Plant Desiccation Tolerance and its Regulation in the Foliage of Resurrection “Flowering-Plant” Species
by
 ,
,
Cecilia K. Blomstedt
1,* ,
,
Cara A. Griffiths
2,
Donald F. Gaff
1,
John D. Hamill
3 and
Alan D. Neale
3 1
School of Biological Sciences, Monash University, Clayton, Victoria 3800, Australia
2
Plant Science, Rothamsted Research, Harpenden, Hertfordshire AL5 2JQ, UK
3
Centre for Regional and Rural Futures, Faculty of Science Engineering & Built Environment, Deakin University, Geelong, Victoria 3217, Australia
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(8), 146; https://doi.org/10.3390/agronomy8080146
Submission received: 29 June 2018
/
Revised: 7 August 2018
/
Accepted: 9 August 2018
/
Published: 14 August 2018
(This article belongs to the Special Issue Gene Expression Regulation during Drought and Salt Stress in Crop Plants)
Abstract
:The majority of flowering-plant species can survive complete air-dryness in their seed and/or pollen. Relatively few species (‘resurrection plants’) express this desiccation tolerance in their foliage. Knowledge of the regulation of desiccation tolerance in resurrection plant foliage is reviewed. Elucidation of the regulatory mechanism in resurrection grasses may lead to identification of genes that can improve stress tolerance and yield of major crop species. Well-hydrated leaves of resurrection plants are desiccation-sensitive and the leaves become desiccation tolerant as they are drying. Such drought-induction of desiccation tolerance involves changes in gene-expression causing extensive changes in the complement of proteins and the transition to a highly-stable quiescent state lasting months to years. These changes in gene-expression are regulated by several interacting phytohormones, of which drought-induced abscisic acid (ABA) is particularly important in some species. Treatment with only ABA induces desiccation tolerance in vegetative tissue of Borya constricta Churchill. and Craterostigma plantagineum Hochstetter. but not in the resurrection grass Sporobolus stapfianus Gandoger. Suppression of drought-induced senescence is also important for survival of drying. Further research is needed on the triggering of the induction of desiccation tolerance, on the transition between phases of protein synthesis and on the role of the phytohormone, strigolactone and other potential xylem-messengers during drying and rehydration.
1. Introduction
The ability of leaves to survive dehydration below 5% relative water content is found in a very small proportion of angiosperm species. In most such ‘resurrection’ species, desiccation tolerance is induced in them in the early stage of drying by the drought stress itself [1]. The occurrence of desiccation tolerance in the foliage of 39 species of grasses (Table 1) is of particular interest in view of the importance of the family Poaceae for agriculture and grazing. (See https://www.youtube.com/watch?v=tGg-_tOiUvA for a summary, including time-lapse video, of resurrection plants reviving).
The mechanisms for implementing desiccation tolerance (constitutive stability, protection, and repair) have often been reviewed [10,11,12,13,14,15]. In addition, stability of extraplastidic lipids is achieved in the resurrection gesneriad Paraisometrum mileense W.T. Wang by maintaining low levels of phospholipase D throughout drying and rehydration [16]. This review focuses on the regulation of drought-induced desiccation tolerance, with particular focus on the resurrection grass S. stapfianus.
Angiosperm species with desiccation-tolerant foliage emerged in largely unrelated families and they do not form a sequence from early-evolved families to late-evolved families. Desiccation-tolerant foliage has not been observed in the large early-evolved angiosperm families, such as Ranunculaceae and Proteaceae, nor in the earlier groups, the cycads and other gymnosperms. Desiccation-tolerant foliage in the angiosperms did not begin in the foliage of early spermatophytes and then progress through continuous lines of related angiosperm families. In the large grass genera Eragrostis and Sporobolus only a small proportion of the species are resurrection grasses. Desiccation-tolerant foliage appears to have originated late in the evolution of these genera and in the family [10].
2. Single Basic Mechanism of Desiccation Tolerance in Seed, Pollen and Resurrection Foliage
The small proportion of species with desiccation-tolerant foliage in the angiosperms contrasts markedly with the 95% of angiosperm species that have desiccation-tolerant seed and the 87% of spermatophyte species that possess desiccation-tolerant pollen [10]. Every angiosperm species in which both seed and pollen have been examined has either desiccation-tolerant seed or desiccation-tolerant pollen. It appears that almost all angiosperm species contain the genetic information for desiccation tolerance, even though it is expressed in the foliage of only a few species. The properties associated with desiccation tolerance are similar in foliage, embryos and pollen [10,17]. This supports the simplest working hypothesis that a single functional module for the mechanism of desiccation tolerance is expressed in the seed and pollen of most angiosperms, and in the foliage of only resurrection species. That is, the appropriate genes are commonly expressed in the reproductive tissues that in each generation are exposed to drying during aerial dispersal-whereas in most angiosperm species, well-developed mechanisms of drought avoidance protect foliage from dehydrating, largely obviating selective pressure for expression in foliage of the seed/pollen mechanism of desiccation tolerance.
3. Predisposing Factors for Evolution of Angiosperm Resurrection Species
Particular angiosperm families appear to have a predisposition for the evolution of foliage-desiccation tolerance. Most resurrection species, for example, occur in the Gesneriaceae, Scrophulariaceae, Velloziaceae and the Poaceae. Some predisposing factors (A–D below) may stem from the importance of shallow soils (often on rock outcrops) in the ecology and evolution of desiccation-tolerant plants. (A) Small seed size (found in all the numerous resurrection angiosperm species inspected by the authors) improves long distance dispersal between rock outcrops. Small assimilate seed-reserves limit the initial depth of root systems, and so plants on shallow soils are liable to drought stress, promoting evolution of desiccation tolerance. (B) Plants have sufficient drought avoidance to colonize shallow soils and to allow sufficient time during drying for drought-induction of desiccation tolerance. (C) For plants with short-stature (linked with small seed size), evolving desiccation tolerance mechanisms may protect them from competition from tall desiccation-sensitive species that succumb to drought on shallow soils. (D) High plasticity in genetic-expression increases scope for drought-driven selection of genes regulating seed/pollen desiccation tolerance to be expressed in the foliage. There is evidence from desiccation-tolerant rotifers that desiccation-prone habitats increase inter-chromosomal transfer of DNA (intra- and inter-species) [18]. Intraspecies proliferation of dehydration-inducible retroelements, such as CDT-1 transcripts in C. plantagineum [19], and conceivably, viral transfer of transposons between species might increase genetic plasticity.
4. Desiccation Tolerance in Plant Taxa That Preceded the Angiosperms
Desiccation tolerance is found in all the extant plant taxa that reflect steps in the evolution of the angiosperms. Although no gymnosperm species is known to have desiccation-tolerant leaves, desiccation-tolerant seed and/or pollen have been reported e.g., in the ‘orthodox seed’ of the early evolved gymnosperm taxa (cycads, Ginkgo), and in the conifers, Gnetum, Ephedra and Welwitschia [20]. The pollen of several conifer species in Pinus and Picea are also desiccation-tolerant [21,22]. The earlier-evolved taxa, lycopods, ferns, bryophytes and green-algae, all have numerous species with vegetative cells and/or spores that are desiccation-tolerant [10].
This distribution pattern suggests that desiccation tolerance was widespread in terrestrial cryptogams with little or no drought avoidance. As survival became less dependent on desiccation tolerance, more complex plant structures and reproductive mechanisms evolved. As further evolution produced stomata, leaf cuticle, water-conducting xylem, cambium and root systems, the effectiveness of drought-avoidance mechanisms increased and desiccation tolerance was needed mainly in the aerially-spread reproductive structures. Thus, the genes producing desiccation tolerance in seed and pollen were carried forward to the angiosperms, but were expressed in the foliage of only the few resurrection plants [10]. In many resurrection angiosperm species, leaf desiccation tolerance is confined to immature leaves and the meristematic base of monocot leaves (Table 2). Evolution of desiccation tolerant leaves appears to commence in young leaves and progresses with further evolution to mature leaves (but, in the authors’ experience, never to senescent leaf tissue), i.e., the expression of genes involved in the regulation of desiccation tolerance is dependent on the developmental stage of the plant.
The main organelles of plant cells are considered to derive from the ‘capture’ of endosymbiotic organisms into the host cell. It is thought that chloroplasts are derived from cyanobacteria [24], mitochondria from bacteria, and that the eukaryote nucleus has origins in the early archaea, with some inflow from bacteria [25,26]. Each of these endosymbionts may have contributed via the derivative organelles to the desiccation tolerance of plants, since desiccation-tolerant species occur in extant archaea (e.g., Methanosarcina barkeri, [27]), bacteria [28,29] and cyanobacteria (see [10]).
5. Regulation of the Induction of Desiccation Tolerance in Angiosperms
Several phytohormones are implicated in the regulation of terrestrial-plant desiccation tolerance (specific hormones are discussed in later sections below). Drought-induction alters transport of some hormones, which are carried in the xylem from the roots to the shoot and may act to signal that the soil is losing moisture content [30]. In detached leaves of eight desiccation-tolerant angiosperm species tested, drought stress did not induce desiccation tolerance suggesting that transported hormone signals may be acting in tolerance-induction in intact plants [31]. Desiccation tolerance, however, appeared to be constitutive in Myrothamnus flabellifolia Welw [31]. Xylem transport is disrupted relatively early during drought stress when the xylem sap cavitates (at water potentials of −0.7 to −3 MPa; [32,33]). Cavitation would curtail xylem-traffic of phytohormones, which are synthesized in the roots-ABA, acetate, CLAVATA3/EMBRYO-surrounding region-related 25 (CLE25) peptide, cytokinins, auxin, strigolactone, and gibberellins. Recent experiments utilizing Arabidopsis suggest that soil water stress elicits a rapid hydraulic response in the shoot that stimulates ABA biosynthesis in the leaves. Hence, the shoot ABA response does not require translocation of ABA from the water stressed roots [34]. Droughted leaves of the resurrection grass S. stapfianus synthesize ABA when they are detached, but ABA contents are higher in leaves that remain attached to droughted plants [35]. Wall associated kinases (WAKs) are transmembrane serine-threonine protein kinases, that have been proposed to act as mechanoreceptors which may link hydraulic signals to physiological responses. Induction of desiccation tolerance first becomes evident at ~60% RWC in S. stapfianus (Table 3), a water content sufficiently low to produce large changes in the hydraulic pressure on the cell contents. A glycine rich protein (GRP) that may interact with WAK kinase is specifically expressed in desiccation tolerant tissue of S. stapfianus [36]. A similar GRP from C. plantagineum has been shown to interact with CpWAK1 [37]. These proteins may be a mechanism for linking hydraulic signals to the desiccation tolerance program. Further water loss decreases transport in the phloem [38], compromising bipolar mass-flow transport of phytohormones in the phloem, including of ABA, azelaic acid, jasmonate, and salicylate [39]. Beyond that point, reactive oxygen species (ROS) movement in the phloem apoplast is feasible, as are also electrical pulses along the phloem cell membrane pathways [39]. Later in drying, vapours of ethylene and of jasmonate may, in theory, still exert effects away from the organs which synthesize them.
The complexity of phytohormones and their transportation raises a potential for development of phased regulation of desiccation tolerance during algal-to-angiosperm evolution. The presence of phases in induction of desiccation tolerance is evidenced by the changes in the complement of leaf proteins observed during drought-induced desiccation tolerance in the resurrection grass S. stapfianus [40]. The proteome is influenced by alterations in synthesis of phytohormones, especially ABA, as drought stress intensifies. Ismail et al. [41] emphasised the importance of the timing of successive phases of the induction of plant stress-tolerance. A phase immediately preceding the quiescent desiccated state has been distinguished from the desiccated state by differences in molecular responses in angiosperms [42].
5.1. ABA Induction Pathway for Desiccation Tolerance
Borya constricta is a poikilochlorophyllous resurrection plant (i.e., the foliage loses all chlorophyll during dehydration). Drought-induction of chlorophyll-loss is tightly linked with the induction of desiccation tolerance. The breakdown of chlorophyll in both homoio- and poikilo-chlorophyllous resurrection plants, is by the same Pheophorbide α Oxygenase (PAO)/phyllobilin pathway found in senescing non-resurrection plants and in maturing embryos [43]. Both chlorophyll loss and desiccation tolerance are induced concurrently in detached Borya leaves by 2 days of drought stress (in 96% RH air) or by ABA treatment of fully hydrated leaves [1]. Increases in endogenous levels of ABA (up to 14-fold) accompany the drought-induction of desiccation tolerance and yellowing in Borya leaves [44]. This suggests that drought-induced accumulation of ABA mediates both responses to drought in intact Borya plants. In intact drying Borya plants, non-senescent leaves yellow and survive the dehydration, but senescing leaves do not survive even though they also become yellow. ABA-induced chlorophyll-loss in drying immature to mature leaves is linked with desiccation tolerance but it is uncoupled from the process of senescence, which drying normally induces in immature/mature leaves [45].
In the homoiochlorophyllous resurrection plant M. flabellifolia (i.e., the foliage retains all chlorophyll during dehydration), 38% of freshly detached leaves survive drying in air of ~0% RH but their final ABA content is not high. Equilibration of detached leaves to 96% RH air raises survival to 80% while endogenous ABA accumulates up to 12-fold at 24 h equilibration [44]. In all these treatments, chlorophyll was retained and ABA appears to be uncoupled from degradation of chlorophyll. On the other hand, senescent leaves lost chlorophyll during drying. In the homoiochlorophyllous resurrection grass S. stapfianus, endogenous ABA accumulates late during drying but exogenous ABA produces little improvement in protoplasmic drought tolerance [35,46]. ABA responses in plants are influenced by interconversion of free ABA to an inactive ‘bound’ glycosylated ABA that is found in the xylem of drought stressed plants [47,48].
The leaves of desiccation-tolerant plant C. plantagineum lose about half their chlorophyll as they dry, i.e., it is intermediate between the homoio- and poikilochlorophyllous behaviour [49]. Exogenous ABA induces desiccation tolerance in well-hydrated callus of this species [50]. Several ABA-inducible proteins accumulate in C. plantagineum and S. stapfianus leaves as they dry, including protective late embryogenesis abundant (LEA) proteins, which become abundant during dehydration in resurrection plants in general and in the embryos of non-resurrection plants [51]. A hydrophilic protein LLA23, which has some similarity to LEA proteins, is induced in pollen by drying or ABA [52]. The Lilium longiflorum Thunberg protein, LLA23 may have a regulatory role in mediating stress responsive ABA signalling [53].
These findings indicate that ABA plays a considerable role in mediating the induction of desiccation tolerance in both chlorophyll -losing and -retaining resurrection foliage, in seed and presumably in pollen. That a single exogenous substance, ABA, induces desiccation tolerance in hydrated B. constricta leaves, suggests that ABA is a major regulatory factor upstream in a transduction pathway controlling the expression of a large number of genes. ABA is one of several potential signals to foliage that soil water is depleted, leading to long-term drought stress rather than short-term stress of foliage due to transient aerial conditions. Drought-induced ABA transported to the foliage triggers a range of drought resistance mechanisms [48]. It controls the expression of several drought-induced genes in S. stapfianus also; but in that species, factors other than ABA appear to be limiting and controlling the transduction pathway for desiccation tolerance [35]. During drought-induction of desiccation tolerance, the action of ABA is uncoupled from drought-induction of senescence. The importance of inhibition of senescence for desiccation tolerance has been raised by [45].
More is known of the ABA induction pathway in seed than in resurrection plants. Lack of endogenous ABA compromises induction of desiccation tolerance in maturing seed and the application of exogenous ABA may induce desiccation tolerance. In the Arabidopsis mutants, ABA deficient (aba) and ABA insensitive (abI3) and the double mutant (aba × abI3), the seed are slow to desiccate, remain green and lose viability upon drying [54]. Desiccation tolerance was not displayed in seed when the plant was both insensitive to ABA and deficient in ABA [54]. Double mutants lack ABA, acquire only low levels of storage proteins and are deficient in many low molecular weight polypeptides (soluble and bound), some of which are heat stable [54]. A major 100 kDa glycoprotein also shows a decrease in size associated with the abI3 mutation.
