Generating Plants with Improved Water Use Efficiency
by
 ,
,
Sonja Blankenagel
1, 
Zhenyu Yang
2,*,
Viktoriya Avramova
1,
Chris-Carolin Schön
1 and
Erwin Grill
2 1
Plant Breeding, School of Life Sciences Weihenstephan, Technical University of Munich, Liesel-Beckmann-Straße 2, 85354 Freising, Germany
2
Botany, School of Life Sciences Weihenstephan, Technical University of Munich, Emil-Ramann-Straße 4, 85354 Freising, Germany
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(9), 194; https://doi.org/10.3390/agronomy8090194
Submission received: 13 August 2018
/
Revised: 13 September 2018
/
Accepted: 14 September 2018
/
Published: 18 September 2018
(This article belongs to the Special Issue Biotic and Abiotic Stress Responses in Crop Plants)
{kind=link}
{kind=link}
{kind=link}
Abstract
:To improve sustainability of agriculture, high yielding crop varieties with improved water use efficiency (WUE) are needed. Despite the feasibility of assessing WUE using different measurement techniques, breeding for WUE and high yield is a major challenge. Factors influencing the trait under field conditions are complex, including different scenarios of water availability. Plants with C3 photosynthesis are able to moderately increase WUE by restricting transpiration, resulting in higher intrinsic WUE (iWUE) at the leaf level. However, reduced CO2 uptake negatively influences photosynthesis and possibly growth and yield as well. The negative correlation of growth and WUE could be partly disconnected in model plant species with implications for crops. In this paper, we discuss recent insights obtained for Arabidopsis thaliana (L.) and the potential to translate the findings to C3 and C4 crops. Our data on Zea mays (L.) lines subjected to progressive drought show that there is potential for improvements in WUE of the maize line B73 at the whole plant level (WUEplant). However, changes in iWUE of B73 and Arabidopsis reduced the assimilation rate relatively more in maize. The trade-off observed in the C4 crop possibly limits the effectiveness of approaches aimed at improving iWUE but not necessarily efforts to improve WUEplant.
1. Introduction
Green Revolution technologies and significant expansion in the use of land, water, and other natural resources for agricultural purposes have led to a tripling in agricultural production between 1960 and 2015 [1]. Despite this success, the high costs to the natural environment that accompany elevated productivity and changes in the food supply chain threaten the sustainability of food production [1]. Global food security is further challenged by climate change, with a predicted increase in frequency of droughts [2,3]. Globally, agriculture accounts for at least 70% of withdrawals from freshwater resources, with large effects on ecosystems [4,5]. Despite this high water deployment, major yield losses due to water deficits are experienced in crops [6]. At the same time, global population growth increases the demand for food, feed, and fuel, which intensifies the pressure to improve water use efficiency (WUE) of crops [7,8]. While better crop and water management practices provide an immediate opportunity to increase crop water productivity, breeding for superior varieties can achieve a medium- and long-term increase [9,10].
Physiologically, water use efficiency can be defined at different scales [11,12,13]. At the plot level, it represents the ratio of grain or biomass yield to water received or evapotranspired. At the single plant level (WUEplant), it is the ratio of biomass to transpiration. The increase in biomass and amount of water transpired over time can be assessed gravimetrically [14]. However, this is destructive and laborious on a long-term basis, especially regarding large crops like maize and sorghum. Therefore, analyses of intrinsic water use efficiency (iWUE) and carbon isotope discrimination (Δ13C) are used as surrogates when evaluating WUE [11,15]. The iWUE is assessed at the leaf level as the ratio of net CO2 assimilation (An) to stomatal conductance (gs) and can be measured noninvasively with portable gas exchange equipment [16]. As transpiration rate (E) is influenced not only by gs but also by the leaf-to-air vapor pressure deficit (VPD) of the air [17], iWUE usually differs from transpiration efficiency (An/E). In addition, VPD affects the stomatal aperture and therefore gs [17,18]. Extrapolation of gas exchange data from single-leaf to whole plant is error-prone due to differences in photosynthesis and transpiration among leaves [19]. Prediction of long-term biomass accumulation and water consumption, WUEplant, based on iWUE is even more uncertain given the possible differences in VPD and additional physiological processes such as dark respiration and photorespiration influencing the resulting biomass increase [19]. Despite these limitations, analysis of iWUE provides a convenient measure for the water efficiency of carbon capture. The throughput of iWUE analyses is quite low as only single, time-consuming measurements per plant can be taken, which impedes large-scale phenotyping.
Analysis of stable carbon isotope discrimination (Δ13C) offers a suitable alternative in C3 plants by providing a read-out for transpiration efficiency integrated over time. Discrimination of the heavier isotope is mainly caused by differences in diffusion rates of the isotopes and enzymatic discrimination during carboxylation reactions [20]. Therefore, Δ13C has been used as an indirect trait to select cultivars with improved WUE [21,22,23,24]. By combining the analysis with oxygen isotope enrichment Δ18O, an estimation for transpiration rate [25,26,27], contributions of water loss, and CO2 assimilation on iWUE could be disentangled [28,29,30]. Stable isotope compositions of leaves or grains, however, represent integrated measures of many processes over a period of plant growth and therefore correlation with iWUE can be limited [31]. In C4 plants, CO2 prefixation, for instance, by phosphoenolpyruvat carboxylase and bundle sheath leakiness restrict the responsiveness of Δ13C to changes in WUE [20,32] and make the relationship between Δ13C, gs and WUE in C4 species less predictable compared to C3 plants [13].
Improving WUE of crops is considered beneficial in very dry climates and in very severe and terminal drought conditions, while growth maintenance traits are advantageous under milder drought conditions [33,34,35]. For crops experiencing water deficit early in their development, traits found to be positive for improving WUE are negative for yield [36]. Enhanced water uptake through investments in the root system can result in reduced plant size and water expenditure for growth maintenance can result in increased drought stress experiences if plants are growing at very low soil water availability [33,34,36,37]. Hence, water-conserving traits as imposed by higher WUE would be beneficial, provided growth and yield are not negatively affected.
2. Disconnecting Improved WUE and Growth Trade-Offs
Being a ratio, iWUE can be improved by reducing gs per amount of CO2 assimilated or by enhancing the assimilation rate at a given gs. Both cases result in lowered intercellular CO2 concentration (Ci) and consequently in an increased stomatal CO2 gradient (Ca − Ci, with external CO2 concentration Ca), which is directly proportional to the ratio of An to gs according to Fick’s law applied to carbon assimilation in leaves, An = gs (Ca − Ci) [16,38,39]. Increased iWUE has been observed in several C3 species under water deficit conditions when plants reduce gs [11,15,40,41,42], although a decrease in gs caused by drought was found to be overridden by heat stress [43]. However, closing stomatal pores to reduce transpiration often results in a reduction of An [41,44]. Lowering gs impinges on Ci and unless this change in Ci is counteracted by an elevated mesophyll conductance (gm), the CO2 concentration at the site of Rubisco-dependent carboxylation (Cc, CO2 concentration in chloroplasts) will be reduced [39,45,46]. A reduction of Cc affects the carboxylation efficiency of Rubisco and favors photorespiration [47,48]. Sustaining net photosynthesis under these conditions might require a higher electron transfer rate (ETR) and/or reduced nonphotochemical quenching to support enhanced carboxylation by Rubisco for compensation of enhanced photorespiration [49,50,51,52]. There are reports that water deficit results in increased gm; however, in most analyses, no change or a reduced gm was observed under drought [44,53,54,55,56,57,58].
Gains in WUE are often associated with growth trade-offs [59,60]. As pointed out by Blum [61], crops with high CO2 assimilation and high biomass accumulation per unit land area require high stomatal conductance. This is supported by the observation of a constant WUE on the field level over a broad range of yields [8]. Nevertheless, there might be ways to achieve elevated WUE and high photosynthesis, namely by exploring CO2 concentrating mechanisms, increased gm, and increased CO2 specificity of Rubisco [10].
