Functions of Rab Proteins at Presynaptic Sites


1
Department of Neurobiology, Max-Planck-Institute for Biophysical Chemistry, Göttingen 37077, Germany
2
Interactomics and Intracellular Trafficking laboratory, Department of Physiology, Yong Loo Lin School of Medicine, National University of Singapore, Singapore 117456, Singapore
3
Neurobiology/Ageing Programme, National University of Singapore, Singapore 117456, Singapore
4
Research Group Protein trafficking in synaptic development and function, Department of Neurobiology, Max-Planck-Institute for Biophysical Chemistry, Göttingen 37077, Germany
*
Author to whom correspondence should be addressed.
Cells 2016, 5(1), 7; https://doi.org/10.3390/cells5010007
Submission received: 12 January 2016
/
Accepted: 3 February 2016
/
Published: 6 February 2016
(This article belongs to the Special Issue Regulation and Function of Small GTPases)
{kind=link}
Abstract
:Presynaptic neurotransmitter release is dominated by the synaptic vesicle (SV) cycle and entails the biogenesis, fusion, recycling, reformation or turnover of synaptic vesicles—a process involving bulk movement of membrane and proteins. As key mediators of membrane trafficking, small GTPases from the Rab family of proteins play critical roles in this process by acting as molecular switches that dynamically interact with and regulate the functions of different sets of macromolecular complexes involved in each stage of the cycle. Importantly, mutations affecting Rabs, and their regulators or effectors have now been identified that are implicated in severe neurological and neurodevelopmental disorders. Here, we summarize the roles and functions of presynaptic Rabs and discuss their involvement in the regulation of presynaptic function.
1. Introduction
Rab proteins comprise the largest subgroup of the Ras family of small GTPases [1]. They are evolutionarily conserved and occur in all eukaryotes. Since their initial discovery in yeast and mammals [2,3], our knowledge about their function and molecular mechanisms of action has grown exponentially. Rab proteins are capable of alternating between a GTP bound and a GDP bound state corresponding to the active and inactive states, respectively. An assortment of GTPase-activating proteins (GAP) and guanine nucleotide exchange factors (GEF) act to regulate GTP hydrolysis or the exchange of GDP with GTP [4,5,6]. In addition, nucleotide cycling of Rab proteins is tightly coupled to their membrane association/dissociation cycle, a process requiring two additional factors, Rab GDP dissociation inhibitor (Rab GDI) and GDI-displacement factor (GDF) [7]. Thus, the switching between the active and inactive state is subjected to an exquisite level of spatial and temporal coordination.
Rab proteins exert their function by recruiting specific effectors from the cytoplasm exclusively to the GTP-state of the protein. GTP binding induces major conformational changes in a specific region called switch I and switch II that are exposed on the surface of the Rab proteins to which effectors bind [8]. Although structurally similar, variations in the interface of switch regions confer unique specificity in the interaction of individual Rabs with their respective effectors, thereby creating the impressive diversity in Rab protein function and localization [9].
Rab proteins are essential regulators of various intracellular membrane trafficking steps in the secretory pathway such as cargo selection during vesicle formation, vesicle transport, tethering, and docking. Indeed, Rab proteins are crucial for defining the identity of subcellular membranes, thus serving as molecular zip codes in membrane traffic [10,11]. Each subcellular compartment or trafficking organelle has its own set of Rab proteins controlling its biogenesis, maturation, and transition as well as interaction with other membranous compartments. This includes not only trafficking steps conserved in all eukaryotes but also specialized adaptations such as regulated exocytosis in neurons that govern synaptic transmission. In this mini review, we will focus on presynaptic Rab proteins and highlight their roles in synaptic organization, synaptic transmission, and protein turnover.
2. An Overview of Membrane Traffic in Presynaptic Nerve Terminals
Synaptic transmission is mediated by the regulated release of neurotransmitters from presynaptic nerve terminals (Figure 1). Before release, neurotransmitters are stored in small trafficking organelles, termed synaptic vesicles, some of which are docked to specialized regions of the presynaptic plasma membrane known as active zones [12,13]. Upon arrival of an action potential, Ca2+ ions enter the nerve terminal and trigger exocytosis of synaptic vesicles. After exocytosis, the SV membrane is retrieved by endocytosis and SV are locally regenerated to allow for another round of exo-endocytotic membrane cycling [14].