5.2. Transcription Factors and the ABA Induction Pathway for Desiccation Tolerance
Two important ABA receptors are PYR/PYL/RCAR phosphatase 2C-ABA complex in the cytosol and the ABA binding protein, ABAR-ABA complex in the bounding membranes of chloroplasts [55]. If ABA perception through PYR/PYL/RCAR is impaired in Arabidopsis, vegetative growth and seed production is reduced and the ABA-insensitive characteristics appear even though other ABA perception mechanisms still function [56]. The large subunit (CHLH) of magnesium-protoporphyrin IX chelatase acts as an ABA receptor in chloroplasts. CHLH catalyzes the ATP-dependent incorporation of Mg into protoporphyrin IX in the biosynthesis of chlorophyll and it also functions in a plastid-to-nucleus signalling pathway [57]. It is possible that Mg-chelatase may be involved in the breakdown of chlorophyll in drying poikilochlorophyllous resurrection plants and in catalysing the resynthesis of chlorophyll in their yellow, rehydrated leaves. The PEBP (phosphatidylethanolamine-binding protein) family is involved in ABA signalling in germinating seeds and in the biogenesis and fluidity of membranes. A putative member of the PEBP family of proteins increased 9-fold in abundance in drying plants of the desiccation-tolerant moss Physcomitrella patens (Hedw.) Bruch & Schimp [58].
Comparison of the transcriptomes of tissues of Medicago truncatula Gaertner undergoing a transition to desiccation tolerance (in drying seeds and in germinating seeds treated with polyethylene glycol −1.7 MPa osmoticum) showed changed abundance of numerous transcripts for transcription factors (TF), including two crucial TFs, MtABI3 and MtABI5 [14]. ABA responsive elements (ABRE) were present in the promoter regions of those genes exhibiting altered transcript levels—as also was found in comparisons with transcriptomes from dry desiccation-tolerant tissue in a diversity of other species [59]. Medicago truncatula mutation studies revealed that MtABI5 was necessary for osmotic stress to re-establish desiccation tolerance in germinating seed but not during drying in maturing seed. MtABI5 and MtABI3 are each upstream of large numbers of genes leading to desiccation tolerance (~14% and ~30% respectively of the genes with changed expression). The ABI3- and ABI5-pathways are considered to be complementary to and interacting with each other. ABI5 acts in the upregulation of two genes (EM1 and EM6) that encode LEA protectants [60]. ABA-responsive genes downstream of MtABI3 include genes for heat stress proteins (HSPs), genes involved in protein-repair, in microtubule-processes and encoding LEAs [14]. LEA proteins appear to be ubiquitous in the tissue of numerous desiccation tolerant life forms [61]. Comparative studies of dehydrated desiccation-tolerant species indicated that the LEA-5 (with higher hydrophobic amino-acid residues than other LEA groups) plays a crucial role in the induction of desiccation tolerance in drying tissue [59]. Increases in abundance of LEA-14 transcripts were correlated with desiccation tolerance in all species compared by Costa et al. [59].
WRKY transcription factors (TFs) bind to the W box, TTGAC(C/T), in promoters of specific genes to activate their expression [62]. The WRKY TFs occur in developing seed and are key agents in the ABA regulatory pathways that respond to environmental stresses, senescence, dormancy and germination of seeds. Various WRKY TFs act in different pathway ‘hubs’ downstream from the reception of ABA by ABAR and by PYR/PYL/RCAR [55]. In Arabidopsis, before drought stress AtWRK40 represses expression of ABI5. ABA binding to receptors removes this repression of ABI5, leading to activation of AtWRKY63, whose encoded protein in turn activates ABA-response genes, for instance RD29A and COR47.
In the foliage of the resurrection gesneriad, Boea hygrometrica (Bunge) R.Br., BhWRKY1, induced by drought stress and by ABA, binds to the promoter of the galactinol synthase gene BhGolS1 [63]. Overexpression of WRKY in rice induced expression of galactinol synthase and raffinose synthase genes and also improved drought tolerance [62]. Raffinose may play a protectant role in drying resurrection plants [64]. WRKY and galactinol synthase genes are possibly involved in an ABA-signalling pathway during the induction of desiccation tolerance in ABA-responsive resurrection plants, such as B. constricta [1]. ABA may aid the formation of protective sugars, especially sucrose, that assist ABA induction of desiccation tolerance in maturing Arabidopsis seed [54].
When desiccation tolerance was re-established in germinating seeds of Arabidopsis thaliana (L.) Heynh. by treatment with polyethylene glycol (−1.7 MPa), the most-upregulated genes in the resulting transcriptomes contained the promoter sequences (ACGT), which is often associated with ABA activity. In the majority of cases this nucleotide sequence was present in 11 different sequence ABRE motifs (ABA responsive elements) [65]. Five additional transcription factor sites (with a dehydration-responsive element (DRE)-motif and MYCATERD1, RY-repeat promoter motifs) may indicate influences of non-ABA-avenues of transcriptional control. ABA-regulated and non-ABA signalling pathways interact and converge in the induction of desiccation tolerance [66,67]. Many genes in Arabidopsis are regulated in an ABA-dependent and ABA-independent manner, e.g., RD29A [68]. In ABA-independent responses to dehydration and cold, the DRE 9bp element is an essential cis-acting element to the regulation of RD29A [68]. The Clp protease regulatory subunit encoding gene, ERD1 (early responsive to dehydration 1) responds to dehydration and salinity stress before the accumulation of ABA, which suggests that regulators other than ABA also control dehydration stress responses of Arabidopsis [69,70].
6. Potential Regulators of Desiccation Tolerance Other Than ABA
In the following sections the information on drought stress is complemented with data on other stresses. There is considerable overlap in tolerances to drying, salinity, low temperature and heat (e.g., [71]), an overlap corroborated by recent studies of the responses of gene expression to both abiotic and biotic stresses, e.g., [55,72,73,74]. Desiccation tolerance was induced in Brassica napus L. microspore-derived embryos by stress other than drought, for example heat shock (10 min) and cold shock for 2 days to the point where desiccated embryos survived 3 weeks in the dry state, before rehydration allowed renewed growth [75].
6.1. Brassinosteroids (BR), Jasmonates (JA) and Strigolactones (SL)
The first evidence that the brassinosteroid, brassinolide (BR), and methyl jasmonate (MeJa) participate in the induction of desiccation tolerance in angiosperm foliage showed that, out of 11 phytohormones and growth substances tested, exogenous BR and MeJa each gave the greatest enhancement of the protoplasmic drought tolerance of S. stapfianus mesophyll cells [46]. Electophoretic partitions (2D-PAGE) of soluble proteins from BR- or MeJa-treated plants registered 10–14 proteins with increased abundance and 3–4 with decreases. Two proteins were both BR- and MeJa-responsive. Evidence of BR synthesis was reported for drying foliage of the resurrection grass S. stapfianus [76]. Rice plants transgenically modified to synthesize elevated cytokinins under drought stress augmented the expression of brassinosteroid-related genes but diminished the expression of jasmonate-related genes [77].
Genes encoding enzymes involved in jasmonate (JA)-biosynthesis were upregulated in droughted non-resurrection plants in the Solanaceae and in salt-stressed barley plants [72,78]. In barley, JA-inducible genes were also upregulated by salinity, suggesting action of endogenous JA formed during salt-stress [78]. Pre-treatment with JA lessened salt-induced inhibition of photosynthesis in barley plants [78]. In rice (Oryza sativa L.), BR induces ABA synthesis and the BR pathway interacts with ABA, with strigolactone and with gibberellins in relation to plant stress reactions, growth and development [79]. As levels of BR increase, a maximum effect is attained but the effect declines with higher BR levels.
Whilst there have been no comprehensive studies on the involvement of strigolactones (SL) in induction of desiccation tolerance, recent reports indicate SL play important roles in responses to drought [80,81,82,83,84]. Analyses of impaired SL biosynthesis and signalling mutants in Arabidopsis suggest that SL has a role in the positive regulation of dehydration stress via both ABA-dependent and ABA-independent mechanisms [80]. The SL mutants exhibited hypersensitivity to drought and salt stress and slower ABA-induced stomatal closure. Strigolactone biosynthetic genes AtMAX3 and AtMAX4 are induced by both ABA and dehydration in wildtype (WT) plants, and the drought tolerance of both mutant and WT plants is enhanced by application exogenous SL. While SL biosynthesis genes are upregulated by drought in Arabidopsis leaves they are repressed in the drought-stressed roots of Lotus japonicus L. and tomato [82,84]. A comparative transcriptome analysis between WT and the Atmax2 SL-signalling mutant under both water-stress and well-hydrated conditions found several interesting differences in the expression profiles of ABA regulatory and transporter proteins, cytokinin-catabolizing CKX genes, as well as genes associated with flavonoid biosynthesis and photosynthetic activity [80]. Somewhat similar results utilizing max2 SL-signalling mutant were reported by Bu et al. [81], although they hypothesized that the drought-sensitivity of the mutant may result from the max2 F-box mutation interacting with other hormonal pathways. The direct role of the SL compound in drought resistance of Arabidopsis as reported by Ha et al. [80] was confirmed in L. japonicus by Lui et al. [82] who found that SL was down-regulated in roots early during osmotic stress and that the SL decrease may be required to allow ABA production. Visentin et al. [83] suggested that a reduction in SL production and its acropetal transport from tomato roots may trigger a stress response in the shoots that does not rely on ABA transport from the roots, but may lead to increased localized SL production in the shoot which could induce stomatal closure and suppress shoot branching.
6.2. Reactive Oxygen Species (ROS)
Reactive oxygen species (ROS) are produced under unstressed conditions from aberrant electron flow related to ATP-producing metabolism. ROS levels accumulate above normal levels as electron flows are perturbed under a wide range of environmental stresses including drought [85]. ROS signaling is widespread in plants [85]. As an early chemical result of stress, ROS accumulation is well placed to be a first or early signalling-trigger for adaptive responses to the stress [86], involving both systems against injury by ROS (catalase, peroxidase, dismutase, glutathione, ascorbate, isoprenoids) and also ROS-signalling toward induction of stress tolerance. Early ROS accumulation in resurrection plants would need to be high enough to trigger tolerance-induction but not so high as to produce cell injury.
In the desiccation-tolerant dicot Haberlea rhodopensis Friv. expression of catalase genes fell to ~40% during plant drying and recovered to initial values on plant rehydration. The 40% catalase expression level remains important since inhibition of catalase activity rendered the plants partially sensitive to dehydration [87]. During drying H. rhodopensis increased its total content of phenols, especially of the antioxidant myconoside, but the plant’s total antioxidant capacity fell to 70% of the level in the hydrated control [88]. In B. hygrometrica (a desiccation-tolerant species) levels of ROS were low in dry viable plants but high in plants killed by rapid drying. Contents of the antioxidant tocopherol and the expression of three enzymes controlling tocopherol biosynthesis were much higher in dry viable plants than in the rapidly-dried nonviable plants [89]. The accumulated tocopherol presumably helps limit oxidative damage in dry viable plants and during early stages of subsequent rehydration.
The desiccation-tolerant poikilochlorophyllous monocot Xerophyta humilis (Baker) Dur. & Schinz released, at moderate drought stress (~80% RWC), the volatile isoprenoid, isoprene, which protects chloroplasts against ozone and ROS [90]. The antioxidant role of volatile isoprenoids in protecting membranes was restricted to early drought stress and was replaced by the action of non-volatile isoprenoids later in dehydration of X. humilis.
Low ROS accumulation and high antioxidant ability are crucial to plant endurance of combinations of abiotic stresses [74]. Combined drought and heat stresses can cause 4-times as much agricultural loss in the USA than drought alone [74]. Cytosolic ascorbate peroxidase 1 (APX1) is needed for Arabidopsis to tolerate combined drought and heat stresses [91]. In Arabidopsis, drought and other stresses, including heat stress and viral pathogens (singly and in combination) greatly down-regulated a transcript Rap2.9, a putative transcriptional repressor. Under non-stress conditions Rap2.9 may suppress expression of stress-adaptive genes, whereas down-regulation of Rap2.9 under drought or other stresses would allow expression of genes inductive of tolerance [92]. Conversely a transcript for a G-box binding factor 3 (GBF3), implicated in ABA-signaling was markedly upregulated.
The ROS transduction pathway is considered to involve oxidative inactivation of specific proteins (by carbonylation or disulfide formation) and activation of mitogen-activated protein MAP kinases and calcium channels [85]. Genes encoding enzymes for cysteine biosynthesis and thence redox-sensitive signalling molecules in abiotically stressed plants promote survival in abiotically-stressed plants [93]. The capacity of the antioxidant defence system determines continued functioning (supporting survival) or progressive photo-oxidation (leading to injury) [94]. Nucleoside diphosphate kinase 1, associated with ROS signalling, was markedly higher in dry and in rehydrated plants than in hydrated control samples in the moss Fontinalis antipyretica Hedwig [95].
The enzyme ascorbate peroxidase (APX6) maintains low levels of ROS until, under environmental stress, ROS overwhelms the capacity of the APX system. Increasing ROS stimulates synthesis of auxin and of ABA, which inhibit germination [86]. ROS can accumulate, under temperature and salt stress, in the phloem apoplast where it mediates a rapid signalling pathway that is independent of other signalling pathways [39]. Ismail et al. [41] propose that during salinity stress, cell uptake of Na+ and Ca++ within minutes, stimulates both accumulation of ROS and rapid removal of Na+ and Ca++ (by vacuolar uptake and extracellular secretion). Removal of the ions curtails the Ca signal. The amplitude of the Ca signal and the speed of its attenuation determine which of two subsequent parallel pathways predominates. Rapid attenuation of the Ca signal favors ROS signalling, its stimulation of ABA signalling leading to increased stress tolerance. Slow Ca signal attenuation diverts ROS signalling to produce an excessive activation of the JA pathway which leads to oxidative damage to membranes. In the resurrection grass S. stapfianus, JA was found to have a beneficial rather than deleterious effect on protoplasmic drought tolerance [46].
ROS, ABA and JA all appear to be involved in the induction of desiccation tolerance. The processes described by Ismail et al. [41] for induction of salt-tolerance may well be operating in resurrection plants during induction of desiccation tolerance as plants progress through drying, but further research is needed in resurrection plants to elucidate this possibility. It is conceivable that since ROS can be transported in phloem, ROS levels rise in the foliage apoplast as part of a signal translocated from drought-stressed roots in drying soil. The primary triggering role that Ismail et al. [41] ascribed to uptake of apoplastic Na+ in signal transduction in salt stress/tolerance is less likely in resurrection angiosperms, which usually grow in habitats with low soil Na+ (at high altitudes and distant from coasts). Immense turgor pressures strain the plasmalemmas of plant cells (commonly up to 2 to 3 MPa) in the early stages of drought stress until leaves wilt. Altered inflow of calcium via mechano-sensitive calcium channels [41] may conceivably result from changes in pressures on membranes and trigger signalling for induction of desiccation tolerance.
Strigolactones and the recently-reported messenger agents, acetate [96] and CLE25 peptide [97] have yet to be examined as possible root-to-shoot signals in the induction of desiccation tolerance: CLE25 synthesized in droughted Arabidopsis roots is transmitted in the vascular system to leaves where it acts with BARELY ANY MERISTEM (BAM) receptors to influence ABA-synthesis and thereby produces closure of stomata [97]. In Arabidopsis plants, drought stimulated the synthesis of acetic acid, while 10–20 mM acetic acid applied to the roots lowered the pH of xylem sap and ultimately promoted the drought resistance of the plants. A similar promotion of drought resistance was found in plants of maize, rice, rapeseed and wheat [96].