Interestingly, several reports of C3 plants have shown enhanced iWUE without the expected negative impact on An or growth [11,15,62,63,64], as postulated by plant physiologists [4]. In these studies with transgenic tomato and Arabidopsis plants, gs was moderately reduced by enhancing the biosynthesis or the responsiveness to the phytohormone abscisic acid (ABA) or by reducing the size and density of leaf stomata [15,63,64,65]. Plants overexpressing distinct ABA receptors—termed ABA-Binding Regulatory Component (RCAR)/Pyrabactin Resistance 1-(like) (PYR1/PYL)—caused increases of 40% in iWUE, integrated WUE based on Δ13C of biomass and cellulose fractions, and WUEplant [15]. Growth rates and biomass accumulation were not significantly different from wild type [15]. Hence, the ABA receptor lines revealed higher water productivity, i.e., WUE per time, both under well-watered growth conditions and under water deficit. Net carbon assimilation was comparable to the wild type, however, at lowered Ci levels and without detectable changes in gm. This report and other studies show that improving WUE is possible without growth trade-offs. The underlying physiological mechanisms are largely unknown and might involve the root system, as grafting experiments have suggested [15], and enzymes of the C4 metabolism, such as PEP carboxylase and its regulatory protein kinase PEPC kinase, which are both upregulated in C3 plants at low CO2 availability [65].
C4 and C3 plants differ in WUE [66,67,68]. At a given gs, C4 plants show higher net carbon assimilation rates and higher WUE [66]. The CO2 concentrating mechanism involving PEP carboxylase results in saturation of C4 photosynthesis at relatively low Ci [69,70]; therefore, lower gs and a steeper CO2 gradient (Ca − Ci) are realized in C4 plants compared to C3 plants [66,70]. C3 plants have Ci values in the range of 300 ± 60 µmol mol−1, while the Ci of the C4 plants is around 150 ± 40 µmol mol−1 [71,72,73] at ambient CO2 of 370–400 µmol mol−1 in well-watered conditions. Under optimal growth conditions, maize and sorghum with C4 metabolism therefore have higher yields per water transpired than the C3 crop wheat [9].
3. Comparative Analysis of Maize and Arabidopsis
The question arises as to whether it is possible to transfer the finding of improved iWUE without having the negative impact on growth to crops. The data on gs-modified tomato plants suggests that it might work for C3 plants [63], but the lower Ci level of C4 plants could preclude such an accomplishment in maize.
To explore the relevance of these findings of uncoupling WUE improvement and yield decreases for the C4 crop maize (Zea mays L.), we analyzed gas exchange data obtained from the maize inbred line B73 and compared them to findings in Arabidopsis. In addition, we analyzed the WUEplant of maize lines subjected to drought. B73 is an inbred line that is commonly used in breeding programs, but is known to be drought-sensitive [74]. B73 was included in a progressive drought stress experiment adapted from Yang et al. [15] in which biomass production with a given amount of water was analyzed and WUEplant was determined. In this experiment, B73 showed the lowest WUEplant (Figure 1a) compared to the maize inbred Mo17 and lines derived from an introgression library described by Gresset et al. [75]. In Figure 1, data are shown of the recurrent parent (RP) of the introgression library as well as two introgression lines differing from RP by reduced (IL-05) or elevated kernel Δ13C (IL-81) [75]. A significantly reduced WUEplant compared with the recurrent parent for IL-05 shows the potential of genetic improvement for this trait. However, the largest difference in WUEplant was observed between B73 and Mo17, with an increase of ~27% (Figure 1a). Our data is in accordance with a previous drought stress experiment conducted on seedlings of maize inbred lines, where Mo17 ranked top in yield per plant [76]. The results indicate genetic variation in the efficiencies of water use among maize lines and a potential for genetic improvement of the WUEplant for B73.
Maize lines showed a difference in water consumption over the five weeks of the experiment (ANOVA, p < 0.001, Figure 1b). However, differences in water consumption cannot explain the differences observed in WUEplant and towards the end of the progressive drought, all genotypes included in the experiment had consumed an equal amount of water (5.8 kg ± 0.02 kg, mean ± SE).
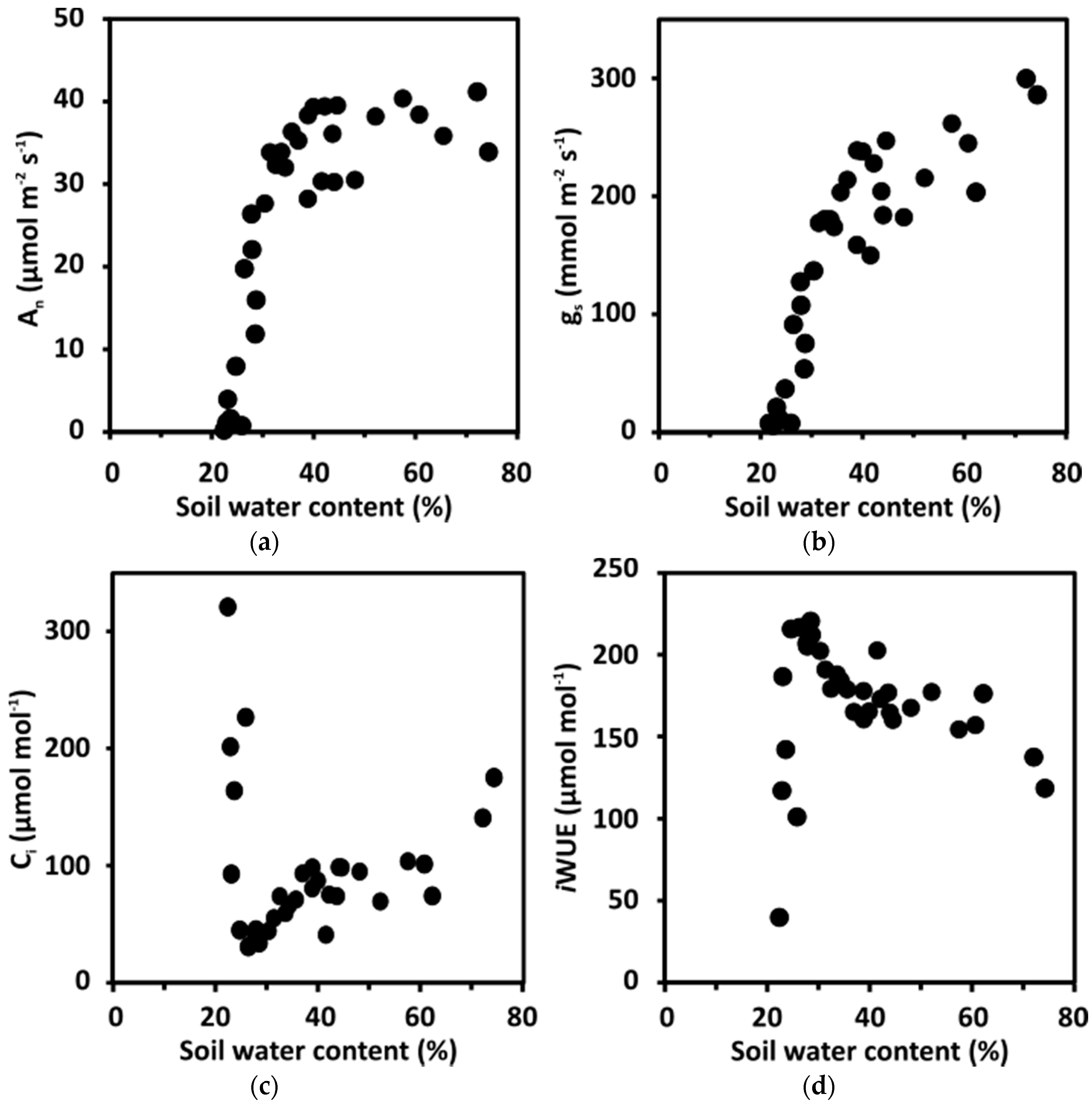
The way in which the change in soil water content (SWC) during the progressive drought experiment affected photosynthesis and iWUE was analyzed by gas exchange measurements. The An of leaves was fairly constant for maize B73 plants exposed to high SWC levels up to 40%, then the An dropped steadily approaching zero at approximately 20% SWC (Figure 2a). In parallel, gs changed moderately between 70% and 40% SWC and declined to zero at 20% SWC (Figure 2b). The Ci values were in the range of 80–100 µmol CO2 per mol between 40–60% SWC. They were somewhat higher in plants from water-saturated soil and were lowered to a minimum of approximately 40 µmol mol−1 at 25% SWC (Figure 2c). Further reduction of the water content in the soil resulted in the steep rise of Ci values, indicating collapsing photosynthesis at very low gs of plants experiencing severe drought stress. As the ambient CO2 concentration (Ca) surrounding the leaf was maintained at 400 µmol mol−1, the CO2 gradient (Ca – Ci) at the stomatal pores increased from approximately 250 µmol mol−1 (Ci of 150 µmol mol−1) at soil water saturation to approximately 360 µmol mol−1 (Ci of 40 µmol mol−1) at the brink of terminal drought. The SWC also influenced iWUE (Figure 2d). Values increased from well-watered conditions to a maximum at 25% SWC, with a plateau around 170 µmol CO2 per mol H2O between 40–60% SWC. Under mild water deficit between 40–60% SWC, there was little variation in An, and gs and, consequently, the iWUE values.
