211]. However, although the precise impact of vitB6 for pathogenicity is not clear at this point, a defined metabolic rate with sufficient PLP levels is discussed as a requirement to support either the mutualistic lifestyle or the ability to infect other organisms [211].
A correlation between vitB6 biosynthesis and pathogenicity has also been reported for three other bacteria, Helicobacter pylori, S. pneumonia, and Mycobacterium tuberculosis [22,212,213]. H. pylori is a flagellated bacterium that can cause stomach infections. Reduced activity in the vitB6 de novo synthesis pathway reduces the bacterium’s virulence and also impairs its ability to make glycosylated flagella, resulting in immobile bacteria cells [212]. S. pneumoniae is a Gram-positive bacterium that is responsible for a variety of severe infections, including pneumonia or meningitis. Loss in de novo biosynthesis genes did not fully prevent pneumonia infection in a mouse model, but it was able to produce significantly attenuated infection rates compared with wild type [22].
Similarly, loss of the de novo pathway in the tuberculosis-causing bacterium, M. tuberculosis, prevents the pathogen from persisting in host tissue [213]. Interestingly, apart from interfering with vitB6 biosynthesis directly in M. tuberculosis, it has also been found that the vitamin itself, taken as a supplement or added to a treatment, can improve the efficacy of tuberculosis treatments. For example, vitB6 (along with 11 other vitamins) was tested as a cofactor for MtbLrpA, a transcriptional regulator that is thought to play an important role in the persistence of M. tuberculosis, and was found to inhibit MtbLrpA binding to DNA [214]. In other words, more vitB6 means more inhibition of MtbLrpA, which in turn causes reduced persistence of the bacterium. In another example, when the M. tuberculosis pdx1 loss of function mutation was added to a tuberculosis vaccine background, as long as supplemental vitB6 was provided, the resulting vaccine was found to be safer for immunodeficient mice and profoundly more effective than the vaccine strain without mutated pdx1 [215].
All of these examples led to the discussion of whether the vitB6 pathway could be used as a potential drug target to develop novel medications for preventing infections by the mentioned bacteria and other pathogens. The absence of the de novo vitB6 biosynthesis pathway in humans makes this a very promising approach; because in order for a potential drug to be effective, it needs to (1) be specific to the pathogen’s enzymes and metabolism without also targeting the host’s processes or causing undue damage to the host, and (2) interfere with or inhibit the pathogen’s metabolic process to such an extent that it cannot survive, reducing or eliminating the pathogen load so that the host can recover. A drug that targets PDX proteins and selective PLP-dependent enzymes that are necessary for pathogen survival can conceivably meet both of these requirements. In this context, we will mention malaria as a eukaryotic pathogen that may be a promising target for development of novel anti-malaria drugs utilizing the de novo vitB6 pathway.
Malaria, which, in severe cases, can cause death, is a devastating disease that continues to affect millions of people worldwide, especially around the equator. There were 214 million new cases of malaria and 438,000 deaths due to the disease in 2015 according to World Health Organization (WHO) estimates [216]. A major problem in the fight against malaria is the growing disease resistance of Plasmodium falciparum to the current anti-malarial drugs, requiring researchers to continually look for new drug targets. A 2014 review identified the PLP-dependent enzymes ornithine decarboxylase (ODX), the P. falciparum aspartate aminotransferase (AAT), and SHMT as potential targets, however, the corresponding host enzymes might also be affected, which must be taken into consideration [132,195]. Kronenburger and coworkers also suggested targeting PLP-related molecules in the Anopheles vector, such as 3-hydroxykynurenine transaminase (HKT), but more information on the structure of HKT is needed to be able to design a drug that targets it. We are not aware of any more recent work that sheds light on the structure of HKT or any drugs that have been developed specifically against vitB6 de novo enzymes. There may be other candidate targets to fight malaria that are currently more appealing to pursue, however, it is likely an oversight if these potential vitB6-related targets are abandoned as a tool to develop novel treatments against P. falciparum and other pathogens.
9. Conclusions
More than 80 years have passed since vitB6 was first described. Since then, we have come to understand how it is synthesized in the cell and its requirement in numerous biochemical reactions. There is also a growing amount of knowledge about the biological and physiological processes it affects across pro- and eukaryotic species. Yet there are still many aspects of the vitamin that need to be explored. Because it is required for so many processes, ranging from amino acid metabolism to stability of certain storage compounds or the biosynthesis of other vitamins, one would expect that its own biosynthesis and homeostasis is tightly controlled in the cell. However, regulatory aspects that control the homeostasis of vitB6 in the cell, on both the transcriptional and the post-translational level, are not well defined. Likewise, little knowledge has been generated about the cross-talk between the salvage and de novo pathways, and how catabolic processes may play a role in controlling vitB6 levels in the cell. We also have a poor understanding of how the biosynthesis of vitB6 is regulated in the metabolic network of a cell in context with environmental conditions, or in relation to developmental aspects. Another underexplored area is its subcellular and long-distance distribution and transport, which will be key to fully understanding the vitamin’s role in the cell.
The many ‘indirect’ aspects that appear to be affected by the vitamin, such as health, abiotic stress tolerance, pathogen susceptibility, or even the virulence of a pathogen, are very promising areas for vitB6 research. Investigations in these fields may lead to new findings and applications that result in novel treatments and medications to cope with stress, depression, or infections.
One interesting aspect to potentially explore deeper could be to use plants as primary producers of the vitamin to cope with potential deficiencies in the human diet, like with the ‘Golden Rice’ [217], where biofortification of rice plants with carotenoids was meant as a tool to improve the nutritional quality. Because UL levels are very high, it is unlikely that one would observe toxic side effects caused by such an approach. In fact, current literature shows that bioengineered plants with elevated vitB6 levels did not exceed, for example, 6 ng/mg fresh weight [26]. Rather, because vitB6 and other essential vitamins have such profound impacts on human health, and can be de novo synthesized by plants, phytonutrient biofortification could be a promising route in the future. In addition, beneficial impacts have also been described for the plants themselves, such as improved abiotic stress tolerance and increased seed size, and likely also affect pathogen tolerance, which are highly valuable traits for agriculture [26,209].
In conclusion, it will be interesting to see how the vitB6 field continues to develop, and what novel findings will add to the understanding and utilization of this important and ubiquitous vitamin.
Funding
This research received no external funding. This research received no external funding.
Acknowledgement
We would like to thank Sutton Mooney for critical reading and commenting on the manuscript and WSU for continuously supporting M.P. and S.S through a teaching fellowship.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| AASA | L-aminoadipate-semialdehyde |
| AAT | aspartate aminotransferase |
| ABC | ATP BINDING CASSETTE |
| ABRE | abscisic acid-responsive elements |
| ACC | 1-aminocyclopropane-1-carboxilic acid |
| ALP | alkaline phosphatase |
| ALS | acetolactate synthase |
| BCAA | branched-chain amino acid |
| BCAT | branched-chain amino acid transaminase |
| BH4 | tetrahydrobiopterin |
| CBS | cystathionine-β-synthase |
| DAP | deoxyxylose 5′-phosphate |
| DXP | deoxyxylose 5′-phosphate |
| ERE | ethylene-responsive elements |
| FMN | flavin mononucleotide |
| GAP | glyceraldehyde-3-phosphate |
| GLN | L-glutamine |
| GLU | L-glutamate |
| HKT | 3-hydroxykynurenine transaminase |
| HTH | helix-turn-helix |
| MAT | methionine adenosyltransferase |
| MS | methionine synthase |
| MT | methyltransferase |
| MTHFR | methylenetetrahydrofolate reductase |
| NR | nitrate reductase |
| NUP1 | NICOTINE UPTAKE PERMEASE 1 |
| ODX | ornithine decarboxylase |
| P6C | 1-piperideine-6-carboxylic acid |
| PDE | pyridoxine-dependent epilepsy |
| PDXA | Pyridoxine Biosynthesis Proteins A |
| PDXH | pyridoxine/pyridoxamine 5′-phosphate oxidase |
| PDXJ | pyridoxine 5′-phosphate synthase |
| PDXK | pyridoxal kinase |
| PHT | 4-phosphohydroxy-L-threonine |
| PL | pyridoxal |
| PLK | pyridoxal kinase specific to PL |
| PLP | pyridoxal 5′-phosphate |
| PLR | pyridoxal reductase |
| PM | pyridoxamine |
| PMP | pyridoxamine 5′-phosphate |
| PN | pyridoxine |
| PNP | pyridoxine 5′-phosphate |
| PNPox | pyridoxine 5′-phosphate oxidase |
| PPL | pyridoxyllysine |
| PUP | purine permease |
| RDA | recommended dietary allowances |
| RIP | ribose-5-phosphate |
| ROS | reactive oxygen species |
| RUP | ribulose-5-phosphate |
| SAH | S-adenosylhomocysteine |
| SAHH | S-adenosylhomocysteine hydrolase |
| SAM | S-adenosylmethionine |
| SHMT | serine hydroxymethyltransferase |
| THF | tetrahydrofolate |
| UL | tolerable upper intake levels |
| vitB12 | vitamin B12 |
| vitB6 | vitamin B6 |
References
- Hellmann, H.; Mooney, S. Vitamin B6: A molecule for human health? Mol. Basel Switz. 2010, 15, 442–459. [Google Scholar] [CrossRef] [PubMed]
- Birch, T.W.; György, P.; Harris, L.J. The vitamin B(2) complex. Differentiation of the antiblacktongue and the “P.-P.” factors from lactoflavin and vitamin B(6) (so-called “rat pellagra” factor). Parts I-VI. Biochem. J. 1935, 29, 2830–2850. [Google Scholar] [CrossRef] [PubMed]
- Mooney, S.; Leuendorf, J.-E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A Long Known Compound of Surprising Complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, J.B.; George, F.; Audhya, T. Abnormally high plasma levels of vitamin B6 in children with autism not taking supplements compared to controls not taking supplements. J. Altern. Complement. Med. (N. Y. NY) 2006, 12, 59–63. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Singh, S.K.; Roy, S.; Sengupta, D.N. An insight into the sequential, structural and phylogenetic properties of banana 1-aminocyclopropane-1-carboxylate synthase 1 and study of its interaction with pyridoxal-5′-phosphate and aminoethoxyvinylglycine. J. Biosci. 2010, 35, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Seijo, J.A.; Ruzanski, C.; Krucewicz, K.; Meier, S.; Hägglund, P.; Svensson, B.; Palcic, M.M. Functional and structural characterization of plastidic starch phosphorylase during barley endosperm development. PLoS ONE 2017, 12, e0175488. [Google Scholar] [CrossRef] [PubMed]
- Geng, M.Y.; Saito, H.; Katsuki, H. Effects of vitamin B6 and its related compounds on survival of cultured brain neurons. Neurosci. Res. 1995, 24, 61–65. [Google Scholar] [CrossRef]
- Plecko, B.; Stöckler, S. Vitamin B6 dependent seizures. Can. J. Neurol. Sci. 2009, 36 (Suppl. 2), S73–S77. [Google Scholar] [PubMed]
- Tsang, E.W.T.; Hu, Z.; Chang, Q.; McGregor, D.I.; Keller, W.A. Expression of a Brassic napus glutamate 1-semialdehyde aminotransferase in Escherichia coli and characterization of the recombinant protein. Protein Expr. Purif. 2003, 29, 193–201. [Google Scholar] [CrossRef]
