Climate Change, Carbon Dioxide, and Pest Biology, Managing the Future: Coffee as a Case Study

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Overview of Climate and CO2 Effects on Pest Biology
2.1. Insects
2.2. Plant Pathogens
2.3. Weeds
3. Coffee as a Case Study
3.1. Why Coffee?
3.2. Climate Impacts on Current Coffee Production Areas
3.3. Climate Change and Coffee Insect Pests
3.3.1. The Coffee Berry Borer (Hypothenemus hampei (Ferrari)
3.3.2. Coffee Leaf Miner (Leucoptera coffeella Guérin-Mèneville & Perrottet)
3.3.3. Coffee White Stem Borer (Monochamus leoconotus (Pascoe))
3.4. Other Coffee Pests
3.4.1. Root-knot nematode (Meloidogyne incognita (Kofoid & White) Chitwood)
3.4.2. Coffee Leaf Rust (Hemileia vastatrix Berk. & Broome)
4. Pest Management Vulnerabilities
4.1. Rapid Demographic Shifts
4.2. Evolution and Fitness
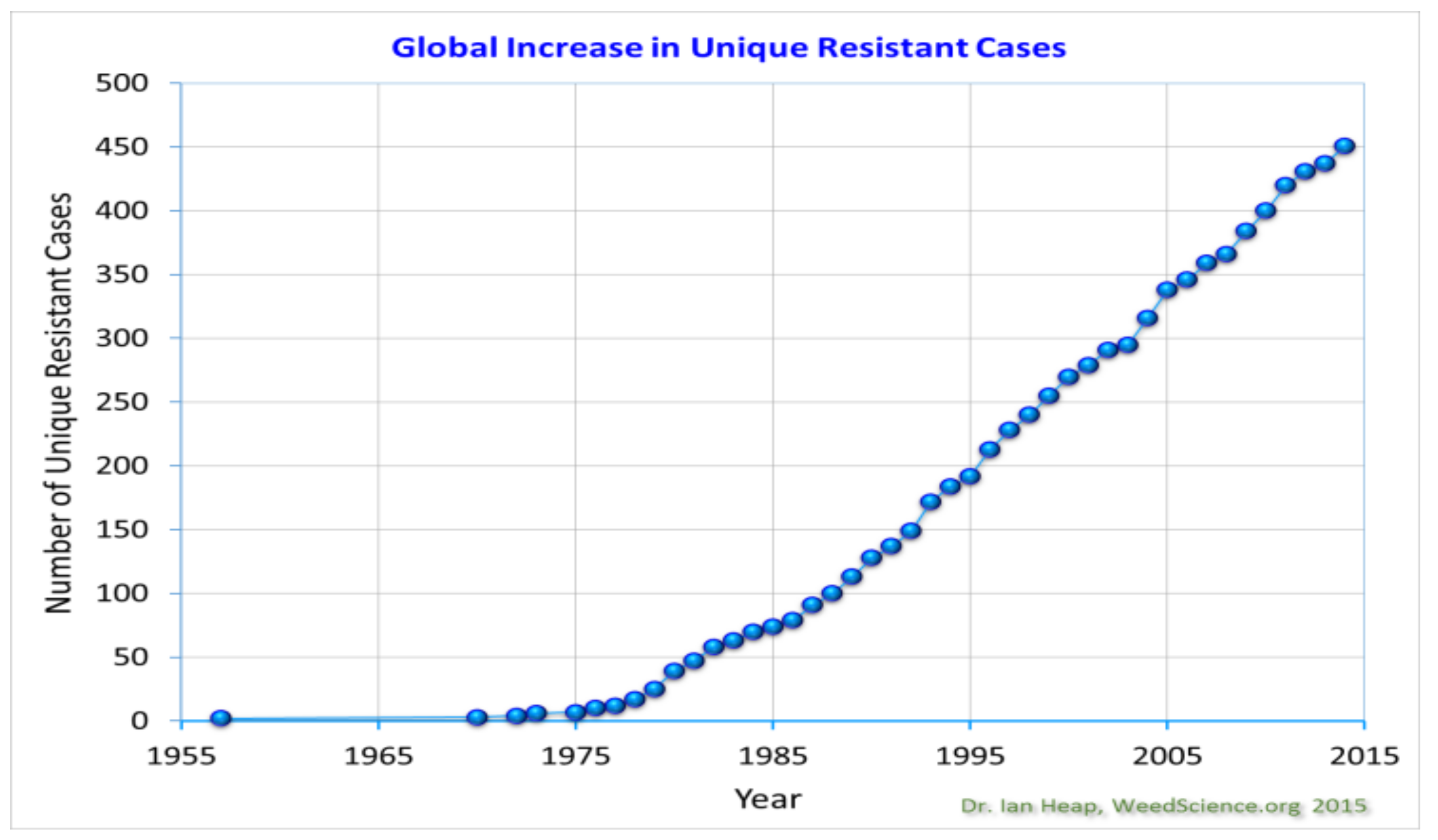
4.3. Pest Management Efficacy
5. How to Reduce Uncertainty in Pest Management: Some Ideas
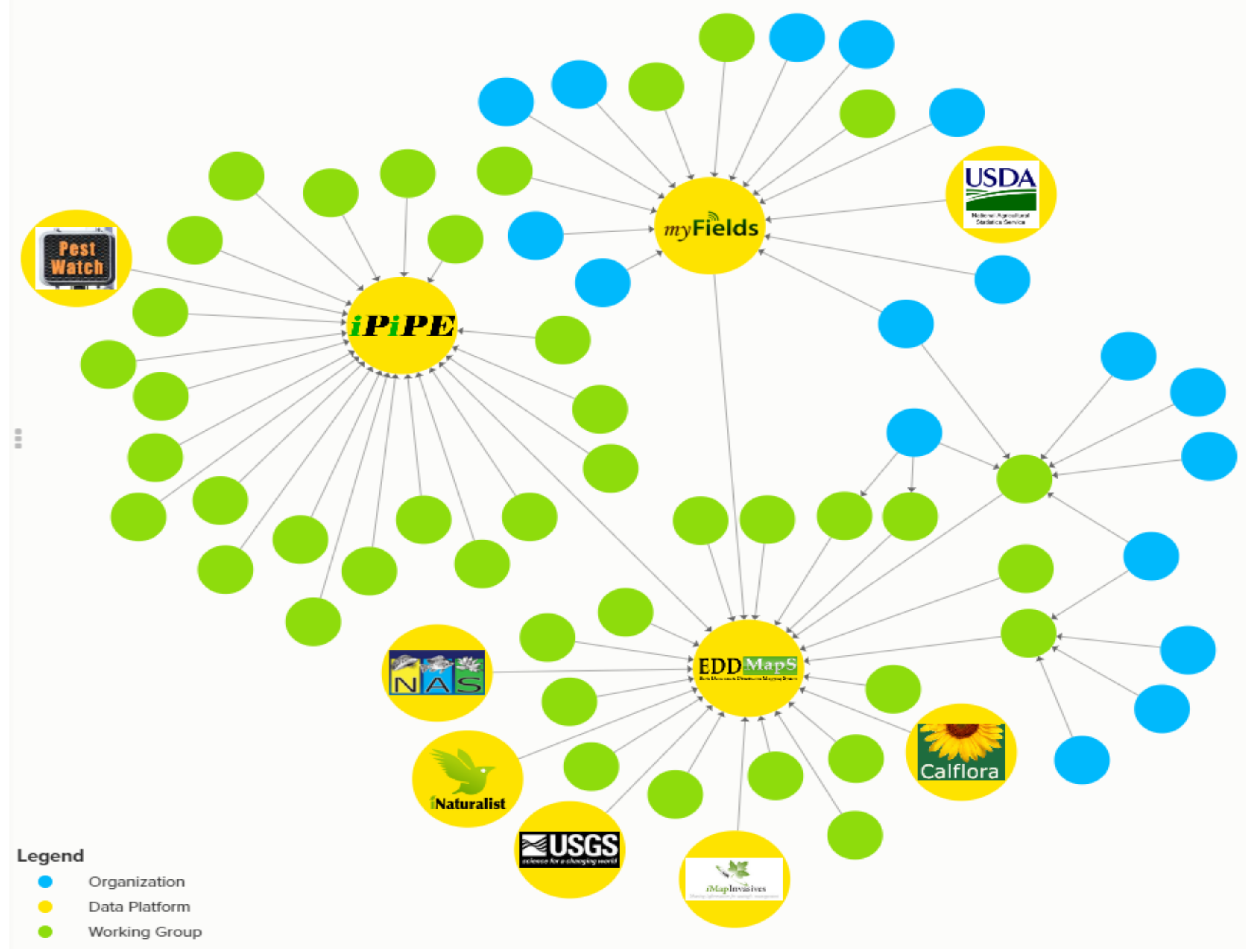
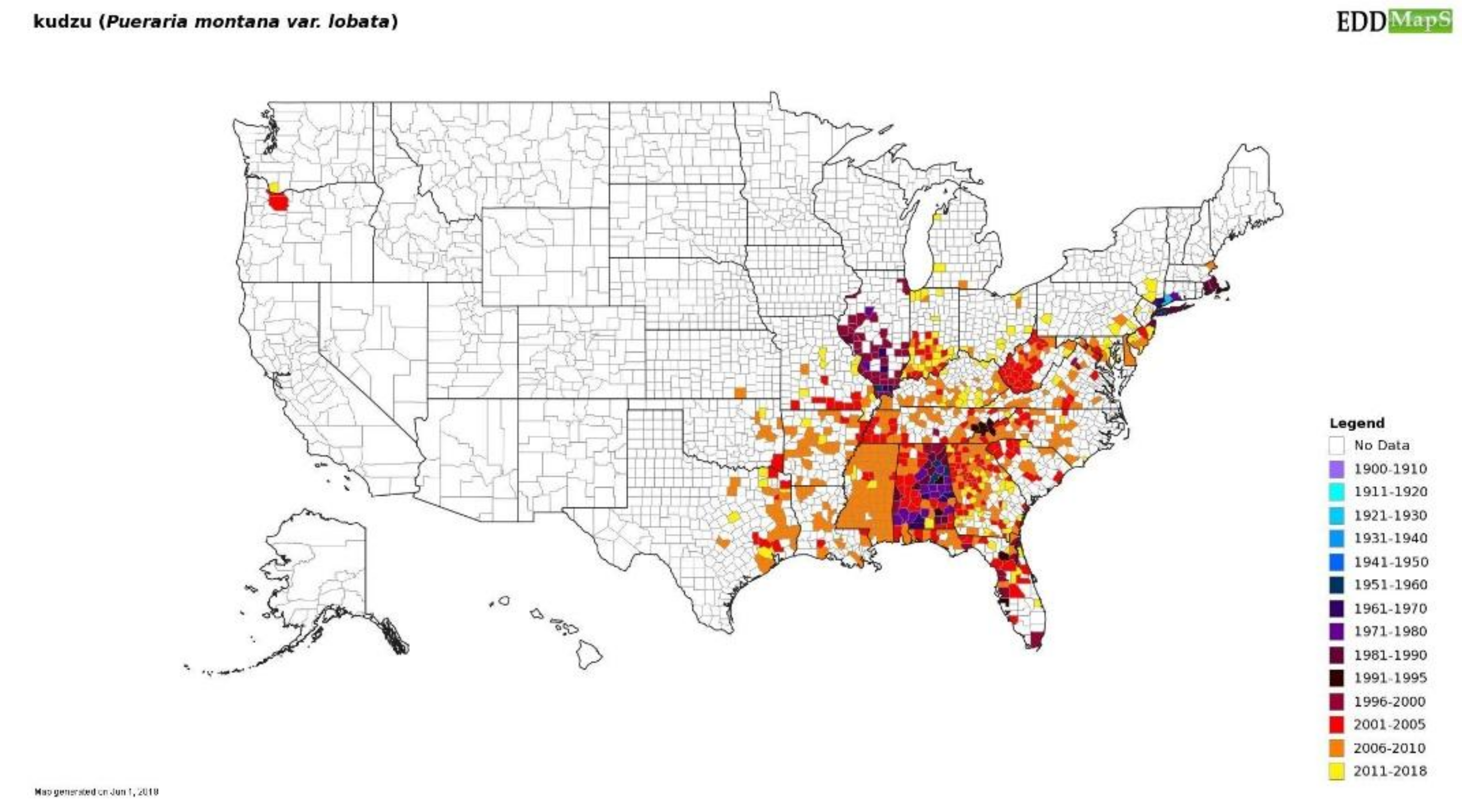
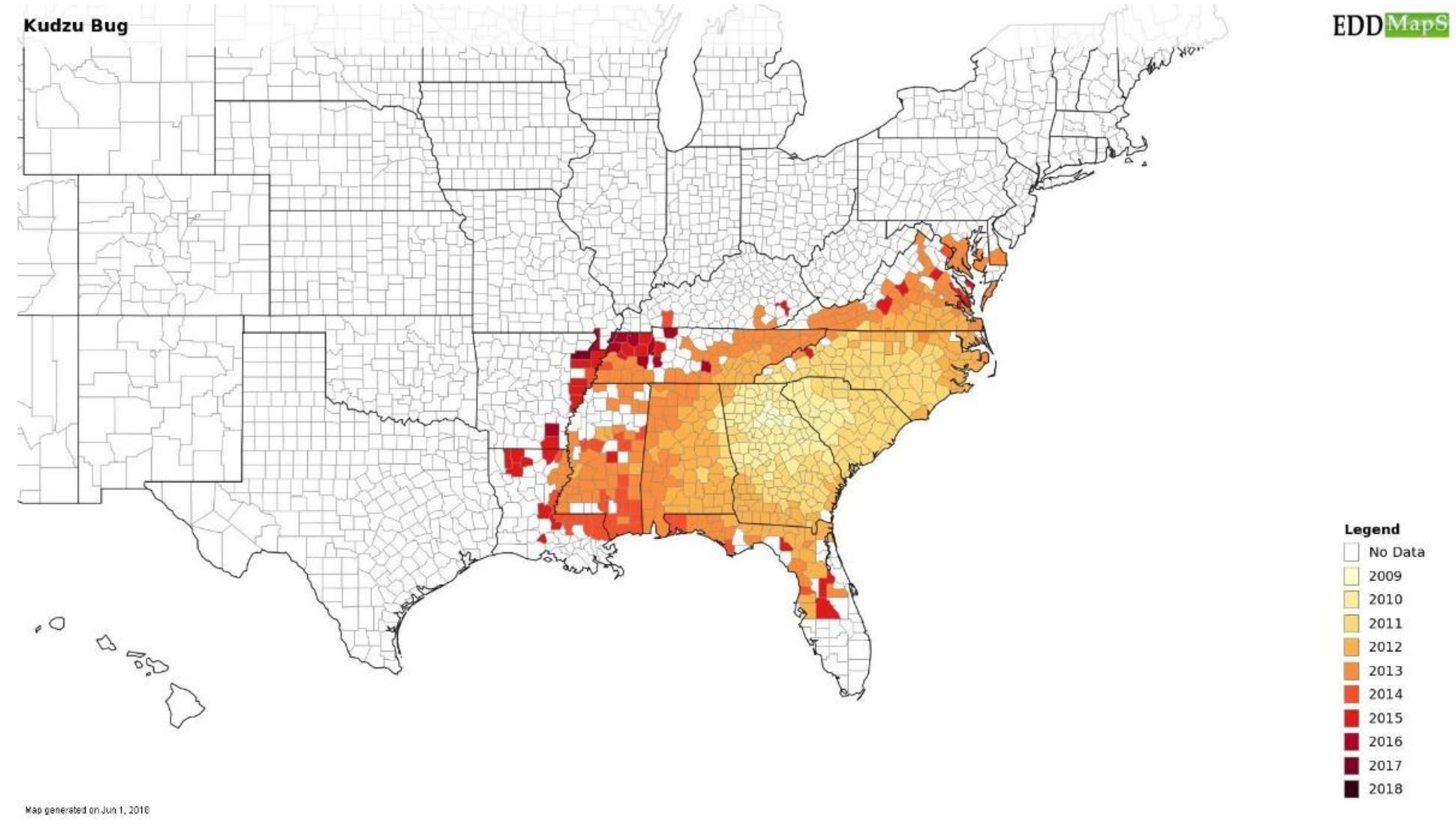
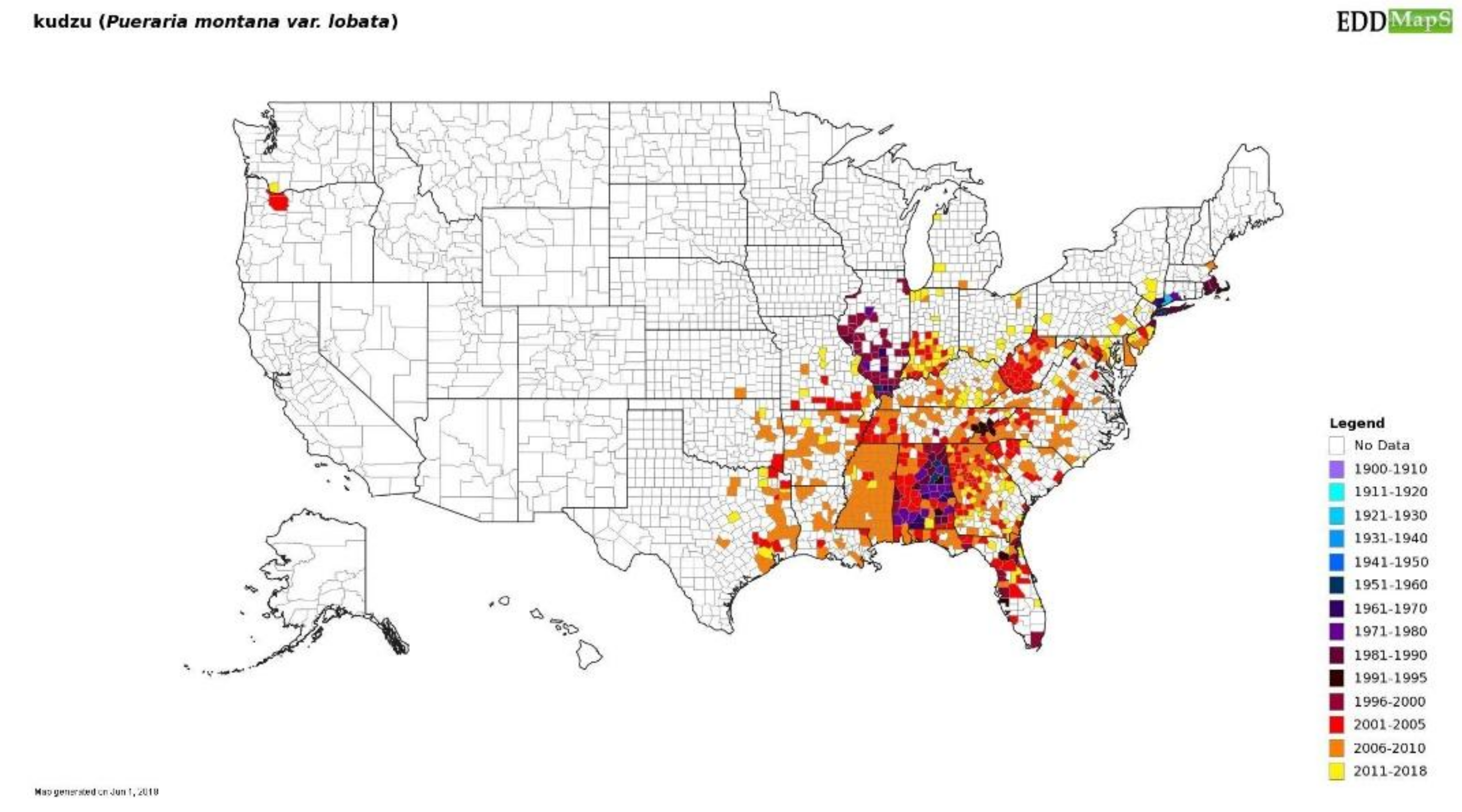
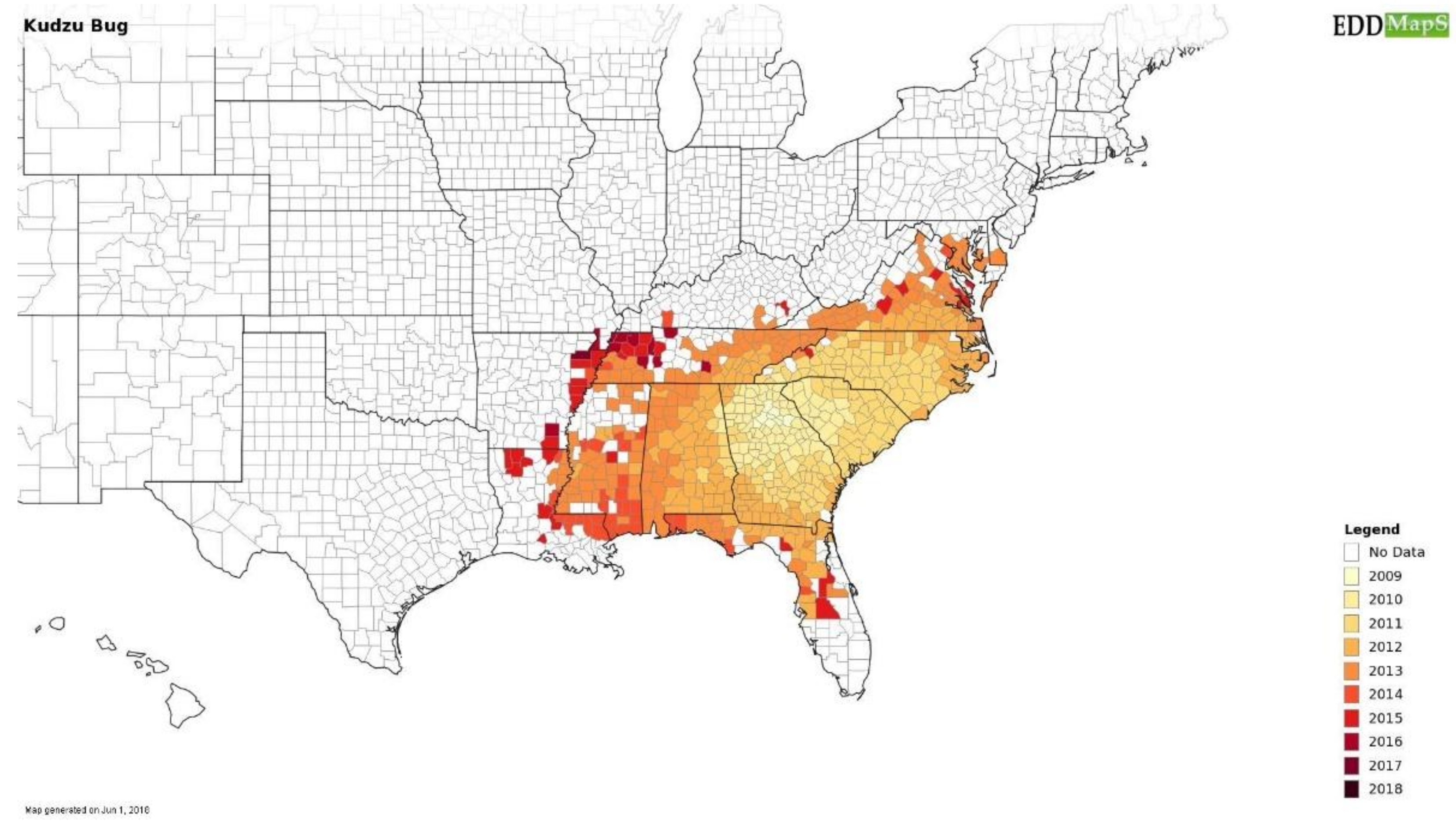
5.1. Increase Monitoring Capacity: EDDMapS
5.2. Chemical Control vs Integrated Pest Management
5.3. Basic Biology
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Booth, B.B.B.; Bernie, D.; McNeall, D.; Hawkins, E.; Caesar, J.; Boulton, C.; Friedlingstein, P.; Sexton, D.M.H. Scenario and modelling uncertainty in global mean temperature change derived from emission-driven global climate models. Earth Syst. Dynam. 2013, 4, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Edenhofer, O.; Pichs-Madruga, R.; Sokona, Y.; Farahani, E.; Kadner, S.; Seyboth, K.; Adler, A.; Baum, I.; Brunner, S.; Eickemeier, P.; et al. (Eds.) Climate change 2014: Mitigation of Climate Change—Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2014; ISBN 978-1-107-65481-5. [Google Scholar]
- Qian, B.; Gregorich, E.G.; Gameda, S.; Hopkins, D.W.; Wang, X.L. Observed soil temperature trends associated with climate change in Canada. J. Geophys. Res. 2011, 116, D02106. [Google Scholar] [CrossRef]
- Dore, M.H. Climate change and changes in global precipitation patterns: What do we know? Environ. Int. 2005, 31, 1167–1181. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Wan, J.; Hao, X. Analysis of diurnal air temperature range change in the continental United States. Weather Clim. Extremes 2014, 4, 86–95. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Iglesias, A.; Yang, X.B.; Epstein, P.R.; Chivian, E. Climate Change and Extreme Weather Events; Implications for Food Production, Plant Diseases, and Pests. Glob. Chang. Hum. Health 2001, 2, 90–104. [Google Scholar] [CrossRef]