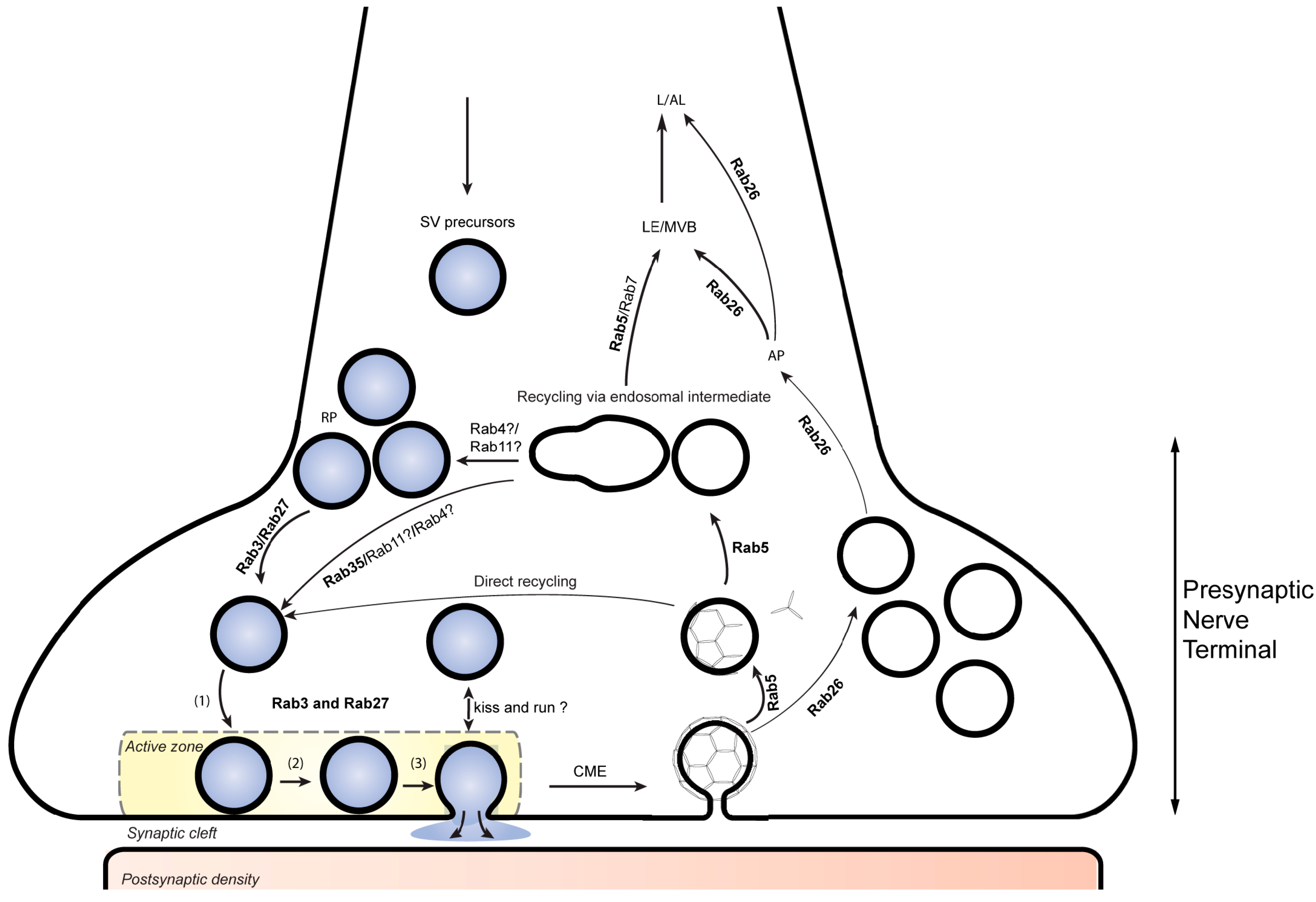
Figure 1.
Rab proteins involved in presynaptic function. RAB3 and RAB27B participate in synaptic vesicle (SV) exocytosis while Rab27b has an addition role in synaptic vesicle recycling at the presynaptic nerve terminal. RAB5 participates in the biogenesis and retrieval of synaptic vesicle components through clathrin-mediate endocytosis. Together with RAB5, RAB35 is also thought to be involved in directing these components through early endosomal compartments that contribute to the reformation of SVs replenishing the SV pool. RAB26 is associated with a subset of recycled SVs and directs them for autophagic degradation. SV, synaptic vesicle; RP, reserve pool (1), docking; (2) priming; (3), fusion. CME, clathrin mediated endocytosis. AP, autophagy; LE/MVB, late endosome/multivesiclular body; L/AL, lysosome/autolysosome.
Figure 1.
Rab proteins involved in presynaptic function. RAB3 and RAB27B participate in synaptic vesicle (SV) exocytosis while Rab27b has an addition role in synaptic vesicle recycling at the presynaptic nerve terminal. RAB5 participates in the biogenesis and retrieval of synaptic vesicle components through clathrin-mediate endocytosis. Together with RAB5, RAB35 is also thought to be involved in directing these components through early endosomal compartments that contribute to the reformation of SVs replenishing the SV pool. RAB26 is associated with a subset of recycled SVs and directs them for autophagic degradation. SV, synaptic vesicle; RP, reserve pool (1), docking; (2) priming; (3), fusion. CME, clathrin mediated endocytosis. AP, autophagy; LE/MVB, late endosome/multivesiclular body; L/AL, lysosome/autolysosome.
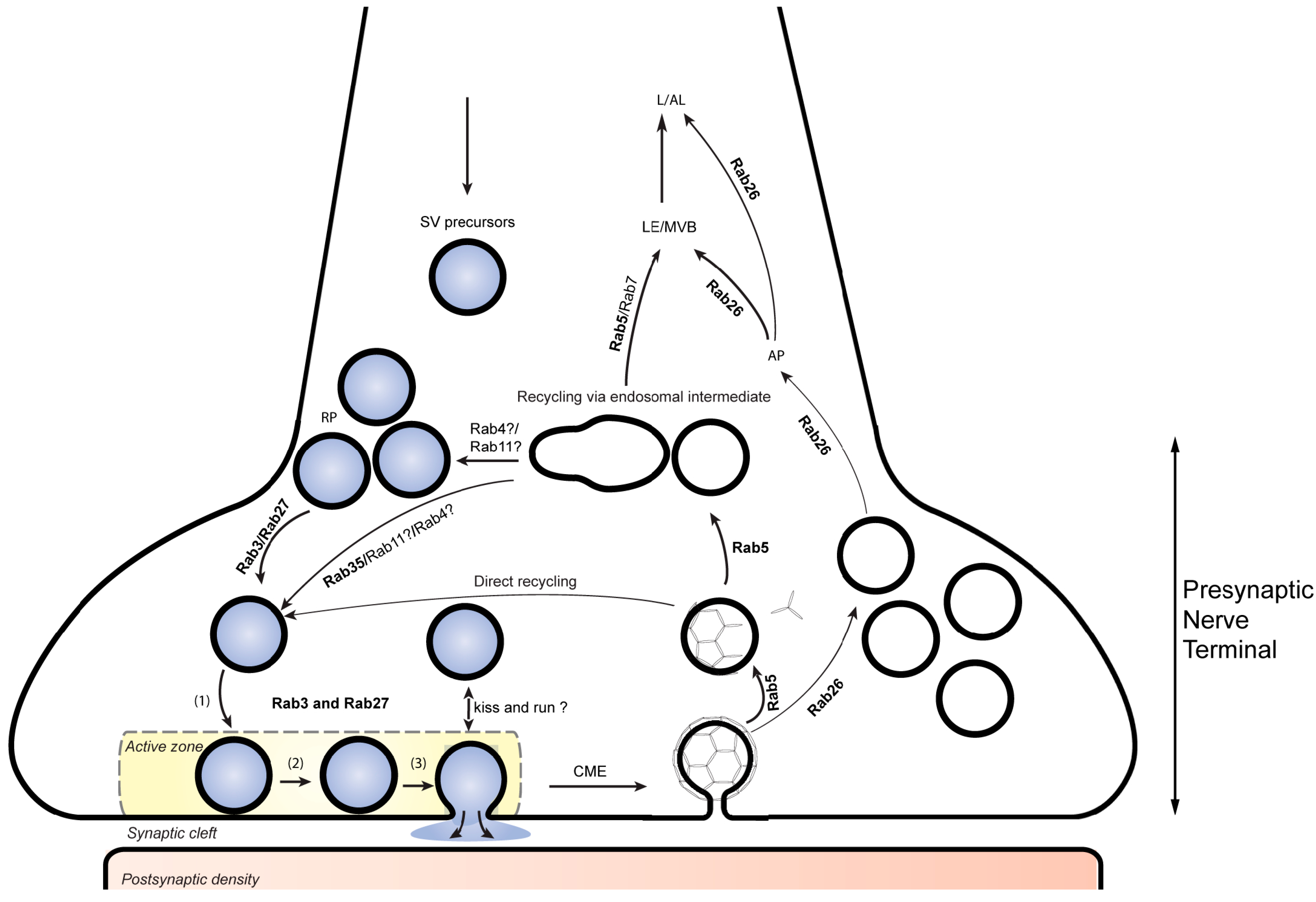
The molecular machineries involved in both Ca2+-dependent exocytosis and endocytosis were intensely studied during the past decades, and most of the molecular components are known. Evidently, the key steps in synaptic vesicle cycling are derived from conserved trafficking steps common to all eukaryotes but contain specialized adaptations. Like most eukaryotic fusion events, exocytotic membrane fusion is mediated by SNARE proteins and requires Sec1/Munc18-like (SM) proteins and the complexes associated with tethering containing helical rods (CATCHR) proteins while Ca2+ regulation is conveyed by the specialist proteins synaptotagmin and complexin [15]. Similarly, endocytosis is dominated by clathrin-mediated endocytosis (CME) that also involves conserved components such as the clathrin components and adaptor complexes, in addition to synapse-specific components such as AP180 and epsin [16]. Along with CME, a fast and clathrin-independent endocytotic pathway operates in the synapse but whose molecular foundation is less clear [17,18]
A unique feature of nerve terminals is the capacity to locally regenerate synaptic vesicles from endocytosed membranes without the involvement of central trafficking platforms such as the Golgi complex. However, the pathways leading to vesicle re-formation are still debated. Synaptic vesicles may be regenerated directly from endocytosed membrane vesicles without an additional trafficking step. For instance, following clathrin uncoating, the resulting vesicles may directly be loaded with neurotransmitter and enter the pool of synaptic vesicles. Alternatively, endocytosed vesicles may need to pass through an early endosomal compartment from which SVs re-form by budding [19]. Also, only limited information is available about the biosynthetic