7. Sucrose and Induction of Desiccation Tolerance
When the aquatic moss F. antipyretica, was dried slowly in 50% relative humidity to reach 25% RWC, the plants survived for one month and an increase in the synthesis of proteins associated with sucrose production was observed [95]. Pretreatment of cell suspensions from S. stapfianus leaves with high concentrations of sucrose for 1 h enhanced the protoplasmic drought tolerance (PDT) markedly from equilibration to <50% RH compared with 86–87% RH for controls. In plants pretreated with separate solutions of glucose, fructose, trehalose or raffinose, the PDT remained at the control levels [98]. The cells absorbed sucrose rapidly, doubling their osmotic pressure, whereas the four other sugars produced no detectable change in osmotic pressure. Sucrose absorption into suspended S. stapfianus cells was suppressed by PCMBS (p-chloromercuribenzenesulfonic acid) an inhibitor of the proton-sucrose symport of the plasmalemma [99].
The improvement induced by sucrose in the PDT of S. stapfianus cell suspensions (~36%RH) was much greater than that induced by pretreatment with methyl jasmonate/jasmonic acid, or brassinolide or ABA (3%, 3% and 1% RH respectively; [46]). The need for high concentrations of pretreatment with sucrose and the rapid absorption of sucrose suggest that in S. stapfianus the stimulation of PDT may be a direct protective effect of the sucrose and/or be a regulatory role by high intracellular sucrose concentrations. These data indicate that sucrose-loading into young leaves of S. stapfianus is important for induction of desiccation tolerance. In intact drying resurrection plants, considerable sucrose would be supplied by remobilization of carbohydrates from senescing leaves via the phloem.
The SUT4 subfamily of sucrose proton co-transporters and sucrose facilitators displays broad physiological involvements and directions of regulation which vary between species-variation that in part, is linked with whether the path of sucrose to the phloem is via the apoplast or via the symplast [100,101]. Moreover, the promoter of the SUT4 homolog in Arabidopsis contains stress-responsive regions whereas the homolog in rice has mainly hormone-related elements [102]. The situation in these two regards is not known in the resurrection angiosperms. The widely separated taxa containing resurrection angiosperms may well differ in the mechanism of sucrose transfer.
Mechanisms of regulation by sugars in non-resurrection plants are reviewed by Smeekens and Hellmann [103]. Target of Rapamycin (TOR) kinase complexes are stimulated by glucose and repress synthesis of sucrose and starch [104]. In S. stapfianus glucose accumulation occurs early during drought stress, while starch and sucrose contents are decreasing [64], consistent with TOR kinase associated sugar accumulation. However, low levels of glucose and high levels of sucrose are seen in later stages of desiccation where drought stress is severe [64]. Sucrose accumulation and transport is for the most part associated with the sucrose-related kinase SnRK1 (addressed in Section 8).
8. The Role of Trehalose-6-Phospate in Desiccation Tolerance
While trehalose is the major protective sugar that accumulates in some desiccation-tolerant species, such as nematodes and tardigrades, sucrose is the predominant protective sugar accumulating in drying resurrection plants, with trehalose and other sugars accumulating at lower levels [64]. The exceptions to this are Selaginella spp. that also accumulate trehalose to high concentrations when dehydrated, however, there is no strong link to desiccation tolerance as other desiccation-sensitive members of Selaginella spp. also accumulate high levels of trehalose [105]. Craterostigma plantagineum and Lindernia brevidens Skan. also accumulate sugars in response to desiccation [42,106]. Fully hydrated leaves of C. plantagineum contain high levels of the unusual sugar 2-octulose which upon dehydration is converted to sucrose [107]. This interconversion of 2-octulose to sucrose is conserved in the close relative, L. brevidens [106].
Several studies have demonstrated that exogenously applied, and genetically enhanced trehalose content results in a higher capacity for withstanding drought [108,109], however, the direct mechanism whereby trehalose fuels this response remains to be elucidated. Trehalose biosynthesis involves the enzymes trehalose-6-phosphate (T6P) synthase (TPS) and T6P phosphatase (TPP). TPS generates T6P from derivatives of sucrose metabolism and other pathways, followed by the action of TPP, which removes the phosphate generating trehalose. T6P and the genes controlling T6P synthesis and conversion to trehalose have been implicated in the regulation of photosynthetic ability, starch synthesis and breakdown, glycolysis-linked hexokinase activity, and embryo maturation [110]. The role of trehalose as a minor sugar is not clear, however trehalose is thought to accumulate at low levels due to the rapid degradation of trehalose into two glucose units by trehalase [111].
Water deficit results in the rapid inhibition of growth yet photosynthesis can continue and result in increases in cellular carbohydrate contents [112]. Key carbon storage molecules, such as sucrose, are generated through photosynthesis and can be stored or transported to where they are required within the plant [113]. T6P has been implicated as an indication of cellular sucrose status where T6P levels are directly proportional to sucrose levels, termed the T6P: sucrose nexus [114]. Importantly, T6P has been shown to inhibit the sucrose homeostasis enzyme SnF-Related Kinase 1 (SnRK1) [115]. Put simply, SnRK1 inhibited by T6P directs the “feast” response, whereas uninhibited SnRK1 directs a “famine” response. T6P-inhibited SnRK1 will up-regulate carbohydrate storage, prime gene expression for growth, growth recovery and signals satiety, whereas uninhibited SnRK1 will signal starvation, catabolism and will slow growth. Indeed, the T6P:SnRK1 relationship is considered a major player in directing carbohydrates between source and sink tissues, and can be a key factor in the survival of environmental stress [116,117,118,119]. This has been addressed in Arabidopsis, where accumulated T6P under sink-limited stress conditions inhibits SnRK1 activity, and thus allows synthesis and growth to resume rapidly once the growth limiting stress conditions were alleviated [118]. A similar acceleration of growth is observed in S. stapfianus that has been through a dehydration cycle [36].
Evidence of T6P affecting changes in carbohydrate storage and desiccation-related processes has been demonstrated in both transgenic Arabidopsis and maize. Arabidopsis with increased T6P content stimulated rapid starch synthesis (requiring SnRK1) following redox-activation of adenosine-5′-diphosphoglucose (ADP-glucose) pyrophosphorylase (AGPase) [120,121]. This rapid increase in starch mirrors that which is seen in the rehydration of most resurrection plant species following the loss of starch in the earlier stages of drought stress [122,123]. Conversely, a decrease in T6P during embryo development in Arabidopsis, demonstrated in an AtTPS1 mutant, prevents the accumulation of storage molecules during the latter stage of seed development [124]. In addition, there is evidence that trehalose phosphate/trehalose interconversion acts as a system regulating the vascular flow of sucrose to Zea mays L. inflorescences and protects florets against drought injury [119]. It is possible that this system, catalysed by TPP, may assist the phloem-transportation of sucrose from remobilized carbohydrate in old leaves on drying resurrection plants to young leaves, so assisting the survival of young plant-parts.
Furthermore, T6P has also been associated with ABA signaling, where low levels of T6P correlated to hypersensitivity to ABA in early stages of embryo development [125]. The induction of the desiccation tolerance pathways in some resurrection plants will occur as relative water content falls below 60% (Table 3), whereas in non-resurrection plants drought-related ABA processes, such as stomatal closure, will occur earlier during water deficit [125]. It seems likely that T6P driven ABA signaling, where high T6P levels presumably instigates ABA hyposensitivity, would be required for the prevention of innate drought-related responses prior to the activation of the ABA-dependent desiccation-tolerance pathway [126].
The trehalose biosynthesis pathway potentially has a significant role in the establishment of intracellular glasses that form within desiccated plant tissues in a process called vitrification. During this process, significant increases in sucrose, in addition to increases in raffinose, trehalose and other sugars come together to form intracellular glass [127]. Sugar accumulation has been observed during drought stress in transgenic tobacco expressing the E. coli TPS gene [128] providing indirect evidence of the potential involvement of the trehalose biosynthesis pathway in some aspects of vitrification observed in desiccation tolerance. Trehalose has a high glass transition temperature, and forms strong hydrogen bonds with water [129]. Furthermore trehalose is thought to provide stabilisation to proteins, and prevents membrane fusion [130]. This may potentially help in the formation of a scaffold during vitrification, which forms the building blocks for T6P-driven sucrose accumulation during the desiccation tolerance network.
While the above mentioned are important during the dehydration cycle, the rehydration cycle is equally as important, however, sparsely covered in scientific literature. Mentioned above, transcripts, proteins and energy-related compounds are essential in desiccation tolerance and as metabolism slows as resurrection plants enter the quiescent state, these sensitive compounds need to be stored. In Tortula ruralis (Hedw.) Gaertn., Meyer & Scherb. mRNPs (micro RiboNuceloProteins) accumulate during water loss, and are possibly involved in mRNA storage and protection [131]. This discovery offers a potential mechanism of rapid protein synthesis post-rehydration.
9. Protein-Synthesis in Resurrection Plants under Drought Stress
As resurrection plants dry, protein synthesis is necessary for the production of the specific proteins associated with implementation of desiccation tolerance and for the specific proteins regulating implementation. The major changes in the protein complement during induction of desiccation tolerance imply changes in gene expression and turnover of degraded protein. In resurrection plants protein synthesis continues to surprisingly low water contents. In the resurrection monocot Xerophyta villosa Th.Dur. & Schinz. protein synthesis rates increase markedly below 60% RWC before declining again at 30% RWC (Table 4). In B. hygrometrica, increased abundance of transcripts related to chaperones, to ubiquitination and to unfolded-protein-stress signaling in dry desiccation-tolerant and desiccation-sensitive plants, indicated unfolded-protein-stress occurred in both cases [89]. Transcripts related to autophagy accumulated in the drying desiccation tolerant plants. Specific staining for autophagosomes in these plants remained at levels seen in the hydrated controls, whereas this staining decreased in drought injured plants [89]. Autophagy may be contributing to switching cells from a growth-related state to a quiescent/dormant dry state.
Transcripts encoding biosynthetic enzymes generally decreased and those encoding catabolic enzymes generally increased in B. hygrometrica plants surviving dehydration [89]. Increased hydrolytic breakdown of cell wall constituents may raise the plasticity of the cell walls. Allowing folding of the cell walls in the resurrection plant during drying reduces the risk of mechanical injury to tissues and may augment growth following subsequent rehydration of plants.
Numerous proteins change in abundance in all the studies of drying resurrection angiosperms and in desiccation-tolerant (orthodox) seed. Transcriptome and proteome studies of desiccation tolerant plants compare the initial hydrated tissue with air-dry phase [58,76,87,95]. All the studies found high numbers of transcripts or proteins with large increases or decreases in their abundance during drying. Specific transcripts with >50% change in abundances during acquisition of desiccation tolerance in drying Arabidopsis seed numbered almost 7000 [12]: 74% of the 7000 transcripts were specific to drying seed. A major shift clearly occurred from a growth/development metabolism before drying to a desiccation-tolerance metabolism during drying.
As the homoiochlorophyllous resurrection grass S. stapfianus dries, the proteomic changes vary with the different RWC levels before and during the appearance of desiccation tolerance [40,51,133,134]. In the poikilochlorophyllous resurrection monocot, B. constricta, complete chlorophyll-destruction in drying leaves is tightly linked with the induction of desiccation tolerance [135]. Following 1D-PAGE partitions of soluble protein extracts from B. constricta leaves, the protein-pattern for yellow-green leaves undergoing induction of desiccation tolerance resembled the pattern for green leaves killed by dehydration (strong emphasis of a peak of mid-molecular-weight proteins in both cases). Whereas the protein-pattern for leaves that had become fully yellow and desiccation-tolerant, showed restoration of a three-peak pattern similar to that for fully hydrated control leaves. It appears that in drought-hardening B. constricta, an early phase moves toward injury and a subsequent phase moves toward reinstatement of a ‘hydrated pattern’ as a basis for successful recovery in subsequent rehydration. In drying plants of X. villosa an early phase of declining protein synthesis is followed by a phase where protein synthesis rises above levels in fully-hydrated control plants, and then a phase of decline to no protein synthesis at ~20% RWC [132].
Transcript for a protein initiation factor eIF1 became abundant in drying S. stapfianus plants at moderate stress (60–79% RWC, comparable to the phase of increasing polysome content in drying X. villosa) and was most abundant in airdry leaves [51]. Thus eIF1 content may limit the rate of protein synthesis during early induction of desiccation tolerance but eIF1 is in good supply for protein synthesis early in rehydrating leaves. The desiccation-tolerant moss F. antipyretica had low abundance of RNA-binding proteins, elongation factor 1-α and translation initiation factor eIF-4A in dry plants; the level of the former protein remained low on rehydration and recovery whereas the eIF-4A protein accumulated to almost twice the control value [95].
Desiccation-tolerant plants maintain their ATP content as they dry, while in several resurrection species the ATP content in the air-dry state exceeded levels in hydrated plants [136] indicating higher ATP synthesis than ATP usage. Drying desiccation-tolerant angiosperms show accumulation of ATP synthases which would support the ATP levels in drying and rehydrating plants (e.g., S. stapfianus, [76]; H. rhodopensis, [87]). Slow-dried moss F. antipyretica accumulated one form of ATP synthase during drying but a second form declined in abundance [87]. Abundance of a putative ATP synthase almost doubled during drying in the desiccation-tolerant moss P. patens [58]. ATP contents do not appear to limit protein synthesis in drying desiccation-tolerant plants.
Processes in Seed and Pollen during Desiccation
There have been many studies of seed- and pollen-specific gene expression relating to the process establishing desiccation tolerance as either the seed or pollen matures [137], but it has been difficult to untangle the expression of genes directly involved in desiccation tolerance from those related to other aspects of seed and pollen development. Molecular studies in Arabidopsis have shown that mutations in the transcription factors ABSCISIC ACID INSENSITIVE 3 (ABI3), LEAFY COTYLEDON2 (LEC2) and FUSCA3 (FUS3) are lethal at the late stages of seed maturity. These transcription factors contain the B3 domain, a highly conserved DNA binding region specific to higher plants [138]. FUSCA3 for example, has a role in regulating ABA and gibberellic acid (GA) levels and is involved in controlling seed dormancy and preventing germination under adverse conditions [138]. Additional transcription factors identified which may have a role, such as DREBs (dehydration responsive element binding) and TaNAC69, are stress-related and expressed in pollen/anthers during development [139].
Costa et al. [59] compared published transcriptomes of plant species following induction of desiccation tolerance (leaves of the resurrection plants C. plantagineum and S. stapfianus, germinating seeds of M. treated with ABA or osmoticum, and the lichen Cladonia rangiferina (L.) Weber ex F.H.Wigg). Of 12 changes in specific transcript abundance with induction of desiccation that were consistent across all five species, only two showed elevated abundance. These, include the gene transcripts for the genes LEA-14 (AT1G01470) and NAD (P)-linked oxidoreductase superfamily protein (AT5G01670) that have putative roles in protein-protection and counter-oxidation, respectively. Comparison of transcripts for all datasets with the genome sequence data for Arabidopsis, yielded a core of 260 Arabidopsis orthologs, of which “77 increased and 184 decreased in abundance”. The categories of those that accumulated were broad in coverage, and included, lipid storage, multicellular organismal processes, seed development, reproduction and response to temperature, whilst those decreasing in abundance related to carbohydrate metabolism and biosynthesis of phenylpropanoids. Data on regulatory motifs in the cis-acting promoter elements in the “77-increased-abundance” core genes highlighted genes responsive to ABA (ABRE binding site motif), light regulation, stress (DRE core motif), circadian rhythm (EveningElement promoter motif) and gibberellin (GADOWNAT), whilst promoter motifs for the “184-declined-abundance” core genes also included the ABA related ABRE binding site. Once again an important role of ABA inter alia is evident.