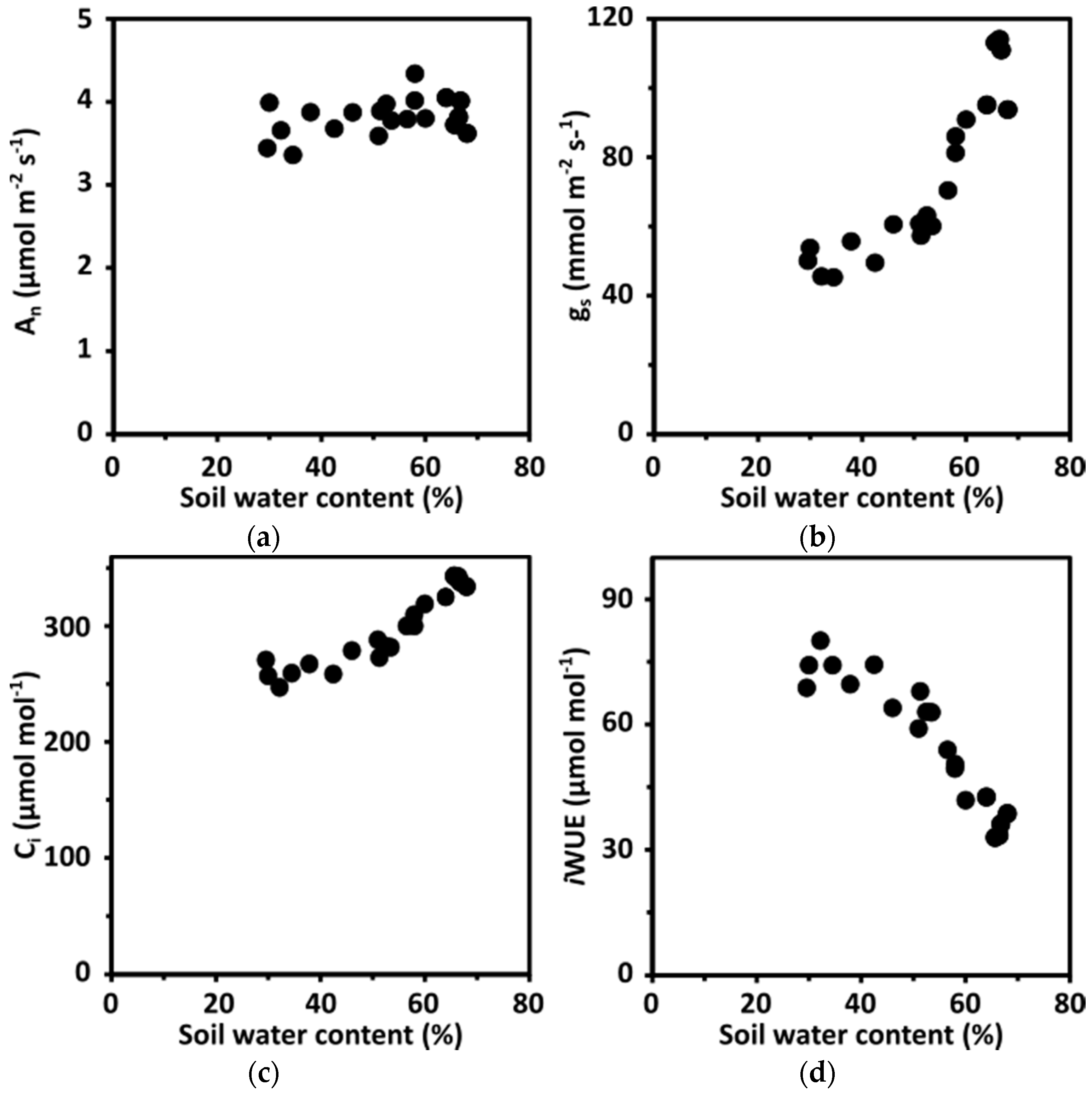
The results for maize B73 differed from data gained by similar analyses of Arabidopsis plants (Figure 3a–d; Reference [15]). The An remained constant between 30–70% SWC, which might be caused by light-limited, but not water-limited, photosynthesis. However, gs and Ci steadily decreased with decreasing SWC and, concomitantly, the iWUE increased by twofold from approximately 35 to 70 µmol mol−1 at 30% SWC. The CO2 gradient at stomata increased more than twofold from approximately 80 µmol mol−1 at soil water saturation to approximately 170 µmol mol−1 at 30%. The data were obtained at light conditions that did not saturate photosynthesis, but analysis at saturating light confirmed the capacity of Arabidopsis to lower Ci and maintain photosynthetic rates unchanged [15]. The improvement in iWUE by limiting gs without major trade-offs in An (Figure 3) was observed for the C3 plants Arabidopsis [15,24] and tomato [63]. A twofold enhancement in iWUE has been reported in different C3 species under drought [11,15,77,78,79]. Besides, considerable differences in WUE in the absence of drought stress have been observed among natural variants [80,81].
Comparing the results from the analysis of maize and Arabidopsis, the potential for increasing iWUE in maize was more limited relative to Arabidopsis. Between 30–60% SWC—reflecting mainly mild water deficit [15]—Arabidopsis responded to the mounting water deficit by a 70% increase in iWUE, while maize showed an increase of less than 20%. Based on the iWUE values obtained at water-saturated soil, the iWUE increase in Arabidopsis and maize was approximately 100% and 40%, respectively, but water logging might be an issue at these high SWC levels. Between 30% and 60% SWC, the stomatal CO2 gradient of maize increased from 300 to 350 µmol mol−1, which corresponded to a 17% increase, whereas in Arabidopsis, the gradient was enhanced by 70%, from 100 to 170 µmol mol−1.
To sum up, mild drought stress (30–60% SWC) had a minor effect on An, gs, and iWUE in maize. However, in Arabidopsis, gs and iWUE changed dynamically, while An was little affected. A reduction in SWC from 35% to 25% led to a rapid decline of An in maize.
These results are in accordance with data from C3 and C4 grass species [77,78] but only partly meet the behavior expected for C3 and C4 plants based on a meta study [82] where decreases in gs and An under mild drought stress were more pronounced in C3 relative to C4 species. Comparing the An and gs curves of maize B73 and Arabidopsis Col-0 (Figure 2a,b and Figure 3a,b), a reduction in stomatal conductance led to an immediate reduction in assimilation rate An for maize but not for Arabidopsis. However, not all C3 plants show the same flat An/gs curve as Arabidopsis [83,84] and therefore the data cannot be translated to C3 crops in general. A previous study on maize lines found differences in gs without trade-offs in An at well-watered conditions [85], and the author noted that Ci values did not become low enough to limit An as it might in high VPD conditions [85] or as it was observed here under drought.