and degradative pathways that are responsible for the creation of new SV and the elimination of old SV or damaged SV components. SV precursors are derived from the trans-Golgi Network and are transported through the axon to the nerve terminal, but it is unclear whether they are fully functional upon reaching synapses or need to undergo fusion-fission events before becoming competent for regulated exocytosis [20,21]. Similarly, the degradation of SVs or SV components is likely to involve proteasomal, lysosomal, and autophagosomal pathways but their relative contribution is not understood.
As all eukaryotic trafficking steps, the synaptic vesicle cycle is governed by specific Rab GTPases. These include sets of specific Rabs involved in exocytosis (RAB3 family), in endosomal function (RAB5, and probably also RAB4, RAB11, and RAB35) and, as recently discovered, Rab proteins involved in autophagosomal function (RAB26 and RAB33).
3. Synaptic Vesicle Docking and Exocytosis by Rab3-Related Proteins
The members of the RAB3 protein subfamily represent the most abundant small GTPases in the brain. They include four closely related homologs (RAB3A, RAB3B, RAB3C, and RAB3D) of which RAB3A, RAB3B, and RAB3C are highly expressed in neurons where they specifically localize to synaptic vesicles [14,22]. RAB3A is best investigated and was shown to be involved in regulating the efficiency of neurotransmitter release in neurons [23,24]. RAB3-GTP is bound to the membrane of SVs from which it dissociates during neurotransmitter release, concomitant with GTP hydrolysis [25,26]. However, though mice deficient in all RAB3 isoforms die immediately after birth, only a surprisingly mild reduction (30%) in neurotransmitter release was observed [27]. This may be due to functional redundancy with the structurally related RAB27B which is also expressed at high levels on the membrane of synaptic vesicles, partially overlapping with RAB3 and which shares with RAB3 a common set of regulators and effectors [28,29,30]. Indeed, overexpression of RAB27B mutants (the GTP and GDP preferring variants) caused a strong reduction in SV recycling [22]. Furthermore, impairment of synaptic function was observed upon knockout of RAB3GEF in mice and in Caenorhabiditis elegans [31,32,33]. RAB3GEF regulates GTP-exchange of both RAB3 proteins and RAB27B [34]. However, it remains to be established whether RAB27B is a true functional isoform of RAB3 or whether it performs additional functions [35,36]
How are the RAB3 family members regulating exocytosis? Two effectors are known: rabphilin and RIM [28,29,37]. Whereas knockout of rabphilin had only minimal effects on neurotransmitter release [38], knockout of RIM impaired neurotransmitter release although it did not result in a complete inhibition [39,40]. RIM1 is one of the core components of the active zone, which forms a trimeric complex with RAB3 and the CATCHR protein MUNC13 [41,42]. In agreement with the canonical function of Rab proteins in tethering vesicles to target membranes, RIM1 and MUNC13 were proposed to recruit SVs to the active zone and ready them for release [42,43]. Unlike in other trafficking steps, however, recruitment of SVs to the active zone is not only dependent on Rabs, but also on several other factors including the SNAREs themselves [44]. This may explain why knockout of the exocytotic Rab machinery in neurons does impair but not abrogate docking and fusion as it does in other trafficking steps such as constitutive exocytosis in yeast [45,46].