- Kimball, B.A.; Mauney, J.R.; Nakayama, F.S.; Idso, S.B. Effects of increasing atmospheric CO2 on vegetation. In CO2 and Biosphere. Advances in vegetation science; Advances in vegetation science; Rozema, J., Lambers, H., Van de Geijn, S.C., Cambridge, M.L., Eds.; Springer: Dordrecht, The Netherlands, 1993; Volume 14, ISBN 978-94-010-4791-3. [Google Scholar]
- Poorter, H.; Navas, M.L. Plant growth and competition at elevated CO2: On winners, losers and functional groups. New Phytol. 2003, 157, 175–198. [Google Scholar] [CrossRef]
- Kimball, B.A. Crop responses to elevated CO2 and interactions with H2O, N, and temperature. Curr. Opin. Plant Biol. 2016, 31, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooi, C.J.; Reich, M.; Löw, M.; De Kok, L.J.; Tausz, M. Growth and yield stimulation under elevated CO2 and drought: A meta-analysis on crops. Environ. Exp. Bot. 2016, 122, 150–157. [Google Scholar] [CrossRef]
- Pautasso, M.; Doring, T.F.; Garbelotto, M.; Pellis, L.; Jeger, M.J. Impacts of climate change on plant diseases-opinions and trends. Eur. J. Plant Pathol. 2012, 133, 295–313. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.D.; Grey, T.L.; Webster, T.M.; Vencill, W.K. Increased purple nutsedge (Cyperus rotundus) tuber sprouting with diurnally fluctuating temperatures. Weed Sci. 2013, 61, 126–130. [Google Scholar] [CrossRef]
- Bebber, D.P. Range-expanding pests and pathogens in a warming world. Annu. Rev. Phytopathol. 2015, 53, 335–356. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.J.; Herms, D.A.; Cardina, J.; Moore, R.H. Climate change and pest management: Unanticipated consequences of trophic dislocation. Agronomy 2018, 8, 7. [Google Scholar] [CrossRef]
- Zavala, J.A.; Gog, L.; Giacometti, R. Anthropogenic increase in carbon dioxide modifies plant-insect interactions. Ann. Appl. Biol. 2017, 170, 68–77. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Bjorkman, C.; Niemela, P. (Eds.) Climate Change and Insect Pests; CABI Climate Change Series 7; CAB International: Wallingford, UK, 2015; ISBN 978-1-78064-378-6. [Google Scholar]
- Zhou, X.; Harrington, R.; Woiwod, I.P.; Perry, J.N.; Bale, J.S.; Clark, S.J. Effects of temperature on aphid phenology. Glob. Chang. Biol. 1995, 1, 303–313. [Google Scholar] [CrossRef]
- Chen, S.; Fleischer, S.J.; Saunders, M.C.; Thomas, M.B. The influence of diurnal temperature variation on degree-day accumulation and insect life history. PLoS ONE 2015, 10, e0120772. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, K.E.; Karl, T.R.; Brooks, H.; Kossin, J.; Lawrimore, J.H.; Arndt, D.; Emanuel, K. Monitoring and understanding trends in extreme storms: State of knowledge. Bull. Am. Meteorol. Soc. 2013, 94, 499–514. [Google Scholar] [CrossRef]