Still, the limitations in improving WUE in maize B73 could be unique to this inbred line, and comparable data from other maize lines is needed before implications can be expanded to maize in general. Additionally, results from maize cannot represent C4 photosynthesis in general because this pathway evolved independently in 19 angiosperm families [67]. However, other C4 grass species have also shown a slight increase in iWUE with progressive depletion of water followed by a steep decline under very severe drought conditions driven by a pronounced decline in An [77,78]. The loss of An in the C4 grass species under drought could partly be attributed to stomatal limitations, while other limitations dominate, including photoinhibition, limitations of CO2 fixation due to desiccation, and decreases in gm [73,77].
It has also been shown that for subspecies of Alloteropsis semialata (R.Br.) Hitchc., An is massively reduced in C4 subspecies under drought in such a way that C4 photosynthesis totally loses its advantage over photosynthesis of the C3 subspecies [73]. This conclusion cannot be drawn from our experiments. However, the observation that C3 plants become more water use efficient during mild and moderate drought while C4 plants show more stable WUE [86] is in accordance with our data on iWUE.
The less potent improvement in WUE observed in maize under drought might be attributable to a limitation in increasing the CO2 gradient (Ca − Ci) further. Maize, like other C4 species, possesses a CO2 concentrating mechanism utilizing precarboxylation of CO2 by PEP carboxylase, which results in Ci values approximately half compared to C3 species [87]. Such a mechanism results in advantages in An and WUE under non- or mild-water-deficit conditions [9,66,69,73,77,78]. However, these advantages cannot be maintained when the drought gets severe, especially for maize and C4 grass species [66,69,73,77,78,86], which is in agreement with our observation for maize B73 at SWC below 27%. The differences can also translate to the field level, where maize has been found to be more sensitive to drought than wheat (C3), with yield reductions of 39.3% compared to 20.6%, respectively, at approximately 40% water reduction [88]. Maize and sorghum are equally or even more sensitive to water stress than many C3 plants [69,73].
Our results show a potential to increase WUE in maize. The inbred lines displayed a broad variation in WUEplant under progressive drought, and iWUE—measured under the same conditions for the least efficient line at whole plant level—still showed a moderate increase in iWUE with declining SWC. However, the potential of WUE improvement is limited in this C4 plant compared to Arabidopsis. This limitation is caused by a very high iWUE and low Ci under well-watered conditions, which provides a minor degree of freedom for further lowering the Ci. The C3 plant is more responsive concerning increases in iWUE under mild water deficit compared to maize. Hence, screening C3 plants for enhanced iWUE in combination with efficient growth is a suitable approach to identify crops with improved WUEplant. This approach is less promising for C4 plants. Establishing higher Ca − Ci gradients in C3 crops at a given soil water potential, e.g., by biotechnical engineering using ABA receptors, has the potential to increase iWUE at the cost of minor reductions in An. Moderate reductions in An do not necessarily influence yield. In barley, improvements in iWUE and WUEplant have been associated with trade-offs in carbon assimilation but without deleterious effects on plant growth or seed yield [89].
4. Conclusions
Our results indicate that the improvement in iWUE without trade-offs in carbon assimilation, as observed for tomato and Arabidopsis [15,63], is less promising for maize and possibly other C4 plants. The large CO2 gradient established by the CO2-concentrating mechanisms of C4 plants limits the potential for further increases in iWUE compared to C3 plants. However, our data show major differences in WUEplant for maize inbred lines and therefore potential for genetic improvement of this trait.
A recent meta-analysis on WUE revealed a tenfold bias in favor of C3 plant studies compared to analyses on C4 plants [86]. We therefore see an urgent need for more studies on C4 crops to shed light on the mechanisms of WUE under water deficit in these important but drought-sensitive crops. Cereals like rice, maize, and wheat contribute largely to global food security [90]; therefore, breeding for and generating water-efficient and high yielding crops are an urgent task to meet future challenges.
Author Contributions
Conceptualization, E.G. and C.-C.S.; Methodology, Z.Y. and V.A.; Formal Analysis, Z.Y., V.A., S.B.; Investigation, Z.Y., V.A., S.B.; Resources, E.G. and C.-C.S.; Data Curation, Z.Y., V.A.; Writing—Original Draft Preparation, S.B.; Writing—Review & Editing, Z.Y., V.A., E.G., C.-C.S.; Visualization, Z.Y., S.B.; Supervision, E.G., C.-C.S.; Project Administration, E.G., C.-C.S.; Funding Acquisition, E.G., C.-C.S.
Funding
This research was funded by the German Research Foundation (Deutsche Forschungsgemeinschaft; DFG) through the Sonderforschungsbereich 924 (SFB924): “Molecular mechanisms regulating yield and yield stability in plants” and the Bavarian State Ministry of the Environment and Consumer Protection within the project network BayKlimaFit (Project TGC01GCUFuE69779).
Acknowledgments
We thank Stefan Schwertfirm and Amalie Fiedler for technical assistance, Anne-Marie Stache for discussion of the statistical analysis, and Farhah Assaad for critical reading and constructive comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO. The Future of Food and Agriculture. Trends and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017. [Google Scholar]
- Spinoni, J.; Naumann, G.; Carrao, H.; Barbosa, P.; Vogt, J. World drought frequency, duration, and severity for 1951–2010. Int. J. Climatol. 2014, 34, 2792–2804. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Dai, A.; van der Schrier, G.; Jones, P.D.; Barichivich, J.; Briffa, K.R.; Sheffield, J. Global warming and changes in drought. Nat. Clim. Chang. 2014, 4, 17–22. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Baker, N.R.; Mullineaux, P.M.; Davies, W.J. Improving water use in crop production. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2008, 363, 639–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobbágy, E.G.; Jackson, R.B. Groundwater use and salinization with grassland afforestation. Glob. Chang. Biol. 2004, 10, 1299–1312. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Spiertz, J.H.J.; Ewert, F. Crop production and resource use to meet the growing demand for food, feed and fuel: Opportunities and constraints. NJAS-Wagen. J. Life Sci. 2009, 56, 281–300. [Google Scholar] [CrossRef]
- Rockström, J.; Lannerstad, M.; Falkenmark, M. Assessing the water challenge of a new green revolution in developing countries. Proc. Natl. Acad. Sci. USA 2007, 104, 6253–6260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadras, V.O.; Grassini, P.; Steduto, P. Status of water use efficiency of main crops. In The State of the World’s Land and Water Resources for Food and Agriculture; F.A.O. Thematic Report No. 7; FAO: Rome, Italy, 2012. [Google Scholar]
- Parry, M.A.J.; Flexas, J.; Medrano, H. Prospects for crop production under drought: Research priorities and future directions. Ann. Appl. Biol. 2005, 147, 211–226. [Google Scholar] [CrossRef]
- Rizza, F.; Ghashghaie, J.; Meyer, S.; Matteu, L.; Mastrangelo, A.M.; Badeck, F.-W. Constitutive differences in water use efficiency between two durum wheat cultivars. Field Crops Res. 2012, 125, 49–60. [Google Scholar] [CrossRef]
- Vadez, V.; Kholova, J.; Medina, S.; Kakkera, A.; Anderberg, H. Transpiration efficiency: New insights into an old story. J. Exp. Bot. 2014, 65, 6141–6153. [Google Scholar] [CrossRef] [PubMed]
- Ellsworth, P.Z.; Cousins, A.B. Carbon isotopes and water use efficiency in C4 plants. Curr. Opin. Plant Biol. 2016, 31, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.C.; Dodd, I.C.; Rothwell, S.A.; Jones, R.; Tardieu, F.; Draye, X.; Davies, W.J. Gravimetric phenotyping of whole plant transpiration responses to atmospheric vapour pressure deficit identifies genotypic variation in water use efficiency. Plant Sci. 2016, 251, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Liu, J.; Tischer, S.V.; Christmann, A.; Windisch, W.; Schnyder, H.; Grill, E. Leveraging abscisic acid receptors for efficient water use in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 6791–6796. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerosa, G.; Mereu, S.; Finco, A.; Marzuoli, R. Stomatal conductance modeling to estimate the evapotranspiration of natural and agricultural ecosystems. In Evapotranspiration: Remote Sensing and Modeling; Irmak, A., Ed.; InTech: Rijeka, Croatia, 2011; pp. 403–420. [Google Scholar]