4. GTPases Involved in Recycling of SVs
In addition to these exocytotic Rabs, careful analyses of the vesicle composition also confirmed the presence of a specific subset of endosomal Rabs on highly purified synaptic vesicles [22]. These include RAB4 and RAB5 (early endosome, EE) as well as RAB4, RAB11, and RAB35 (recycling endosomes, RE) [47,48]. In addition, Rab proteins associated with autophagy were also identified on these vesicles (e.g., RAB26 and RAB33) suggesting their involvement in eliminating old/damaged synaptic vesicle proteins via the autophagy pathway (see the next section).
Recycling of SVs via the clathrin-mediated pathway presumably occurs via an endosomal intermediate [16,19]. However, the origins and identity of this compartment remains a subject of debate. Of the endosomal Rab proteins, RAB5 coordinates trafficking in the early endocytic events by recruiting a host of RAB5-specific effectors to endocytic vesicles targeted for fusion [49,50]. The involvement of RAB5 in SV recycling was uncovered when the GTPase was found to label an endosomal compartment present at nerve terminals [51,52]. Subsequent experiments additionally showed that RAB5 was directly involved in synaptic vesicle recycling and SV morphology, with RAB5 mutants affecting the formation of these compartments and impairing SV recycling [53,54].
In addition to RAB5, RAB35 is another small GTPase that has been more recently shown to participate in the recycling of SVs via the presynaptic endosomal compartment. In Drosophila, expression of constitutively active RAB35 or loss of function of its corresponding GAP, Skywalker, promotes retrieval of synaptic vesicles via endosomal intermediates and increases synaptic transmission indicating that RAB35, like RAB5, is important for regeneration of new synaptic vesicles [55].
5. Degradation and Turnover of SVs
Regulated turnover of proteins is essential for synaptic plasticity and synaptic remodeling. Periodic turnover of synaptic vesicles and their proteins is also necessary to remove damaged components. While proteasomal degradation of proteinaceous synaptic material is documented, less is known about how synaptic vesicle membranes and their associated proteins are removed [56,57]. Apart from proteasomal degradation of membrane-associated proteins, it is conceivable that membrane constituents are delivered to lysosomes after sorting into multivesicular bodies by the ESCRT pathway. Indeed, multivesicular bodies containing synaptic proteins were found in axons, suggesting that this pathway also operates in nerve terminals [58].
Besides the proteasome pathway, autophagic clearance of cellular components also plays important roles in neurons [59,60]. Autophagosomes can form in distal axons and are transported back to neuronal cell bodies for degradation following fusion with lysosomes [61,62]. Several Rab proteins (such as RAB7, RAB8, and RAB33) and their effectors are known to be involved in regulating various steps in the autophagy pathway [63]. Recently, we discovered a direct link between the turnover of recycled SV and the autophagic pathway that involves RAB26.
In neurons, RAB26 is enriched in SVs and adorns a subset of SVs that have undergone recycling [64]. Overexpression of constitutively active RAB26 causes the appearance of large intracellular vesicular aggregates containing synaptic vesicle proteins such as synaptobrevin and RAB3. Remarkably, these aggregates are positive for autophagosomal markers such as ATG16L1 and LC3B. Strikingly, RAB26 interacts with a complex of ATG16L1-ATG5, indicating that the GTPase plays a direct role in earmarking synaptic vesicles for degradation via the autophagyc pathway.
In addition to RAB26, the two isoforms of RAB33 have also been shown to bind to ATG16L1 [65]. While RAB33A is highly expressed in the brain, Rab33B is ubiquitously present. Interestingly, the interaction of RAB33B with Atg16L1 regulates autophagosome formation. In comparison to this, the interaction of RAB33A with Atg16L1 does not appear to play a direct role in autophagy but serves rather to regulate hormone secretion and mediate anterograde transport of Golgi-derived vesicles involved in the exocytosis in an autophagy-independent manner [66,67]. Since RAB26 and RAB33B both share a common effector, it will be important to determine if RAB26 and RAB33 potentially overlap in their roles in autophagy in neurons to regulate the turnover of SVs.
6. Presynaptic Rabs and Their Involvement in the Neuronal-Associated Disorders
Many mutations and physiological abnormalities affecting synaptic Rabs have now been identified that contribute to neurodegenerative and neurodevelopmental disorders. Aggregates of α-synuclein trap RAB3A and prevent its interaction with rabphilin, suggesting that exocytosis and neurotransmitter release are affected in Lewy´s Body diseases [68]. Patients affected by the Warburg-Micro-syndrome and the Martsolf-syndrome, which is a more attenuated form of the former, carry mutations in the catalytic and non-catalytic domains of RAB3GAP. Both are inheritable forms of neurological diseases exhibiting developmental abnormalities and intellectual disability [69,70]. Patients affected by X-linked, non-specific mental retardation harbor mutations in αGDI (Rab GDP-dissociator inhibitor). One such mutation (L92P) affects RAB3A recycling which might indicate that proper SV cycling is fundamental for mental development [71]. Interestingly, loss of αGDI in mice affects short term memory and social behavior and Rab3a null mice also exibit impaired learning, albeit with different phenotypes compared to the ones observed for αGDI [72,73]. Recent work also revealed that knock out of Rab3b gene affects long term depression (LTD) of inhibitory synapses and enhanced reversal learning in mice [74].