- Ercan-Fang, N.; Taylor, M.R.; Treadway, J.L.; Levy, C.B.; Genereux, P.E.; Gibbs, E.M.; Rath, V.L.; Kwon, Y.; Gannon, M.C.; Nuttall, F.Q. Endogenous effectors of human liver glycogen phosphorylase modulate effects of indole-site inhibitors. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E366–E372. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M.; Ksas, B.; Szewczyk, A.; Rumeau, D.; Franck, F.; Caffarri, S.; Triantaphylidès, C. Vitamin B6 deficient plants display increased sensitivity to high light and photo-oxidative stress. BMC Plant Biol. 2009, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Drewke, C.; Klein, M.; Clade, D.; Arenz, A.; Müller, R.; Leistner, E. 4-O-phosphoryl-L-threonine, a substrate of the pdxC(serC) gene product involved in vitamin B6 biosynthesis. FEBS Lett. 1996, 390, 179–182. [Google Scholar] [CrossRef]
- Notheis, C.; Drewke, C.; Leistner, E. Purification and characterization of the pyridoxol-5′-phosphate:oxygen oxidoreductase (deaminating) from Escherichia coli. Biochim. Biophys. Acta 1995, 1247, 265–271. [Google Scholar] [CrossRef]
- Mittenhuber, G. Phylogenetic analyses and comparative genomics of vitamin B6 (pyridoxine) and pyridoxal phosphate biosynthesis pathways. J. Mol. Microbiol. Biotechnol. 2001, 3, 1–20. [Google Scholar] [PubMed]
- Tambasco-Studart, M.; Tews, I.; Amrhein, N.; Fitzpatrick, T.B. Functional analysis of PDX2 from Arabidopsis, a glutaminase involved in vitamin B6 biosynthesis. Plant Physiol. 2007, 144, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Tambasco-Studart, M.; Titiz, O.; Raschle, T.; Forster, G.; Amrhein, N.; Fitzpatrick, T.B. Vitamin B6 biosynthesis in higher plants. Proc. Natl. Acad. Sci. USA 2005, 102, 13687–13692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titiz, O.; Tambasco-Studart, M.; Warzych, E.; Apel, K.; Amrhein, N.; Laloi, C.; Fitzpatrick, T.B. PDX1 is essential for vitamin B6 biosynthesis, development and stress tolerance in Arabidopsis. Plant J. Cell Mol. Biol. 2006, 48, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Bernhardt, A.; Leuendorf, J.E.; Drewke, C.; Lytovchenko, A.; Mujahed, N.; Gurgui, C.; Frommer, W.B.; Leistner, E.; Fernie, A.R.; et al. Analysis of the Arabidopsis rsr4-1/pdx1-3 mutant reveals the critical function of the PDX1 protein family in metabolism, development, and vitamin B6 biosynthesis. Plant Cell 2006, 18, 1722–1735. [Google Scholar] [CrossRef] [PubMed]
- Boycheva, S.; Dominguez, A.; Rolcik, J.; Boller, T.; Fitzpatrick, T.B. Consequences of a Deficit in Vitamin B6 Biosynthesis de Novo for Hormone Homeostasis and Root Development in Arabidopsis. Plant Physiol. 2015, 167, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Nakamura, Y.; Dong, Y.-X.; Nikawa, J.; Sueda, S. Pyridoxine biosynthesis in yeast: Participation of ribose 5-phosphate ketol-isomerase. Biochem. J. 2004, 379, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Jochmann, N.; Götker, S.; Tauch, A. Positive transcriptional control of the pyridoxal phosphate biosynthesis genes pdxST by the MocR-type regulator PdxR of Corynebacterium glutamicum ATCC 13032. Microbiology (Read. Engl.) 2011, 157, 77–88. [Google Scholar] [CrossRef] [PubMed]
- El Qaidi, S.; Yang, J.; Zhang, J.-R.; Metzger, D.W.; Bai, G. The vitamin B6 biosynthesis pathway in Streptococcus pneumoniae is controlled by pyridoxal 5′-phosphate and the transcription factor PdxR and has an impact on ear infection. J. Bacteriol. 2013, 195, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Kita, M.; Katsuragi, T.; Ogasawara, N.; Tani, Y. yaaD and yaaE are involved in vitamin B6 biosynthesis in Bacillus subtilis. J. Biosci. Bioeng. 2002, 93, 309–312. [Google Scholar] [CrossRef]
- Apostolakos, D.; Birge, E.A. A thermosensitive pdxJ mutation affecting vitamin B6 biosynthesis in Escherichia coli K-12. Curr. Microbiol. 1979, 2, 39. [Google Scholar] [CrossRef]
- Chen, H.; Xiong, L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005, 44, 396–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raschke, M.; Boycheva, S.; Crèvecoeur, M.; Nunes-Nesi, A.; Witt, S.; Fernie, A.R.; Amrhein, N.; Fitzpatrick, T.B. Enhanced levels of vitamin B(6) increase aerial organ size and positively affect stress tolerance in Arabidopsis. Plant J. 2011, 66, 414–432. [Google Scholar] [CrossRef] [PubMed]
- González, E.; Danehower, D.; Daub, M.E. Vitamer Levels, Stress Response, Enzyme Activity, and Gene Regulation of Arabidopsis Lines Mutant in the Pyridoxine/Pyridoxamine 5′-Phosphate Oxidase (PDX3) and the Pyridoxal Kinase (SOS4) Genes Involved in the Vitamin B6 Salvage Pathway. Plant Physiol. 2007, 145, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tsui, H.C.; Man, T.K.; Winkler, M.E. Identification and function of the pdxY gene, which encodes a novel pyridoxal kinase involved in the salvage pathway of pyridoxal 5′-phosphate biosynthesis in Escherichia coli K-12. J. Bacteriol. 1998, 180, 1814–1821. [Google Scholar] [PubMed]
- Tang, L.; Li, M.-H.; Cao, P.; Wang, F.; Chang, W.-R.; Bach, S.; Reinhardt, J.; Ferandin, Y.; Galons, H.; Wan, Y.; et al. Crystal structure of pyridoxal kinase in complex with roscovitine and derivatives. J. Biol. Chem. 2005, 280, 31220–31229. [Google Scholar] [CrossRef] [PubMed]
- Di Salvo, M.L.; Contestabile, R.; Safo, M.K. Vitamin B(6) salvage enzymes: Mechanism, structure and regulation. Biochim. Biophys. Acta 2011, 1814, 1597–1608. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, G.; Winkler, M.E. Identification of the pdxK gene that encodes pyridoxine (vitamin B6) kinase in Escherichia coli K-12. FEMS Microbiol. Lett. 1996, 141, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Biswal, B.K.; Au, K.; Cherney, M.M.; Garen, C.; James, M.N.G. The molecular structure of Rv2074, a probable pyridoxine 5′-phosphate oxidase from Mycobacterium tuberculosis, at 1.6 angstroms resolution. Acta Crystallograph. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Safo, M.K.; Musayev, F.N.; Schirch, V. Structure of Escherichia coli pyridoxine 5′-phosphate oxidase in a tetragonal crystal form: Insights into the mechanistic pathway of the enzyme. Acta Crystallogr. D Biol. Crystallogr. 2005, 61, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Herrero, S.; González, E.; Gillikin, J.W.; Vélëz, H.; Daub, M.E. Identification and characterization of a pyridoxal reductase involved in the vitamin B6 salvage pathway in Arabidopsis. Plant Mol. Biol. 2011, 76, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, E.; Nocek, B.; Brown, G.; Makarova, K.S.; Flick, R.; Wolf, Y.I.; Khusnutdinova, A.; Evdokimova, E.; Jin, K.; Tan, K.; et al. Functional Diversity of Haloacid Dehalogenase Superfamily Phosphatases from Saccharomyces cerevisiae: Biochemical, structural, and evolutionary insights. J. Biol. Chem. 2015, 290, 18678–18698. [Google Scholar] [CrossRef] [PubMed]
- Ueland, P.M.; Ulvik, A.; Rios-Avila, L.; Midttun, Ø.; Gregory, J.F. Direct and Functional Biomarkers of Vitamin B6 Status. Annu. Rev. Nutr. 2015, 35, 33–70. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, M.C.; Sampson, D.A.; Skala, J.H.; Gietzen, D.W.; Grier, R.E. Evaluation of vitamin B-6 status and function of rats fed excess pyridoxine. J. Nutr. 1989, 119, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Merrill, A.H.; Horiike, K.; McCormick, D.B. Evidence for the regulation of pyridoxal 5′-phosphate formation in liver by pyridoxamine (pyridoxine) 5′-phosphate oxidase. Biochem. Biophys. Res. Commun. 1978, 83, 984–990. [Google Scholar] [CrossRef]
- Narisawa, S.; Wennberg, C.; Millán, J.L. Abnormal vitamin B6 metabolism in alkaline phosphatase knock-out mice causes multiple abnormalities, but not the impaired bone mineralization. J. Pathol. 2001, 193, 125–133. [Google Scholar] [CrossRef]
- Whyte, M.P.; Mahuren, J.D.; Vrabel, L.A.; Coburn, S.P. Markedly increased circulating pyridoxal-5′-phosphate levels in hypophosphatasia. Alkaline phosphatase acts in vitamin B6 metabolism. J. Clin. Investig. 1985, 76, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Whyte, M.P.; Mahuren, J.D.; Fedde, K.; Cole, F.; McCabe, E.R.B.; Coburn, S.P. Perinatal hypophosphatasia: Tissue levels of vitamin B6 are unremarkable despite markedly increased circulating concentrations of pyridoxal-5′-phosphate. Evidence for an ectoenzyme role for tissue-nonspecific alkaline phosphatase. J. Clin. Investig. 1988, 81, 1234–1239. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Zhang, L.; Du, W.; Zhuang, X. A nutritional conditional lethal mutant due to pyridoxine 5′-phosphate oxidase deficiency in Drosophila melanogaster. G3 Genes Genomes Genet. 2014, 4, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Barbosa, J.M.; Wu, H.; Locy, R.D.; Singh, N.K. Identification of a pyridoxine (pyridoxamine) 5′-phosphate oxidase from Arabidopsis thaliana. FEBS Lett. 2007, 581, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Xiong, L.; Stevenson, B.; Lu, T.; Zhu, J.-K. The Arabidopsis salt overly sensitive 4 Mutants Uncover a Critical Role for Vitamin B6 in Plant Salt Tolerance. Plant Cell 2002, 14, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Colinas, M.; Eisenhut, M.; Tohge, T.; Pesquera, M.; Fernie, A.R.; Weber, A.P.M.; Fitzpatrick, T.B. Balancing of B6 Vitamers Is Essential for Plant Development and Metabolism in Arabidopsis. Plant Cell 2016, 28, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.R.; Leech, R.M. Amino Acid Biosynthesis by Isolated Chloroplasts during Photosynthesis. Plant Physiol. 1972, 50, 228–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueschhoff, E.E.; Gillikin, J.W.; Sederoff, H.W.; Daub, M.E. The SOS4 pyridoxal kinase is required for maintenance of vitamin B6-mediated processes in chloroplasts. Plant Physiol. Biochem. 2013, 63, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Brown, W.C.; Harms, E.; Smith, J.L. Crystal Structures Capture Three States in the Catalytic Cycle of a Pyridoxal Phosphate (PLP) Synthase. J. Biol. Chem. 2015, 290, 5226–5239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Burgner, J.W.; Harms, E.; Belitsky, B.R.; Smith, J.L. A new arrangement of (beta/alpha)8 barrels in the synthase subunit of PLP synthase. J. Biol. Chem. 2005, 280, 27914–27923. [Google Scholar] [CrossRef] [PubMed]
- Strohmeier, M.; Raschle, T.; Mazurkiewicz, J.; Rippe, K.; Sinning, I.; Fitzpatrick, T.B.; Tews, I. Structure of a bacterial pyridoxal 5′-phosphate synthase complex. Proc. Natl. Acad. Sci. USA 2006, 103, 19284–19289. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R. Physical and enzymological interaction of Bacillus subtilis proteins required for de novo pyridoxal 5′-phosphate biosynthesis. J. Bacteriol. 2004, 186, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Zein, F.; Zhang, Y.; Kang, Y.-N.; Burns, K.; Begley, T.P.; Ealick, S.E. Structural insights into the mechanism of the PLP synthase holoenzyme from Thermotoga maritima. Biochemistry (Mosc.) 2006, 45, 14609–14620. [Google Scholar] [CrossRef] [PubMed]
- Neuwirth, M.; Strohmeier, M.; Windeisen, V.; Wallner, S.; Deller, S.; Rippe, K.; Sinning, I.; Macheroux, P.; Tews, I. X-ray crystal structure of Saccharomyces cerevisiae Pdx1 provides insights into the oligomeric nature of PLP synthases. FEBS Lett. 2009, 583, 2179–2186. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.C.; Kaufmann, M.; Roux, C.; Fitzpatrick, T.B. Structural definition of the lysine swing in Arabidopsis thaliana PDX1: Intermediate channeling facilitating vitamin B6 biosynthesis. Proc. Natl. Acad. Sci. USA 2016, 113, E5821–E5829. [Google Scholar] [CrossRef] [PubMed]
- Guédez, G.; Hipp, K.; Windeisen, V.; Derrer, B.; Gengenbacher, M.; Böttcher, B.; Sinning, I.; Kappes, B.; Tews, I. Assembly of the eukaryotic PLP-synthase complex from Plasmodium and activation of the Pdx1 enzyme. Structure 2012, 20, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.J.; Windeisen, V.; Zhang, Y.; Guédez, G.; Weber, S.; Strohmeier, M.; Hanes, J.W.; Royant, A.; Evans, G.; Sinning, I.; et al. Lysine relay mechanism coordinates intermediate transfer in vitamin B6 biosynthesis. Nat. Chem. Biol. 2017, 13, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Moccand, C.; Boycheva, S.; Surriabre, P.; Tambasco-Studart, M.; Raschke, M.; Kaufmann, M.; Fitzpatrick, T.B. The Pseudoenzyme PDX1.2 Boosts Vitamin B6 Biosynthesis under Heat and Oxidative Stress in Arabidopsis. J. Biol. Chem. 2014, 289, 8203–8216. [Google Scholar] [CrossRef] [PubMed]
- Leuendorf, J.E.; Mooney, S.L.; Chen, L.; Hellmann, H.A. Arabidopsis thaliana PDX1.2 is critical for embryo development and heat shock tolerance. Planta 2014, 240, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Denslow, S.A.; Rueschhoff, E.E.; Daub, M.E. Regulation of the Arabidopsis thaliana vitamin B6 biosynthesis genes by abiotic stress. Plant Physiol. Biochem. 2007, 45, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Dezfulian, M.H.; Foreman, C.; Jalili, E.; Pal, M.; Dhaliwal, R.K.; Roberto, D.K.A.; Imre, K.M.; Kohalmi, S.E.; Crosby, W.L. Acetolactate synthase regulatory subunits play divergent and overlapping roles in branched-chain amino acid synthesis and Arabidopsis development. BMC Plant Biol. 2017, 17, 71. [Google Scholar] [CrossRef] [PubMed]