Studies of mutant lines by Costa et al. [59] apply to the full period of induction of desiccation tolerance. Analysis of T-DNA mutant Arabidopsis lines for 13 genes chosen from the 260 DT set, revealed mutations in three genes (LEA 5-1, PCK1, and RCI2H) which resulted in reduced seedling ability to re-induce desiccation tolerance. In further studies of the T-DNA mutant lines, dormancy was increased by disruption in 5 genes (AT3G10020, LEA7, PCK1, RCI2H and SAG29) and seed longevity was lowered in the PCK1 line.
10. Identification of Genes Involved in Desiccation Tolerance
Protection against the damaging effects of desiccation is essential to enable the vegetative parts of resurrection species to remain viable and intact in their typically dry, high light and otherwise hostile environments, often for extended periods of time [11,140]. Most of the initial reports dealing with the molecular genetics of desiccation tolerance involved the use of a range of differential cDNA screening procedures to identify genes with altered (mainly increasing, though some decreasing) transcript abundance in RNA extracted from leaf tissues at various stages in the dehydration process. Although the number of desiccation-related genes identified in such studies was relatively small, their characterization did provide valuable insights into the molecular processes that underpin the capacity of plants to survive desiccation. These included upregulation of genes encoding enzymes likely to reduce negative effects of reactive oxygen species, glycine-rich components of cell walls and membrane intrinsic aquaporin proteins that facilitate transport of water molecules across cellular membranes in response to moderate drought stress [51,133,134,141,142,143,144,145,146,147,148,149,150].
Dramatic increases in transcript abundance of LEA-type dehydrin proteins were also observed in many of these studies. This broad group of hydrophilins is generally regarded as providing a broadly protective role against drought and related stress-induced denaturation of cellular proteins [151,152,153]. Furthermore, in C. plantagineum changes in lipid metabolism have been associated with the establishment of desiccation tolerance. The removal of MGDG (MonoGalactosylDiacylGlycerol) a conic-shaped non-bilayer-forming chloroplast lipid was strongly associated with desiccation tolerance. A fraction of MGDG is converted to oligogalactolipids in the chloroplast envelope, which are then hydrolysed and sent to the ER (Endoplasmic Reticulum), these are then used for the synthesis of phospholipids. Remaining MGDG may be used for the synthesis of phosphatidylinositol (PI), a key lipid involved in signalling and membrane trafficking. PI only accumulated during the dehydration of desiccation-tolerant species [154].
As methodology and cost effectiveness of DNA analysis has steadily improved over recent years, involving larger scale transcriptomic studies and entire genomic analysis of monocotyledenous and dicotyledonous species from different regions of the globe, so there has been a steady increase in the number and range of gene transcripts that appear to be involved in the onset of desiccation tolerance [59,87,101,155,156,157,158,159,160,161]. Although there are differences in the specific molecular responses of different types resurrection plants, probably reflecting lineage-specific adaptations to widely variable geographic areas and localised responses to environmental stresses, together analysis of the predicted proteins encoded by many of these genes does point to them playing a role in protection of cellular components from the otherwise destructive effects of water loss and oxidative damage upon cellular biochemical processes and structural integrity of cell walls and entire tissues.
Interestingly, in general, these ‘omic’ studies are also supportive of the earlier hypotheses that vegetative desiccation tolerance has independently evolved in different taxa via reactivation of pathways normally restricted to seed maturation in plants (e.g., [137,155,159,162]. A major challenge over the next few years will undoubtedly be to understand the nature of regulatory genes, and their activation, which control the expression of these ‘protective protein’ genes at appropriate stages of the desiccation process. Some promising results have been reported here (e.g., [73,163,164,165]) but much remains to be discovered in this area of investigation.
11. Direct DNA Manipulation of Desiccation-Related Genes
Effective gene transfer procedures in many plant species have enabled DNA disruption, RNAi-mediated or viral-induced gene silencing protocols to be developed for researchers wishing to study the role of any given gene in a developmental or biochemical process (e.g., [166] and references therein). Transformation procedures have been reported for some resurrection species, such as the well-studied dicotyledenous species C. plantagineum [167] and also L. brevidens [168]. The latter species is a more recently described resurrection plant, also from within the family Linderniaceae, and interestingly is found naturally in the montane rainforests of Tanzania and Kenya [106]. However, reliable DNA transformation protocols for other desiccation-tolerant plants have not been reported and development of such methods would be extremely beneficial to the subject area, particularly involving desiccation-tolerant monocotyledonous species. Such procedures may also allow CRISPR/cas9-mediated genome editing technology to be employed in direct functional genomics studies to investigate the role of particular genes in conferring desiccation tolerance in such species, as has recently been demonstrated with other genes in monocotyledonous crops [169,170]. Another powerful functional genetics methodology, which does not necessitate the development of DNA transformation technology, involves targeting induced local lesions in genomes (TILLING) [171,172]. This approach has been successfully used to identify mutants in monocotyledonous crops that possess disruptions in specific genes [172,173,174]. An alternative approach which has proven valuable and which may be of direct interest to agricultural crops, is the ectopic expression of genes from resurrection plants in otherwise desiccation-sensitive species where reproducible transformation protocols do exist.
One of the first reports of successful ectopic expression of drought activated genes from the resurrection plant C. plantagineum involved transgenic tobacco [175]. Here three genes, encoding a serine and lysine-rich repeat motifs protein, a LEA-type protein and a novel sequence, were introduced under control of a modified version of the powerful Cauliflower mosaic virus 35S (CaMV35S) promoter. Although the introduced proteins were detected, these transgenic plants did not appear to show enhanced tolerance to drought stress [175]. However, ectopic expression of LEA-4 group genes from the resurrection plant B. hygrometrica did confer elevated dehydration tolerance in transgenic tobacco [176]. In separate studies, ectopic expression of the dehydration inducible XvSAP1 gene from Xerophyta viscosa Baker in tobacco increased osmotic-, salinity- and high-temperature-stress tolerance in transgenic plants compared to controls. The XvSAP1 gene encodes a highly hydrophobic and likely membrane-bound protein with significant identity to a cold-induced dehydrin-like gene from wheat [145,146]. Also using X. viscosa as a source of stress-related genetic information, it was observed that ectopic expression of aldose reductase (ALDRXV4) conferred enhanced drought and also salinity tolerance on transgenic tobacco plants [177], most likely due to enhanced scavenging of methylglyoxal and reduction in the level of abiotic stress-induced membrane damage. Over expression of a MYB-type transcription factor gene (CpMYB10) from C. plantagineum in Arabidopis increased drought (and salt) tolerance of transgenic plants, apparently via elevated sensitivity to endogenous ABA levels [164]. Constitutive expression of the dehydration-induced family 1 uridine diphosphate (UDP)-glycosyltransferse (UGT) gene SDG8i, from S. stapfianus in Arabidopsis led to altered growth characteristics in transgenic plants, which possessed significantly enhanced drought tolerance compared to non-transgenic plants [73]. The underlying reasons for the increased drought tolerance here may be related to altered hormonal metabolism in transformants, which, in turn, may have activated or enhanced endogenous protection mechanisms dealing with abiotic stress in general [73]. Interestingly here, overexpression of a related Arabidopsis family 1 UDP-glycosyltransferase gene UGT87A2, in Arabidopsis, yielded transgenic plants also exhibiting enhanced tolerance to drought and other abiotic stresses. Analysis of these transgenic plants showed increased expression of a number of stress-related genes compared to control Arabidopsis plants [178]. Focussing on crop species of direct relevance to global food security, ectopic expression of a drought induced mRNA Splicing-Activated Endoplasmic Reticulum Stress Regulator gene (BhbZIP60) from B. hygrometrica in rice [160] and over-expression of an osmotin gene (TIOsm) from the resurrection grass Tripogon loliiformis F. Muell, also in rice, produced transgenic plants which displayed improved tolerance to drought compared to their non-transgenic counterparts [179].
As the 21st century unfolds, plant breeders, farmers and the wider public will need to consider the role of biotechnological approaches to combat deleterious effects of drought stress which are predicted to steadily increase in the face of ongoing climatic change, diminishing available arable land and increasing global population pressures [180,181,182]. The few examples noted above, albeit largely laboratory based and obviously quite limited in scope, are encouraging for future applications of resurrection plant molecular genetics and genomic studies to agricultural production and environmental sustainability.
12. Stress-Induction of Senescence in Mature and Premature Leaves
Griffiths et al. [45] postulate that drought-invoked senescence is a critical cause of cell degeneration and death in the pre-mature/mature leaves of non-resurrection plants and that suppression of such drought-induced senescence is crucial for resurrection plants to survive desiccation. The immature and mature leaves of homoiochlorophyllous resurrection plants undergo little change in organelle microstructure during drying [123,183]. In drying poikilochlorophyllous resurrection plants, the leaves lose chloroplast grana and thylakoids transform into vesicles [122], i.e., the resulting plastids resemble the proplastids of meristematic cells. This indicates that drought-induced post-mitotic senescence is uncoupled from processes that would lead to irreversible cell injury in resurrection plants.
The desiccation-tolerance-correlated gene SDG8i (from the resurrection grass S. stapfianus) encodes an enzyme UDP glycosyltransferase, which glycosylates an analogue of strigolactone (SL) [45,73]. SL is a phytohormone that in non-resurrection species inhibits growth of buds, promotes reallocation of resources from senescing leaves, promotes growth of interfascicular cambium and alters root architecture [184]. Some of these effects are mediated by interactions of SL with auxin and cytokinin. Glycosylation of indole-3-acetic acid (IAA) and ABA is known to produce pools of the reversibly deactivated hormones [185]. This suggests that glycosylation of SL may deactivate SL. Transcripts of SDG8i accumulate during induction of desiccation tolerance in young leaves of S. stapfianus, possibly leading to glycosylation of SL and preventing SL-stimulation of senescence in young leaves as they dry and rehydrate [45]. SDG8i transcripts remain high in dry foliage [134]. Deactivation of the SL during drying would also allow the rapid growth observed in desiccation-tolerant shoots of S. stapfianus following their subsequent rehydration [36].
Exogenous SL increases the overall drought resistance of plants, including by stimulating root growth and stomatal responsiveness [80,184]. SL biosynthesis from carotenoid is regulated by genes in the MAX (more axillary growth) gene family [186]. Whereas SL inhibits bud growth, cytokinins promote it. The actions of both hormones target the gene for a transcription factor BRC1 that leads to suppression of bud growth [187]. Comparison of the transcriptomes of desiccation tolerant tissues in diverse plant species, led Costa et al. [59] to emphasize the overlap of gene expression changes related to desiccation tolerance with those related to dormancy. It is noteworthy then that SL appears to be implicated in bud-dormancy and possibly desiccation tolerance. The SL signalling pathway involves the protein ORE9/MAX2, which promotes senescence [188,189] whereas expression of SDG8i UGT inhibits senescence, possibly by altering ORE9/MAX activity [45].
Some breakdown products and reactive oxidants that accumulate during senescence may accelerate senescence [190]. Products of lipid catabolism include precursors for synthesis of jasmonic acid, a senescence-promoting hormone [191]. Exogenous methyl jasmonate and brassinolide improve the protoplasmic drought tolerance of non-senescent leaf cells from hydrated S. stapfianus plants [46]. Sugars accumulate from starch hydrolysis during senescence and during drought stress. Transgenic studies using the promoter of the senescence-associated gene SAG12 (an Arabidopsis gene encoding a cysteine proteinase), fused to a gene catalysing cytokinin synthesis, linked overproduction of cytokinin with the early onset of senescence and high hexose contents [192,193]. Sugar accumulation is general during drought stress in both resurrection and non-resurrection plants. Drying resurrection plants appear to have either a mechanism for preventing high-sugar-induced senescence or from uncoupling senescence from production of cell injury.
Senescence involves differential expression of a large number of genes (many already expressed to some degree in non-senescent tissue) in monocot and dicot species [194,195]. Roles include hydrolysis of macromolecules, remobilization of nutrients, protection against oxidation and against pathogen [192,196,197,198].
13. Epigenetic Modifications
Epigenetic regulation has also been linked with differentiation and development in plants as well as with the responses of plants to their environment, including reproduction, pathogen invasion and cold stress [199,200,201]. Differentiation (organ-type), development (juvenility, maturity or senescence) and environment (drought stress) together determine the ability of resurrection plant organs to become desiccation tolerant [10]. Epigenetic regulation thus appears to lie at the intersection of these three factors and so presents as a likely over-arching mode of regulation of the expression of desiccation tolerance. The major epigenetic chromosomal ‘tags’ controlling availability for expression are DNA methylation, histone modifications and RNA mediated regulation (small interfering (si) RNA, micro (mi) RNA and long non-coding (lnc) RNA) [202]. Epigenetic regulation allows rapid and flexible responses to both short- and long-term stresses, resulting in increased plant plasticity in response to environmental conditions. In recent years there have been extensive reviews of epigenetic regulation of gene expression in plants and the reader is referred to these [202,203,204,205,206]. Here we focus on the potential role of epigenetics in the evolution of extreme desiccation tolerance.
There are only a few published reports on the identification of epigenetic mechanisms associated with desiccation tolerance in resurrection plants. In C. plantagineum an ABA-inducible retrotransposon, CDT-1, has been shown to control the synthesis of a 21 bp double stranded siRNA that has some sequence similarity to miR159 [19,207]. miR159 has been implicated in both drought and ABA responses in plants [208]. To show that the CDT-1 siRNA was involved in gene regulation, C. plantagineum protoplasts were transfected with the siRNA, resulting in increased expression of desiccation-associated genes compared to control protoplasts, indicating a role for this siRNA in the induction of the desiccation tolerance pathway [19]. siRNA and miRNA play an important role in stress signalling networks and are induced by abiotic and biotic stresses in many plant species [209]. The Craterostigma siRNA may act as a signal of stress, passing through plasmodesmata, and may initiate desiccation tolerance pathways, though the specific target sequence of CDT-1 is unknown [19]. A screen of a C. plantagineum genomic library showed that the CDT-1 retrotransposon is present at several loci but flanked by different sequences. This suggests that through transposition of CDT-1, there has been an increase in the synthesis of siRNA and as the stress increases greater amounts of transcripts are induced, which leads to an increase in the transcription of desiccation tolerance-related genes. Further screens of C. plantagineum identified an additional mutant, CDT-2, which showed similarities to CDT-1 [210]. CDT-2 also leads to increased expression of desiccation associated genes and results in callus that is desiccation tolerant without application of ABA. This suggests the involvement of the CDT-1/2 gene family in ABA signal transduction. A similar retrotransposon to CDT-1/2 has also been identified in B. hygrometrica [211]. Within the 49 kb LTR-retrotransposon a smaller element of ~2 kb was identified and named Osmotic and Alkaline Resistance 1 (OAR1). Transformation of Arabidopsis with AOR1 conferred tolerance to both osmotic and alkaline stress. When stressed, short transcripts originating from OAR1 increased in both B. hygrometrica and the transformed Arabidopsis. In addition, stressing B. hygrometrica resulted in an increase in the OAR1 copy number [211]. With the advances in molecular techniques the genomes of three resurrection plants have been sequenced in recent years; B. hygrometrica [161], Oropetium thomaeum (L.f.) Trin. [158] and X. viscosa [155]. The proportion of transposable elements (TE) in each genome varied considerably, with 18% in X. viscosa, compared to 43% in B. hygrometrica and 75% in O. thomaeum. This suggests that the level of TEs in a genome is not directly linked to desiccation tolerance, but they appear to play important roles, possibly in the evolution of desiccation tolerance in these unique plants.