- Pasquarella, V.J.; Bradley, B.A.; Woodcock, C.E. Near-real-time monitoring of insect defoliation using Landsat time series. Forests 2017, 8, 275. [Google Scholar] [CrossRef]
- Taub, D.R.; Miller, B.; Allen, H. Effects of elevated CO2 on the protein concentration of food crops: A meta-analysis. Glob. Chang. Biol. 2008, 14, 565–575. [Google Scholar] [CrossRef]
- Zavala, J.A.; Casteel, C.L.; DeLucia, E.H.; Berenbaum, M.R. Anthropogenic increase in carbon dioxide compromises plant defense against invasive insects. Proc. Natl. Acad. Sci. USA 2008, 105, 5129–5133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Calonnec, A.; Burie, J.B.; Langlais, M.; Guyader, S.; Saint-Jean, S.; Sache, I.; Tivoli, B. Impacts of plant growth and architecture on pathogen processes and their consequences for epidemic behaviour. Eur. J. Plant Pathol. 2013, 135, 479–497. [Google Scholar] [CrossRef]
- Ziska, L.H.; Runion, G.B. Future weed, pest and disease problems for plants. In Agroecosystems in a Changing Climate; Newton, P.C.D., Carran, R.A., Edwards, G.R., Niklaus, P.A., Eds.; CRC (Chemical Rubber Company) Press: Boca Raton, FL, USA, 2007; pp. 261–287. ISBN 978-0-8493-2088-0. [Google Scholar]
- Patterson, D.T.; Westbrook, J.K.; Joyce, R.J.V.; Lingren, P.D.; Rogasik, J. Weeds, insects, and diseases. Clim. Chang. 1999, 43, 711–727. [Google Scholar] [CrossRef]
- Shakya, S.K.; Goss, E.M.; Dufailt, N.S.; van Bruggen, A.H.C. Potential effects of diurnal temperature oscillations on potato late blight with special reference to climate change. Phytopathology 2015, 105, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.W.; Munkvold, G.P. Associations of planting date, drought stress, and insects with Fusarium ear rot and fumonisin B1 contamination in California maize. Food Addit. Contam. 2010, 27, 591–607. [Google Scholar] [CrossRef] [PubMed]
- Garrett, K.A.; Dendy, S.P.; Frank, E.E.; Rouse, M.N.; Travers, S.E. Climate change effects on plant disease: Genomes to ecosystems. Annu. Rev. Phytopathol. 2006, 44, 489–509. [Google Scholar] [CrossRef] [PubMed]
- Trębicki, P.; Nancarrow, N.; Cole, E.; Bosque-Pérez, N.A.; Constable, F.E.; Freeman, A.J.; Rodoni, B.; Yen, A.L.; Luck, J.E.; Fitzgerald, G.J. Virus disease in wheat predicted to increase with a changing climate. Glob. Chang. Biol. 2015, 21, 3511–3519. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Datta, S. How will plant pathogens adapt to host plant resistance at elevated CO2 under a changing climate? New Phytol. 2003, 159, 733–742. [Google Scholar] [CrossRef]
- Wolf, J.; O’Neill, N.R.; Rogers, C.A.; Muilenberg, M.L.; Ziska, L.H. Elevated atmospheric carbon dioxide concentrations amplify Alternaria alternata sporulation and total antigen production. Environ. Health Perspect. 2010, 118, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Mcelrone, A.J.; Reid, C.D.; Hoye, K.A.; Hart, E.; Jackson, R.B. Elevated CO2 reduces disease incidence and severity of a red maple fungal pathogen via changes in host physiology and leaf chemistry. Glob. Chang. Biol. 2005, 11, 1828–1836. [Google Scholar] [CrossRef]