- Amorim Franco, T.M.; Hegde, S.; Blanchard, J.S. Chemical Mechanism of the Branched-Chain Aminotransferase IlvE from Mycobacterium tuberculosis. Biochemistry (Mosc.) 2016, 55, 6295–6303. [Google Scholar] [CrossRef] [PubMed]
- Prunetti, L.; El Yacoubi, B.; Schiavon, C.R.; Kirkpatrick, E.; Huang, L.; Bailly, M.; ElBadawi-Sidhu, M.; Harrison, K.; Gregory, J.F.; Fiehn, O.; et al. Evidence That COG0325 Proteins are involved in PLP Homeostasis. Microbiology (Read. Engl.) 2016, 162, 694–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tramonti, A.; Milano, T.; Nardella, C.; di Salvo, M.L.; Pascarella, S.; Contestabile, R. Salmonella typhimurium PtsJ is a novel MocR-like transcriptional repressor involved in regulating the vitamin B6 salvage pathway. FEBS J. 2017, 284, 466–484. [Google Scholar] [CrossRef] [PubMed]
- Milano, T.; Contestabile, R.; Lo, A.P.; Ciccozzi, M.; Pascarella, S. The aspartate aminotransferase-like domain of Firmicutes MocR transcriptional regulators. Comput. Biol. Chem. 2015, 58, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Yang, H.; Yao, L.; Zhang, J.; Huang, L. Effect of exogenous hormones on transcription levels of pyridoxal 5′-phosphate biosynthetic enzymes in the silkworm (Bombyx mori). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 194–195, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Locy, R.D.; Goertzen, L.R.; Rashotte, A.M.; Si, Y.; Kang, K.; Singh, N.K. Expression, in vivo localization and phylogenetic analysis of a pyridoxine 5′-phosphate oxidase in Arabidopsis thaliana. Plant Physiol. Biochem. 2011, 49, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Samsatly, J.; Chamoun, R.; Gluck-Thaler, E.; Jabaji, S. Genes of the de novo and Salvage Biosynthesis Pathways of Vitamin B6 are Regulated under Oxidative Stress in the Plant Pathogen Rhizoctonia solani. Front. Microbiol. 2015, 6, 1429. [Google Scholar] [CrossRef] [PubMed]
- Govrin, E.M.; Levine, A. The hypersensitive response facilitates plant infection by the necrotrophic pathogen Botrytis cinerea. Curr. Biol. 2000, 10, 751–757. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Di, M.S.; Hunt, S.; Schirch, V. Expression, purification, and kinetic constants for human and Escherichia coli pyridoxal kinases. Protein Expr. Purif. 2004, 36, 300–306. [Google Scholar] [CrossRef]
- Li, M.; Kwok, F.; Chang, W.; Liu, S.; Lo, S.C.L.; Zhang, J.; Jiang, T.; Liang, D. Conformational Changes in the Reaction of Pyridoxal Kinase. J. Biol. Chem. 2004, 279, 17459–17465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musayev, F.N.; di Salvo, M.L.; Ko, T.-P.; Gandhi, A.K.; Goswami, A.; Schirch, V.; Safo, M.K. Crystal Structure of human pyridoxal kinase: Structural basis of M+ and M2+ activation. Protein Sci. Publ. Protein Soc. 2007, 16, 2184–2194. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Gong, Y.; Tang, L.; Leung, Y.-C.; Jiang, T. Crystal structure of human pyridoxal kinase. J. Struct. Biol. 2006, 154, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Safo, M.K.; Musayev, F.N.; di Salvo, M.L.; Hunt, S.; Claude, J.-B.; Schirch, V. Crystal structure of pyridoxal kinase from the Escherichia coli pdxK gene: Implications for the classification of pyridoxal kinases. J. Bacteriol. 2006, 188, 4542–4552. [Google Scholar] [CrossRef] [PubMed]
- Musayev, F.N.; Di Salvo, M.L.; Ko, T.-P.; Schirch, V.; Safo, M.K. Structure and properties of recombinant human pyridoxine 5′-phosphate oxidase. Protein Sci. Publ. Protein Soc. 2003, 12, 1455–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Winkler, M.E. Kinetic limitation and cellular amount of pyridoxine (pyridoxamine) 5′-phosphate oxidase of Escherichia coli K-12. J. Bacteriol. 1995, 177, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.-F.; di Salvo, M.; Schirch, V. Distribution of B6 Vitamers in Escherichia coli as Determined by Enzymatic Assay. Anal. Biochem. 2001, 298, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Churchich, J.E.; Zaiden, E.; Kwok, F. Brain pyridoxine-5-phosphate oxidase. Modulation of its catalytic activity by reaction with pyridoxal 5-phosphate and analogs. J. Biol. Chem. 1987, 262, 12013–12017. [Google Scholar] [PubMed]
- Choi, J.-D.; Bowers-Komro, M.; Davis, M.D.; Edmondson, D.; Mccormick, D. Kinetic properties of pyridoxamie (pyridoxine) 5′-phosphate oxidase from rabbit liver. J. Biol. Chem. 1983, 258, 840–845. [Google Scholar] [PubMed]
- Pease, A.J.; Roa, B.R.; Luo, W.; Winkler, M.E. Positive growth rate-dependent regulation of the pdxA, ksgA, and pdxB genes of Escherichia coli K-12. J. Bacteriol. 2002, 184, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Finkel, S.E.; Johnson, R.C. The Fis protein: It’s not just for DNA inversion anymore. Mol. Microbiol. 1992, 6, 3257–3265. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, H.; Nilsson, L.; Bosch, L. The mechanism of trans-activation of the Escherichia coli operon thrU(tufB) by the protein FIS. A model. Nucleic Acids Res. 1992, 20, 4077–4081. [Google Scholar] [CrossRef] [PubMed]
- Braun, E.L.; Fuge, E.K.; Padilla, P.A.; Werner-Washburne, M. A stationary-phase gene in Saccharomyces cerevisiae is a member of a novel, highly conserved gene family. J. Bacteriol. 1996, 178, 6865–6872. [Google Scholar] [CrossRef] [PubMed]
- Tramonti, A.; Fiascarelli, A.; Milano, T.; di Salvo, M.L.; Nogués, I.; Pascarella, S.; Contestabile, R. Molecular mechanism of PdxR—A transcriptional activator involved in the regulation of vitamin B6 biosynthesis in the probiotic bacterium Bacillus clausii. FEBS J. 2015, 282, 2966–2984. [Google Scholar] [CrossRef] [PubMed]
- Belitsky, B.R. Role of PdxR in the activation of vitamin B6 biosynthesis in Listeria monocytogenes. Mol. Microbiol. 2014, 92, 1113–1128. [Google Scholar] [CrossRef] [PubMed]
- Milano, T.; Angelaccio, S.; Tramonti, A.; Salvo, D.; Luigi, M.; Contestabile, R.; Pascarella, S. A Bioinformatics Analysis Reveals a Group of MocR Bacterial Transcriptional Regulators Linked to a Family of Genes Coding for Membrane Proteins. Available online: https://www.hindawi.com/journals/bri/2016/4360285/abs/ (accessed on 11 June 2018).
- Oka, T. Modulation of gene expression by vitamin B6. Nutr. Res. Rev. 2001, 14, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Cake, M.H.; DiSorbo, D.M.; Litwack, G. Effect of pyridoxal phosphate on the DNA binding site of activated hepatic glucocorticoid receptor. J. Biol. Chem. 1978, 253, 4886–4891. [Google Scholar] [PubMed]
- Allgood, V.E.; Cidlowski, J.A. Vitamin B6 modulates transcriptional activation by multiple members of the steroid hormone receptor superfamily. J. Biol. Chem. 1992, 267, 3819–3824. [Google Scholar] [PubMed]
- Somvanshi, V.S.; Sloup, R.E.; Crawford, J.M.; Martin, A.R.; Heidt, A.J.; Kim, K.; Clardy, J.; Ciche, T.A. A single promoter inversion switches Photorhabdus between pathogenic and mutualistic states. Science 2012, 337, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Somvanshi, V.S.; Kaufmann-Daszczuk, B.; Kim, K.-S.; Mallon, S.; Ciche, T.A. Photorhabdus phase variants express a novel fimbrial locus, mad, essential for symbiosis. Mol. Microbiol. 2010, 77, 1021–1038. [Google Scholar] [CrossRef] [PubMed]
- Benabdellah, K.; Azcón-Aguilar, C.; Valderas, A.; Speziga, D.; Fitzpatrick, T.B.; Ferrol, N. GintPDX1 encodes a protein involved in vitamin B6 biosynthesis that is up-regulated by oxidative stress in the arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 2009, 184, 682–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antelmann, H.; Bernhardt, J.; Schmid, R.; Mach, H.; Völker, U.; Hecker, M. First steps from a two-dimensional protein index towards a response-regulation map for Bacillus subtilis. Electrophoresis 1997, 18, 1451–1463. [Google Scholar] [CrossRef] [PubMed]
- Brosché, M.; Schuler, M.A.; Kalbina, I.; Connor, L.; Strid, A. Gene regulation by low level UV-B radiation: Identification by DNA array analysis. Photochem. Photobiol. Sci. 2002, 1, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Dawes, I.W.; Roe, J.H. Adaptive response of Schizosaccharomyces pombe to hydrogen peroxide and menadione. Microbiology (Read. Engl.) 1995, 141 Pt 12, 3127–3132. [Google Scholar] [CrossRef] [PubMed]
- Padilla, P.A.; Fuge, E.K.; Crawford, M.E.; Errett, A.; Werner-Washburne, M. The highly conserved, coregulated SNO and SNZ gene families in Saccharomyces cerevisiae respond to nutrient limitation. J. Bacteriol. 1998, 180, 5718–5726. [Google Scholar] [PubMed]
- Roy, S. Function of MYB domain transcription factors in abiotic stress and epigenetic control of stress response in plant genome. Plant Signal. Behav. 2016, 11, e1117723. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.-L.; Ma, N.-N.; Meng, X.; Zhang, S.; Wang, J.-R.; Chai, S.; Meng, Q.-W. A novel tomato MYC-type ICE1-like transcription factor, SlICE1a, confers cold, osmotic and salt tolerance in transgenic tobacco. Plant Physiol. Biochem. 2013, 73, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, M.; Johnson, J.M.; Hieno, A.; Tokizawa, M.; Nomoto, M.; Tada, Y.; Godfrey, R.; Obokata, J.; Sherameti, I.; Yamamoto, Y.Y.; et al. High REDOX RESPONSIVE TRANSCRIPTION FACTOR1 Levels Result in Accumulation of Reactive Oxygen Species in Arabidopsis thaliana Shoots and Roots. Mol. Plant 2015, 8, 1253–1273. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Sharoni, A.M.; Satoh, K.; Kumar, A.; Leung, H.; Kikuchi, S. Comparative transcriptome profiles of the WRKY gene family under control, hormone-treated, and drought conditions in near-isogenic rice lines reveal differential, tissue specific gene activation. J. Plant Physiol. 2014, 171, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.; Hellmann, H.; Schmidt, R.; Willmitzer, L.; Frommer, W.B. Identification of mutants in metabolically regulated gene expression. Plant J. Cell Mol. Biol. 1997, 11, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, J.W. Intracellular trafficking of the pyridoxal cofactor. Implications for health and metabolic disease. Arch. Biochem. Biophys. 2016, 592, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolz, J.; Wöhrmann, H.J.P.; Vogl, C. Amiloride uptake and toxicity in fission yeast are caused by the pyridoxine transporter encoded by bsu1+ (car1+). Eukaryot. Cell 2005, 4, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Stolz, J.; Vielreicher, M. Tpn1p, the plasma membrane vitamin B6 transporter of Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 18990–18996. [Google Scholar] [CrossRef] [PubMed]
- Rossier, B.C.; Staub, O.; Hummler, E. Genetic dissection of sodium and potassium transport along the aldosterone-sensitive distal nephron: Importance in the control of blood pressure and hypertension. FEBS Lett. 2013, 587, 1929–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, L.; Bernhardt, R.; Bernhardt, I. Nongenomic effect of aldosterone on ion transport pathways of red blood cells. Cell. Physiol. Biochem. 2008, 22, 269–278. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L.; Olivieri, O.; Girelli, D.; Lupo, A.; Bernich, P.; Corrocher, R. Red blood cell cation transports in uraemic anaemia: Evidence for an increased K/Cl co-transport activity. Effects of dialysis and erythropoietin treatment. Eur. J. Clin. Investig. 1995, 25, 762–768. [Google Scholar] [CrossRef]
- Said, Z.M.; Subramanian, V.S.; Vaziri, N.D.; Said, H.M. Pyridoxine uptake by colonocytes: A specific and regulated carrier-mediated process. Am. J. Physiol. Cell Physiol. 2008, 294, C1192–C1197. [Google Scholar] [CrossRef] [PubMed]
- Bürkle, L.; Cedzich, A.; Döpke, C.; Stransky, H.; Okumoto, S.; Gillissen, B.; Kühn, C.; Frommer, W.B. Transport of cytokinins mediated by purine transporters of the PUP family expressed in phloem, hydathodes, and pollen of Arabidopsis. Plant J. Cell Mol. Biol. 2003, 34, 13–26. [Google Scholar] [CrossRef]
- Kato, K.; Shitan, N.; Shoji, T.; Hashimoto, T. Tobacco NUP1 transports both tobacco alkaloids and vitamin B6. Phytochemistry 2015, 113, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Szydlowski, N.; Bürkle, L.; Pourcel, L.; Moulin, M.; Stolz, J.; Fitzpatrick, T.B. Recycling of pyridoxine (vitamin B6) by PUP1 in Arabidopsis. Plant J. Cell Mol. Biol. 2013, 75, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, A.; LaBrie, D.A.; Wagner, R.P. Amino acid synthesis by the mitochondria of Neurospora crassa: I. Dependence on respiration of mitochondria. Arch. Biochem. Biophys. 1969, 134, 401–407. [Google Scholar] [CrossRef]