Histone modifications have been identified as important for gene regulation in response to abiotic stress in non-resurrection species. The promoter of a LEA gene from C. plantagineum was used in a yeast one hybrid screen to isolate binding proteins resulting in the identification of three histone H3 variants [212]. Characterizations of these proteins suggest that they may be involved in gene regulation via histone modifications, but more studies are required to confirm this. Histone variants replace core histones in the nucleosome resulting in structural and functional variations. Proteome analysis of S. stapfianus leaf tissue identified a protein with similarity to SNF2P that doubled in abundance in drying leaves [76]. SNF2 proteins are nucleus-located ATP-dependent helicases that are involved in chromatin remodelling and have also been implicated in conserving domains for specific histone variants, thereby altering chromosome accessibility and hence gene expression [213].
A similar regulatory role to that of miRNA is exerted by RNA-binding proteins that modify mRNA activity and turnover of mRNA [214]. More than 1000 RNA-binding proteins have been identified from eukaryotes, but as yet none have been examined in relation to desiccation tolerance to the authors’ knowledge. The rice ‘alba’ protein (‘acetylation lowers binding affinity’), which binds to nuclear DNA and responds to drought stress and to oxidative stress, could be implicated in epigenetic regulation of stress responses [215]. This protein appears to have ancient origins since a related alba-like protein binds with DNA and RNA in the Archaea [215].
It is hypothesised that the mechanisms that operate in seed and pollen, resulting in desiccation tolerance, were re-activated in the vegetative tissue of resurrection plants [155,159,162]. To date, several genes involved in seed maturation have been linked to desiccation tolerance in resurrection plants. However, support for this theory and the involvement of epigenetic regulation in desiccation tolerance would come from the identification in resurrection plants of epigenetic mechanisms characterised in seeds. Epigenetic involvement in seed desiccation tolerance is suggested by the identification of DME-family glycosylase genes in orthodox desiccation-tolerant seed of barley (HvDME) and Arabidopsis (e.g., AtDME) [216]. HvDME transcripts accumulated in maturing seed and drought-stressed vegetative organs. Differences in methylation levels of HvDME genes in two barley cultivars were indicative of species-related epigenetic distinctions. Other modifications include histone acetylation, which has been shown to mediate ABA sensitivity during seed germination [217], whilst the specific histone deacetylase 6 (HDA6) affects the expression of some of the genes induced during acclimation to freezing [218]. The expression of the phas gene, encoding the seed storage protein phaseolin, is induced during seed embryogenesis but is silent in vegetative tissue. Analysis of the promoter of phas indicated that the chromatin structure of the gene represses expression in leaf tissue. However, during embryogenesis the ABA insensitive 3 (ABI3)-like factor (PvALF, a putative activator), in conjunction with ABA, disrupts the chromatin structure leading to phas expression [219]. DNA-methylating/demethylating enzymes (‘DNMT’ DNA methyltransferases/DNA glycoylases) or histone-acetylating/deacetylating enzymes (‘HAT’ histone acetyl transferases/‘HDAC’ histone deacetylases) are not evident in proteome-investigations of dehydrating desiccation-tolerant plants [58,76,87,95], suggesting their abundances may be too low to be readily detected.
Epigenetic modifications can be reset during mitosis/meiosis but it has also been shown that these changes can be passed to subsequent generations and provide a mechanism for short-term stress memory (acclimation) as well as long-term stress (adaptation) [220]. Priming of Arabidopsis seedlings with mild salinity, followed by a period of non-saline growth conditions, increased the salt resistance and drought resistance of the plants to later salt-stress. This stress priming was accompanied by tissue-specific epigenomic changes [220]. Changes in profiles of four histones across the genome were particularly evident with tri-methylated lysine in histone 3 (H3K27me3), which persisted during seedling growth in normal non-saline conditions [220]. The histone modification, H3K4me3 also appears to function as an epigenetic mark of stress memory [201].
14. Conclusions
We suggest the following tentative hypothesis for the regulation of desiccation tolerance in the flowering-plants, the group that provides the great majority of our food. Since the information on responses to drought-stress by species with desiccation tolerant-foliage is sparse compared to the complex situation, findings from non-resurrection species and for other environmental stresses have also been considered to fill gaps in our information.
Fully hydrated flowering-plants, including resurrection plants, are generally desiccation-sensitive; desiccation-tolerance is induced in resurrection plants by drought stress as the plant is drying. In most resurrection plants, induction of desiccation-tolerance in the foliage depends on the foliage being attached to the plant, which indicates that hormonal messages from the drought-stressed root system are necessary for induction. We propose the following phases in the induction of desiccation tolerance in resurrection plants (Figure 1). The phases should not be regarded as discrete compartments since plant responses to drought are a network of progressive interacting processes. Phases overlap and intergrade in the field. For example, Phase 1 processes overlap in time and also in drought stress levels with Phase 2 processes in field plants as diurnal fluctuations in foliage water potential occur throughout the drying. Foliage is stressed during the day, resulting in temporary wilting, but becomes well hydrated during the night. After the soil water potential falls below −1.5 MPa the foliage becomes permanently wilted. These overlapping phases last several days overall, depending on the resurrection species and on environmental circumstances. The values given below are not meant to delimit the phases but only to give a rough idea of their duration and RWC at some points.
Phase 1 Processes: Initial Perception of Drought Stress: Commencing with fully hydrated cells, small reductions in cell water content produce large reductions (often 10 to 30 times atmospheric pressure) in the wall pressure on the cell contents and the cell membrane becomes less stretched. WAK kinases spanning across the cell membrane from the wall to the cytosol register the change in pressure and initiate intracellular signals in the cell. Concurrently, increasing concentration of the cell sap augments the formation of ROS that initiate signal pathways to both restrict further ROS production and to trigger mechanisms contributing to desiccation tolerance. Gupta et al. [221] present a schema at a cell and organelle level for stress-induced triggering and signal transduction.
Phase 2 Processes: Signalling: Drying of soil around even only a part of a root system generally improves the drought resistance of the shoot in flowering-plant species; a mix of hormones rather than a single hormone seem to be involved. Important potential plant-hormones for this messaging in resurrection plants are ABA (in those species accumulating ABA under mild stress), jasmonate and strigolactone and also hormones not yet shown for resurrection plants, acetate (generated in drought-stressed roots from glycolysis products) and CLE25 polypeptide (drought-enhanced synthesis; CLE25 stimulates ABA synthesis in leaves). At the leaf-cell surfaces, hormones combine with specific receptor molecules at the cell membrane or are taken into the cell, where they initiate their specific transduction pathways, which interact together and with other pathways in the leaf cells (see hormone-interaction schema for drying resurrection plants in [45]). Complex changes in the protein complement begin at mild drought stress and continue as stress intensifies. Ismail et al. [41] consider that stress-signalling should be regarded as a message-pulse passing along the signal-pathway, and they stress the importance of timing and crosstalk between the various signalling-pathways. Starch grains disappear as they are hydrolysed into sugars; trehalose may influence starch breakdown and phloem relocation of sugars. Leaves in which senescence has already been triggered, senesce rapidly as they dehydrate and their remobilized minerals, amino acids and sucrose are translocated to younger leaves. In resurrection plants, younger leaves are not subject to drought-induced senescence during appropriate drying. This avoidance or suppression of drought-induced senescence has been proposed as a crucial distinction between resurrection and non-resurrection plants [45]. Much of the mechanisms for cell survival may be in place by the end of Phase 2 in resurrection plants, since leaves detached at mid-phase-3 survive full drying.
Phase 3 Processes: Zero Cell Turgor: When cell water is too low to stretch the cell wall, cell turgor becomes zero and the cell sap solutes concentrate more markedly as cell water potential declines further. Mesomorphic leaves wilt at about this point, after which vascular transport ceases and with it inter-organ signalling stops. Each cell is independently generating desiccation tolerance as drying continues. Glucose and fructose contents decline and sucrose begins to accumulate as drying progresses. Sucrose protects proteins and lipoprotein membranes from degrading. Sucrose signalling presumably commences in this phase. ABA content rises markedly. ABA is a major controller directly and indirectly of metabolism: in the resurrection monocot B. constricta exogenous ABA induces desiccation tolerance in detached hydrated leaves. The ABA signalling pathway interacts with a number of other pathways. A high proportion of major proteins expressed in Phases 1, 2 & 3 are ABA-inducible. ABA-induced protective LEA proteins particularly attain high levels. Protein synthesis increases: increases in the abundance of initiation factor eIF1 may have a key role in stimulating protein synthesis [51,76,222]. Numerous changes occur in the proteome. As drying continues, water contents approach the critical levels at which detached (desiccation-sensitive) leaves would be injured and die (roughly 30–40% RWC). The desiccation tolerance inducing process must be well advanced before these low water levels are reached and surpassed in resurrection plant foliage.
Phase 4 Processes: Nearing Desiccation: In this phase, termed by Zhang and Bartels [42] the ‘Dehydration’ stage (~25% RWC and below), protein synthesis slows. Changes in the proteome at this stage are thought to prepare for metabolic recovery during rehydration, to allow restoration of ‘normal’ metabolism in fully rehydrated plants and eventual resumption of growth. At very low water contents metabolism slows (as metabolite diffusion slows in the increasing viscosity of sucrose-rich cell sap) and appears to cease in the desiccated plants.
Phase 5: Desiccated State: The desiccated state (most commonly 5–8% RWC) is very stable, a situation promoted by trehalose and high sucrose concentrations. In some species dry resurrection plants survive 6 months in the field, but most resurrection species recover if they are rehydrated after up to 2 years storage in the desiccated condition (Figure 1).
Phase 6 Processes: Rehydrating: In good contact with water, resurrection plants usually rehydrate in a shorter time (usually <24 h) than they dehydrate (usually 2 to 6 days after their soil dries). At 20 °C and above, root pressure may assist the rise of water from the roots to the foliage and so hasten leaf rehydration. Vital processes commence at staggered times: respiration recommences within minutes; protein synthesis commences after 4–6 h. In homoiochlorophyllous resurrection plants photosynthesis commences at 5–8 h after watering [31,44,223].
Phase 7 Processes: Post-rehydration: On full rehydration, some vital processes may continue increasing as recovery proceeds. Poikilochlorophyllous resurrection plants are yellow when first fully rehydrated; they commence synthesizing chlorophyll when illuminated; photosynthesis may begin in 30 h and growth recommences [31].
There is a dearth of information on the regulation of the ordered recovery that takes place in Phase 6 and Phase 7.
The extensive alterations in the proteome during the phases 1–5 and the progression from a growth-based state to a quiescent/survival state lead us to the view that changes in epigenetic regulation are involved in the control of some inductive gene expression in most phases. Further studies into epigenetic mechanisms and the genes involved may lead to a greater understanding of extreme desiccation tolerance and allow manipulation of important crop plants to improve yield under adverse environmental conditions.
Author Contributions
The authors have all contributed equally to the discussion, the literature searches and the writing of this review.
Acknowledgments
The authors thank Janet L. Gaff for corrections to the text of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gaff, D.F.; Churchill, D.M. Borya nitida Labill.—An Australian species in the Liliaceae with desiccation-tolerant leaves. Aust. J. Bot. 1976, 24, 209–224. [Google Scholar] [CrossRef]
- Gaff, D.F. Desiccation tolerant vascular plants of Southern Africa. Oecologia 1977, 1, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Gaff, D.F. The Biology of Resurrection Plants; Western Australia Press: Nedlands, Australia, 1981; pp. 114–146. [Google Scholar]
- Gaff, D.F. Desiccation tolerant ‘resurrection’ grasses from Kenya and West Africa. Oecologia 1986, 70, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Gaff, D.F.; Latz, P.K. The occurrence of resurrection plants in the Australian flora. Aust. J. Bot. 1978, 26, 485–492. [Google Scholar] [CrossRef]
- Gaff, D.F.; Bole, P.V. Resurrection grasses in India. Oecologia 1986, 71, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Gaff, D.F.; Ellis, R.P. Southern African grasses with foliage that revives after dehydration. Bothalia Afr. Biodivers. Conserv. 1974, 11, 305–308. [Google Scholar] [CrossRef]
- Iturriaga, G.; Gaff, D.F.; Zentella, R. New desiccation-tolerant plants, including a grass, in the central highlands of Mexico. Aust. J. Bot. 2000, 48, 153–158. [Google Scholar] [CrossRef]
- Gaff, D.F.; Sutaryono, Y.A.; Miszalski, Z. Resurrecting desiccation-tolerant resurrection grasses from dryland areas. Wiad. Bot. 1990, 34, 17–22. [Google Scholar]
- Gaff, D.F.; Oliver, M. The evolution of desiccation tolerance in angiosperm plants: A rare yet common phenomenon. Funct. Plant Biol. 2013, 40, 315–328. [Google Scholar] [CrossRef]
- Farrant, J.; Brandt, W.; Lindsey, G.G. An Overview of Mechanisms of Desiccation Tolerance in Selected Angiosperm Resurrection Plants; Plant Stress: Rondebosch, South Africa, 2007; pp. 72–84. [Google Scholar]
- Angelovici, R.; Galili, G.; Fernie, A.R.; Fait, A. Seed desiccation: A bridge between maturation and germination. Trends Plant Sci. 2010, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Dinakar, C.; Benina, M.; Toneva, V.; Bartels, D. Molecular mechanisms of desiccation tolerance in resurrection plants. Cell. Mol. Life Sci. 2012, 69, 3175–3186. [Google Scholar] [CrossRef] [PubMed]