Abnormalities in the endosomal pathway are found in some of the commonly known neurodegeneratve diseases such as Alzheimer’s (AD), Huntington’s (HD), and Parkinson’s disease (PD) (for review see [75]). For instance, RAB5 appears to play a role in the internalization of exogenous α-Synuclein and overexpression of constitutively active RAB5 or its effector Rabaptin leads to the formation of intracellular inclusions in cultured neurons resembling Lewy´s bodies [76]. ALS2/alsin is a RAB5GEF and its depletion is responsible for different recessive forms of motor neuron diseases. This implies that RAB5 activity, and by inference the endosome pathway, plays a crucial role in contributing to severe motor neurons disorders such as Amyotrophic Lateral Sclerosis (ALS) [77,78]. Huntingtin-HAP40, a RAB5 effector, is upregulated in Huntington’s disease (HD) suggesting that RAB5 is also implicated in this disorder [79,80].
Many neurological diseases show defects in recycling endosomes [81]. Recent studies uncovered mutations in RAB35GAP (also known as Skywalker or TBC1D24) in patients with familial infantile myoclonic epilepsy or focal epilepsy with intellectual disability syndrome [82,83]. RAB11 family (including the ubiquitously expressed RAB11A, the neuronal RAB11B and the epithelial RAB25) is also an important member of the recycling pathway and deregulated activity of RAB11 is found in many neuronal disorders [11,84,85]. For instance, the function of RAB11 was found to be inhibited in many models of HD [86]. RAB11 is also implicated in Alzheimer’s disease by its ability to bind presenilin 1 and presenilin 2, which are components of the γ-secretease responsible for the generation of Aβ peptides [87].
Intracellular membrane trafficking pathologies might be causative or a consequence of many neurodegenerative diseases. Studying the role of the small GTPases and their effectors and regulators at the molecular level would improve our understanding the pathology of these human diseases and may in turn offer new avenues for therapeutic intervention.
Acknowledgments
The work in J.C’s lab is supported by funding from the Deutsche Forschungsgemeinschaft (grant no. CH 1385/1-1) and by a startup grant from the National University of Singapore.
Author Contributions
J.C. conceived the manuscript; B.B., R.J and J.C wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stenmark, H.; Olkkonen, V.M. The rab gtpase family. Genome Biol. 2001, 2, S3007. [Google Scholar] [CrossRef]
- Gallwitz, D.; Donath, C.; Sander, C. A yeast gene encoding a protein homologous to the human c-has/bas proto-oncogene product. Nature 1983, 306, 704–707. [Google Scholar] [CrossRef] [PubMed]
- Zahraoui, A.; Touchot, N.; Chardin, P.; Tavitian, A. The human rab genes encode a family of gtp-binding proteins related to yeast ypt1 and sec4 products involved in secretion. J. Biol. Chem. 1989, 264, 12394–12401. [Google Scholar] [PubMed]
- Cherfils, J.; Zeghouf, M. Regulation of small gtpases by gefs, gaps, and gdis. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef] [PubMed]
- Goody, R.S.; Rak, A.; Alexandrov, K. The structural and mechanistic basis for recycling of rab proteins between membrane compartments. Cell. Mol. Life Sci. 2005, 62, 1657–1670. [Google Scholar] [CrossRef] [PubMed]
- Barr, F.; Lambright, D.G. Rab gefs and gaps. Curr. Opin. Cell. Biol. 2010, 22, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.; Aivazian, D. Targeting rab gtpases to distinct membrane compartments. Nat. Rev. Mol. Cell Biol. 2004, 5, 886–896. [Google Scholar] [CrossRef] [PubMed]
- Vetter, I.R.; Wittinghofer, A. The guanine nucleotide-binding switch in three dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef] [PubMed]
- Ostermeier, C.; Brunger, A.T. Structural basis of rab effector specificity: Crystal structure of the small g protein rab3a complexed with the effector domain of rabphilin-3a. Cell 1999, 96, 363–374. [Google Scholar] [CrossRef]
- Barr, F.A. Review series: Rab gtpases and membrane identity: Causal or inconsequential? J. Cell Biol. 2013, 202, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Zerial, M.; McBride, H. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell. Biol. 2001, 2, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Sudhof, T.C. Neurotransmitter release: The last millisecond in the life of a synaptic vesicle. Neuron 2013, 80, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Chua, J.J. Macromolecular complexes at active zones: Integrated nano-machineries for neurotransmitter release. Cell. Mol. Life Sci. 2014, 71, 3903–3916. [Google Scholar] [CrossRef] [PubMed]
- Sudhof, T.C. The synaptic vesicle cycle. Annu. Rev. Neurosci. 2004, 27, 509–547. [Google Scholar] [CrossRef] [PubMed]
- Jahn, R.; Fasshauer, D. Molecular machines governing exocytosis of synaptic vesicles. Nature 2012, 490, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Saheki, Y.; De Camilli, P. Synaptic vesicle endocytosis. Cold Spring Harb. Perspect. Biol. 2012, 4, a005645. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Liu, Q.; Davis, M.W.; Hollopeter, G.; Thomas, N.; Jorgensen, N.B.; Jorgensen, E.M. Ultrafast endocytosis at caenorhabditis elegans neuromuscular junctions. Elife 2013, 2, e00723. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, S.O.; Jahn, R. Kiss-and-run, collapse and 'readily retrievable' vesicles. Traffic 2007, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Jahne, S.; Rizzoli, S.O.; Helm, M.S. The structure and function of presynaptic endosomes. Exp. Cell Res. 2015, 335, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Hannah, M.J.; Schmidt, A.A.; Huttner, W.B. Synaptic vesicle biogenesis. Annu. Rev. Cell Dev. Biol. 1999, 15, 733–798. [Google Scholar] [CrossRef] [PubMed]