- Turner, N.C.; Schulze, E.-D.; Gollan, T. The responses of stomata and leaf gas exchange to vapour pressure deficits and soil water content: I. Species comparisons at high soil water contents. Oecologia 1984, 63, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Medrano, H.; Tomás, M.; Martorell, S.; Flexas, J.; Hernández, E.; Rosselló, J.; Pou, A.; Escalona, J.-M.; Bota, J. From leaf to whole-plant water use efficiency (WUE) in complex canopies: Limitations of leaf WUE as a selection target. Crop J. 2015, 3, 220–228. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon Isotope Discrimination and Photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust. J. Plant Physiol. 1984, 11, 539. [Google Scholar] [CrossRef]
- Zhang, C.-Z.; Zhang, J.-B.; Zhao, B.-Z.; Zhang, H.; Huang, P. Stable isotope studies of crop carbon and water relations: A review. Agric. Sci. China 2009, 8, 578–590. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Condon, A.G.; Richards, R.A.; Farquhar, G.D. Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat. Crop Sci. 2002, 42, 739. [Google Scholar] [CrossRef]
- Masle, J.; Gilmore, S.R.; Farquhar, G.D. The ERECTA gene regulates plant transpiration efficiency in Arabidopsis. Nature 2005, 436, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Sheshshayee, M.S.; Bindumadhava, H.; Ramesh, R.; Prasad, T.G.; Lakshminarayana, M.R.; Udayakumar, M. Oxygen isotope enrichment (Δ18O) as a measure of time-averaged transpiration rate. J. Exp. Bot. 2005, 56, 3033–3039. [Google Scholar] [CrossRef] [PubMed]
- Barbour, M.M.; Farquhar, G.D. Relative humidity- and ABA-induced variation in carbon and oxygen isotope ratios of cotton leaves. Plant Cell Environ. 2000, 23, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Barbour, M.M.; Schurr, U.; Henry, B.K.; Wong, S.C.; Farquhar, G.D. Variation in the oxygen isotope ratio of phloem sap sucrose from castor bean. Evidence in support of the Péclet effect. Plant Physiol. 2000, 123, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, G.D.; Lloyd, J. Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere. In Stable Isotopes and Plant Carbon-Water Relations; Ehleringer, J.R., Hall, A.E., Farquhar, G.D., Eds.; Elsevier Science: Burlington, NJ, USA, 1993; pp. 47–70. [Google Scholar]
- Battipaglia, G.; Saurer, M.; Cherubini, P.; Calfapietra, C.; McCarthy, H.R.; Norby, R.J.; Cotrufo, F.M. Elevated CO2 increases tree-level intrinsic water use efficiency: Insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites. New Phytol. 2013, 197, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Schnyder, H.; Cuntz, M.; Keitel, C.; Zeeman, M.J.; Dawson, T.E.; Badeck, F.-W.; Brugnoli, E.; Ghashghaie, J.; Grams, T.E.E.; et al. Progress and challenges in using stable isotopes to trace plant carbon and water relations across scales. Biogeosciences 2012, 9, 3083–3111. [Google Scholar] [CrossRef] [Green Version]
- Von Caemmerer, S.; Ghannoum, O.; Pengelly, J.J.L.; Cousins, A.B. Carbon isotope discrimination as a tool to explore C4 photosynthesis. J. Exp. Bot. 2014, 65, 3459–3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, M.S.; Araus, J.L.; van Heerden, P.D.R.; Foyer, C.H. Enhancing drought tolerance in C4 crops. J. Exp. Bot. 2011, 62, 3135–3153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardieu, F. Any trait or trait-related allele can confer drought tolerance: Just design the right drought scenario. J. Exp. Bot. 2012, 63, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Parent, B.; Caldeira, C.F.; Welcker, C. Genetic and physiological controls of growth under water deficit. Plant Physiol. 2014, 164, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, F.; Simonneau, T.; Muller, B. The physiological basis of drought tolerance in crop plants: A Scenario-Dependent Probabilistic Approach. Annu. Rev. Plant Biol. 2018, 69, 733–759. [Google Scholar] [CrossRef] [PubMed]
- van Oosterom, E.J.; Yang, Z.; Zhang, F.; Deifel, K.S.; Cooper, M.; Messina, C.D.; Hammer, G.L. Hybrid variation for root system efficiency in maize: Potential links to drought adaptation. Funct. Plant Biol. 2016, 43, 502. [Google Scholar] [CrossRef]
- Williams, M.; Woodward, F.I.; Baldocchi, D.D.; Ellsworth, D. CO2 capture from the leaf to the landscape. In Photosynthetic Adaptation: Chloroplast to Landscape; Smith, W.K., Vogelmann, T.C., Critchley, C., Eds.; Springer: Berlin, Germany, 2004; pp. 133–168. [Google Scholar]
- Flexas, J.; Diaz-Espejo, A.; Galmés, J.; Kaldenhoff, R.; Medrano, H.; Ribas-Carbo, M. Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant Cell Environ. 2007, 30, 1284–1298. [Google Scholar] [CrossRef] [PubMed]
- Medrano, H. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Díaz-Espejo, A.; Conesa, M.A.; Coopman, R.E.; Douthe, C.; Gago, J.; Gallé, A.; Galmés, J.; Medrano, H.; Ribas-Carbo, M.; et al. Mesophyll conductance to CO2 and Rubisco as targets for improving intrinsic water use efficiency in C3 plants. Plant Cell Environ. 2016, 39, 965–982. [Google Scholar] [CrossRef] [PubMed]
- Monneveux, P.; Rekika, D.; Acevedo, E.; Merah, O. Effect of drought on leaf gas exchange, carbon isotope discrimination, transpiration efficiency and productivity in field grown durum wheat genotypes. Plant Sci. 2006, 170, 867–872. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Loreto, F.; Cornic, G.; Sharkey, T.D. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol. 2004, 6, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Hassiotou, F.; Ludwig, M.; Renton, M.; Veneklaas, E.J.; Evans, J.R. Influence of leaf dry mass per area, CO2, and irradiance on mesophyll conductance in sclerophylls. J. Exp. Bot. 2009, 60, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Barbour, M.M.; Brendel, O.; Cabrera, H.M.; Carriquí, M.; Díaz-Espejo, A.; Douthe, C.; Dreyer, E.; Ferrio, J.P.; Gago, J.; et al. Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis. Plant Sci. 2012, 193–194, 70–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, T.D. Estimating the rate of photorespiration in leaves. Physiol. Plant 1988, 73, 147–152. [Google Scholar] [CrossRef]
- Flexas, J. Genetic improvement of leaf photosynthesis and intrinsic water use efficiency in C3 plants: Why so much little success? Plant Sci. 2016, 251, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Zhu, X.-G.; Naidu, S.L.; Ort, D.R. Can improvement in photosynthesis increase crop yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, M.A.J.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.-G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.P.; Davison, P.A.; Ruban, A.V.; Horton, P. The xanthophyll cycle pool size controls the kinetics of non-photochemical quenching in Arabidopsis thaliana. FEBS Lett. 2008, 582, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461. [Google Scholar] [CrossRef]
- Perez-Martin, A.; Michelazzo, C.; Torres-Ruiz, J.M.; Flexas, J.; Fernández, J.E.; Sebastiani, L.; Diaz-Espejo, A. Regulation of photosynthesis and stomatal and mesophyll conductance under water stress and recovery in olive trees: Correlation with gene expression of carbonic anhydrase and aquaporins. J. Exp. Bot. 2014, 65, 3143–3156. [Google Scholar] [CrossRef] [PubMed]
- Olsovska, K.; Kovar, M.; Brestic, M.; Zivcak, M.; Slamka, P.; Shao, H.B. Genotypically Identifying Wheat Mesophyll Conductance Regulation under Progressive Drought Stress. Front. Plant Sci. 2016, 7, 1111. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Struik, P.C.; Yin, X.; Yang, J. Stomatal conductance, mesophyll conductance, and transpiration efficiency in relation to leaf anatomy in rice and wheat genotypes under drought. J. Exp. Bot. 2017, 68, 5191–5205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flexas, J.; Ribas-Carbó, M.; Bota, J.; Galmés, J.; Henkle, M.; Martínez-Cañellas, S.; Medrano, H. Decreased Rubisco activity during water stress is not induced by decreased relative water content but related to conditions of low stomatal conductance and chloroplast CO2 concentration. New Phytol. 2006, 172, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Galmés, J.; Medrano, H.; Flexas, J. Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms. New Phytol. 2007, 175, 81–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159. [Google Scholar] [CrossRef]