- Whittaker, M.M.; Penmatsa, A.; Whittaker, J.W. The Mtm1p carrier and pyridoxal 5′-phosphate cofactor trafficking in yeast mitochondria. Arch. Biochem. Biophys. 2015, 568, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodionov, D.A.; Hebbeln, P.; Eudes, A.; ter Beek, J.; Rodionova, I.A.; Erkens, G.B.; Slotboom, D.J.; Gelfand, M.S.; Osterman, A.L.; Hanson, A.D.; et al. A novel class of modular transporters for vitamins in prokaryotes. J. Bacteriol. 2009, 191, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Cellini, B.; Montioli, R.; Oppici, E.; Astegno, A.; Borri Voltattorni, C. The chaperone role of the pyridoxal 5′-phosphate and its implications for rare diseases involving B6-dependent enzymes. Clin. Biochem. 2014, 47, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Cellini, B.; Bertoldi, M.; Montioli, R.; Laurents, D.V.; Paiardini, A.; Voltattorni, C.B. Dimerization and Folding Processes of Treponema denticola Cystalysin: The Role of Pyridoxal 5′-Phosphate. Biochemistry (Mosc.) 2006, 45, 14140–14154. [Google Scholar] [CrossRef] [PubMed]
- Deu, E.; Kirsch, J.F. Cofactor-directed reversible denaturation pathways: The cofactor-stabilized Escherichia coli aspartate aminotransferase homodimer unfolds through a pathway that differs from that of the apoenzyme. Biochemistry (Mosc.) 2007, 46, 5819–5829. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, A.N.; Bhakuni, V. Characterization of pyridoxal 5′- phosphate-binding domain and folding intermediate of Bacillus subtilis serine hydroxymethyltransferase: An autonomous folding domain. J. Biochem. (Tokyo) 2008, 144, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Mooney, S.; Hellmann, H. Vitamin B6: Killing two birds with one stone? Phytochemistry 2010, 71, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Leuendorf, J.E.; Osorio, S.; Szewczyk, A.; Fernie, A.R.; Hellmann, H. Complex assembly and metabolic profiling of Arabidopsis thaliana plants overexpressing vitamin B6 biosynthesis proteins. Mol. Plant 2010, 3, 890–903. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kim, S.; Sagong, H.-Y.; Son, H.F.; Jin, K.S.; Kim, I.-K.; Kim, K.-J. Structural basis for cytokinin production by LOG from Corynebacterium glutamicum. Sci. Rep. 2016, 6, 31390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriechbaumer, V.; Botchway, S.W.; Hawes, C. Localization and interactions between Arabidopsis auxin biosynthetic enzymes in the TAA/YUC-dependent pathway. J. Exp. Bot. 2016, 67, 4195–4207. [Google Scholar] [CrossRef] [PubMed]
- Fujino, A.; Ose, T.; Yao, M.; Tokiwano, T.; Honma, M.; Watanabe, N.; Tanaka, I. Structural and enzymatic properties of 1-aminocyclopropane-1-carboxylate deaminase homologue from Pyrococcus horikoshii. J. Mol. Biol. 2004, 341, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; González-Lucán, M.; Donapetry-García, C.; Fernández-Fernández, C.; Ameneiros-Rodríguez, E. Glycogen metabolism in humans. BBA Clin. 2016, 5, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Bahaji, A.; Li, J.; Sánchez-López, Á.M.; Baroja-Fernández, E.; Muñoz, F.J.; Ovecka, M.; Almagro, G.; Montero, M.; Ezquer, I.; Etxeberria, E.; et al. Starch biosynthesis, its regulation and biotechnological approaches to improve crop yields. Biotechnol. Adv. 2014, 32, 87–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kossmann, J.; Lloyd, J. Understanding and influencing starch biochemistry. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 141–196. [Google Scholar] [CrossRef] [PubMed]
- David, E.S.; Crerar, M.M. Quantitation of muscle glycogen phosphorylase mRNA and enzyme amounts in adult rat tissues. Biochim. Biophys. Acta BBA Gen. Subj. 1986, 880, 78–90. [Google Scholar] [CrossRef]
- Shin, Y.S. Glycogen storage disease: Clinical, biochemical, and molecular heterogeneity. Semin. Pediatr. Neurol. 2006, 13, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Oikonomakos, N.G. Glycogen phosphorylase as a molecular target for type 2 diabetes therapy. Curr. Protein Pept. Sci. 2002, 3, 561–586. [Google Scholar] [CrossRef] [PubMed]
- Immenschuh, S.; Vijayan, V.; Janciauskiene, S.; Gueler, F. Heme as a Target for Therapeutic Interventions. Front. Pharmacol. 2017, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Ankar, A.; Bhimji, S.S. Vitamin, B12 (Cobalamin), Deficiency. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Taniguchi, M.; Lindsey, J.S. Synthetic Chlorins, Possible Surrogates for Chlorophylls, Prepared by Derivatization of Porphyrins. Chem. Rev. 2017, 117, 344–535. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Voloshin, R.A.; Korol’kova, D.V.; Tomo, T.; Shen, J.-R. Chlorophylls d and f and Their Role in Primary Photosynthetic Processes of Cyanobacteria. Biochem. Biokhimiia 2016, 81, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Hedtke, B.; Alawady, A.; Albacete, A.; Kobayashi, K.; Melzer, M.; Roitsch, T.; Masuda, T.; Grimm, B. Deficiency in riboflavin biosynthesis affects tetrapyrrole biosynthesis in etiolated Arabidopsis tissue. Plant Mol. Biol. 2012, 78, 77–93. [Google Scholar] [CrossRef] [PubMed]
- Haust, H.L.; Poon, H.C.; Carson, R.; VanDeWetering, C.; Peter, F. Protoporphyrinaemia and decreased activities of 5-aminolevulinic acid dehydrase and uroporphyrinogen I synthetase in erythrocytes of a Vitamin B6-deficient epileptic boy given valproic acid and carbamazepine. Clin. Biochem. 1989, 22, 201–211. [Google Scholar] [CrossRef]
- Davison, K.M.; Kaplan, B.J. Nutrient intakes are correlated with overall psychiatric functioning in adults with mood disorders. Can. J. Psychiatry 2012, 57, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Herbison, C.E.; Hickling, S.; Allen, K.L.; O’Sullivan, T.A.; Robinson, M.; Bremner, A.P.; Huang, R.-C.; Beilin, L.J.; Mori, T.A.; Oddy, W.H. Low intake of B-vitamins is associated with poor adolescent mental health and behaviour. Prev. Med. 2012, 55, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Parletta, N.; Milte, C.M.; Meyer, B.J. Nutritional modulation of cognitive function and mental health. J. Nutr. Biochem. 2013, 24, 725–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaner, G.; Soylu, M.; Yüksel, N.; Inanç, N.; Ongan, D.; Başmısırlı, E. Evaluation of Nutritional Status of Patients with Depression. BioMed Res. Int. 2015, 2015, 521481. [Google Scholar] [CrossRef] [PubMed]
- Rechenberg, K. Nutritional Interventions in Clinical Depression. Clin. Psychol. Sci. 2016, 4, 144–162. [Google Scholar] [CrossRef]
- Goldstein, D.S. Adrenal responses to stress. Cell. Mol. Neurobiol. 2010, 30, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Obeid, A.I.; Johnson, L.; Potts, J.; Mookherjee, S.; Eich, R.H. Fluid therapy in severe systemic reaction to radiopaque dye. Ann. Intern. Med. 1975, 83, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, W.; Obeid, R. Hyperhomocysteinemia and response of methionine cycle intermediates to vitamin treatment in renal patients. Clin. Chem. Lab. Med. 2005, 43, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Purves-Tyson, T.D.; Owens, S.J.; Rothmond, D.A.; Halliday, G.M.; Double, K.L.; Stevens, J.; McCrossin, T.; Shannon Weickert, C. Putative presynaptic dopamine dysregulation in schizophrenia is supported by molecular evidence from post-mortem human midbrain. Transl. Psychiatry 2017, 7, e1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.; Kozielska, M.; Pilla Reddy, V.; Vermeulen, A.; Barton, H.A.; Grimwood, S.; de Greef, R.; Groothuis, G.M.M.; Danhof, M.; Proost, J.H. Translational Modeling in Schizophrenia: Predicting Human Dopamine D2 Receptor Occupancy. Pharm. Res. 2016, 33, 1003–1017. [Google Scholar] [CrossRef] [PubMed]
- Zanatta, G.; Nunes, G.; Bezerra, E.M.; da Costa, R.F.; Martins, A.; Caetano, E.W.S.; Freire, V.N.; Gottfried, C. Antipsychotic haloperidol binding to the human dopamine D3 receptor: Beyond docking through QM/MM refinement toward the design of improved schizophrenia medicines. ACS Chem. Neurosci. 2014, 5, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, K.M.; Raschke, J.; Emborg, M.E. Cell-based therapies for Parkinson’s disease: Past, present, and future. Antioxid. Redox Signal. 2009, 11, 2189–2208. [Google Scholar] [CrossRef] [PubMed]
- Weir, M.R.; Keniston, R.C.; Enriquez, J.I.; McNamee, G.A. Depression of vitamin B6 levels due to dopamine. Vet. Hum. Toxicol. 1991, 33, 118–121. [Google Scholar] [PubMed]
- Rawson, K.S.; Dixon, D.; Nowotny, P.; Ricci, W.M.; Binder, E.F.; Rodebaugh, T.L.; Wendleton, L.; Doré, P.; Lenze, E.J. Association of functional polymorphisms from brain-derived neurotrophic factor and serotonin-related genes with depressive symptoms after a medical stressor in older adults. PLoS ONE 2015, 10, e0120685. [Google Scholar] [CrossRef] [PubMed]
- Bundeff, A.W.; Woodis, C.B. Selective serotonin reuptake inhibitors for the treatment of irritable bowel syndrome. Ann. Pharmacother. 2014, 48, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.; Ridgewell, A.; Ashwin, C. Looking on the bright side: Biased attention and the human serotonin transporter gene. Proc. Biol. Sci. 2009, 276, 1747–1751. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.A. The Public Health Impact of Major Depression: A Call for Interdisciplinary Prevention Efforts. Prev. Sci. 2011, 12, 361–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, C.F.; Ward, M.; Tracey, F.; Hoey, L.; Molloy, A.M.; Pentieva, K.; McNulty, H. B-Vitamin Intake and Biomarker Status in Relation to Cognitive Decline in Healthy Older Adults in a 4-Year Follow-Up Study. Nutrients 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.-H.; Chang, Y.-P.; Yeh, W.-T.; Guei, Y.-S.; Lin, B.-F.; Wei, I.-L.; Yang, F.L.; Liaw, Y.-P.; Chen, K.-J.; Chen, W.J. Co-occurrence of anemia, marginal vitamin B6, and folate status and depressive symptoms in older adults. J. Geriatr. Psychiatry Neurol. 2012, 25, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Abali, E.E.; Skacel, N.E.; Celikkaya, H.; Hsieh, Y.-C. Regulation of human dihydrofolate reductase activity and expression. Vitam. Horm. 2008, 79, 267–292. [Google Scholar] [CrossRef] [PubMed]
- Cortese, C.; Motti, C. MTHFR gene polymorphism, homocysteine and cardiovascular disease. Public Health Nutr. 2001, 4, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Kamen, B. Folate and antifolate pharmacology. Semin. Oncol. 1997, 24 (Suppl. 18), S18-30–S18-39. [Google Scholar]
- Brown, R.R.; Rose, D.P.; Leklem, J.E.; Linkswiler, H.M. Effects of oral contraceptives on tryptophan metabolism and vitamin B6 requirements in women. Acta Vitaminol. Enzymol. 1975, 29, 151–157. [Google Scholar] [PubMed]
- Fryar-Williams, S. Fundamental Role of Methylenetetrahydrofolate Reductase 677 C→ T Genotype and Flavin Compounds in Biochemical Phenotypes for Schizophrenia and Schizoaffective Psychosis. Front. Psychiatry 2016, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, R.A.; Nicolia, V.; Fiorenza, M.T.; Scarpa, S.; Fuso, A. S-Adenosylmethionine and Superoxide Dismutase 1 Synergistically Counteract Alzheimer’s Disease Features Progression in TgCRND8 Mice. Antioxid. Basel Switz. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.J.; Mansoor, M.A.; Pentieva, K.D.; Hamer, M.; Mishra, G.D. Biochemical risk indices, including plasma homocysteine, that prospectively predict mortality in older British people: The National Diet and Nutrition Survey of People Aged 65 Years and Over. Br. J. Nutr. 2010, 104, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Chiang, P.K.; Gordon, R.K.; Tal, J.; Zeng, G.C.; Doctor, B.P.; Pardhasaradhi, K.; McCann, P.P. S-Adenosylmethionine and methylation. FASEB J. 1996, 10, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Brocardo, P.S.; Budni, J.; Kaster, M.P.; Santos, A.R.S.; Rodrigues, A.L.S. Folic acid administration produces an antidepressant-like effect in mice: Evidence for the involvement of the serotonergic and noradrenergic systems. Neuropharmacology 2008, 54, 464–473. [Google Scholar] [CrossRef] [PubMed]
- di Salvo, M.L.; Safo, M.K.; Contestabile, R. Biomedical aspects of pyridoxal 5′-phosphate availability. Front. Biosci. Elite Ed. 2012, 4, 897–913. [Google Scholar] [PubMed]