- Shupliakov, O.; Brodin, L. Recent insights into the building and cycling of synaptic vesicles. Exp. Cell Res. 2010, 316, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Pavlos, N.J.; Gronborg, M.; Riedel, D.; Chua, J.J.; Boyken, J.; Kloepper, T.H.; Urlaub, H.; Rizzoli, S.O.; Jahn, R. Quantitative analysis of synaptic vesicle rabs uncovers distinct yet overlapping roles for rab3a and rab27b in ca2+-triggered exocytosis. J. Neurosci. 2010, 30, 13441–13453. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Sasaki, T.; Shirataki, H.; Nakanishi, H. Rab3a small gtp-binding protein in ca2+-dependent exocytosis. Genes Cells 1996, 1, 615–632. [Google Scholar] [CrossRef] [PubMed]
- Geppert, M.; Goda, Y.; Stevens, C.F.; Sudhof, T.C. The small gtp-binding protein rab3a regulates a late step in synaptic vesicle fusion. Nature 1997, 387, 810–814. [Google Scholar] [PubMed]
- Fischer von Mollard, G.; Sudhof, T.C.; Jahn, R. A small gtp-binding protein dissociates from synaptic vesicles during exocytosis. Nature 1991, 349, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Stahl, B.; von Mollard, G.F.; Walch-Solimena, C.; Jahn, R. Gtp cleavage by the small gtp-binding protein rab3a is associated with exocytosis of synaptic vesicles induced by alpha-latrotoxin. J. Biol. Chem. 1994, 269, 24770–24776. [Google Scholar] [PubMed]
- Schluter, O.M.; Schmitz, F.; Jahn, R.; Rosenmund, C.; Sudhof, T.C. A complete genetic analysis of neuronal rab3 function. J. Neurosci. 2004, 24, 6629–6637. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M. Distinct rab binding specificity of rim1, rim2, rabphilin, and noc2. Identification of a critical determinant of rab3a/rab27a recognition by rim2. J. Biol. Chem. 2003, 278, 15373–15380. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M. Rab27 effectors, pleiotropic regulators in secretory pathways. Traffic 2013, 14, 949–963. [Google Scholar] [CrossRef] [PubMed]
- Pavlos, N.J.; Jahn, R. Distinct yet overlapping roles of rab gtpases on synaptic vesicles. Small GTPases 2011, 2, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Staunton, J.; Saifee, O.; Nonet, M.; Thomas, J.H. Aex-3 encodes a novel regulator of presynaptic activity in c. Elegans. Neuron 1997, 18, 613–622. [Google Scholar] [CrossRef]
- Nonet, M.L.; Staunton, J.E.; Kilgard, M.P.; Fergestad, T.; Hartwieg, E.; Horvitz, H.R.; Jorgensen, E.M.; Meyer, B.J. Caenorhabditis elegans rab-3 mutant synapses exhibit impaired function and are partially depleted of vesicles. J. Neurosci. 1997, 17, 8061–8073. [Google Scholar] [PubMed]
- Figueiredo, A.C.; Wasmeier, C.; Tarafder, A.K.; Ramalho, J.S.; Baron, R.A.; Seabra, M.C. Rab3gep is the non-redundant guanine nucleotide exchange factor for Rab27a in melanocytes. J. Biol. Chem. 2008, 283, 23209–23216. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, T.R.; Liu, Q.; Itoh, T.; Luo, S.; Hadwiger, G.; Vincent, R.; Wang, Z.W.; Fukuda, M.; Nonet, M.L. Regulation of synaptic transmission by rab-3 and rab-27 in caenorhabditis elegans. Mol. Boil. Cell 2006, 17, 2617–2625. [Google Scholar] [CrossRef] [PubMed]
- Itzen, A.; Goody, R.S. Key determinants of rab specificity. Structure 2008, 16, 1437–1439. [Google Scholar] [CrossRef] [PubMed]
- Handley, M.T.; Haynes, L.P.; Burgoyne, R.D. Differential dynamics of rab3a and rab27a on secretory granules. J. Cell Sci. 2007, 120, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M. Membrane traffic in the secretory pathway. Cell. Mol. Life Sci. 2008, 65, 2801–2813. [Google Scholar] [CrossRef] [PubMed]
- Schluter, O.M.; Schnell, E.; Verhage, M.; Tzonopoulos, T.; Nicoll, R.A.; Janz, R.; Malenka, R.C.; Geppert, M.; Sudhof, T.C. Rabphilin knock-out mice reveal that rabphilin is not required for rab3 function in regulating neurotransmitter release. J. Neurosci. 1999, 19, 5834–5846. [Google Scholar] [PubMed]
- Schoch, S.; Mittelstaedt, T.; Kaeser, P.S.; Padgett, D.; Feldmann, N.; Chevaleyre, V.; Castillo, P.E.; Hammer, R.E.; Han, W.; Schmitz, F.; et al. Redundant functions of rim1alpha and rim2alpha in ca2+-triggered neurotransmitter release. EMBO J. 2006, 25, 5852–5863. [Google Scholar] [CrossRef] [PubMed]
- Schoch, S.; Castillo, P.E.; Jo, T.; Mukherjee, K.; Geppert, M.; Wang, Y.; Schmitz, F.; Malenka, R.C.; Sudhof, T.C. Rim1alpha forms a protein scaffold for regulating neurotransmitter release at the active zone. Nature 2002, 415, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Kaeser, P.S.; Xu, W.; Südhof, T.C. Rim proteins activate vesicle priming by reversing autoinhibitory homodimerization of munc13. Neuron 2011, 69, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Dulubova, I.; Lou, X.; Lu, J.; Huryeva, I.; Alam, A.; Schneggenburger, R.; Sudhof, T.C.; Rizo, J. A munc13/rim/rab3 tripartite complex: From priming to plasticity? EMBO J. 2005, 24, 2839–2850. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Okamoto, M.; Schmitz, F.; Hofmann, K.; Sudhof, T.C. Rim is a putative rab3 effector in regulating synaptic-vesicle fusion. Nature 1997, 388, 593–598. [Google Scholar] [PubMed]