- Kenney, A.M.; McKay, J.K.; Richards, J.H.; Juenger, T.E. Direct and indirect selection on flowering time, water-use efficiency (WUE, δ (13)C), and WUE plasticity to drought in Arabidopsis thaliana. Ecol. Evolut. 2014, 4, 4505–4521. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Franks, P.J.; Doheny-Adams, T.; Britton-Harper, Z.J.; Gray, J.E. Increasing water-use efficiency directly through genetic manipulation of stomatal density. New Phytol. 2015, 207, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, A.J.; Andrews, J.; Mulholland, B.J.; McKee, J.M.T.; Hilton, H.W.; Horridge, J.S.; Farquhar, G.D.; Smeeton, R.C.; Smillie, I.R.A.; Black, C.R.; et al. Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol. 2007, 143, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Yoo, C.Y.; Pence, H.E.; Jin, J.B.; Miura, K.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. The Arabidopsis GTL1 transcription factor regulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1. Plant Cell 2010, 22, 4128–4141. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Haq, N.U.; Zhang, H.; Zhu, X.-G. Was low CO2 a driving force of C4 evolution: Arabidopsis responses to long-term low CO2 stress. J. Exp. Bot. 2014, 65, 3657–3667. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P. Environmental responses. In C4 Plant Biology; Sage, R.F., Monson, R.K., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 215–242. [Google Scholar]
- Sage, R.F. The evolution of C4 photosynthesis. New Phytol. 2004, 161, 341–370. [Google Scholar] [CrossRef] [Green Version]
- Downes, R.W. Differences in transpiration rates between tropical and temperate grasses under controlled conditions. Planta 1969, 88, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, O. C4 photosynthesis and water stress. Ann. Bot. 2009, 103, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Ku, M.; Edwards, G.E. C4 Photosynthesis: The CO2-concentrating mechanism and photorespiration. Plant Physiol. 1993, 103, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.C.; Cowan, I.R.; Farquhar, G.D. Stomatal conductance correlates with photosynthetic capacity. Nature 1979, 282, 424–426. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Uribelarrea, M.; Ainsworth, E.A.; Naidu, S.L.; Rogers, A.; Ort, D.R.; Long, S.P. Photosynthesis, productivity, and yield of maize are not affected by open-air elevation of CO2 concentration in the absence of drought. Plant Physiol. 2006, 140, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Ripley, B.S.; Gilbert, M.E.; Ibrahim, D.G.; Osborne, C.P. Drought constraints on C4 photosynthesis: Stomatal and metabolic limitations in C3 and C4 subspecies of Alloteropsis semialata. J. Exp. Bot. 2007, 58, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, W.; Velten, J.; Xin, Z.; Stout, J. Characterization of maize inbred lines for drought and heat tolerance. J. Soil Water Conserv. 2012, 67, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Gresset, S.; Westermeier, P.; Rademacher, S.; Ouzunova, M.; Presterl, T.; Westhoff, P.; Schön, C.-C. Stable carbon isotope discrimination is under genetic control in the C4 species maize with several genomic regions influencing trait expression. Plant Physiol. 2014, 164, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Tahir, M.H.N. Morpho-physiological response of maize inbred lines under drought environment. Asian J. Plant Sci. 2003, 2, 952–954. [Google Scholar] [CrossRef]
- Ripley, B.S.; Frole, K.; Gilbert, M. Differences in drought sensitivities and photosynthetic limitations between co-occurring C3 and C4 (NADP-ME) Panicoid grasses. Ann. Bot. 2010, 105, 493–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.H.; Ripley, B.S.; Woodward, F.I.; Osborne, C.P. Drought limitation of photosynthesis differs between C3 and C4 grass species in a comparative experiment. Plant Cell Environ. 2011, 34, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ranney, T.G.; Bir, R.E.; Skroch, W.A. Comparative drought resistance among six species of birch (Betula): Influence of mild water stress on water relations and leaf gas exchange. Tree Physiol. 1991, 8, 351–360. [Google Scholar] [CrossRef]
- Easlon, H.M.; Nemali, K.S.; Richards, J.H.; Hanson, D.T.; Juenger, T.E.; McKay, J.K. The physiological basis for genetic variation in water use efficiency and carbon isotope composition in Arabidopsis thaliana. Photosyn. Res. 2014, 119, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Des Marais, D.L.; Razzaque, S.; Hernandez, K.M.; Garvin, D.F.; Juenger, T.E. Quantitative trait loci associated with natural diversity in water-use efficiency and response to soil drying in Brachypodium distachyon. Plant Sci. 2016, 251, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Zhong, Y.; Shangguan, Z. A meta-analysis of leaf gas exchange and water status responses to drought. Sci. Rep. 2016, 6, 20917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flexas, J.; Niinemets, U.; Gallé, A.; Barbour, M.M.; Centritto, M.; Diaz-Espejo, A.; Douthe, C.; Galmés, J.; Ribas-Carbo, M.; Rodriguez, P.L.; et al. Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.E.; Schulze, E.-D. Stomatal response to environment and a possible interrelation between stomatal effects on transpiration and CO2 assimilation. Plant Cell Environ. 1980, 3, 467–474. [Google Scholar] [CrossRef]
- Bunce, J.A. Leaf transpiration efficiency of some drought-resistant maize lines. Crop Sci. 2010, 50, 1409. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, H.; Song, X.; Jin, J.; Zhang, X. The responses of plant leaf CO2/H2O exchange and water use efficiency to drought: A meta-analysis. Sustainability 2018, 10, 551. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Hubick, K.T.; Condon, A.G.; Richards, R.A. Carbon isotope fractionation and plant water-use efficiency. In Stable Isotopes in Ecological Research; Billings, W.D., Golley, F., Lange, O.L., Olson, J.S., Remmert, H., Rundel, P.W., Ehleringer, J.R., Nagy, K.A., Eds.; Springer: New York, NY, USA, 1989; pp. 21–40. [Google Scholar]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing stomatal density in barley improves drought tolerance without impacting on yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAO Statistical Yearbook 2013. World Food and Agriculture; FAO: Rome, Italy, 2013. [Google Scholar]
Figure 1.
Water use efficiency and water consumption of maize lines under progressive drought. (a) Water use efficiency (WUEplant) and (b) whole plant water consumption were assessed over the course of a progressive drought stress experiment adapted from Reference [15]. In the greenhouse, 23 maize genotypes were grown in a randomized complete block design, including the two maize inbred lines Mo17 and B73 and introgression lines described by Gresset et al. [75]. The maize inbred line RP and introgression lines derived therefrom (IL-81, IL-05) were kindly provided KWS Saat SE (Einbeck, Germany). Inbred lines B73 and Mo17 were kindly provided by the Chair of Genetics, Technical University of Munich, Freising, Germany. Prior to the experiment, maize seedlings were established in small pots in the growth chamber (16 h day at 25 °C, 650 µE m−2 s−1 photosynthetically active radiation [PAR], 8 h night at 20 °C; 75% relative humidity [RH]) for two weeks after germination under well-watered conditions. Plants of RP harvested at this age had an aboveground dry matter of 0.62 g ± 0.27 g, and plants of an introgression line derived from IL-05 weighed 0.62 g ± 0.26 g. The influence of initial biomass on the biomass at the end of the experiment (28.74 g ± 2.22 g and 25.5 g ± 2.39 g, respectively) was approximately 2%. The plants were transplanted into 10 L pots containing 8 L water-saturated soil (85% v/v soil water content; CL ED73, Einheitserdewerke Patzer, Germany, particle diameter <15 mm). A cover of polyethylene foil was used to prevent evaporation, and the progressive drought experiment was initiated by no further watering. The experiment was conducted in the greenhouse (Gewächshauslaborzentrum Dürnast in Freising, Germany) in Oct–Nov 2017 at full sunlight plus supplemental light at 25–33 °C, 19–20 °C day/night, 400 μmol m−2 s−1 PAR, 40% RH. Soil water content declined progressively during the course of the experiment until the plants used all available water. The water consumed was determined gravimetrically (means ± SE of n ≥ 4 biological replicates). WUEplant was calculated as final aboveground biomass per water consumed (means ± SE of n ≥ 4 biological replicates). The increase in WUE in Mo17 compared to B73 is indicated with an arrow. Student’s paired t-tests of the maize lines were adjusted for multiple comparisons with the Bonferroni method and lines, which did not differ significantly (p < 0.01), and are marked with common letters.