- Morris, M.S.; Picciano, M.F.; Jacques, P.F.; Selhub, J. Plasma pyridoxal 5′-phosphate in the US population: The National Health and Nutrition Examination Survey, 2003–2004. Am. J. Clin. Nutr. 2008, 87, 1446–1454. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.J.; Pentieva, K.D.; Prentice, A.; Mansoor, M.A.; Finch, S. Plasma pyridoxal phosphate and pyridoxic acid and their relationship to plasma homocysteine in a representative sample of British men and women aged 65 years and over. Br. J. Nutr. 1999, 81, 191–201. [Google Scholar] [PubMed]
- Institute of Medicine (US) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes and Its Panel on Folate, Other B Vitamins, and Choline. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline; The National Academies Collection: Reports Funded by National Institutes of Health; National Academies Press (US): Washington, DC, USA, 1998; ISBN 978-0-309-06411-8. [Google Scholar]
- Brown, M.J.; Beier, K. Vitamin, B6 (Pyridoxine), Deficiency. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Glória, L.; Cravo, M.; Camilo, M.E.; Resende, M.; Cardoso, J.N.; Oliveira, A.G.; Leitão, C.N.; Mira, F.C. Nutritional deficiencies in chronic alcoholics: Relation to dietary intake and alcohol consumption. Am. J. Gastroenterol. 1997, 92, 485–489. [Google Scholar] [PubMed]
- Lin, G.W. Effect of ethanol and vitamin B6 deficiency on pyridoxal 5-phosphate levels and fetal growth in rat. Alcohol. Clin. Exp. Res. 1989, 13, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.; Göbert, A.; Franke, S.; Ott, U.; Gerth, J.; Müller, A.; Stein, G.; Bitsch, R.; Wolf, G. Vitamin B6 metabolism in chronic kidney disease--relation to transsulfuration, advanced glycation and cardiovascular disease. Nephron Clin. Pract. 2010, 114, c38–c46. [Google Scholar] [CrossRef] [PubMed]
- Reid, E.S.; Williams, H.; Stabej, P.L.Q.; James, C.; Ocaka, L.; Bacchelli, C.; Footitt, E.J.; Boyd, S.; Cleary, M.A.; Mills, P.B.; et al. Seizures Due to a KCNQ2 Mutation: Treatment with Vitamin B6. JIMD Rep. 2016, 27, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-G.; Lee, Y.; Shin, H.; Kang, K.; Park, J.-M.; Kim, B.-K.; Kwon, O.; Lee, J.-J. Seizures Related to Vitamin B6 Deficiency in Adults. J. Epilepsy Res. 2015, 5, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Mintzer, S.; Skidmore, C.T.; Sperling, M.R. B-vitamin deficiency in patients treated with antiepileptic drugs. Epilepsy Behav. 2012, 24, 341–344. [Google Scholar] [CrossRef] [PubMed]
- McCormick, D.B. Present Knowledge in Nutrition Volume I; Bowman, B.A., Russel, R., Eds.; International Life Sciences Institue: Washington, DC, USA, 2006. [Google Scholar]
- Apeland, T.; Mansoor, M.A.; Pentieva, K.; McNulty, H.; Strandjord, R.E. Fasting and Post-Methionine Loading Concentrations of Homocysteine, Vitamin B2, and Vitamin B6 in Patients on Antiepileptic Drugs. Clin. Chem. 2003, 49, 1005–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagianos, K.; Bernstein, C.N. Homocysteinemia and B vitamin status among adult patients with inflammatory bowel disease: A one-year prospective follow-up study. Inflamm. Bowel Dis. 2012, 18, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Hess, O.M. Homocysteine and B vitamins. Handb. Exp. Pharmacol. 2005, 325–338. [Google Scholar]
- Merrill, A.H.; Henderson, J.M. Diseases Associated with Defects in Vitamin B6 Metabolism or Utilization. Annu. Rev. Nutr. 1987, 7, 137–156. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, E.; Striano, S.; Andria, G. Electroencephalographic abnormalities in homocystinuria due to cystathionine synthase deficiency. Clin. Neurol. Neurosurg. 1983, 85, 165–168. [Google Scholar] [CrossRef]
- Toriyama, T.; Matsuo, S.; Fukatsu, A.; Takahashi, H.; Sato, K.; Mimuro, N.; Kawahara, H. Effects of high-dose vitamin B6 therapy on microcytic and hypochromic anemia in hemodialysis patients. Nihon Jinzo Gakkai Shi 1993, 35, 975–980. [Google Scholar] [PubMed]
- Merete, C.; Falcon, L.M.; Tucker, K.L. Vitamin B6 is associated with depressive symptomatology in Massachusetts elders. J. Am. Coll. Nutr. 2008, 27, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Malouf, R.; Grimley Evans, J. The effect of vitamin B6 on cognition. Cochrane Database Syst. Rev. 2003, CD004393. [Google Scholar] [CrossRef]
- Qian, B.; Shen, S.; Zhang, J.; Jing, P. Effects of Vitamin B6 Deficiency on the Composition and Functional Potential of T Cell Populations. J. Immunol. Res. 2017, 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ueland, P.M.; McCann, A.; Midttun, Ø.; Ulvik, A. Inflammation, vitamin B6 and related pathways. Mol. Aspects Med. 2017, 53, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, R.; Girija, A.S. Pyridoxine-dependent seizures: A review. Indian Pediatr. 2003, 40, 633–638. [Google Scholar] [PubMed]
- Schaumburg, H.; Kaplan, J.; Windebank, A.; Vick, N.; Rasmus, S.; Pleasure, D.; Brown, M.J. Sensory neuropathy from pyridoxine abuse. A new megavitamin syndrome. N. Engl. J. Med. 1983, 309, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Parry, G.J.; Bredesen, D.E. Sensory neuropathy with low-dose pyridoxine. Neurology 1985, 35, 1466–1468. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.; Zeris, S.; Kothari, M.J. Elevated B6 levels and peripheral neuropathies. Electromyogr. Clin. Neurophysiol. 2008, 48, 219–223. [Google Scholar] [PubMed]
- Baer, R.L. Cutaneous skin changes probably due to pyridoxine abuse. J. Am. Acad. Dermatol. 1984, 10, 527–528. [Google Scholar] [CrossRef]
- Friedman, M.A.; Resnick, J.S.; Baer, R.L. Subepidermal vesicular dermatosis and sensory peripheral neuropathy caused by pyridoxine abuse. J. Am. Acad. Dermatol. 1986, 14, 915–917. [Google Scholar] [CrossRef]
- De Zegher, F.; Przyrembel, H.; Chalmers, R.A.; Wolff, E.D.; Huijmans, J.G.M. Successful treatment of infantile type I primary hyperoxaluria complicated by pyridoxine toxicity. Lancet 1985, 326, 392–393. [Google Scholar] [CrossRef]
- Clayton, P.T. B6-responsive disorders: A model of vitamin dependency. J. Inherit. Metab. Dis. 2006, 29, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Amadasi, A.; Bertoldi, M.; Contestabile, R.; Bettati, S.; Cellini, B.; di Salvo, M.L.; Borri-Voltattorni, C.; Bossa, F.; Mozzarelli, A. Pyridoxal 5′-phosphate enzymes as targets for therapeutic agents. Curr. Med. Chem. 2007, 14, 1291–1324. [Google Scholar] [CrossRef] [PubMed]
- Kronenberger, T.; Lindner, J.; Meissner, K.A.; Zimbres, F.M.; Coronado, M.A.; Sauer, F.M.; Schettert, I.; Wrenger, C. Vitamin B6-dependent enzymes in the human malaria parasite Plasmodium falciparum: A druggable target? BioMed Res. Int. 2014, 2014, 108516. [Google Scholar] [CrossRef] [PubMed]
- Schnell, R.; Sriram, D.; Schneider, G. Pyridoxal-phosphate dependent mycobacterial cysteine synthases: Structure, mechanism and potential as drug targets. Biochim. Biophys. Acta 2015, 1854, 1175–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, P. Pyridoxine-dependent and pyridoxine-responsive seizures. Dev. Med. Child Neurol. 2001, 43, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Stockler, S.; Plecko, B.; Gospe, S.M.; Coulter-Mackie, M.; Connolly, M.; van Karnebeek, C.; Mercimek-Mahmutoglu, S.; Hartmann, H.; Scharer, G.; Struijs, E.; et al. Pyridoxine dependent epilepsy and antiquitin deficiency: Clinical and molecular characteristics and recommendations for diagnosis, treatment and follow-up. Mol. Genet. Metab. 2011, 104, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.B.; Struys, E.; Jakobs, C.; Plecko, B.; Baxter, P.; Baumgartner, M.; Willemsen, M.A.A.P.; Omran, H.; Tacke, U.; Uhlenberg, B.; et al. Mutations in antiquitin in individuals with pyridoxine-dependent seizures. Nat. Med. 2006, 12, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Pena, I.A.; Roussel, Y.; Daniel, K.; Mongeon, K.; Johnstone, D.; Mendes, H.W.; Bosma, M.; Saxena, V.; Lepage, N.; Chakraborty, P.; et al. Pyridoxine-Dependent Epilepsy in Zebrafish Caused by Aldh7a1 Deficiency. Genetics 2017, 207, 1501–1518. [Google Scholar] [CrossRef] [PubMed]
- Elstner, M.; Morris, C.M.; Heim, K.; Lichtner, P.; Bender, A.; Mehta, D.; Schulte, C.; Sharma, M.; Hudson, G.; Goldwurm, S.; et al. Single-cell expression profiling of dopaminergic neurons combined with association analysis identifies pyridoxal kinase as Parkinson’s disease gene. Ann. Neurol. 2009, 66, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Guella, I.; Asselta, R.; Tesei, S.; Zini, M.; Pezzoli, G.; Duga, S. The PDXK rs2010795 variant is not associated with Parkinson disease in Italy. Ann. Neurol. 2010, 67, 411–412. [Google Scholar] [CrossRef] [PubMed]
- Vilariño-Güell, C.; Wider, C.; Aasly, J.O.; White, L.R.; Rajput, A.; Rajput, A.H.; Lynch, T.; Krygowska-Wajs, A.; Jasinska-Myga, B.; Opala, G.; et al. Association of pyridoxal kinase and Parkinson disease. Ann. Neurol. 2010, 67, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Ehrenshaft, M.; Chung, K.R.; Jenns, A.E.; Daub, M.E. Functional characterization of SOR1, a gene required for resistance to photosensitizing toxins in the fungus Cercospora nicotianae. Curr. Genet. 1999, 34, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Ehrenshaft, M.; Jenns, A.E.; Chung, K.R.; Daub, M.E. SOR1, a gene required for photosensitizer and singlet oxygen resistance in Cercospora fungi, is highly conserved in divergent organisms. Mol. Cell 1998, 1, 603–609. [Google Scholar] [CrossRef]
- Daub, M.E.; Ehrenshaft, M. The photoactivated Cercospora toxin Cercosporin: Contributions to Plant Disease and Fundamental Biology. Annu. Rev. Phytopathol. 2000, 38, 461–490. [Google Scholar] [CrossRef] [PubMed]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Bilski, P.; Li, M.Y.; Chignell, C.F.; Daub, M.E. A highly conserved sequence is a novel gene involved in de novo vitamin B6 biosynthesis. Proc. Natl. Acad. Sci. USA 1999, 96, 9374–9378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jin, X.; Ouyang, Z.; Li, X.; Liu, B.; Huang, L.; Hong, Y.; Zhang, H.; Song, F.; Li, D. Vitamin B6 contributes to disease resistance against Pseudomonas syringae pv. tomato DC3000 and Botrytis cinerea in Arabidopsis thaliana. J. Plant. Physiol. 2015, 175, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yoshiga, T.; Hasegawa, K. Activated and inactivated immune responses in Caenorhabditis elegans against Photorhabdus luminescens TT01. SpringerPlus 2014, 3, 274. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yoshiga, T.; Hasegawa, K. Involvement of Vitamin B6 Biosynthesis Pathways in the Insecticidal Activity of Photorhabdus luminescens. Appl. Environ. Microbiol. 2016, 82, 3546–3553. [Google Scholar] [CrossRef] [PubMed]
- Grubman, A.; Phillips, A.; Thibonnier, M.; Kaparakis-Liaskos, M.; Johnson, C.; Thiberge, J.-M.; Radcliff, F.J.; Ecobichon, C.; Labigne, A.; de Reuse, H.; et al. Vitamin B6 is required for full motility and virulence in Helicobacter pylori. mBio 2010, 1, e00112-10. [Google Scholar] [CrossRef] [PubMed]
- Dick, T.; Manjunatha, U.; Kappes, B.; Gengenbacher, M. Vitamin B6 biosynthesis is essential for survival and virulence of Mycobacterium tuberculosis. Mol. Microbiol. 2010, 78, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Cui, Y.; Li, Z.; Chen, L.; Liu, S. New Targets and Cofactors for the Transcription Factor LrpA from Mycobacterium tuberculosis. DNA Cell Biol. 2016, 35, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Gengenbacher, M.; Vogelzang, A.; Schuerer, S.; Lazar, D.; Kaiser, P.; Kaufmann, S.H.E. Dietary Pyridoxine Controls Efficacy of Vitamin B6 -Auxotrophic Tuberculosis Vaccine Bacillus Calmette-Guérin ΔureC::hly Δpdx1 in Mice. mBio 2014, 5, e01262-14. [Google Scholar] [CrossRef] [PubMed]
- WHO. World Malaria Report 2015. Available online: http://www.who.int/malaria/publications/world-malaria-report-2015/report/en/ (accessed on 27 December 2017).
- Mayer, J.E. Delivering golden rice to developing countries. J. AOAC Int. 2007, 90, 1445–1449. [Google Scholar] [PubMed]
Figure 1.
Chemical structures of the six common B6 vitamers. (a,b): Pyridoxal (a) and its phosphorylated form pyridoxal 5′-phosphate (b), (c,d): pyridoxamine (c) and pyridoxamine 5′-phosphate (d), (e,f): pyridoxine (e) and pyridoxine 5′-phosphate (f).
Figure 1.
Chemical structures of the six common B6 vitamers. (a,b): Pyridoxal (a) and its phosphorylated form pyridoxal 5′-phosphate (b), (c,d): pyridoxamine (c) and pyridoxamine 5′-phosphate (d), (e,f): pyridoxine (e) and pyridoxine 5′-phosphate (f).
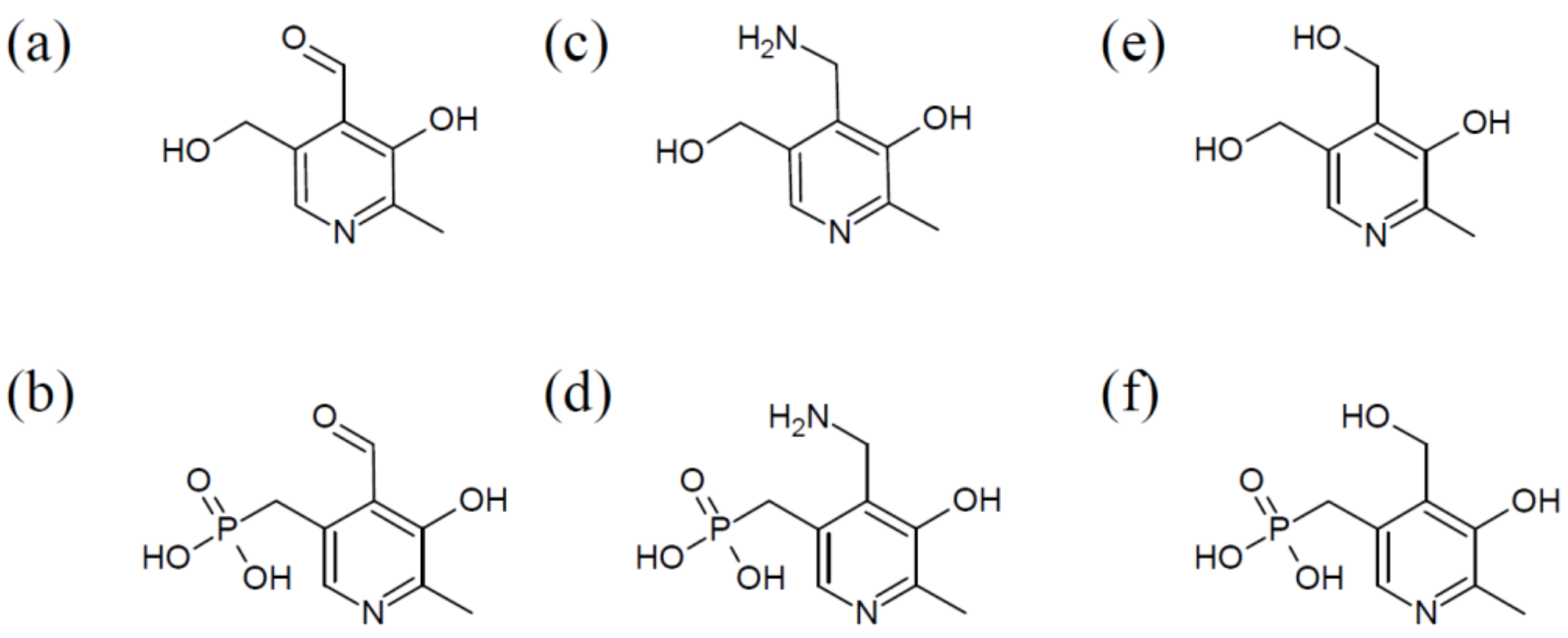
Figure 2.