- Terrasson, E.; Buitink, J.; Righetti, K.; Ly Vu, B.; Pelletier, S.; Lalanne, D.; Zinsmeister, J.; Leprince, O. An emerging picture of the seed desiccome: Confirmed regulators and newcomers identified using transcriptome comparison. Front. Plant Sci. 2013, 4, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrant, J.M.; Cooper, K.; Hilgart, A.; Abdalla, K.O.; Bentley, J.; Thomson, J.A.; Dace, H.J.W.; Peton, N.; Mundree, S.G.; Rafudeen, M.S. A molecular physiological review of vegetative desiccation tolerance in the resurrection plant Xerophyta viscosa (Baker). Planta 2015, 242, 407–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Wang, D.; Yu, B.; Yu, X.; Li, W. Maintenance or collapse: Responses of extraplastidic membrane lipid composition to desiccation in the resurrection plant Paraisometrum mileense. PLoS ONE 2014, 9, e103430. [Google Scholar] [CrossRef] [PubMed]
- Franchi, G.G.; Piotto, B.; Nepi, M.; Baskin, C.C.; Baskin, J.M.; Pacini, E. Pollen and seed desiccation tolerance in relation to degree of developmental arrest, dispersal, and survival. J. Exp. Bot. 2011, 62, 5267–5281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyres, I.; Boschetti, C.; Crisp, A.; Smith, T.P.; Fontaneto, D.; Tunnacliffe, A.; Barraclough, T.G. Horizontal gene transfer in Bdelloid rotifers is ancient, ongoing and more frequent in species from desiccating habitats. BMC Biol. 2015, 13, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilbricht, T.; Varotto, S.; Sgaramella, V.; Bartels, D.; Salamini, F.; Furini, A. Retrotransposons and siRNA have a role in the evolution of desiccation tolerance leading to resurrection of the plant Craterostigma plantagineum. New Phytol. 2008, 179, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Royal Botanic Gardens, Kew. Seed Information Database (SID), version 7.1; Royal Botanic Gardens, Kew: Richmond, UK, 2008. [Google Scholar]
- Holman, R.M.; Brubaker, F. On the Longevity of Pollen; University of California Press: Oakland, CA, USA, 1926. [Google Scholar]
- Visser, T. Germination and Storage of Pollen. Ph.D. Thesis, Wageningen University & Research, Wageningen, The Netherlands, 1955. [Google Scholar]
- Evenari, M.; Shanan, L.; Tadmor, N. The Negev. The Challenge of a Desert; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- McFadden, G.I. Origin and evolution of plastids and photosynthesis in eukaryotes. Cold Spring Harb. Perspect. Biol. 2014, 6, a016105. [Google Scholar] [CrossRef] [PubMed]
- Gribaldo, S.; Poole, A.M.; Daubin, V.; Forterre, P.; Brochier-Armanet, C. The origin of eukaryotes and their relationship with the Archaea: Are we at a phylogenomic impasse? Nat. Rev. Microbiol. 2010, 8, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Rochette, N.C.; Brochier-Armanet, C.; Gouy, M. Phylogenomic test of the hypotheses for the evolutionary origin of eukaryotes. Mol. Biol. Evol. 2014, 31, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.L.; Apolinario, E.E.; Sowers, K.R. Desiccation as a long-term survival mechanism for the Archaeon Methanosarcina barkeri. Appl. Environ. Microbiol. 2012, 78, 1473–1479. [Google Scholar] [CrossRef] [PubMed]
- Billi, D.; Potts, M. Life and death of dried prokaryotes. Res. Microbiol. 2002, 153, 7–12. [Google Scholar] [CrossRef]
- Garcia, A.H. Anhydrobiosis in bacteria: From physiology to applications. J. Biosci. 2011, 36, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Stoll, M.; Loveys, B.; Dry, P. Hormonal changes induced by partial rootzone drying of irrigated grapevine. J. Exp. Bot. 2000, 51, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Gaff, D.F. Protoplasmic tolerance of extreme stress. In Adaptation of Plants to Water and High Temperature Stress; Turner, N.C., Kramer, P.J., Eds.; John Wiley & Sons: New York, NY, USA, 1980; pp. 207–230. [Google Scholar]
- Hsiao, T.C.; Acevedo, E.; Fereres, E.; Henderson, D.W. Water stress, growth and osmotic adjustment. Philos. Trans. R. Soc. Lond. B 1976, 273, 479–500. [Google Scholar] [CrossRef]
- Sperry, J.S.; Tyree, M.T. Mechanism of water stress-induced xylem embolism. Plant Physiol. 1988, 88, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Christmann, A.; Weiler, E.W.; Steudle, E.; Grill, E. A hydraulic signal in root-to-shoot signalling of water shortage. Plant J. 2007, 52, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaff, D.F.; Loveys, B.R. Abscisic acid levels in drying plants of a resurrection grass. Trans. Malays. Soc. Plant Physiol. 1993, 3, 286–287. [Google Scholar]
- Blomstedt, C.K.; Griffiths, C.A.; Fredericks, D.P.; Hamill, J.D.; Gaff, D.F.; Neale, A.D. The resurrection plant Sporobolus stapfianus: An unlikely model for engineering enhanced plant biomass? Plant Growth Regul. 2010, 62, 217–232. [Google Scholar] [CrossRef]
- Giarola, V.; Krey, S.; Driesch, B.; Bartels, D. The Craterostigma plantagineum glycine-rich protein CpGRP1 interacts with a cell wall-associated protein kinase 1 (CpWAK1) and accumulates in leaf cell walls during dehydration. New Phytol. 2016, 210, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.S.; Merkle, M.G.; Bovey, R.W. Effect of moisture stress on the absorption and transport of herbicides in woody plants. Bot. Gaz. 1968, 129, 183–189. [Google Scholar] [CrossRef]
- Dinant, S.; Lemoine, R. The phloem pathway: New issues and old debates. C. R. Biol. 2010, 333, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.; Gaff, D.; Gianello, R.; Blomstedt, C.; Neale, A.; Hamill, J. Changes in in vivo protein complements in drying leaves of the desiccation-tolerant grass Sporobolus stapfianus and the desiccation-sensitive grass Sporobolus pyramidalis. Funct. Plant Biol. 1995, 22, 1027–1034. [Google Scholar]
- Ismail, A.; Takeda, S.; Nick, P. Life and death under salt stress: Same players, different timing? J. Exp. Bot. 2014, 65, 2963–2979. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Bartels, D. Molecular responses to dehydration and desiccation in desiccation-tolerant angiosperm plants. J. Exp. Bot. 2018, 69, 3211–3222. [Google Scholar] [CrossRef] [PubMed]
- Bastien, C.; Aurélie, E.; Iris, S.; Bernhard, K.; Dorothea, B.; Shaun, P.; Stefan, H. Water deficit induces chlorophyll degradation via the ‘pao/phyllobilin’ pathway in leaves of homoio-(Craterostigma pumilum) and poikilochlorophyllous (Xerophyta viscosa) resurrection plants. Plant Cell Environ. 2014, 37, 2521–2531. [Google Scholar]
- Gaff, D.F.; Loveys, B.R. Abscisic acid content and effects during dehydration of detached leaves of desiccation tolerant plants. J. Exp. Bot. 1984, 35, 1350–1358. [Google Scholar] [CrossRef]
- Griffiths, C.; Gaff, D.; Neale, A. Drying without senescence in resurrection plants. Front. Plant Sci. 2014, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Ghasempour, H.R.; Anderson, E.M.; Gaff, D.F. Effects of growth substances on the protoplasmic drought tolerance of leaf cells of the resurrection grass Sporobolus stapfianus. Aust. J. Plant Physiol. 2001, 28, 1115–1120. [Google Scholar]
- Wright, S.T.C. Seasonal changes in the levels of free and bound abscisic acid in blackcurrant (Ribes nigrum) buds and beech (Fagus sylvatica) buds. J. Exp. Bot. 1975, 26, 161–174. [Google Scholar] [CrossRef]
- Bano, A.; Hansen, H.; Dörffling, K.; Hahn, H. Changes in the contents of free and conjugated abscisic acid, phaseic acid and cytokinins in xylem sap of drought stressed sunflower plants. Phytochemistry 1994, 37, 345–347. [Google Scholar] [CrossRef]
- Gaff, D.F.; McGregor, G.R. The effect of dehydration and re-hydration on the nitrogen content of various fractions from resurrection plants. Biol. Plant. 1979, 21, 92–99. [Google Scholar] [CrossRef]
- Bartels, D.; Schneider, K.; Terstappen, G.; Piatowski, D.F.S. Molecular cloning of abscisic acid-modulated genes which are induced during desiccation of the resurrection plant Craterostigma plantagineum. Planta 1990, 181, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Neale, A.; Blomstedt, C.; Bronson, P.; Le, T.N.; Guthridge, K.; Evans, J.; Gaff, D.; Hamill, J. The isolation of genes from the resurrection grass Sporobolus stapfianus which are induced during severe drought stress. Plant Cell Environ. 2000, 23, 265–277. [Google Scholar] [CrossRef]
- Huang, J.-C.; Lin, S.-M.; Wang, C.-S. A pollen-specific and desiccation-associated transcript in Lilium longiflorum during development and stress. Plant Cell Physiol. 2000, 41, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-Y.; Chen, Y.-C.; Jauh, G.Y.; Wang, C.-S. A lily ASR protein involves abscisic acid signaling and confers drought and salt resistance in Arabidopsis. Plant Physiol. 2005, 139, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Meurs, C.; Basra, A.S.; Karssen, C.M.; van Loon, L.C. Role of abscisic acid in the induction of desiccation tolerance in developing seeds of Arabidopsis thaliana. Plant Physiol. 1992, 98, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Rushton, D.L.; Tripathi, P.; Rabara, R.C.; Lin, J.; Ringler, P.; Boken, A.K.; Langum, T.J.; Smidt, L.; Boomsma, D.D.; Emme, N.J.; et al. WRKY transcription factors: Key components in abscisic acid signalling. Plant Biotechnol. J. 2012, 10, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.; Dupeux, F.; Betz, K.; Antoni, R.; Gonzalez-Guzman, M.; Rodriguez, L.; Márquez, J.A.; Rodriguez, P.L. Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs. Plant Sci. 2012, 182, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.-Q.; Xin, Q.; Cao, Z.; Liu, Z.-Q.; Du, S.-Y.; Mei, C.; Zhao, C.-X.; Wang, X.-F.; Shang, Y.; Jiang, T.; et al. The magnesium-chelatase h subunit binds abscisic acid and functions in abscisic acid signaling: New evidence in Arabidopsis. Plant Physiol. 2009, 150, 1940–1954. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Hu, J.; Guo, S.; Wang, J.; Cheng, Y.; Dang, X.; Wu, L.; He, Y. Proteome analysis of Physcomitrella patens exposed to progressive dehydration and rehydration. J. Exp. Bot. 2012, 63, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.C.D.; Farrant, J.M.; Oliver, M.J.; Ligterink, W.; Buitink, J.; Hilhorst, H.M.W. Key genes involved in desiccation tolerance and dormancy across life forms. Plant Sci. 2016, 251, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.; Gampala, S.S.; Lynch, T.J.; Thomas, T.L.; Rock, C.D. Redundant and distinct functions of the ABA response loci ABA-insensitive (ABI) 5 and abre-binding factor (ABF) 3. Plant Mol. Biol. 2005, 59, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Hand, S.C.; Menze, M.A.; Toner, M.; Boswell, L.; Moore, D. LEA proteins during water stress: Not just for plants anymore. Annu. Rev. Physiol. 2011, 73, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Kishitani, S.; Ito, Y.; Toriyama, K. Accumulation of raffinose in rice seedlings overexpressing OsWRKY11 in relation to desiccation tolerance. Plant Biotechnol. 2009, 26, 431–434. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, Y.; Wang, L.; Liu, X.; Liu, Y.; Phillips, J.; Deng, X. A WRKY transcription factor participates in dehydration tolerance in Boea hygrometrica by binding to the W-box elements of the galactinol synthase (BhGOLS1) promoter. Planta 2009, 230, 1155. [Google Scholar] [CrossRef] [PubMed]
- Ghasempour, H.R.; Gaff, D.F.; Williams, R.P.W.; Gianello, R.D. Contents of sugars in leaves of drying desiccation tolerant flowering plants, particularly grasses. Plant Growth Regul. 1998, 24, 185–191. [Google Scholar] [CrossRef]
- Maia, J.; Dekkers, B.J.W.; Provart, N.J.; Ligterink, W.; Hilhorst, H.W.M. The re-establishment of desiccation tolerance in germinated Arabidopsis thaliana seeds and its associated transcriptome. PLoS ONE 2011, 6, e29123. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, M.; Xiong, L.; Stevenson, B.; Zhu, J.-K. Genetic analysis of osmotic and cold stress signal transduction in Arabidopsis: Interactions and convergence of abscisic acid-dependent and abscisic acid-independent pathways. Plant Cell 1997, 9, 1935–1949. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Kiyosue, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K.A. nuclear gene, erd1, encoding a chloroplast-targeted clp protease regulatory subunit homolog is not only induced by water stress but also developmentally up-regulated during senescence in Arabidopsis thaliana. Plant J. 1997, 12, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Levitt, J. The Hardiness of Plants; Academic Press Inc.: New York, NY, USA, 1956. [Google Scholar]
- Dhawan, S.S.; Sharma, A. Analysis of differentially expressed genes in abiotic stress response and their role in signal transduction pathways. Protoplasma 2014, 251, 81–91. [Google Scholar]
- Islam, S.; Griffiths, C.A.; Blomstedt, C.K.; Le, T.-N.; Gaff, D.F.; Hamill, J.D.; Neale, A.D. Increased biomass, seed yield and stress tolerance is conferred in Arabidopsis by a novel enzyme from the resurrection grass Sporobolus stapfianus that glycosylates the strigolactone analogue GR24. PLoS ONE 2013, 8, e80035. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anandarajah, K.; Kott, L.; Beversdorf, W.D.; McKersie, B.D. Induction of desiccation tolerance in microspore-derived embryos of Brassica napus L. By thermal stress. Plant Sci. 1991, 77, 119–123. [Google Scholar] [CrossRef]
- Oliver, M.J.; Jain, R.; Balbuena, T.S.; Agrawal, G.; Gasulla, F.; Thelen, J.J. Proteome analysis of leaves of the desiccation-tolerant grass, Sporobolus stapfianus, in response to dehydration. Phytochemistry 2011, 72, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Peleg, Z.; Reguera, M.; Tumimbang, E.; Walia, H.; Blumwald, E. Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol. J. 2011, 9, 747–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walia, H.; Wilson, C.; Wahid, A.; Condamine, P.; Cui, X.; Close, T.J. Expression analysis of barley (Hordeum vulgare L.) during salinity stress. Funct. Integr. Genom. 2006, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bai, M.-Y.; Chong, K. Brassinosteroid-mediated regulation of agronomic traits in rice. Plant Cell Rep. 2014, 33, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, C.V.; Leyva-González, M.A.; Osakabe, Y.; Tran, U.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Yamaguchi, S.; Dong, N.V.; et al. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. USA 2014, 111, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Bu, Q.; Lv, T.; Shen, H.; Luong, P.; Wang, J.; Wang, Z.; Huang, Z.; Xiao, L.; Engineer, C.; Kim, T.H.; et al. Regular of drought tolerance by the F-box protein max2 in Arabidopsis. Plant Physiol. 2014, 164, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, H.; Vitali, M.; Visentin, I.; Charnikhova, T.; Haider, I.; Schubert, A.; Ruyter-Spira, C.; Bouwmeester, H.J.; Lovisolo, C.; et al. Osmotic stress represses strigolactone biosynthesis in Lotus japonicus roots: Exploring the interaction between strigolactones and ABA under abiotic stress. Planta 2015, 241, 1435–1451. [Google Scholar] [CrossRef] [PubMed]