- Hovmøller, M.S.; Sørensen, C.K.; Walter, S.; Justesen, A.F. Diversity of Puccinia striiformis on cereals and grasses. Annu. Rev. Phytopathol. 2011, 49, 197–217. [Google Scholar] [CrossRef] [PubMed]
- Forcella, F.; Wilson, R.G.; Dekker, J.; Kremer, R.T.; Cardina, J.; Anderson, R.L.; Alm, D.; Renner, K.A.; Harvey, R.G.; Clay, S.; et al. Weed seed bank emergence across the Corn Belt. Weed Sci. 1997, 45, 67–76. [Google Scholar]
- Liu, Y.; Oduor, A.M.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Glob. Chang. Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Bradley, B.A.; Blumenthal, D.M.; Wilcove, D.S.; Ziska, L.H. Predicting plant invasions in an era of global change. Trends Ecol. Evol. 2010, 25, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A.; Ziska, L.H. Crop ecosystem responses to climatic change: Crop/weed interactions. In Climate Change and Global Crop Productivity; Reddy, K.R., Hodges, H.F., Eds.; CABI (Centre for Agriculture and Bioscience International) Publishing: New York, NY, USA, 2000; ISBN 0851994393. [Google Scholar]
- McDonald, A.; Riha, S.; DiTommaso, A.; DeGaetano, A. Climate change and the geography of weed damage: Analysis of US maize systems suggests the potential for significant range transformations. Agric. Ecosyst. Environ. 2009, 130, 131–140. [Google Scholar] [CrossRef]
- Ziska, L.H.; Dukes, J.S. (Eds.) Weed Biology and Climate Change; John Wiley & Sons: New York, NY, USA, 2011; ISBN 978-0-813-81417-9. [Google Scholar]
- Bridges, D.C. Crop Losses Due to Weeds in the United States, 1992; Weed Science Society of America: Champaign, IL, USA, 1992; ISBN 0911733159. [Google Scholar]
- Alberto, A.M.; Ziska, L.H.; Cervancia, C.R.; Manalo, P.A. The influence of increasing carbon dioxide and temperature on competitive interactions between a C3 crop, rice (Oryza sativa) and a C4 weed (Echinochloa glabrescens). Funct. Plant Biol. 1996, 23, 795–802. [Google Scholar] [CrossRef]
- Valerio, M.; Tomecek, M.B.; Lovelli, S.; Ziska, L.H. Quantifying the effect of drought on carbon dioxide-induced changes in competition between a C3 crop (tomato) and a C4 weed (Amaranthus retroflexus). Weed Res. 2011, 51, 591–600. [Google Scholar] [CrossRef]
- Ziska, L.H.; Tomecek, M.B.; Gealy, D.R. Competitive interactions between cultivated and red rice as a function of recent and projected increases in atmospheric carbon dioxide. Agron. J. 2010, 102, 118–123. [Google Scholar] [CrossRef]