The two known de novo pathways leading to the biosynthesis of PLP. The two de novo pathways, DXP-dependent and -independent, use pyridoxine 5′-phosphate synthase (PDXJ) and 4-hydroxythreonine-4-phosphate dehydrogenase (A) and PDX1 and 2 enzyme, respectively, to synthesize PLP. DXP, Deoxyxylose 5′-phosphate; DAP, Dihydroxyacetone phosphate; DXP, 1-Deoxyxylulose-5-phosphate; GAP, Glyceraldehyde-3-phosphate; GLN, L-Glutamine; GLU, L-Glutamate; PHT, 4-Phosphohydroxy-L-threonine; PLP, Pyridoxal-5-phosphate; PNP, Pyridoxine-5-phosphate; RIP, Ribose-5-phosphate; RUP, Ribulose-5-phosphate.
Figure 2.
The two known de novo pathways leading to the biosynthesis of PLP. The two de novo pathways, DXP-dependent and -independent, use pyridoxine 5′-phosphate synthase (PDXJ) and 4-hydroxythreonine-4-phosphate dehydrogenase (A) and PDX1 and 2 enzyme, respectively, to synthesize PLP. DXP, Deoxyxylose 5′-phosphate; DAP, Dihydroxyacetone phosphate; DXP, 1-Deoxyxylulose-5-phosphate; GAP, Glyceraldehyde-3-phosphate; GLN, L-Glutamine; GLU, L-Glutamate; PHT, 4-Phosphohydroxy-L-threonine; PLP, Pyridoxal-5-phosphate; PNP, Pyridoxine-5-phosphate; RIP, Ribose-5-phosphate; RUP, Ribulose-5-phosphate.
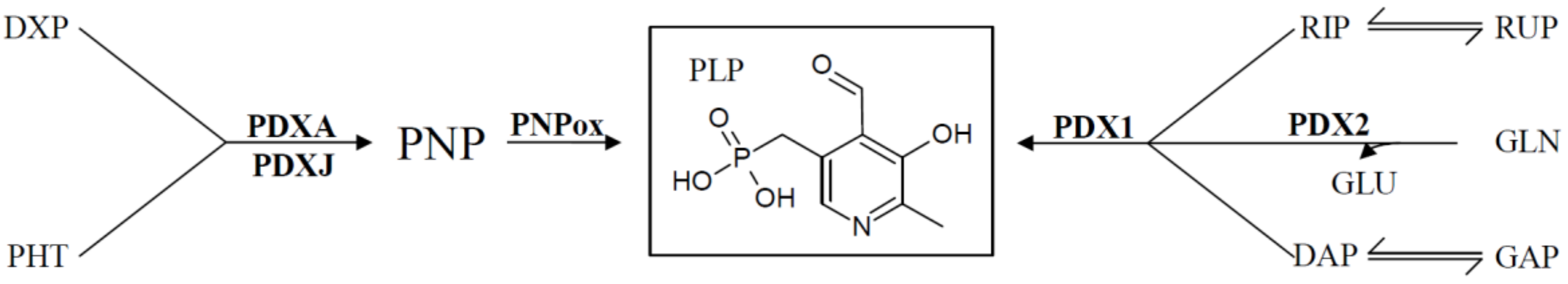
Figure 3.
The VitB6 salvage pathway. The salvage pathway is used to derivatize and phosphorylate B6 vitamers, leading to PLP synthesis via oxidase (PDXH), kinase (K), and pyridoxal reductase (PRL) enzymes.
Figure 3.
The VitB6 salvage pathway. The salvage pathway is used to derivatize and phosphorylate B6 vitamers, leading to PLP synthesis via oxidase (PDXH), kinase (K), and pyridoxal reductase (PRL) enzymes.
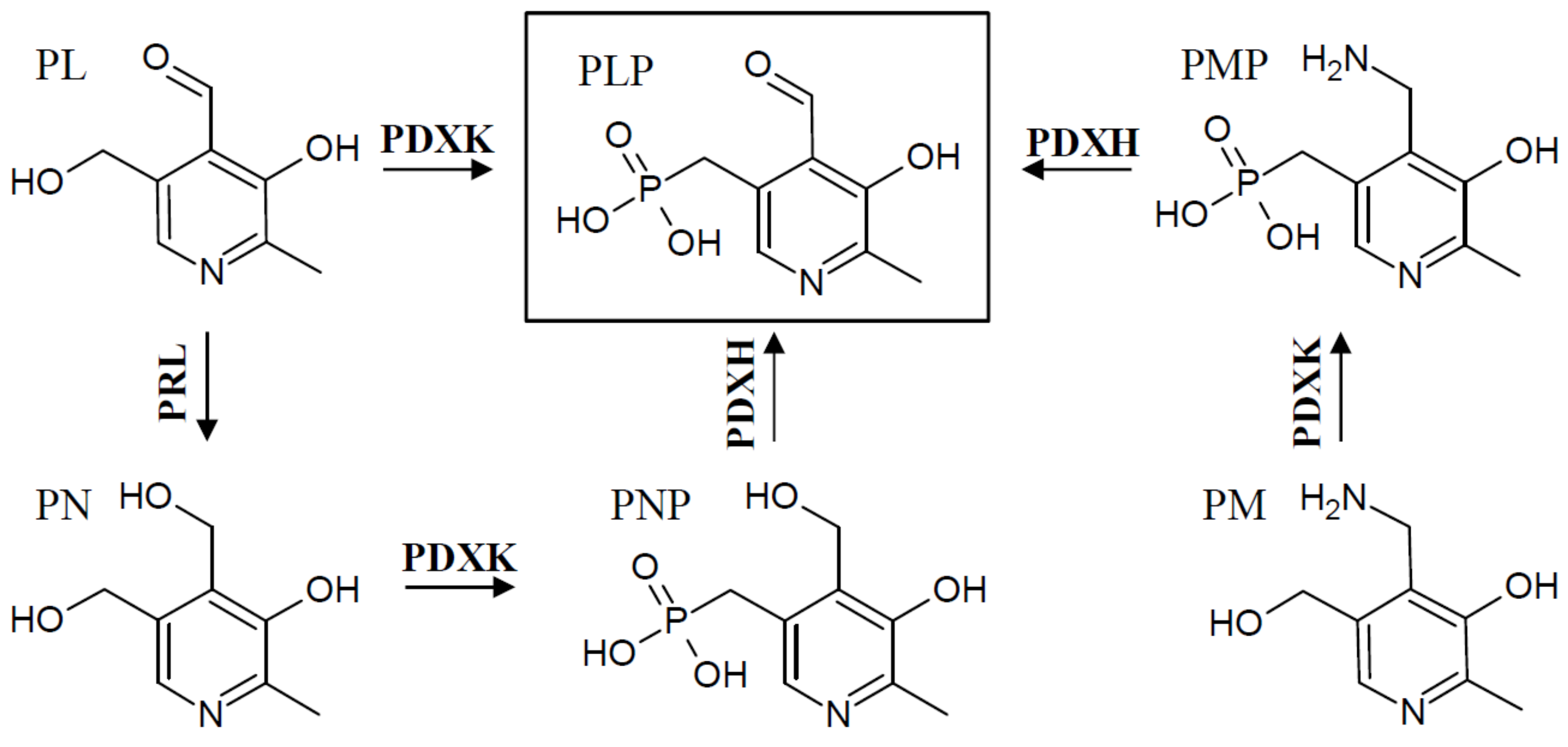
Figure 4.
Schematic of known and potential vitB6 transporters. In the yeasts S. cerevisiae and S. pombe, import of vitB6 across the plasma membrane is facilitated by Tpn1p and Bsu1, respectively. In plants, members of the amino acid permease family (here PUP1) perform this function. In humans, there is no confirmed transporter for vitB6, but this may be accomplished by BsuI orthologs. In the bacterial phylum firmicutes, a modular transporter called PdxT transports B6 vitamers into the cell. Mtm1p is a carrier from S. cerevisiae that imports PLP into the mitochondria. De novo and salvage pathways are labeled according to their presumed subcellular localization and presence in the respective organism.
Figure 4.
Schematic of known and potential vitB6 transporters. In the yeasts S. cerevisiae and S. pombe, import of vitB6 across the plasma membrane is facilitated by Tpn1p and Bsu1, respectively. In plants, members of the amino acid permease family (here PUP1) perform this function. In humans, there is no confirmed transporter for vitB6, but this may be accomplished by BsuI orthologs. In the bacterial phylum firmicutes, a modular transporter called PdxT transports B6 vitamers into the cell. Mtm1p is a carrier from S. cerevisiae that imports PLP into the mitochondria. De novo and salvage pathways are labeled according to their presumed subcellular localization and presence in the respective organism.
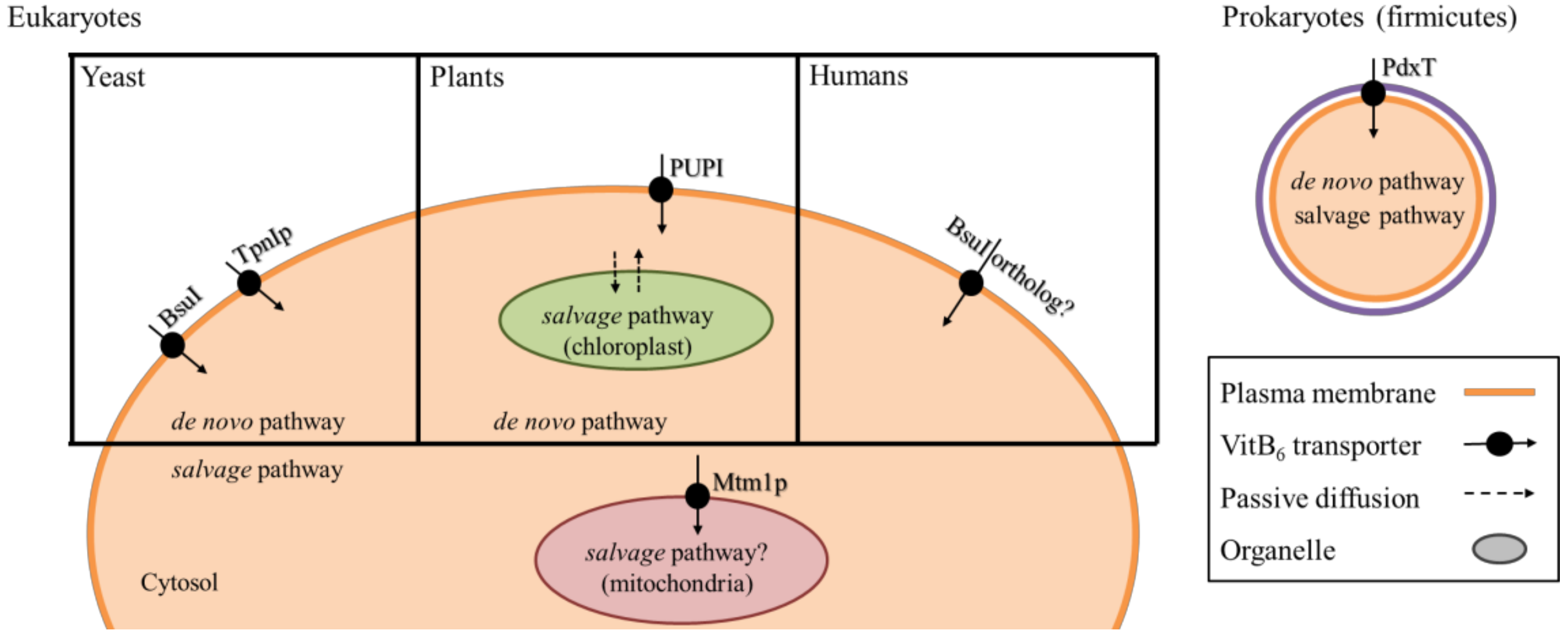
Figure 5.
Schematic drawing of neurotransmitter biosynthesis that involve PLP-dependent enzymes. B6, vitB6; L-DOPA, L-3′-4′-dihydroxyphenylalanine; 5′-HTP, 5′-hydroxytryptophan; 5′-HT, 5′-hydroxytryptamine.
Figure 5.
Schematic drawing of neurotransmitter biosynthesis that involve PLP-dependent enzymes. B6, vitB6; L-DOPA, L-3′-4′-dihydroxyphenylalanine; 5′-HTP, 5′-hydroxytryptophan; 5′-HT, 5′-hydroxytryptamine.