- Imig, C.; Min, S.-W.; Krinner, S.; Arancillo, M.; Rosenmund, C.; Südhof, T.C.; Rhee, J.; Brose, N.; Cooper, B.H. The morphological and molecular nature of synaptic vesicle priming at presynaptic active zones. Neuron 2014, 84, 416–431. [Google Scholar] [CrossRef] [PubMed]
- Novick, P.; Ferro, S.; Schekman, R. Order of events in the yeast secretory pathway. Cell 1981, 25, 461–469. [Google Scholar] [CrossRef]
- Novick, P.; Schekman, R. Secretion and cell-surface growth are blocked in a temperature-sensitive mutant of saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1979, 76, 1858–1862. [Google Scholar] [CrossRef] [PubMed]
- Villarroel-Campos, D.; Gastaldi, L.; Conde, C.; Caceres, A.; Gonzalez-Billault, C. Rab-mediated trafficking role in neurite formation. J. Neurochem. 2014, 129, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.; Kiger, A.A. Coordination between rab gtpase and phosphoinositide regulation and functions. Nat. Rev. Mol. Cell Biol. 2012, 13, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, H. Rab gtpases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Fischer von Mollard, G.; Stahl, B.; Walch-Solimena, C.; Takei, K.; Daniels, L.; Khoklatchev, A.; De Camilli, P.; Sudhof, T.C.; Jahn, R. Localization of rab5 to synaptic vesicles identifies endosomal intermediate in synaptic vesicle recycling pathway. Eur. J. Cell. Biol. 1994, 65, 319–326. [Google Scholar] [PubMed]
- de Hoop, M.J.; Huber, L.A.; Stenmark, H.; Williamson, E.; Zerial, M.; Parton, R.G.; Dotti, C.G. The involvement of the small gtp-binding protein rab5a in neuronal endocytosis. Neuron 1994, 13, 11–22. [Google Scholar] [CrossRef]
- Wucherpfennig, T.; Wilsch-Brauninger, M.; Gonzalez-Gaitan, M. Role of drosophila rab5 during endosomal trafficking at the synapse and evoked neurotransmitter release. J. Cell Boil. 2003, 161, 609–624. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Kawamura, S.; Ozaki, K. An essential role of Rab5 in uniformity of synaptic vesicle size. J. Cell Sci. 2003, 116, 3583–3590. [Google Scholar] [CrossRef] [PubMed]
- Uytterhoeven, V.; Kuenen, S.; Kasprowicz, J.; Miskiewicz, K.; Verstreken, P. Loss of skywalker reveals synaptic endosomes as sorting stations for synaptic vesicle proteins. Cell 2011, 145, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Haas, K.F.; Broadie, K. Roles of ubiquitination at the synapse. Biochim. Biophys. Acta 2008, 1779, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Cajigas, I.J.; Will, T.; Schuman, E.M. Protein homeostasis and synaptic plasticity. EMBO J. 2010, 29, 2746–2752. [Google Scholar] [CrossRef] [PubMed]
- Von Bartheld, C.S.; Altick, A.L. Multivesicular bodies in neurons: Distribution, protein content, and trafficking functions. Prog. Neurobiol. 2011, 93, 313–340. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Yue, Z. Autophagy and its normal and pathogenic states in the brain. Annu. Rev. Neurosci. 2014, 37, 55–78. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.L.; Cuervo, A.M. Autophagy and human disease: Emerging themes. Curr. Opin. Genet. Dev. 2014, 26, 16–23. [Google Scholar]
- Cheng, X.T.; Zhou, B.; Lin, M.Y.; Cai, Q.; Sheng, Z.H. Axonal autophagosomes recruit dynein for retrograde transport through fusion with late endosomes. J. Cell Biol. 2015, 209, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Maday, S.; Holzbaur, E.L. Autophagosome biogenesis in primary neurons follows an ordered and spatially regulated pathway. Dev. Cell 2014, 30, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Szatmari, Z.; Sass, M. The autophagic roles of rab small gtpases and their upstream regulators: A review. Autophagy 2014, 10, 1154–1166. [Google Scholar] [CrossRef] [PubMed]
- Binotti, B.; Pavlos, N.J.; Riedel, D.; Wenzel, D.; Vorbruggen, G.; Schalk, A.M.; Kuhnel, K.; Boyken, J.; Erck, C.; Martens, H.; Chua, J.J.; et al. The gtpase rab26 links synaptic vesicles to the autophagy pathway. Elife 2015, 4, e00597. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Itoh, T. Direct link between atg protein and small gtpase rab: Atg16l functions as a potential rab33 effector in mammals. Autophagy 2008, 4, 824–826. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Uemura, T.; Waguri, S.; Fukuda, M. Atg16l1, an essential factor for canonical autophagy, participates in hormone secretion from pc12 cells independently of autophagic activity. Mol. Biol. Cell 2012, 23, 3193–3202. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, H.; Sada, T.; Toriyama, M.; Tago, K.; Sugiura, T.; Fukuda, M.; Inagaki, N. Rab33a mediates anterograde vesicular transport for membrane exocytosis and axon outgrowth. J. Neurosci. 2012, 32, 12712–12725. [Google Scholar] [PubMed]
- Dalfo, E.; Barrachina, M.; Rosa, J.L.; Ambrosio, S.; Ferrer, I. Abnormal alpha-synuclein interactions with rab3a and rabphilin in diffuse lewy body disease. Neurobiol. Dis. 2004, 16, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Aligianis, I.A.; Johnson, C.A.; Gissen, P.; Chen, D.; Hampshire, D.; Hoffmann, K.; Maina, E.N.; Morgan, N.V.; Tee, L.; Morton, J.; et al. Mutations of the catalytic subunit of rab3gap cause warburg micro syndrome. Nat. Genet. 2005, 37, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Aligianis, I.A.; Morgan, N.V.; Mione, M.; Johnson, C.A.; Rosser, E.; Hennekam, R.C.; Adams, G.; Trembath, R.C.; Pilz, D.T.; Stoodley, N.; et al. Mutation in Rab3 GTPase-activating protein (rab3gap) noncatalytic subunit in a kindred with martsolf syndrome. Am. J. Hum. Genet. 2006, 78, 702–707. [Google Scholar] [CrossRef] [PubMed]