Figure 1.
Water use efficiency and water consumption of maize lines under progressive drought. (a) Water use efficiency (WUEplant) and (b) whole plant water consumption were assessed over the course of a progressive drought stress experiment adapted from Reference [15]. In the greenhouse, 23 maize genotypes were grown in a randomized complete block design, including the two maize inbred lines Mo17 and B73 and introgression lines described by Gresset et al. [75]. The maize inbred line RP and introgression lines derived therefrom (IL-81, IL-05) were kindly provided KWS Saat SE (Einbeck, Germany). Inbred lines B73 and Mo17 were kindly provided by the Chair of Genetics, Technical University of Munich, Freising, Germany. Prior to the experiment, maize seedlings were established in small pots in the growth chamber (16 h day at 25 °C, 650 µE m−2 s−1 photosynthetically active radiation [PAR], 8 h night at 20 °C; 75% relative humidity [RH]) for two weeks after germination under well-watered conditions. Plants of RP harvested at this age had an aboveground dry matter of 0.62 g ± 0.27 g, and plants of an introgression line derived from IL-05 weighed 0.62 g ± 0.26 g. The influence of initial biomass on the biomass at the end of the experiment (28.74 g ± 2.22 g and 25.5 g ± 2.39 g, respectively) was approximately 2%. The plants were transplanted into 10 L pots containing 8 L water-saturated soil (85% v/v soil water content; CL ED73, Einheitserdewerke Patzer, Germany, particle diameter <15 mm). A cover of polyethylene foil was used to prevent evaporation, and the progressive drought experiment was initiated by no further watering. The experiment was conducted in the greenhouse (Gewächshauslaborzentrum Dürnast in Freising, Germany) in Oct–Nov 2017 at full sunlight plus supplemental light at 25–33 °C, 19–20 °C day/night, 400 μmol m−2 s−1 PAR, 40% RH. Soil water content declined progressively during the course of the experiment until the plants used all available water. The water consumed was determined gravimetrically (means ± SE of n ≥ 4 biological replicates). WUEplant was calculated as final aboveground biomass per water consumed (means ± SE of n ≥ 4 biological replicates). The increase in WUE in Mo17 compared to B73 is indicated with an arrow. Student’s paired t-tests of the maize lines were adjusted for multiple comparisons with the Bonferroni method and lines, which did not differ significantly (p < 0.01), and are marked with common letters.
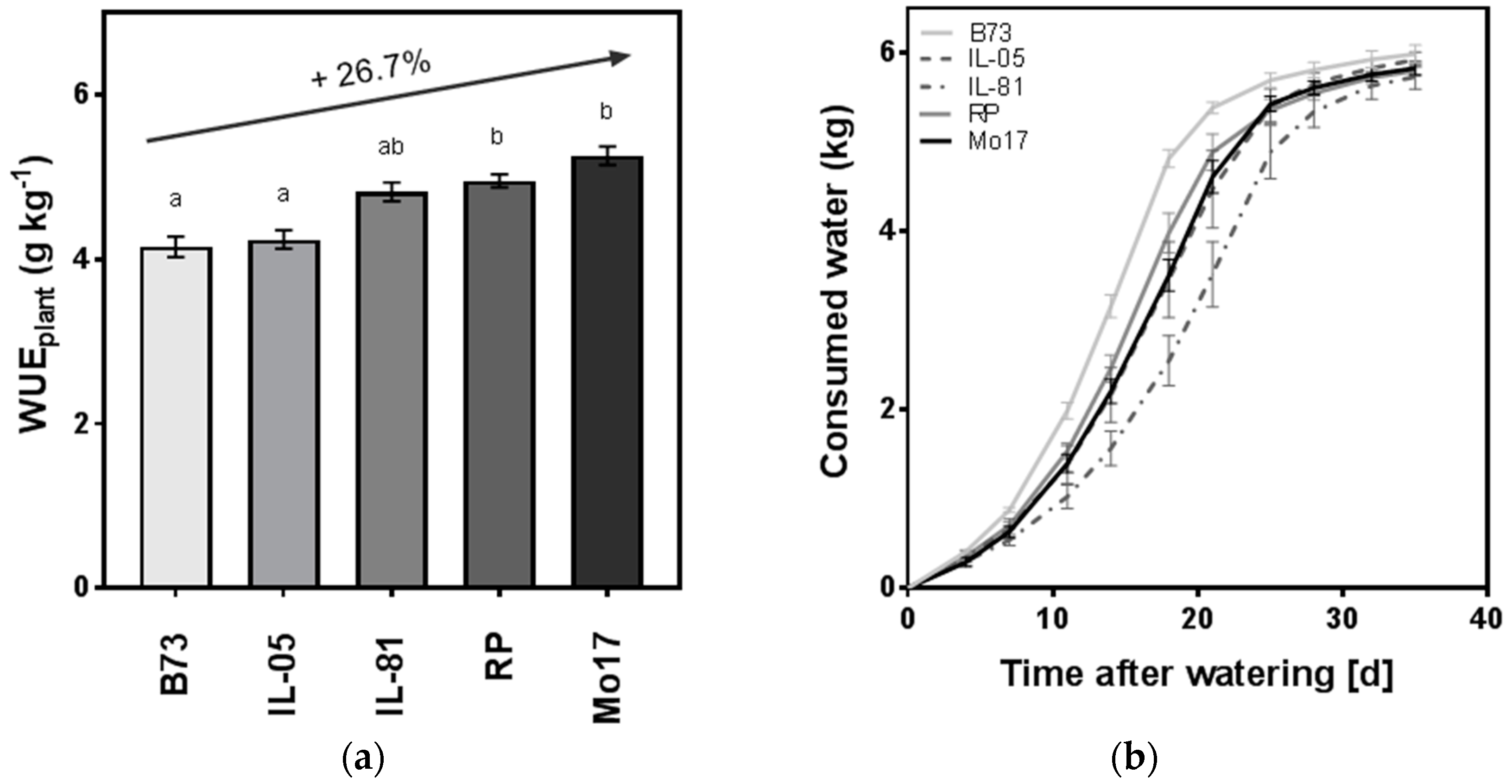
Figure 2.
Changes in intrinsic water use efficiency of maize exposed to a progressive depletion of soil water content. (a) Net carbon assimilation rate (An), (b) stomatal conductance (gs), (c) intercellular CO2 concentration (Ci), and (d) intrinsic WUE (iWUE; defined as the ratio of An to gs) of B73 plants at different soil water content. Gas exchange measurements using the GFS-3000 gas exchange system (Heinz, Walz GmbH, Effeltrich, Germany) were conducted at a photon flux density of 1000 μmol m−2 s−1, an external CO2 (Ca) of 400 μmol mol−1 CO2, and vapor pressure deficit (VPD) of 26 Pa kPa−1 ± 2 Pa kPa−1. The first fully expanded leaf counting from the top of the plants was clamped into an 8 cm2 cuvette for measurements, and plants were subjected to progressive drought as detailed in Figure 1. Plants were grown in soil (Classic Profi Substrate Einheitserde Werkverband) as described in Reference [15]. The experiment was conducted in a greenhouse in the Department of Botany in Freising, Germany from June to August. The maize plants were exposed to full sunlight, at an average temperature of 27 °C, and an average relative humidity of 55% in the experimental period. (a–d) five biological replicates and each data point represents single measurements with five technical replicates.