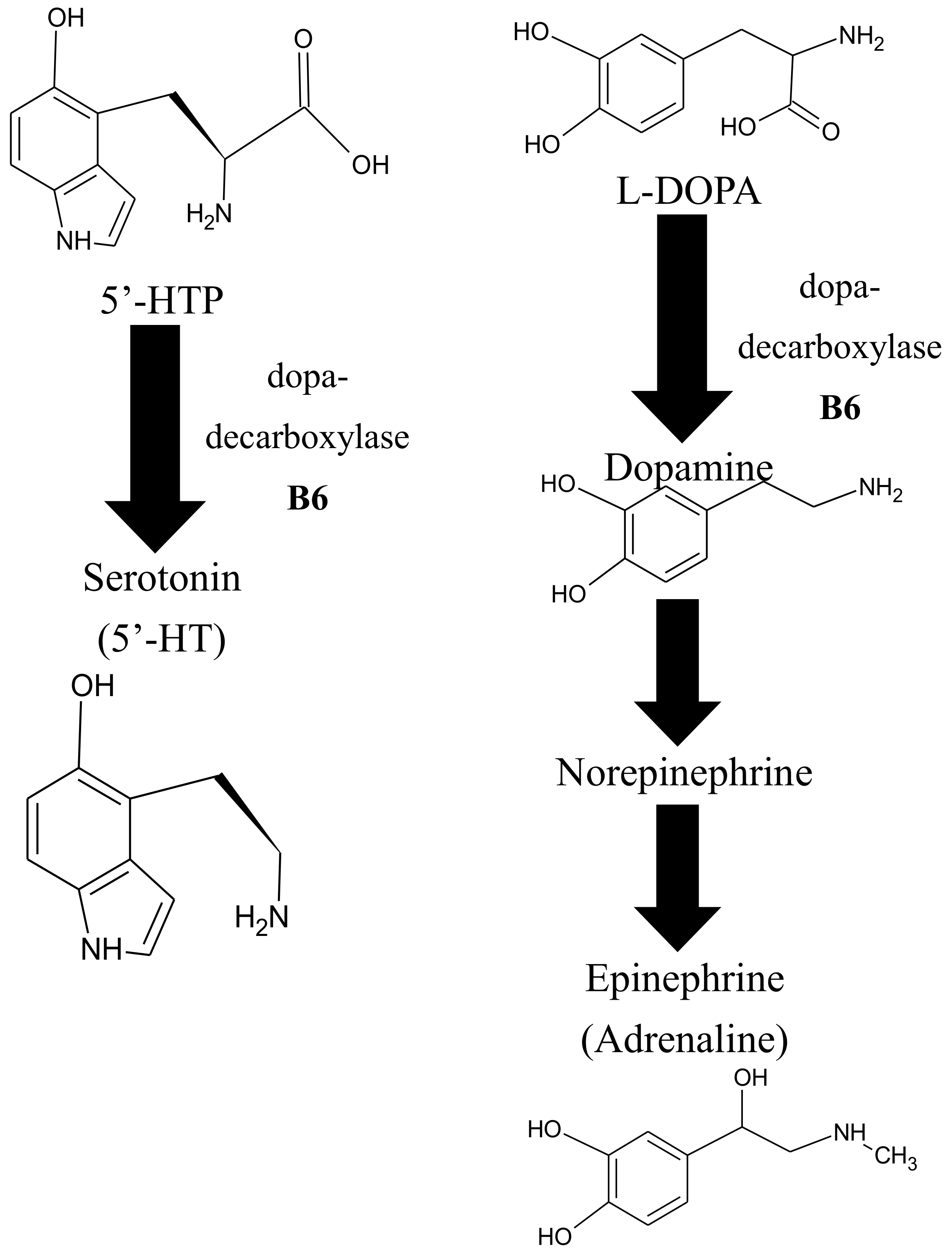
Figure 6.
Schematic drawing of the 1-carbon methylation cycle. The methionine and the folate cycle, as well as the transulfuration pathway, are three interrelated routes involved in one-carbon metabolism, which is required for the synthesis of amino acids, neurotransmitters, and methylation of DNA and proteins. B6, vitB6; B12, vitB12; BH4, tetrahydrobiopterin; CBS, cystathionine-β-synthase; MAT, methionine adenosyltransferase; MS, methionine synthase; MT, methyltransferase; MTHFR, methylenetetrahydrofolate reductase; SAH, S-adenosylhomocysteine; SAHH, S-adenosylhomocysteine hydrolase; SAM, S-adenosylmethionine; SHMT, serine hydroxymethyltransferase; THF, tetrahydrofolate.
Figure 6.
Schematic drawing of the 1-carbon methylation cycle. The methionine and the folate cycle, as well as the transulfuration pathway, are three interrelated routes involved in one-carbon metabolism, which is required for the synthesis of amino acids, neurotransmitters, and methylation of DNA and proteins. B6, vitB6; B12, vitB12; BH4, tetrahydrobiopterin; CBS, cystathionine-β-synthase; MAT, methionine adenosyltransferase; MS, methionine synthase; MT, methyltransferase; MTHFR, methylenetetrahydrofolate reductase; SAH, S-adenosylhomocysteine; SAHH, S-adenosylhomocysteine hydrolase; SAM, S-adenosylmethionine; SHMT, serine hydroxymethyltransferase; THF, tetrahydrofolate.
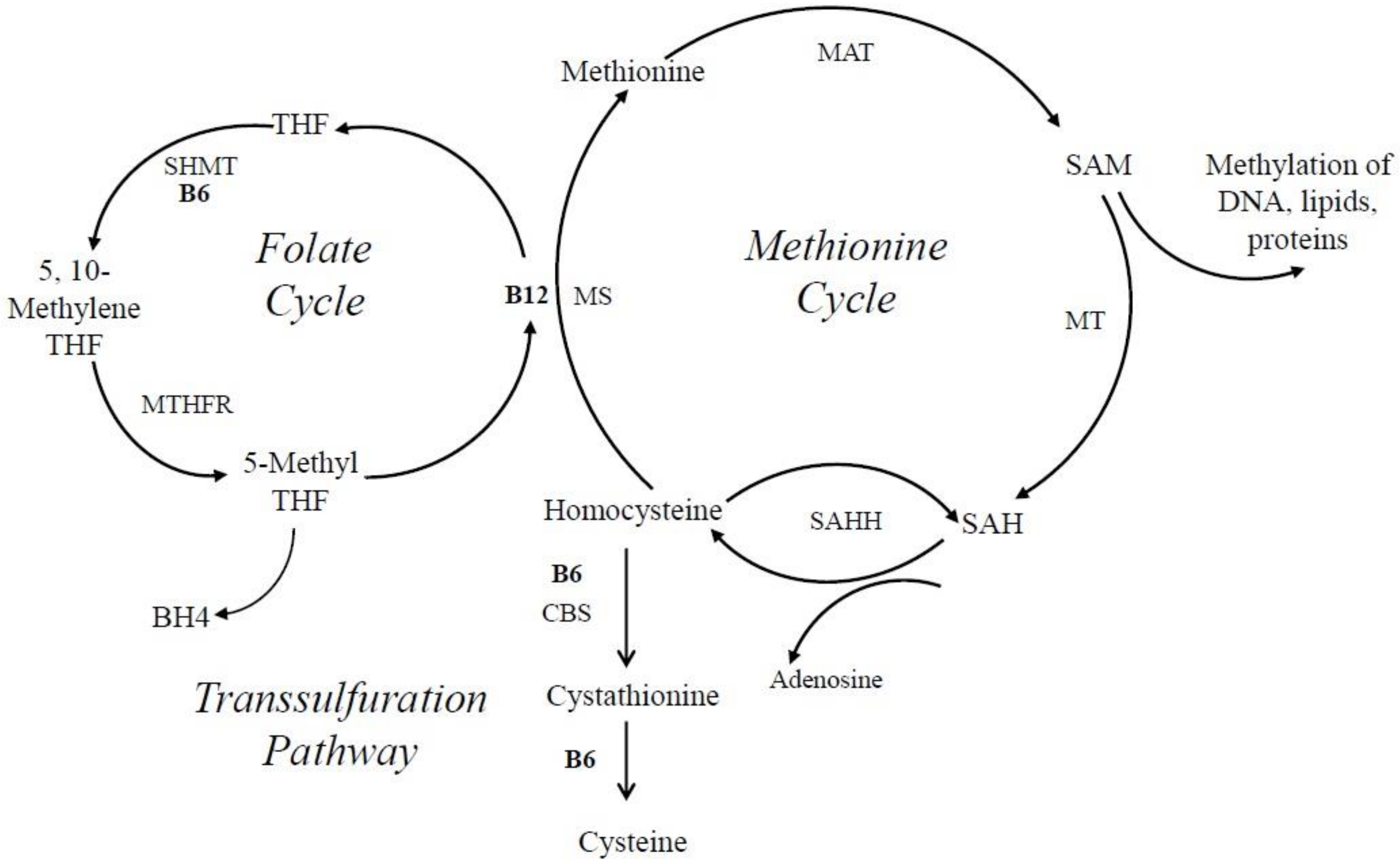

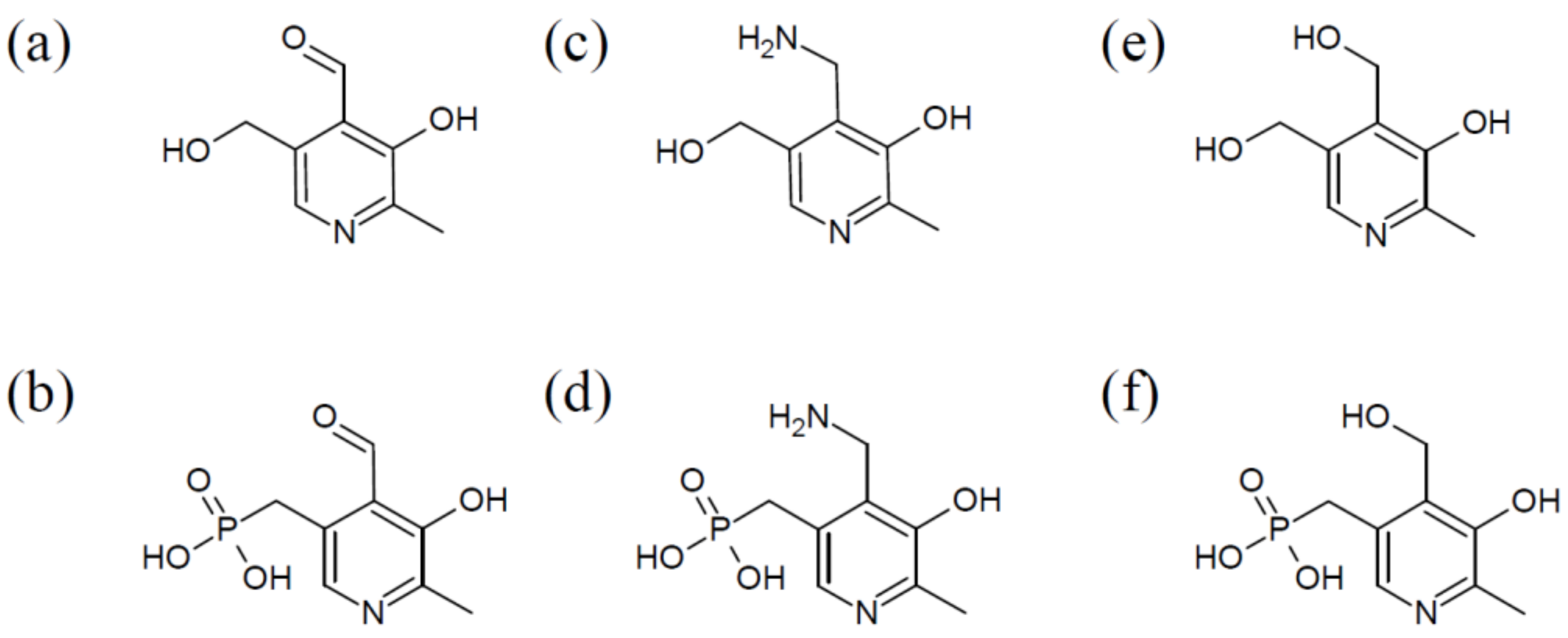{kind=link}
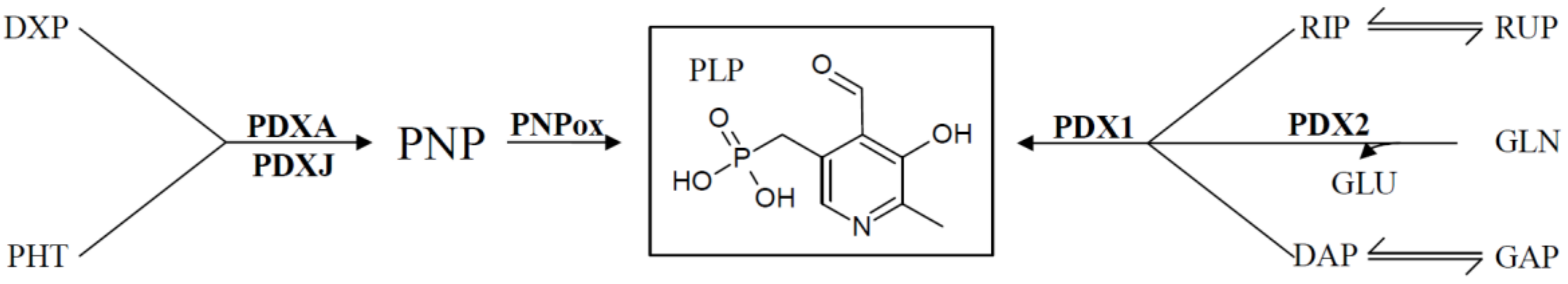{kind=link}
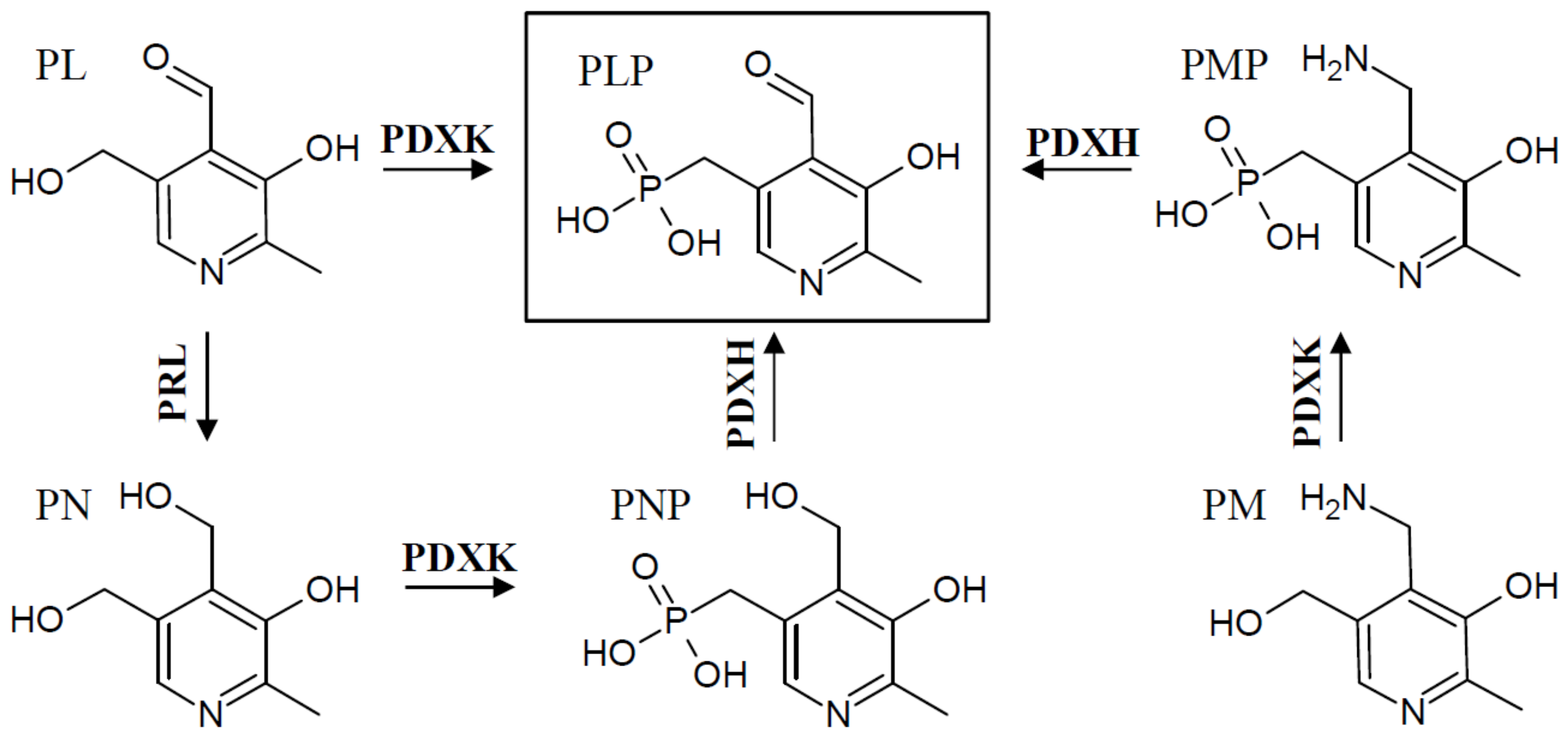{kind=link}