- Ziska, L.H.; McClung, A. Differential response of cultivated and weedy (red) rice to recent and projected increases in atmospheric carbon dioxide. Agron. J. 2008, 100, 1259–1263. [Google Scholar] [CrossRef]
- Polley, H.W.; Johnson, H.B.; Mayeux, H.S. Increasing CO2: Comparative responses of the C4 grass Schizachyrium and grassland invader Prosopis. Ecology 1994, 75, 976–988. [Google Scholar] [CrossRef]
- Dukes, J.S.; Chiariello, N.R.; Loarie, S.R.; Field, C.B. Strong response of an invasive plant species (Centaurea solstitialis L.) to global environmental changes. Ecol. Appl. 2011, 21, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.; Breitsameter, L.; Gerowitt, B. Impact of climate change on weeds in agriculture: A review. Agron. Sustain. Dev. 2014, 34, 707–721. [Google Scholar] [CrossRef]
- Läderach, P.; Haggar, J.; Lau, C.; Eitzinger, A.; Ovalle, O.; Baca, M.; Jarvis, A.; Lundy, M. Mesoamerican Coffee: Building a Climate Change Adaptation Strategy. CIAT Policy Brief No. 2; Centro Internacional de Agricultural Tropical (CIAT): Cali, Colombia, 2010; p. 4. [Google Scholar]
- Craparo, A.C.W.; Van Asten, P.J.A.; Läderach, P.; Jassogne, L.T.P.; Grab, S.W. Coffea arabica yields decline in Tanzania due to climate change: Global implications. Agric. For. Meteorol. 2015, 207, 1–10. [Google Scholar] [CrossRef]
- Minten, B.; Tamru, S.; Kuma, T.; Nyarko, Y. Structure and # of Ethiopia’s Coffee Export Sector; Ethiopia Strategy Support Program; Ethiopian Development Research Institute (EDRI): Addis Ababa, Ethiopia; International Food Policy Research Institute (IFPRI): Washington, DC, USA; Volume 66, June 2014; p. 30. [Google Scholar]
- Cheatham, M.R.; Rouse, M.N.; Esker, P.D.; Ignacio, S.; Pradel, W.; Raymundo, R.; Sparks, A.H.; Forbes, G.A.; Gordon, T.R.; et al. Beyond yield: Plant disease in the context of ecosystem services. Phytopathology 2009, 99, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Vandermeer, J.; Perfecto, I.; Philpott, S. Ecological complexity and pest control in organic coffee production: Uncovering an autonomous ecosystem service. BioScience 2010, 60, 527–537. [Google Scholar] [CrossRef]
- FAOSTAT, Statistics Division, (Food and Agriculture Organization of the United Nations): FAO Publication, Rome, Italy. 2018. Available online: http://faostat.fao.org/ (accessed on 26 June 2018).
- Coffee: World Markets and Trade. USDA (United States Department of Agriculture)-FAS (Foreign Agricultural Service): Washington, DC, USA, December 2017.
- ICO. International Coffee Organization. World Coffee Trade (1963–2013): A Review of the Markets, Challenges and opportunities Facing the Sector, ICC 111-5 Rev. 1, French, 24 February 2014; International Coffee Council: London, UK, 3–7 March 2014; 112th Session. 2014, p. 29. Available online: http://www.ico.org/news/icc-111-5-r1e-world-coffee-outlook.pdf (accessed on 26 June 2018).