- Visentin, I.; Vitali, M.; Ferrero, M.; Zhang, Y.; Ruyter-Spira, C.; Novák, O.; Strnad, M.; Lovisolo, C.; Schubert, A.; Cardinale, F. Low levels of strigolactones in roots as a component of the systemic signal of drought stress in tomato. New Phytol. 2016, 212, 954–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Lozano, J.M.; Aroca, R.; Zamarreño, A.M.; Molina, S.; Andreo-Jiméne, B.; Porcel, R.; García-Mina, J.M.; Ruyter-Spira, C.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis induces strigolactone biosynthesis under drought and improves drought tolerance in lettuce and tomato. Plant Cell Environ. 2016, 39, 441–452. [Google Scholar] [CrossRef] [PubMed]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Twito, S.; Miller, G. New cross talk between ROS, ABA and auxin controlling seed maturation and germination unraveled in APX6 deficient Arabidopsis seeds. Plant Signal. Behav. 2014, 9, e976489. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.-R.; Marriott, A.S.; Bergström, E.; et al. Molecular mechanisms of desiccation tolerance in the resurrection glacial relic Haberlea rhodopensis. Cell. Mol. Life Sci. 2013, 70, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic profiling of the resurrection plant Haberlea rhodopensis during desiccation and recovery. Physiol. Plant. 2014, 152, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, B.; Phillips, J.; Zhang, Z.-N.; Du, H.; Xu, T.; Huang, L.-C.; Zhang, X.-F.; Xu, G.-H.; Li, W.-L.; et al. Global transcriptome analysis reveals acclimation-primed processes involved in the acquisition of desiccation tolerance in Boea hygrometrica. Plant Cell Physiol. 2015, 56, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Beckett, M.; Loreto, F.; Velikova, V.; Brunetti, C.; DiFerdinando, M.; Tattini, M.; Calfapietra, C.; Farrant, J.M. Photosynthetic limitations and volatile and non-volatile isoprenoids in the poikilochlorophyllous resurrection plant Xerophyta humilis during dehydration and rehydration. Plant Cell Environ. 2012, 35, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Koussevitzky, S.; Suzuki, N.; Huntington, S.; Armijo, L.; Sha, W.; Cortes, D.; Shulaev, V.; Mittler, R. Ascorbate peroxidase 1 plays a key role in the response of Arabidopsis thaliana to stress combination. J. Biol. Chem. 2008, 283, 34197–34203. [Google Scholar] [CrossRef] [PubMed]
- Prasch, C.M.; Sonnewald, U. Simultaneous application of heat, drought, and virus to Arabidopsis plants reveals significant shifts in signaling networks. Plant Physiol. 2013, 162, 1849–1866. [Google Scholar] [CrossRef] [PubMed]
- Tahir, J.; Dijkwel, P. B-substituting alanine synthases: Roles in cysteine metabolism and abiotic and biotic stress signalling in plants. Funct. Plant Biol. 2016, 43, 307–323. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lelandais, M.; Kunert, K.J. Photooxidative stress in plants. Physiol. Plant. 1994, 92, 696–717. [Google Scholar] [CrossRef]
- Cruz de Carvalho, R.; Bernardes Da Silva, A.; Soares, R.; Almeida, A.M.; Coelho, A.V.; Marques Da Silva, J.; Branquinho, C. Differential proteomics of dehydration and rehydration in bryophytes: Evidence towards a common desiccation tolerance mechanism. Plant Cell Environ. 2014, 37, 1499–1515. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; To, T.K.; Matsui, A.; Tanoi, K.; Kobayashi, N.I.; Matsuda, F.; Habu, Y.; Ogawa, D.; Sakamoto, T.; Matsunaga, S. Acetate-mediated novel survival strategy against drought in plants. Nat. Plants 2017, 3, 17097. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M. Studies of Sugars in Relation to Induction of Desiccation Tolerance in Resurrection Grass Sporobolus stapfianus. Ph.D. Thesis, Monash University, Clayton, Australia, 2001. [Google Scholar]
- Bourquin, S.; Bonnemain, J.-L.; Delrot, S. Inhibition of loading of 14C assimilates by p-chloromercuribenzenesulfonic acid: Localization of the apoplastic pathway in Vicia faba. Plant Physiol. 1990, 92, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Qu, H.; Dibley, K.E.; Offler, C.E.; Patrick, J.W. A suite of sucrose transporters expressed in coats of developing legume seeds includes novel pH-independent facilitators. Plant J. 2007, 49, 750–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chincinska, I.; Gier, K.; Krügel, U.; Liesche, J.; He, H.; Grimm, B.; Harren, F.J.M.; Cristescu, S.M.; Kühn, C. Photoperiodic regulation of the sucrose transporter StSUT4 affects the expression of circadian-regulated genes and ethylene production. Front. Plant Sci. 2013, 4, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibraheem, O.; Botha, C.E.J.; Bradley, G. In silico analysis of cis-acting regulatory elements in 5′ regulatory regions of sucrose transporter gene families in rice (Oryza sativa Japonica) and Arabidopsis thaliana. Comput. Biol. Chem. 2010, 34, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Smeekens, S.; Hellmann, H.A. Sugar sensing and signaling in plants. Front. Plant Sci. 2014, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Dobrenel, T.; Marchive, C.; Azzopardi, M.; Clément, G.; Moreau, M.; Sormani, R.; Robaglia, C.; Meyer, C. Sugar metabolism and the plant target of rapamycin kinase: A sweet operator? Front. Plant Sci. 2013, 4, 93. [Google Scholar] [CrossRef] [PubMed]
- Yobi, A.; Wone, B.W.; Xu, W.; Alexander, D.C.; Guo, L.; Ryals, J.A.; Oliver, M.J.; Cushman, J.C. Comparative metabolic profiling between desiccation-sensitive and desiccation-tolerant species of Selaginella reveals insights into the resurrection trait. Plant J. 2012, 72, 983–999. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R.; Fischer, E.; Baron, M.; Van Den Dries, N.; Facchinelli, F.; Kutzer, M.; Rahmanzadeh, R.; Remus, D.; Bartels, D. Lindernia brevidens: A novel desiccation-tolerant vascular plant, endemic to ancient tropical rainforests. Plant J. 2008, 54, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, G.; Gamba, A.; Murelli, C.; Salamini, F.; Bartels, D. Novel carbohydrate metabolism in the resurrection plant Craterostigma plantagineum. Plant J. 1991, 1, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.; Bellés, J.M.; Vayá, J.L.; Serrano, R.; Culiáñez-Macià, F.A. Expression of the yeast trehalose-6-phosphate synthase gene in transgenic tobacco plants: Pleiotropic phenotypes include drought tolerance. Planta 1997, 201, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.K.; Kim, J.-K.; Owens, T.G.; Ranwala, A.P.; Choi, Y.D.; Kochian, L.V.; Wu, R.J. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grennan, A.K. The role of trehalose biosynthesis in plants. Plant Physiol. 2007, 144, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A. The function of trehalose biosynthesis in plants. Phytochemistry 2002, 60, 437–440. [Google Scholar] [CrossRef]
- Muller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C. content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [PubMed]
- Chiou, T.-J.; Bush, D.R. Sucrose is a signal molecule in assimilate partitioning. Proc. Natl. Acad. Sci. USA 1998, 95, 4784–4788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, U.P.; Ivakov, A.; Feil, R.; Duan, G.Y.; Walther, D.; Giavalisco, P.; Piques, M.; Carillo, P.; Hubberten, H.-M.; Stitt, M.; et al. The sucrose–trehalose 6-phosphate (Tre6P) nexus: Specificity and mechanisms of sucrose signalling by Tre6P. J. Exp. Bot. 2014, 65, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Primavesi, L.F.; Jhurreea, D.; Andralojc, P.J.; Mitchell, R.A.; Powers, S.J.; Schluepmann, H.; Delatte, T.; Wingler, A.; Paul, M.J. Inhibition of SNF1-related protein kinase1 activity and regulation of metabolic pathways by trehalose-6-phosphate. Plant Physiol. 2009, 149, 1860–1871. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.A.; Sagar, R.; Geng, Y.; Primavesi, L.F.; Patel, M.K.; Passarelli, M.K.; Gilmore, I.S.; Steven, R.T.; Bunch, J.; Paul, M.J. Chemical intervention in plant sugar signalling increases yield and resilience. Nature 2016, 540, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.A.; Paul, M.J.; Foyer, C.H. Metabolite transport and associated sugar signalling systems underpinning source/sink interactions. Biochim. Biophys. Acta (BBA)-Bioenerg. 2016, 1857, 1715–1725. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; O’Hara, L.E.; Primavesi, L.F.; Delatte, T.L.; Schluepmann, H.; Somsen, G.W.; Silva, A.B.; Fevereiro, P.S.; Wingler, A.; Paul, M.J. The trehalose 6-phosphate/SnRK1 signaling pathway primes growth recovery following relief of sink limitation. Plant Physiol. 2013, 162, 1720–1732. [Google Scholar] [CrossRef] [PubMed]
- Nuccio, M.L.; Wu, J.; Mowers, R.; Zhou, H.P.; Meghji, M.; Primavesi, L.F.; Paul, M.J.; Chen, X.; Gao, Y.; Haque, E.; et al. Expression of trehalose-6-phosphate phosphatase in maize ears improves yield in well-watered and drought conditions. Nat. Biotechnol. 2015, 33, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Schluepmann, H.; Pellny, T.; van Dijken, A.; Smeekens, S.; Paul, M. Trehalose 6-phosphate is indispensable for carbohydrate utilization and growth in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6849–6854. [Google Scholar] [CrossRef] [PubMed]
- Lunn, J.E.; Feil, R.; Hendriks, J.H.; Gibon, Y.; Morcuende, R.; Osuna, D.; Scheible, W.R.; Carillo, P.; Hajirezaei, M.R.; Stitt, M. Sugar-induced increases in trehalose 6-phosphate are correlated with redox activation of ADPglucose pyrophosphorylase and higher rates of starch synthesis in Arabidopsis thaliana. Biochem. J. 2006, 397, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Hallam, N.D.; Gaff, D.F. Re-organization of fine structure during rehydration of desiccated leaves of Xerophyta villosa. New Phytol. 1978, 81, 349–355. [Google Scholar] [CrossRef]
- Hallam, N.D.; Gaff, D.F. Regeneration of chloroplast structure in Talbotia elegans: A desiccation-tolerant plant. New Phytol. 1978, 81, 657–662. [Google Scholar] [CrossRef]
- Eastmond, P.J.; van Dijken, A.J.; Spielman, M.; Kerr, A.; Tissier, A.F.; Dickinson, H.G.; Jones, J.D.; Smeekens, S.C.; Graham, I.A. Trehalose-6-phosphate synthase 1, which catalyses the first step in trehalose synthesis, is essential for Arabidopsis embryo maturation. Plant J. 2002, 29, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Gómez, L.D.; Gilday, A.; Feil, R.; Lunn, J.E.; Graham, I.A. AtTPS1-mediated trehalose 6-phosphate synthesis is essential for embryogenic and vegetative growth and responsiveness to aba in germinating seeds and stomatal guard cells. Plant J. 2010, 64, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Avonce, N.; Leyman, B.; Mascorro-Gallardo, J.O.; Van Dijck, P.; Thevelein, J.M.; Iturriaga, G. The Arabidopsis trehalose-6-p synthase AtTPS1 gene is a regulator of glucose, abscisic acid, and stress signaling. Plant Physiol. 2004, 136, 3649–3659. [Google Scholar] [CrossRef] [PubMed]
- Koster, K.L. Glass formation and desiccation tolerance in seeds. Plant Physiol. 1991, 96, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, E.A.; Terry, N.; Sears, T.; Kim, H.; Zayed, A.; Hwang, S.; van Dun, K.; Voogd, E.; Verwoerd, T.C.; Krutwagen, R.W. Trehalose-producing transgenic tobacco plants show improved growth performance under drought stress. J. Plant Physiol. 1998, 152, 525–532. [Google Scholar] [CrossRef]
- Magazù, S.; Migliardo, F.; Gonzalez, M.; Mondelli, C.; Parker, S.; Vertessy, B. Molecular mechanisms of survival strategies in extreme conditions. Life 2012, 2, 364. [Google Scholar] [CrossRef] [PubMed]
- Crowe, J.H.; Carpenter, J.F.; Crowe, L.M. The role of vitrification in anhydrobiosis. Ann. Rev. Physiol. 1998, 60, 73–103. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.J.; Oliver, M.J. Translational control in plant stress: The formation of messenger ribonucleoprotein particles (mRNPs) in response to desiccation of Tortula ruralis gametophytes. Plant J. 1999, 18, 359–370. [Google Scholar] [CrossRef]
- Tymms, M.J.; Gaff, D.F.; Hallam, N.D. Protein synthesis in the desiccation tolerant angiosperm Xerophyta villosa during dehydration. J. Exp. Bot. 1982, 33, 332–343. [Google Scholar] [CrossRef]
- Blomstedt, C.K.; Gianello, R.D.; Gaff, D.F.; Hamill, J.D.; Neale, A.D. Differential gene expression in desiccation-tolerant and desiccation-sensitive tissue of the resurrection grass, Sporobolus stapfianus. Funct. Plant Biol. 1998, 25, 937–946. [Google Scholar] [CrossRef]
- Le, T.N.; Blomstedt, C.K.; Kuang, J.; Tenlen, J.; Gaff, D.F.; Hamill, J.D.; Neale, A.D. Desiccation-tolerance specific gene expression in leaf tissue of the resurrection plant Sporobolus stapfianus. Funct. Plant Biol. 2007, 34, 589–600. [Google Scholar] [CrossRef]
- Daniel, V.; Gaff, D.F. Dessication-induced changes in the protein complement of soluble extracts from leaves of resurrection plants and related desiccation-sensitive species. Annu. Bot. 1980, 45, 173–181. [Google Scholar] [CrossRef]
- Gaff, D.F.; Ziegler, H. ATP and ADP contents in leaves of drying and rehydrating desiccation tolerant plants. Oecologia 1989, 78, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Leprince, O.; Buitink, J. Desiccation tolerance: From genomics to the field. Plant Sci. 2010, 179, 554–564. [Google Scholar] [CrossRef] [Green Version]
- Chiu, R.S.; Nahal, H.; Provart, N.J.; Gazzarrini, S. The role of the Arabidopsis FUSCA3 transcription factor during inhibition of seed germination at high temperature. BMC Plant Biol. 2012, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.-P.; Way, H.M.; Richardson, T.; Drenth, J.; Joyce, P.A.; McIntyre, C.L. Overexpression of TaNAC69 leads to enhanced transcript levels of stress up-regulated genes and dehydration tolerance in bread wheat. Mol. Plant 2011, 4, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Zia, A.; Walker, B.J.; Oung, H.M.O.; Charuvi, D.; Jahns, P.; Cousins, A.B.; Farrant, J.M.; Reich, Z.; Kirchhoff, H. Protection of the photosynthetic apparatus against dehydration stress in the resurrection plant Craterostigma pumilum. Plant J. 2016, 87, 664–680. [Google Scholar] [CrossRef] [PubMed]
- Piatkowski, D.; Schneider, K.; Salamini, F.; Bartels, D. Characterization of five abscisic acid-responsive cDNA clones isolated from the desiccation-tolerant plant Craterostigma plantagineum and their relationship to other water-stress genes. Plant Physiol. 1990, 94, 1682–1688. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.; Bartels, D. The molecular basis of dehydration tolerance in plants. Annu. Rev. Plant Biol. 1996, 47, 377–403. [Google Scholar] [CrossRef] [PubMed]
- Mariaux, J.-B.; Bockel, C.; Salamini, F.; Bartels, D. Desiccation-and abscisic acid-responsive genes encoding major intrinsic proteins (MIPs) from the resurrection plant Craterostigma plantagineum. Plant Mol. Biol. 1998, 38, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Salamini, F. Desiccation tolerance in the resurrection plant Craterostigma plantagineum. A contribution to the study of drought tolerance at the molecular level. Plant Physiol. 2001, 127, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Garwe, D.; Thomson, J.A.; Mundree, S.G. Molecular characterization of XvSAP1, a stress-responsive gene from the resurrection plant Xerophyta viscosa Baker. J. Exp. Bot. 2003, 54, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Garwe, D.; Thomson, J.A.; Mundree, S.G. Xvsap1 from Xerophyta viscosa improves osmotic-, salinity-and high-temperature-stress tolerance in Arabidopsis. Biotechnol. J. 2006, 1, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Ndima, T.; Farrant, J.; Thomson, J.; Mundree, S. Molecular characterization of XvT8, a stress-responsive gene from the resurrection plant Xerophyta viscosa Baker. Plant Growth Regul. 2001, 35, 137–145. [Google Scholar] [CrossRef]
- Ramanjulu, S.; Bartels, D. Drought-and desiccation-induced modulation of gene expression in plants. Plant Cell Environ. 2002, 25, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Van den Dries, N.; Facchinelli, F.; Giarola, V.; Phillips, J.R.; Bartels, D. Comparative analysis of LEA-like 11-24 gene expression and regulation in related plant species within the Linderniaceae that differ in desiccation tolerance. New Phytol. 2011, 190, 75–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Shang, H.; Liu, Y.; Zheng, M.; Wu, R.; Phillips, J.; Bartels, D.; Deng, X. A role for a cell wall localized glycine-rich protein in dehydration and rehydration of the resurrection plant Boea hygrometrica. Plant Biol. 2009, 11, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortee, S.; Boschetti, C.; Walton, L.J.; Sarkar, S.; Rubinsztein, D.C.; Tunnacliffe, A. Hydrophilic protein associated with desiccation tolerance exhibits broad protein stabilization function. Proc. Natl. Acad. Sci. USA 2007, 104, 18073–18078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Close, T.J. Dehydrins: A commonalty in the response of plants to dehydration and low temperature. Physiol. Plant. 1997, 100, 291–296. [Google Scholar] [CrossRef]