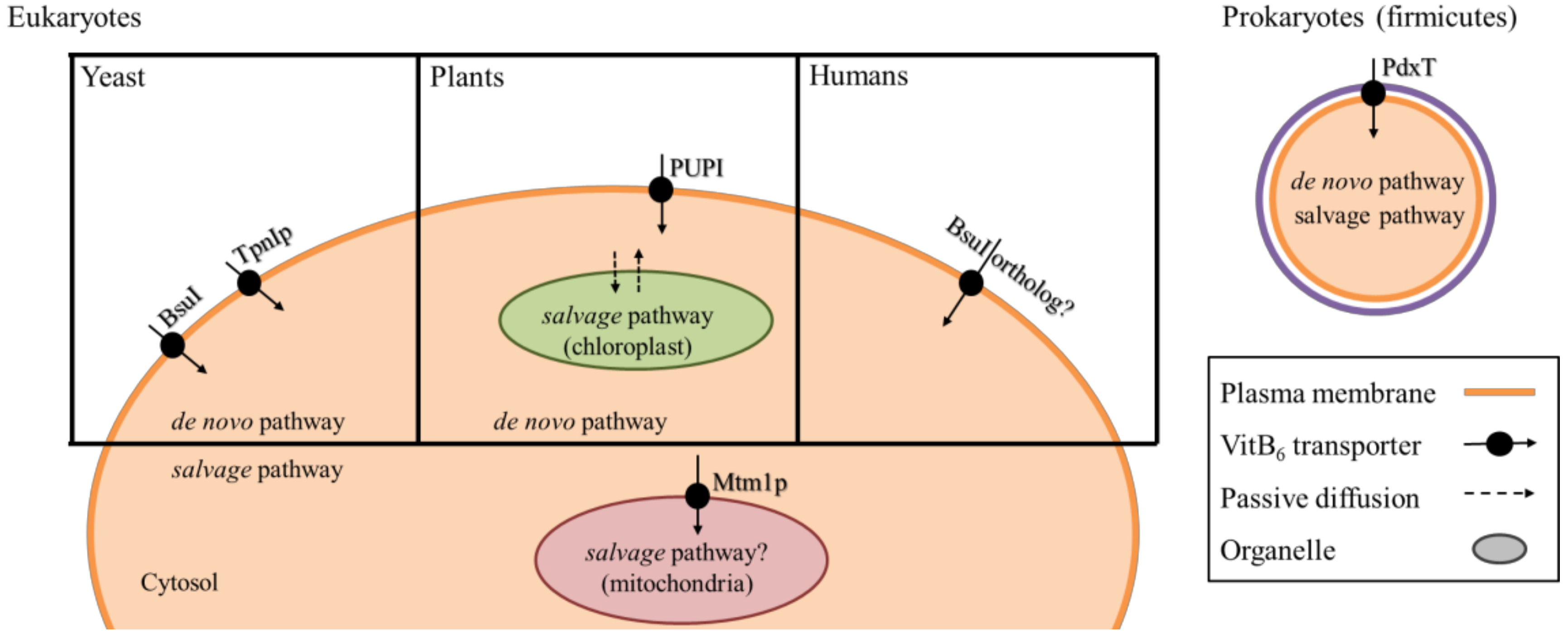{kind=link}
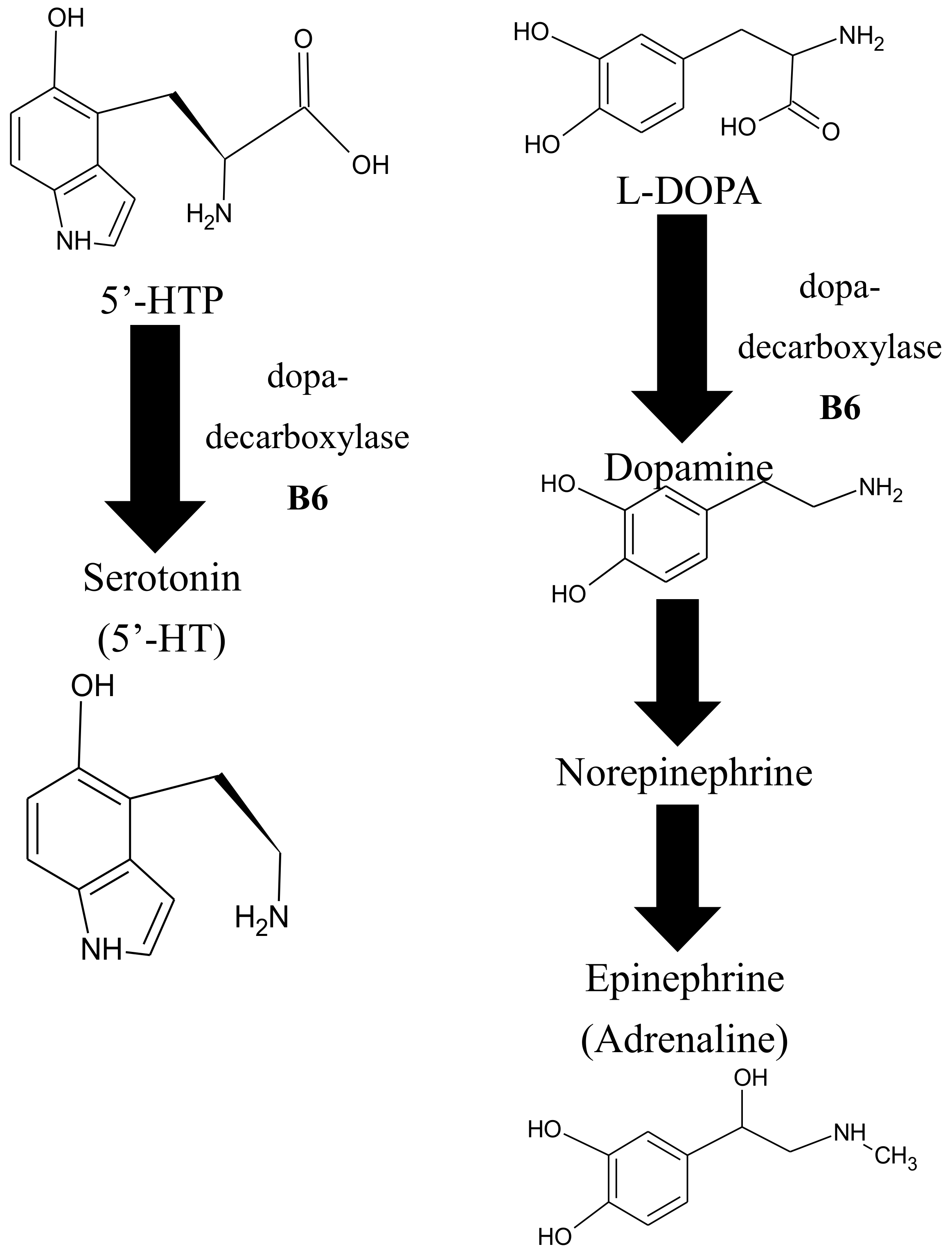{kind=link}
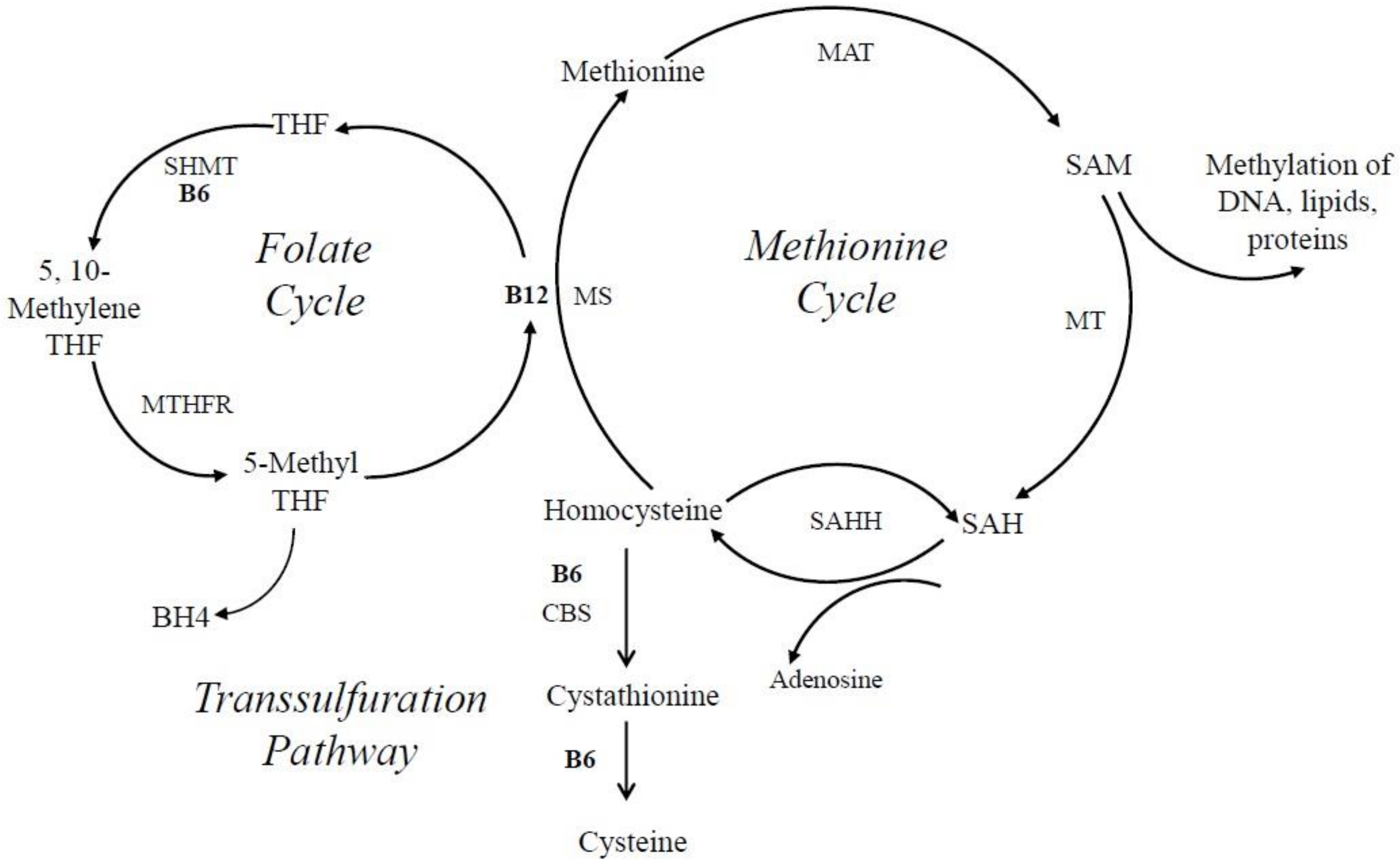{kind=link}
Table 1.
Selected Foods and VitB6 Content based on the USDA Food Composition Database (https://ndb.nal.usda.gov).
Table 1.
Selected Foods and VitB6 Content based on the USDA Food Composition Database (https://ndb.nal.usda.gov).
| Food Source | VitB6 [mg/100 g] |
|---|---|
| Crude Rice Bran | 4.07 |
| Vegetable Oil Spread (60% Fat) | 3.75 |
| Raw Garlic | 1.235 |
| Cooked Chicken Breast | 1.157 |
| Cooked Beef Liver | 1.083 |
| Roasted Pistachio Nuts | 1.07 |
| Cooked Yellow Fin Tuna | 1.04 |
| Top Round Boneless Steak | 0.891 |
| Cooked Sockeye Salmon | 0.83 |
| Potato Chips | 0.8 |
| Roasted Hazelnuts | 0.62 |
| Baked Potato | 0.614 |
| American Cheese | 0.567 |
| Flaxseeds | 0.47 |
| Feta Cheese | 0.424 |
| Raw Bananas | 0.367 |
| Raw Avocado | 0.257 |
| Hard-boiled Egg | 0.121 |
| American Cheddar | 0.12 |
| Dried Pine Nuts | 0.09 |
| Milk (2% Fat) | 0.051 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Parra, M.; Stahl, S.; Hellmann, H. Vitamin B6 and Its Role in Cell Metabolism and Physiology. Cells 2018, 7, 84. https://doi.org/10.3390/cells7070084
AMA Style
Parra M, Stahl S, Hellmann H. Vitamin B6 and Its Role in Cell Metabolism and Physiology. Cells. 2018; 7(7):84. https://doi.org/10.3390/cells7070084
Chicago/Turabian StyleParra, Marcelina, Seth Stahl, and Hanjo Hellmann. 2018. "Vitamin B6 and Its Role in Cell Metabolism and Physiology" Cells 7, no. 7: 84. https://doi.org/10.3390/cells7070084
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.