- Läderach, P.; Eitzinger, A.; Ovalle, O.; Ramírez, J.; Jarvis, A. Climate Change Adaptation and Mitigation in the Kenyan Coffee Sector; Final Report; Centro Internacional de Agricultural Tropical (CIAT): Cali, Colombia, 2010. [Google Scholar]
- Davis, A.P.; Gole, T.W.; Baena, S.; Moat, J. The impact of climate change on indigenous arabica coffee (Coffea arabica): Predicting future trends and identifying priorities. PLoS ONE 2012, 7, e47981. [Google Scholar] [CrossRef] [PubMed]
- Fain, S.J.; Quiñones, M.; Álvarez-Berríos, N.L.; arés-Ramos, I.K.; Gould, W.A. Climate change and coffee: Assessing vulnerability by modeling future climate suitability in the Caribbean island of Puerto Rico. Clim. Chang. 2018, 146, 175–186. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle Rivera, O.; Kirschke, D. A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Clim. Chang. 2015, 129, 89–101. [Google Scholar] [CrossRef]
- Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M.; Schroth, G. Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS ONE 2015, 10, e0124155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjitkar, S.; Sujakhu, N.M.; Merz, J.; Kindt, R.; Xu, J.; Matin, M.A.; Ali, M.; Zomer, R.J. Suitability analysis and projected climate change impact on banana and coffee productions zones in Nepal. PLoS ONE 2016, 11, e0163916. [Google Scholar] [CrossRef] [PubMed]
- Robinet, C.; Roques, A. Direct impacts of recent climate warming on insect populations. Integr. Zool. 2010, 5, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.C. Climate change effects on insects: Implications for crop protection and food security. J. Crop Improv. 2014, 28, 229–259. [Google Scholar] [CrossRef]
- Vega, F.E.; Infante, F.; Johnson, A.J. Chapter 11—The genus Hypothenemus, with emphasis on H. hampei, the coffee berry borer. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: San Diego, CL, USA, 2015; pp. 427–494. ISBN 978-0-12-417156-5. [Google Scholar]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Economic impact of exotic insect pests in Brazilian agriculture. J. Appl. Entomol. 2013, 137, 1–15. [Google Scholar] [CrossRef]
- Jaramillo, J.; Chabi-Olaye, A.; Kamonjo, C.; Jaramillo, A.; Vega, F.E.; Poehling, H.-M.; Borgemeister, C. Thermal tolerance of the coffee berry borer Hypothenemus hampei: Predictions of climate change on a tropical insect pest. PLoS ONE 2009, 4, e6487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magina, F.L.; Makundi, R.H.; Maerere, A.P.; Maro, G.P.; Teri, J.M. Temporal variations in The abundance of three important insect pests of coffee in Kilimanjaro Region, Tanzania. In Proceedings of the 23rd International Conference on Coffee Science, Bali, Indonesia, 3–8 October 2010; pp. 1114–1118. [Google Scholar]
- Jaramillo, J.; Muchugu, E.; Vega, F.E.; Davis, A.; Borgemeister, C.; Chabi-Olaye, A. Some like it hot: The influence and implications of climate change on coffee berry borer (Hypothenemus hampei) and coffee production in East Africa. PLoS ONE 2011, 6, e24528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magrach, A.; Ghazoul, J. Climate and pest-driven geographic shifts in global coffee production: Implications for forest cover, biodiversity and carbon storage. PLoS ONE 2015, 10, e0133071. [Google Scholar] [CrossRef] [PubMed]
- David-Rueda, G.; Constantino, L.M.; Montoya, E.C.; Ortega, M.; Gil, Z.N.; Benavides-Machado, P. Diagnóstico de Leucoptera coffeella (Lepidoptera: Lyonetiidae) y sus parasitoides en el departamento de Antioquia, Colombia. Rev. Colomb. Entomol. 2016, 42, 4–11. [Google Scholar]
- Pereira, E.J.G.; Picanço, M.C.; Bacci, L.; Crespo, A.L.B.; Guedes, R.N.C. Seasonal mortality factors of the coffee leaf miner, Leucoptera coffeella. Bull. Entomol. Res. 2007, 97, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Ghini, R.; Hamada, E.; Pedro, M.J., Jr.; Marengo, J.A.; do Valle Gonçalves, R.R. Risk analysis of climate change on coffee nematodes and leaf miner in Brazil. Pesq. Agropec. Bras. 2008, 43, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Le Pelley, R.H. Pests of Coffee; Longmans, Green and Co., Ltd.: London, UK, 1968. [Google Scholar]
- Waller, J.M.; Bigger, M.; Hillocks, R.J. (Eds.) Coffee Pests, Diseases and Their Management; CABI (Centre for Agriculture and Biosciences International): Wallingford, UK, 2007; ISBN 978-1-84593-129-2. [Google Scholar]
- Kutywayo, D.; Chemura, A.; Kusena, W.; Chidoko, P.; Mahoya, C. The impact of climate change on the potential distribution of agricultural pests: The case of the coffee white stem borer (Monochamus leuconotus P.) in Zimbabwe. PLoS ONE 2013, 8, e73432. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.F.A.; Salgado, S.M.L.; Silva, J.G.P.; Correa, V.R.; Mendonça, J.S.F.; Carneiro, R.M.D.G. Meloidogyne incognita parasitizing coffee plants in southern Minas Gerais, Brazil. Trop. Plant Pathol. 2018, 43, 95–98. [Google Scholar] [CrossRef]
- Barbosa, D.H.S.G.; Vieira, H.D.; Souza, R.M.; Viana, A.P.; Silva, C.P. Field estimates of coffee yield losses and damage threshold by Meloidogyne exigua. Nematol. Bras. 2004, 28, 49–54. [Google Scholar]
- Waller, J.M. Control of coffee diseases. In Coffee: Botany, Biochemistry and Production of Beans and Beverage; Clifford, M.N., Willson, K.C., Eds.; Publishing Company of Westport: Connecticutt, USA, 1985; pp. 219–229. ISBN 0-7099-O787-7. [Google Scholar]
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.J.; Morales, C. The coffee rust crises in Colombia and Central America (2008-2013): Impacts, plausible causes and proposed solutions. Food Sec. 2015, 7, 303–321. [Google Scholar] [CrossRef] [Green Version]
- Bebber, D.P.; Delgado Castillo, A.; Gurr, S.J. Modelling coffee leaf rust risk in Colombia with climate reanalysis data. Phil. Trans. R. Soc. B. 2016, 371, 20150458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghini, R.; Hamada, E.; Pedro, M.J., Jr.; do Valle Gonçalves, R.R. Incubation period of Hemileia vastatrix in coffee plants in Brazil simulated under climate change. Summa Phytopathol. 2011, 37, 85–93. [Google Scholar] [CrossRef]
- Soltani, N.; Dille, J.A.; Burke, I.C.; Everman, W.J.; VanGessel, M.J.; Davis, V.M.; Sikkema, P.H. Potential corn yield losses from weeds in North America. Weed Technol. 2016, 30, 979–984. [Google Scholar] [CrossRef]
- Lesk, C.; Coffel, E.; D’Amato, A.W.; Dodds, K.; Horton, R. Threats to North American forests from southern pine beetle with warming winters. Nat. Clim. Chang. 2017, 7, 713–717. [Google Scholar] [CrossRef]
- Creeden, E.P.; Hicke, J.A.; Buotte, P.C. Climate, weather, and recent mountain pine beetle outbreaks in the western United States. For. Ecol. Manag. 2014, 312, 239–251. [Google Scholar] [CrossRef]
- Coiner, H.A.; Hayhoe, K.; Ziska, L.H.; Van Dorn, J.; Sage, R.F. Tolerance of subzero winter cold in kudzu (Pueraria montana var. lobata). Oecologia 2018, 187, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Langenbach, C.; Schultheiss, H.; Rosenahl, M.; Tresch, N.; Conrath, U.; Goellner, K. Interspecies gene transfer provides soybean resistance to a fungal pathogen. Plant Biotechnol. J. 2016, 14, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Mundt, C.C.; Wallace, L.D.; Allen, T.W.; Hollier, C.A.; Kemerait, R.C.; Sikora, E.J. Initial epidemic area is strongly associated with the yearly extent of soybean rust spread in North America. Biol. Invasions 2013, 15, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Sanatkar, M.R.; Scoglio, C.; Natarajan, B.; Isard, S.; Garrett, K.A. History, epidemic evolution, and model burn-in for a network of annual invasion: Soybean rust. Phytopathology 2015, 105, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Garrett, K.A.; Dobson, A.D.M.; Kroschel, J.; Natarajan, B.; Orlandini, S.; Tonnang, H.E.Z.; Valdivia, C. The effects of climate variability and the color of weather time series on agricultural diseases and pests, and decision-making for their management. Agric. For. Meteorol. 2013, 170, 216–227. [Google Scholar] [CrossRef]
- Hill, J.K.; Griffiths, H.M.; Thomas, C.D. Climate change and evolutionary adaptations at species’ range margins. Annu. Rev. Entomol. 2011, 56, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S. Migrate or evolve: Options for plant pathogens under climate change. Glob. Chang. Biol. 2013, 19, 1985–2000. [Google Scholar] [CrossRef] [PubMed]
- Novy, A.; Flory, S.L.; Hartman, J.M. Evidence for rapid evolution of phenology in an invasive grass. J. Evol. Biol. 2013, 26, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Franks, S.J.; Sim, S.; Weis, A.E. Rapid evolution of flowering time by an annual plant in response to a climate fluctuation. Proc. Nat. Acad. Sci. USA 2007, 104, 1278–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziska, L.H.; Gealy, D.R.; Tomecek, M.B.; Jackson, A.K.; Black, H.L. Recent and projected increases in atmospheric CO2 concentration can enhance gene flow between wild and genetically altered rice (Oryza sativa). PLoS ONE 2012, 7, e37522. [Google Scholar] [CrossRef] [PubMed]
- Garrett, K.A.; Andersen, K.; Bowden, R.L.; Forbes, G.A.; Kulakow, P.A.; Zhou, B. Resistance genes in global crop breeding networks. Phytopathology 2017, 107, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- World Coffee Research. Available online: https://worldcoffeeresearch.org/work/breeding-future/ (accessed on 15 August 2018).