- D'Adamo, P.; Menegon, A.; Lo Nigro, C.; Grasso, M.; Gulisano, M.; Tamanini, F.; Bienvenu, T.; Gedeon, A.K.; Oostra, B.; Wu, S.K.; et al. Mutations in gdi1 are responsible for x-linked non-specific mental retardation. Nat. Genet. 1998, 19, 134–139. [Google Scholar] [CrossRef] [PubMed]
- D'Adamo, P.; Welzl, H.; Papadimitriou, S.; Raffaele di Barletta, M.; Tiveron, C.; Tatangelo, L.; Pozzi, L.; Chapman, P.F.; Knevett, S.G.; Ramsay, M.F.; et al. Deletion of the mental retardation gene gdi1 impairs associative memory and alters social behavior in mice. Hum. Mol. Genet. 2002, 11, 2567–2580. [Google Scholar] [CrossRef] [PubMed]
- D'Adamo, P.; Wolfer, D.P.; Kopp, C.; Tobler, I.; Toniolo, D.; Lipp, H.P. Mice deficient for the synaptic vesicle protein rab3a show impaired spatial reversal learning and increased explorative activity but none of the behavioral changes shown by mice deficient for the rab3a regulator gdi1. Eur. J. Neurosci. 2004, 19, 1895–1905. [Google Scholar] [CrossRef] [PubMed]
- Tsetsenis, T.; Younts, T.J.; Chiu, C.Q.; Kaeser, P.S.; Castillo, P.E.; Sudhof, T.C. Rab3b protein is required for long-term depression of hippocampal inhibitory synapses and for normal reversal learning. Proc. Natl. Acad. Sci. USA 2011, 108, 14300–14305. [Google Scholar] [CrossRef] [PubMed]
- Baskys, A.; Bayazitov, I.; Zhu, E.; Fang, L.; Wang, R. Rab-mediated endocytosis: Linking neurodegeneration, neuroprotection, and synaptic plasticity? Ann. N. Y. Acad. Sci. 2007, 1122, 313–329. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Kim, J.; Paik, S.R.; Park, J.H.; Ahn, Y.S.; Chung, K.C. Induction of neuronal cell death by rab5a-dependent endocytosis of alpha-synuclein. J. Boil. Chem. 2001, 276, 27441–27448. [Google Scholar] [CrossRef] [PubMed]
- Otomo, A.; Hadano, S.; Okada, T.; Mizumura, H.; Kunita, R.; Nishijima, H.; Showguchi-Miyata, J.; Yanagisawa, Y.; Kohiki, E.; Suga, E.; et al. Als2, a novel guanine nucleotide exchange factor for the small gtpase rab5, is implicated in endosomal dynamics. Hum. Mol. Genet. 2003, 12, 1671–1687. [Google Scholar] [CrossRef] [PubMed]
- Kenna, K.P.; McLaughlin, R.L.; Byrne, S.; Elamin, M.; Heverin, M.; Kenny, E.M.; Cormican, P.; Morris, D.W.; Donaghy, C.G.; Bradley, D.G.; et al. Delineating the genetic heterogeneity of als using targeted high-throughput sequencing. J. Med. Genet. 2013, 50, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Severin, F.; Lommer, B.; Shevchenko, A.; Zerial, M. Huntingtin-hap40 complex is a novel rab5 effector that regulates early endosome motility and is up-regulated in huntington's disease. J. Cell Boil. 2006, 172, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Severin, F.; Hopfner, S.; Zerial, M. Regulation of endosome dynamics by rab5 and huntingtin-hap40 effector complex in physiological versus pathological conditions. Methods Enzymol. 2008, 438, 239–257. [Google Scholar] [PubMed]
- Li, X.; DiFiglia, M. The recycling endosome and its role in neurological disorders. Prog. Neurobiol. 2012, 97, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Guven, A.; Tolun, A. Tbc1d24 truncating mutation resulting in severe neurodegeneration. J. Med. Genet. 2013, 50, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.C.; Uytterhoeven, V.; Kuenen, S.; Wang, Y.C.; Slabbaert, J.R.; Swerts, J.; Kasprowicz, J.; Aerts, S.; Verstreken, P. Reduced synaptic vesicle protein degradation at lysosomes curbs tbc1d24/sky-induced neurodegeneration. J. Cell Boil. 2014, 207, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Casanova, J.E.; Wang, X.; Kumar, R.; Bhartur, S.G.; Navarre, J.; Woodrum, J.E.; Altschuler, Y.; Ray, G.S.; Goldenring, J.R. Association of rab25 and rab11a with the apical recycling system of polarized madin-darby canine kidney cells. Mol. Boil. Cell 1999, 10, 47–61. [Google Scholar] [CrossRef]
- Schlierf, B.; Fey, G.H.; Hauber, J.; Hocke, G.M.; Rosorius, O. Rab11b is essential for recycling of transferrin to the plasma membrane. Exp. Cell Res. 2000, 259, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Giorgini, F.; Steinert, J.R. Rab11 as a modulator of synaptic transmission. Commun. Integr. Biol. 2013, 6, e26807. [Google Scholar] [CrossRef] [PubMed]
- Dumanchin, C.; Czech, C.; Campion, D.; Cuif, M.H.; Poyot, T.; Martin, C.; Charbonnier, F.; Goud, B.; Pradier, L.; Frebourg, T. Presenilins interact with rab11, a small gtpase involved in the regulation of vesicular transport. Hum. Mol. Genet. 1999, 8, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Binotti, B.; Jahn, R.; Chua, J.J.E. Functions of Rab Proteins at Presynaptic Sites. Cells 2016, 5, 7. https://doi.org/10.3390/cells5010007
AMA Style
Binotti B, Jahn R, Chua JJE. Functions of Rab Proteins at Presynaptic Sites. Cells. 2016; 5(1):7. https://doi.org/10.3390/cells5010007
Chicago/Turabian StyleBinotti, Beyenech, Reinhard Jahn, and John Jia En Chua. 2016. "Functions of Rab Proteins at Presynaptic Sites" Cells 5, no. 1: 7. https://doi.org/10.3390/cells5010007
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.