Figure 2.
Changes in intrinsic water use efficiency of maize exposed to a progressive depletion of soil water content. (a) Net carbon assimilation rate (An), (b) stomatal conductance (gs), (c) intercellular CO2 concentration (Ci), and (d) intrinsic WUE (iWUE; defined as the ratio of An to gs) of B73 plants at different soil water content. Gas exchange measurements using the GFS-3000 gas exchange system (Heinz, Walz GmbH, Effeltrich, Germany) were conducted at a photon flux density of 1000 μmol m−2 s−1, an external CO2 (Ca) of 400 μmol mol−1 CO2, and vapor pressure deficit (VPD) of 26 Pa kPa−1 ± 2 Pa kPa−1. The first fully expanded leaf counting from the top of the plants was clamped into an 8 cm2 cuvette for measurements, and plants were subjected to progressive drought as detailed in Figure 1. Plants were grown in soil (Classic Profi Substrate Einheitserde Werkverband) as described in Reference [15]. The experiment was conducted in a greenhouse in the Department of Botany in Freising, Germany from June to August. The maize plants were exposed to full sunlight, at an average temperature of 27 °C, and an average relative humidity of 55% in the experimental period. (a–d) five biological replicates and each data point represents single measurements with five technical replicates.
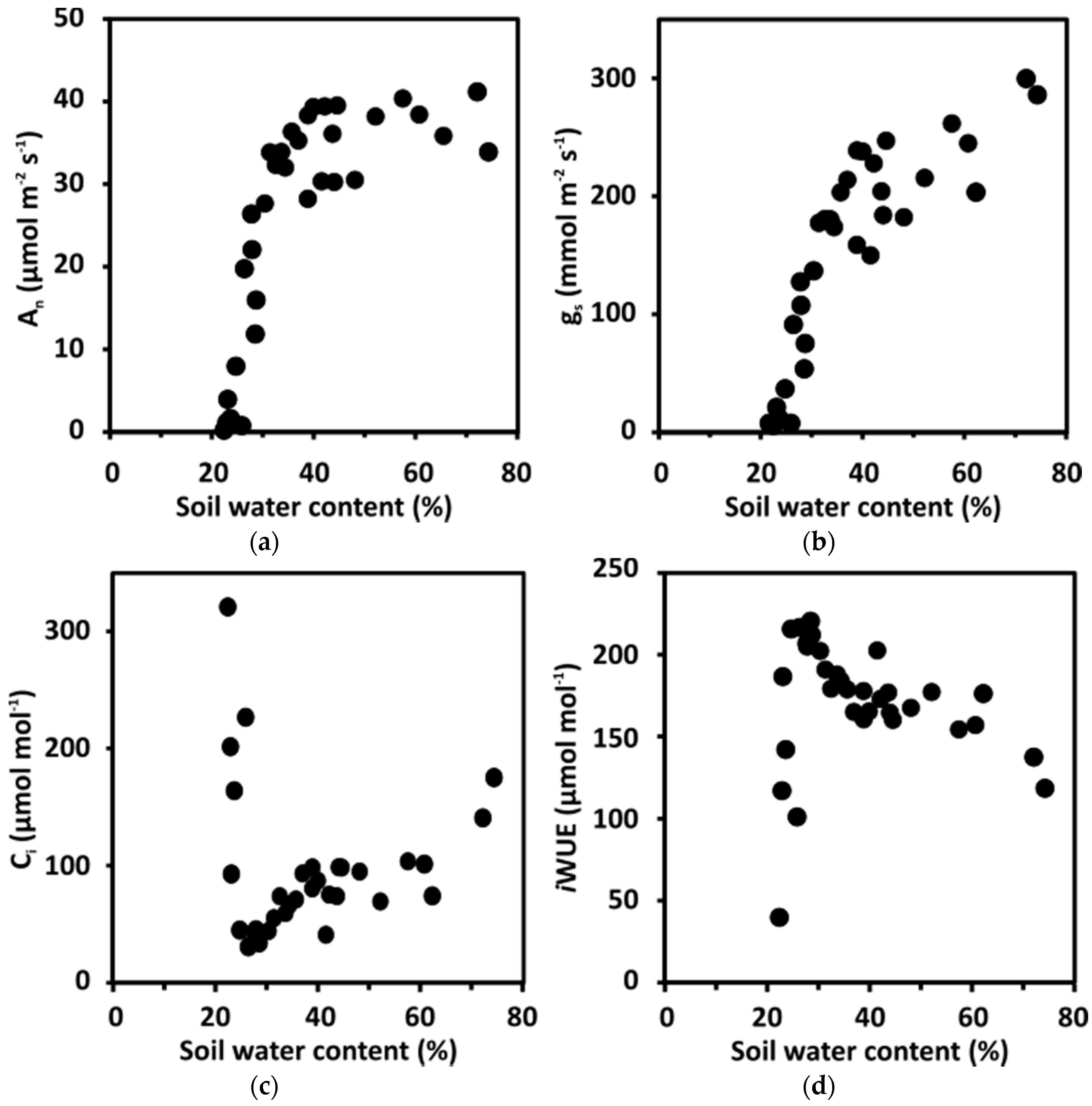
Figure 3.
Changes in intrinsic water use efficiency of Arabidopsis exposed to a progressive depletion of soil water content (SWC). (a) Net carbon assimilation rate (An), (b) stomatal conductance (gs), (c) CO2 concentration in intercellular space (Ci) and (d) intrinsic WUE (iWUE) of whole leaf rosettes of Arabidopsis accession Columbia (Col-0; kindly provided by the Nottingham Arabidopsis Stock Center, Nottingham, UK). The measurements were performed with the device mentioned in Figure 2 at a photon flux density of 150 μmol m−2 s−1, an ambient CO2 (Ca) level of 420 μmol mol−1 and vapor pressure deficit of 13 ± 1 Pa kPa−1. The plants were grown under short day conditions (8 h light/16 h dark photoperiod) at a photon flux density of 150 μmol m−2 s−1 and 22 °C and 50% relative humidity in the day time and 17 °C and 60% relative humidity at night. Data presented in (a–d) consists of three biological replicates and single measurements for each data point consist of 10 technical replicates. Data and the correlation between SWC and water potentials are presented in Reference [15].
Figure 3.
Changes in intrinsic water use efficiency of Arabidopsis exposed to a progressive depletion of soil water content (SWC). (a) Net carbon assimilation rate (An), (b) stomatal conductance (gs), (c) CO2 concentration in intercellular space (Ci) and (d) intrinsic WUE (iWUE) of whole leaf rosettes of Arabidopsis accession Columbia (Col-0; kindly provided by the Nottingham Arabidopsis Stock Center, Nottingham, UK). The measurements were performed with the device mentioned in Figure 2 at a photon flux density of 150 μmol m−2 s−1, an ambient CO2 (Ca) level of 420 μmol mol−1 and vapor pressure deficit of 13 ± 1 Pa kPa−1. The plants were grown under short day conditions (8 h light/16 h dark photoperiod) at a photon flux density of 150 μmol m−2 s−1 and 22 °C and 50% relative humidity in the day time and 17 °C and 60% relative humidity at night. Data presented in (a–d) consists of three biological replicates and single measurements for each data point consist of 10 technical replicates. Data and the correlation between SWC and water potentials are presented in Reference [15].

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Blankenagel, S.; Yang, Z.; Avramova, V.; Schön, C.-C.; Grill, E. Generating Plants with Improved Water Use Efficiency. Agronomy 2018, 8, 194. https://doi.org/10.3390/agronomy8090194
AMA Style
Blankenagel S, Yang Z, Avramova V, Schön C-C, Grill E. Generating Plants with Improved Water Use Efficiency. Agronomy. 2018; 8(9):194. https://doi.org/10.3390/agronomy8090194
Chicago/Turabian StyleBlankenagel, Sonja, Zhenyu Yang, Viktoriya Avramova, Chris-Carolin Schön, and Erwin Grill. 2018. "Generating Plants with Improved Water Use Efficiency" Agronomy 8, no. 9: 194. https://doi.org/10.3390/agronomy8090194
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.