- McErlich, A.F.; Boydston, R.A. Current state of weed management in organic and conventional cropping systems. In Automation: The Future of Weed Control in Cropping Systems; Young, S.L., Pierce, F.J., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 11–32. ISBN 9400775113. [Google Scholar]
- Ziska, L.H. The role of climate change and increasing atmospheric carbon dioxide on weed management: Herbicide efficacy. Agric. Ecosys. Environ. 2016, 231, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Gealy, D.R.; Burgos, N.; Caicedo, A.L.; Gressel, J.; Lawton-Rauh, A.L.; Avila, L.A.; Theisen, G.; Norsworthy, J.; Ferrero, A.; et al. Weedy (red) rice: An emerging constraint to global rice production. Adv. Agron. 2015, 129, 181–228. [Google Scholar] [CrossRef]
- Early Detection & Distribution Mapping Syatem. Available online: www.EDDMapS.org (accessed 15 August 2018).
- Wallace, R.D.; Bargeron, C.T.; LaForest, J.H. Data and knowledge preservation. Earthzine. 2007. Available online: https://earthzine.org/2017/05/22/data-and-knowledge-preservation/ (accessed on 26 June 2018).
- Duke, C.S.; Porter, J.H. The ethics of data sharing and reuse in biology. BioScience 2013, 63, 483–489. [Google Scholar] [CrossRef]
- Van der Eynden, V.; Corti, L.; Woollard, M.; Bishop, L.; Horton, L. Managing and Sharing Data—Best Practices for Researchers, third ed. UK Data Archive, University of Essex: Colchester, UK, May 2011; p. 40. Available online: http://www.data-archive.ac.uk/media/2894/managingsharing.pdf (accessed on 26 June 2018).
- Tenopir, C.; Allard, S.; Douglass, K.; Aydinoglu, A.U.; Wu, L.; Read, E.; Manoff, M.; Frame, M. Data sharing by scientists: Practices and perceptions. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
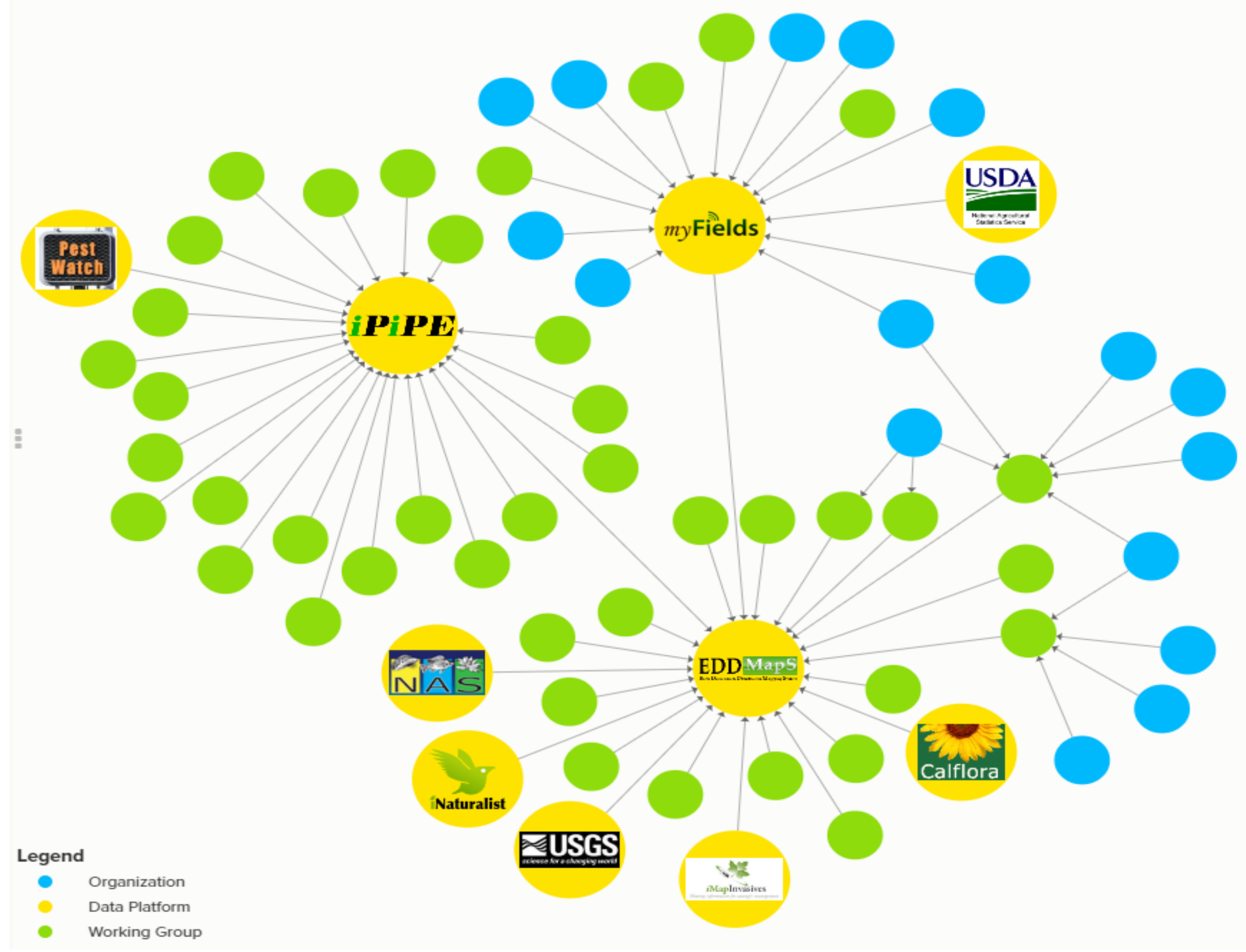
- Isard, S.A.; Russo, J.M.; Magarey, R.D.; Golod, J.; VanKirk, J.R. Integrated Pest Information Platform for Extension and Education (iPiPe): Progress through sharing. J. Integr. Pest Manage. 2015, 6, 15–22. [Google Scholar] [CrossRef]
- Integrated Pest Information Platform for Extension and Education. Available online: http://www.ipipe.org (accessed on15 August 2018).
- Occurrence Data Sharing. Available online: https://kumu.io/IPMC/data-sharing-networks (accessed on 15 August 2018).
- Kudzu, B. Megacopta Working Group. Available online: https://www.kudzubug.org/ (accessed on 26 June 2018).
- Dhammi, A.; van Krestchmar, J.B.; Ponnusamy, L.; Bacheler, J.S.; Reisig, D.D.; Herbert, A.; Del Pozo-Valdivia, A.I.; Roe, R.M. Biology, pest status, microbiome and control of kudzu bug (Hemiptera: Heteroptera: Plataspidae): A new invasive pest in the U.S. Int. J. Mol. Sci. 2016, 17, 1570. [Google Scholar] [CrossRef]
- Thompson, C. Kudzu bugs’ decline is attributed to two factors; CAES (College of Agricultural & Environmental Sciences) News of University of Georgia: Athens, GA, USA, 10 May 2017; Available online: http://www.caes.uga.edu/news/story.html?storyid=6203&story=Kudzu-Bug-Decline (accessed on 26 June 2018).
- Lepczyk, C.A.; Boyle, O.D.; Vargo, T.L.; Gould, P.; Jordan, R.; Liebenberg, L.; Masi, S.; Mueller, W.P.; Pryspy, M.D.; Vaughan, H. Symposium 18: Citizen science in ecology: The intersection of research and education. Bull. Ecol. Soc. Am. 2009, 15, 308–317. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen science as an ecological research tool: Challenges and benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef]
- National Research Council; Division on Engineering and Physical Sciences; Board on Mathematical Sciences and Their Applications; Committee on Applied and Theoretical Statistics; Committee on the Analysis of Massive Data. Massive data in science, technology, commerce, national defense, telecommunications, and other endeavors. In Frontiers in Massive Data Analysis; The National Academies Press: Washington, DC, USA, 2013; pp. 22–40. Available online: http://nap.edu/18374 (accessed on 26 June 2018)ISBN 978-0-309-28778-4.
- Ericsson Mobility Report June 2018. Available online: https://www.ericsson.com/en/mobility-report/reports/june-2018 (accessed 15 August 2018).
- Jurat-Fuentes, J.L.; Gould, F.L.; Adang, M.J. Dual resistance to Bacillus thuringiensis Cry1Ac and Cry2Aa toxins in Heliothis virescens suggests multiple mechanisms of resistance. Appl. Environ. Microbiol. 2003, 69, 5898–5906. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. Herbicide resistant weeds. In Integrated Pest Management; Pimentel, D., Peshin, R., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 3, ISBN 978-94-007-7795-8. [Google Scholar]
- Fernández-Ortuño, D.; Grabke, A.; Bryson, P.K.; Amiri, A.; Peres, N.A.; Schnabel, G. Fungicide resistance profiles in Botrytis cinerea from strawberry fields of seven southern U.S. states. Plant Dis. 2014, 98, 825–833. [Google Scholar] [CrossRef]
- Coll, M.; Wajnberg, E. Environmental pest management: A call to shift from a pest-centric to a system-centric approach. In Environmental Pest Management: Challenges for Agronomists, Ecologists, Economists and Policymakers; Coll, M., Wajnberg, E., Eds.; John Wiley & Sons Ltd.: New Jersey, USA, 2017; pp. 1–17. ISBN 9781119255550. [Google Scholar]
- Culpepper, A.S.; York, A.C.; MacRae, A.W.; Kichler, J. Glyphosate-resistant Palmer amaranth response to weed management programs in Roundup Ready and Liberty Link cotton. In Proceedings of the Beltwide Cotton Conferences, National Cotton Council: Memphis, Tennessee, January 2008; pp. 1689–1690. [Google Scholar]
- Sosnoskie, L.M.; Culpepper, A.S. Glyphosate-resistant palmer amaranth (Amaranthus palmeri) increases herbicide use, tillage, and hand-weeding in Georgia cotton. Weed Sci. 2014, 62, 393–402. [Google Scholar] [CrossRef]
- Livingston, M.; Fernandez-Cornejo, J.; Frisvold, G.B. Economic returns to herbicide resistance management in the short and long run: The role of neighbor effects. Weed Sci. 2016, 64, 595–608. [Google Scholar] [CrossRef]
- Garrett, K.A.; Forbes, G.A.; Savary, S.; Skelsey, P.; Sparks, A.H.; Valdivia, C.; van Bruggen, A.H.C.; Willocquet, L.; Djurle, A.; Duveiller, E.; et al. Complexity in climate-change impacts: An analytical framework for effects mediated by plant disease. Plant Pathol. 2011, 60, 15–30. [Google Scholar] [CrossRef]
- Castex, V.; Beniston, M.; Calanca, P.; Fleury, D.; Moreau, J. Pest management under climate change: The importance of understanding tritrophic relations. Sci. Total Environ. 2018, 616, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Alivisatos, A.P.; Blaser, M.; Brodie, E.L.; Chun, M.; Dangl, J.L.; Donohue, T.J.; Dorrestein, P.C.; Gilbert, J.A.; Green, J.L.; et al. A unified initiative to harness Earth’s microbiomes. Science 2015, 350, 507–508. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Sgrò, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Trelles, F.; Rodríguez, M.A. Rapid micro-evolution and loss of chromosomal diversity in Drosophila in response to climate warming. Evol. Ecol. 1998, 12, 829–838. [Google Scholar] [CrossRef]
- Franks, S.J.; Weis, A.E. A change in climate causes rapid evolution of multiple life-history traits and their interactions in an annual plant. J. Evol. Biol. 2008, 21, 1321–1334. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H. Could recent increases in atmospheric CO2 have acted as a selection factor in Avena fatua populations? A case study of cultivated and wild oat competition. Weed Res. 2017, 57, 399–405. [Google Scholar] [CrossRef]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Heltberg, R.; Siegel, P.B.; Jorgensen, S.L. Addressing human vulnerability to climate change: Toward a ‘no-regrets’ approach. Global Environ. Change 2009, 19, 89–99. [Google Scholar] [CrossRef]
- Lobell, D.B.; Burke, M.B.; Tebaldi, C.; Mastrandrea, M.D.; Falcon, W.P.; Naylor, R.L. Prioritizing climate change adaptation needs for food security in 2030. Science 2018, 319, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.R.; Xie, L.A.; Challinor, A.L.; Cochrane, K.; Howden, D.M.; Iqbal, M.M.; Lobell, D.B.; Travasso, M.I.; Netra, C.; Garrett, K.; et al. Food security and food production systems. In Climate Change 2014: Impacts, Adaptation and Vulnerability—Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Billier, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 485–533. [Google Scholar]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziska, L.H.; Bradley, B.A.; Wallace, R.D.; Bargeron, C.T.; LaForest, J.H.; Choudhury, R.A.; Garrett, K.A.; Vega, F.E. Climate Change, Carbon Dioxide, and Pest Biology, Managing the Future: Coffee as a Case Study. Agronomy 2018, 8, 152. https://doi.org/10.3390/agronomy8080152
Ziska LH, Bradley BA, Wallace RD, Bargeron CT, LaForest JH, Choudhury RA, Garrett KA, Vega FE. Climate Change, Carbon Dioxide, and Pest Biology, Managing the Future: Coffee as a Case Study. Agronomy. 2018; 8(8):152. https://doi.org/10.3390/agronomy8080152
Chicago/Turabian StyleZiska, Lewis H., Bethany A. Bradley, Rebekah D. Wallace, Charles T. Bargeron, Joseph H. LaForest, Robin A. Choudhury, Karen A. Garrett, and Fernando E. Vega. 2018. "Climate Change, Carbon Dioxide, and Pest Biology, Managing the Future: Coffee as a Case Study" Agronomy 8, no. 8: 152. https://doi.org/10.3390/agronomy8080152