- Dure, L. A repeating 11-mer amino acid motif and plant desiccation. Plant J. 1993, 3, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasulla, F.; vom Dorp, K.; Dombrink, I.; Zähringer, U.; Gisch, N.; Dörmann, P.; Bartels, D. The role of lipid metabolism in the acquisition of desiccation tolerance in Craterostigma plantagineum: A comparative approach. Plant J. 2013, 75, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.-C.D.; Artur, M.A.; Maia, J.; Jonkheer, E.; Derks, M.F.; Nijveen, H.; Williams, B.; Mundree, S.G.; Jiménez-Gómez, J.M.; Hesselink, T. A footprint of desiccation tolerance in the genome of Xerophyta viscosa. Nat. Plants 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Giarola, V.; Bartels, D. What can we learn from the transcriptome of the resurrection plant Craterostigma plantagineum? Planta 2015, 242, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Giarola, V.; Hou, Q.; Bartels, D. Angiosperm plant desiccation tolerance: Hints from transcriptomics and genome sequencing. Trends Plant Sci. 2017, 22, 705–717. [Google Scholar] [CrossRef] [PubMed]
- VanBuren, R.; Bryant, D.; Edger, P.P.; Tang, H.; Burgess, D.; Challabathula, D.; Spittle, K.; Hall, R.; Gu, J.; Lyons, E. Single-molecule sequencing of the desiccation-tolerant grass Oropetium thomaeum. Nature 2015, 527, 508. [Google Scholar] [CrossRef] [PubMed]
- VanBuren, R.; Wai, C.M.; Zhang, Q.; Song, X.; Edger, P.P.; Bryant, D.; Michael, T.P.; Mockler, T.C.; Bartels, D. Seed desiccation mechanisms co-opted for vegetative desiccation in the resurrection grass Oropetium thomaeum. Plant Cell Environ. 2017, 40, 2292–2306. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, K.; Bi, D.; Zhou, S.; Shao, J. Characterization of the transcriptome and EST-SSR development in Boea clarkeana, a desiccation-tolerant plant endemic to China. PeerJ 2017, 5, e3422. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Yang, G.; Zhang, L.; Yang, X.; Zhao, S.; Ji, Z.; Zhou, Q.; Hu, M.; Wang, Y.; Chen, M.; et al. The resurrection genome of Boea hygrometrica: A blueprint for survival of dehydration. Proc. Natl. Acad. Sci. USA 2015, 112, 5833–5837. [Google Scholar] [CrossRef] [PubMed]
- Farrant, J.M.; Moore, J.P. Programming desiccation-tolerance: From plants to seeds to resurrection plants. Curr. Opin. Plant Biol. 2011, 14, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Frank, W.; Phillips, J.; Salamini, F.; Bartels, D. Two dehydration-inducible transcripts from the resurrection plant Craterostigma plantagineum encode interacting homeodomain-leucine zipper proteins. Plant J. 1998, 15, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Villalobos, M.A.; Bartels, D.; Iturriaga, G. Stress tolerance and glucose insensitive phenotypes in Arabidopsis overexpressing the CpMYB10 transcription factor gene. Plant Physiol. 2004, 135, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Giarola, V.; Jung, N.U.; Singh, A.; Satpathy, P.; Bartels, D. Analysis of pcc13-62 promoters predicts a link between cis-element variations and desiccation tolerance in Linderniaceae. J. Exp. Bot. 2018, 69, 3773–3784. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.L.; Blomstedt, C.K.; Neale, A.D.; Gleadow, R.; DeBoer, K.D.; Hamill, J.D. Effects of down-regulating ornithine decarboxylase upon putrescine-associated metabolism and growth in Nicotiana tabacum L. J. Exp. Bot. 2016, 67, 3367–3381. [Google Scholar] [CrossRef] [PubMed]
- Furini, A.; Koncz, C.; Salamini, F.; Bartels, D. Agrobacterium-mediated transformation of the desiccation-tolerant plant Craterostigma plantagineum. Plant Cell Rep. 1994, 14, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Smith-Espinoza, C.; Bartels, D.; Phillips, J. Analysis of a LEA gene promoter via agrobacterium-mediated transformation of the desiccation tolerant plant Lindernia brevidens. Plant Cell Rep. 2007, 26, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y. Efficient DNA-free genome editing of bread wheat using CRISPR/Cas9 ribonucleoprotein complexes. Nat. Commun. 2017, 8, 14261. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Chen, K.; Yan, Y.; Zhang, Y.; Gao, C. Genotyping genome-edited mutations in plants using CRISPR ribonucleoprotein complexes. Plant Biotechnol. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeted screening for induced mutations. Nat. Biotechnol. 2000, 18, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Till, B.J.; Cooper, J.; Tai, T.H.; Colowit, P.; Greene, E.A.; Henikoff, S.; Comai, L. Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol. 2007, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Blomstedt, C.K.; Gleadow, R.M.; O’Donnell, N.; Naur, P.; Jensen, K.; Laursen, T.; Olsen, C.E.; Stuart, P.; Hamill, J.D.; Møller, B.L. A combined biochemical screen and TILLING approach identifies mutations in Sorghum bicolor L. Moench resulting in acyanogenic forage production. Plant Biotechnol. J. 2012, 10, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Blomstedt, C.K.; O’Donnell, N.H.; Bjarnholt, N.; Neale, A.D.; Hamill, J.D.; Møller, B.L.; Gleadow, R.M. Metabolic consequences of knocking out UGT85B1, the gene encoding the glucosyltransferase required for synthesis of dhurrin in Sorghum bicolor (L. Moench). Plant Cell Physiol. 2016, 57, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, G.; Schneider, K.; Salamini, F.; Bartels, D. Expression of desiccation-related proteins from the resurrection plant Craterostigma plantagineum in transgenic tobacco. Plant Mol. Biol. 1992, 20, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Z.; Wang, L.; Wu, R.; Phillips, J.; Deng, X. LEA 4 group genes from the resurrection plant Boea hygrometrica confer dehydration tolerance in transgenic tobacco. Plant Sci. 2009, 176, 90–98. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, P.; Yusuf, M.A.; Upadhyaya, C.P.; Roy, S.D.; Hohn, T.; Sarin, N.B. The Xerophyta viscosa aldose reductase (aldrxv4) confers enhanced drought and salinity tolerance to transgenic tobacco plants by scavenging methylglyoxal and reducing the membrane damage. Mol. Biotechnol. 2013, 54, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.J.; Wang, B.; Yu, H.M.; Li, Q.; Hou, B.K. The arabidopsis UGT87A2, a stress-inducible family 1 glycosyltransferase, is involved in the plant adaptation to abiotic stresses. Physiol. Plant. 2017, 159, 416–432. [Google Scholar] [CrossRef] [PubMed]
- Le, T.T.; Williams, B.; Mundree, S.G. An osmotin from the resurrection plant tripogon loliiformis (TlOSM) confers tolerance to multiple abiotic stresses in transgenic rice. Physiol. Plant. 2018, 162, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Dai, A. Increasing drought under global warming in observations and models. Nat. Clim. Chang. 2013, 3, 52. [Google Scholar] [CrossRef]
- Deikman, J.; Petracek, M.; Heard, J.E. Drought tolerance through biotechnology: Improving translation from the laboratory to farmers’ fields. Curr. Opin. Biotechnol. 2012, 23, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, F.A.M.; Wellburn, A.R. Novel chloroplasts and unusual cellular ultrastructure in the “resurrection” plant Myrothamnus flabellifolia Welw. (Myrothamnaceae). Bot. J. Linn. Soc. 1976, 72, 51–54. [Google Scholar] [CrossRef]
- Brewer, P.B.; Koltai, H.; Beveridge, C.A. Diverse roles of strigolactones in plant development. Mol. Plant 2013, 6, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.K.; Bowles, D.J. A class of plant glycosyltransferases involved in cellular homeostasis. EMBO J. 2004, 23, 2915–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, C.; Zhang, Y.; Jamil, M.; Hepworth, J.; Charnikhova, T.; Dimkpa, S.O.N.; Meharg, C.; Wright, M.H.; Liu, J.; Meng, X.; et al. Natural variation of rice strigolactone biosynthesis is associated with the deletion of two max1 orthologs. Proc. Natl. Acad. Sci. USA 2014, 111, 2379–2384. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Martínez, J.A.; Poza-Carrión, C.; Cubas, P. Arabidopsis branched1 acts as an integrator of branching signals within axillary buds. Plant Cell 2007, 19, 458–472. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.R.; Chung, K.M.; Park, J.-H.; Oh, S.A.; Ahn, T.; Hong, S.H.; Jang, S.K.; Nam, H.G. Ore9, an F-box protein that regulates leaf senescence in Arabidopsis. Plant Cell 2001, 13, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Stirnberg, P.; Furner, I.J.; Ottoline, L.H.M. Max2 participates in an scf complex which acts locally at the node to suppress shoot branching. Plant J. 2007, 50, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Zentgraf, U. Oxidative stress and leaf senescence. In Annual Plant Reviews: Senescence Processes in Plants; Gan, S., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2007; Volume 26. [Google Scholar]
- He, Y.; Fukushige, H.; Hildebrand, D.F.; Gan, S. Evidence supporting a role of jasmonic acid in Arabidopsis leaf senescence. Plant Physiol. 2002, 128, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Lohman, K.N.; Gan, S.; John, M.C.; Amasino, R.M. Molecular analysis of natural leaf senescence in Arabidopsis thaliana. Physiol. Plant. 1994, 92, 322–328. [Google Scholar] [CrossRef]
- McCabe, M.S.; Garratt, L.C.; Schepers, F.; Jordi, W.J.R.M.; Stoopen, G.M.; Davelaar, E.; van Rhijn, J.H.A.; Power, J.B.; Davey, M.R. Effects of p(SAG12)-IPT gene expression on development and senescence in transgenic lettuce. Plant Physiol. 2001, 127, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.M.; Gan, S.; Quirino, B.; Amasino, R.M. A comparison of the expression patterns of several senescence-associated genes in response to stress and hormone treatment. Plant Mol. Biol. 1998, 37, 455–469. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Gan, S. A gene encoding an acyl hydrolase is involved in leaf senescence in Arabidopsis. Plant Cell 2002, 14, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.D.; Jones, J.T.; Mullet, J.E. Turgor-responsive gene transcription and RNA levels increase rapidly when pea shoots are wilted. Sequence and expression of three inducible genes. Plant Mol. Biol. 1990, 15, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Wollaston, V.; Ainsworth, C. Leaf senescence in Brassica napus: Cloning of senescence related genes by subtractive hybridisation. Plant Mol. Biol. 1997, 33, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; Song, H.R.; Han, S.K.; Han, M.; Kim, C.Y.; Park, J.; Lee, Y.H.; Jeon, J.S.; Noh, Y.S.; Noh, B. Hda19 is required for the repression of salicylic acid biosynthesis and salicylic acid-mediated defense responses in Arabidopsis. Plant J. 2012, 71, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.-K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 2015, 6, 114. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; To, T.K.; Seki, M. An epigenetic integrator: New insights into genome regulation, environmental stress responses and developmental controls by histone deacetylase 6. Plant Cell Physiol. 2012, 53, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Bej, S.; Basak, J. Abiotic stress induced epigenetic modifications in plants: How much do we know? In Plant Epigenetics; Rajewsky, N., Jurga, S., Barciszewski, J., Eds.; Springer: New York, NY, USA, 2017; pp. 493–512. [Google Scholar]
- Diez, C.M.; Roessler, K.; Gaut, B.S. Epigenetics and plant genome evolution. Curr. Opin. Plant Biol. 2014, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Meyer, P. Epigenetic variation and environmental change. J. Exp. Bot. 2015, 66, 3541–3548. [Google Scholar] [CrossRef] [PubMed]
- Matzke, M.A.; Mosher, R.A. RNA-directed DNA methylation: An epigenetic pathway of increasing complexity. Nat. Rev. Genet. 2014, 15, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Niederhuth, C.E.; Schmitz, R.J. Putting DNA methylation in context: From genomes to gene expression in plants. Biochim. Biophys. Acta 2017, 1860, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Furini, A.; Koncz, C.; Salamini, F.; Bartels, D. High level transcription of a member of a repeated gene family confers dehydration tolerance to callus tissue of Craterostigma plantagineum. EMBO J. 1997, 16, 3599–3608. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, J.; Hussain, S.S.; Shi, B.J. Role of microRNAs in plant drought tolerance. Plant Biotechnol. J. 2015, 13, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khraiwesh, B.; Zhu, J.-K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith-Espinoza, C.; Phillips, J.; Salamini, F.; Bartels, D. Identification of further Craterostigma plantagineum cdt mutants affected in abscisic acid mediated desiccation tolerance. Mol. Genet. Genom. 2005, 274, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, T.; Shen, C.-Y.; Xu, G.-H.; Chen, S.-X.; Song, L.-Z.; Li, M.-J.; Wang, L.-L.; Zhu, Y.; Lv, W.-T.; et al. Identification of a retroelement from the resurrection plant Boea hygrometrica that confers osmotic and alkaline tolerance in Arabidopsis thaliana. PLoS ONE 2014, 9, e98098. [Google Scholar] [CrossRef] [PubMed]
- Ditzer, A.; Bartels, D. Identification of a dehydration and ABA-responsive promoter regulon and isolation of corresponding DNA binding proteins for the group 4 LEA gene cpc2 from C. plantagineum. Plant Mol. Biol. 2006, 61, 643–663. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Owen-Hughes, T. Snf2-family proteins: Chromatin remodellers for any occasion. Curr. Opin. Chem. Biol. 2011, 15, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Strein, C.; Alleaume, A.-M.; Rothbauer, U.; Hentze, M.W.; Castello, A. A versatile assay for RNA-binding proteins in living cells. RNA 2014, 20, 721–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, J.K.; Gayali, S.; Dass, S.; Kumar, A.; Parveen, S.; Chakraborty, S.; Chakraborty, N. Osalba1, a dehydration-responsive nuclear protein of rice (Oryza sativa L. ssp. Indica), participates in stress adaptation. Phytochemistry 2014, 100, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Kapazoglou, A.; Drosou, V.; Argiriou, A.; Tsaftaris, A.S. The study of a barley epigenetic regulator, HvDME, in seed development and under drought. BMC Plant Biol. 2013, 13, 172. [Google Scholar] [CrossRef] [PubMed]
- Song, C.-P.; Agarwal, M.; Ohta, M.; Guo, Y.; Halfter, U.; Wang, P.; Zhu, J.-K. Role of an arabidopsis ap2/erebp-type transcriptional repressor in abscisic acid and drought stress responses. Plant Cell 2005, 17, 2384–2396. [Google Scholar] [CrossRef] [PubMed]
- To, T.K.; Nakaminami, K.; Kim, J.-M.; Morosawa, T.; Ishida, J.; Tanaka, M.; Yokoyama, S.; Shinozaki, K.; Seki, M. Arabidopsis HDA6 is required for freezing tolerance. Biochem. Biophys. Res. Commun. 2011, 406, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-Y.; Chen, S.-Y. Inducible expression of translation elongation factor 1a gene in rice seedlings in response to environmental stresses. Acta Bot. Sin. 1999, 41, 800–806. [Google Scholar]
- Sani, E.; Herzyk, P.; Perrella, G.; Colot, V.; Amtmann, A. Hyperosmotic priming of Arabidopsis seedlings establishes a long-term somatic memory accompanied by specific changes of the epigenome. Genome Biol. 2013, 14, R59. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.B.; Rai, Y.; Gayali, S.; Chakraborty, S.; Chakraborty, N. Plant organellar proteomics in response to dehydration: Turning protein repertoire into insights. Front. Plant Sci. 2016, 7, 460. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.J.; Guo, L.; Alexander, D.C.; Ryals, J.A.; Wone, B.W.M.; Cushman, J.C. A sister group contrast using untargeted global metabolomic analysis delineates the biochemical regulation underlying desiccation tolerance in Sporobolus stapfianus. Plant Cell 2011, 23, 1231–1248. [Google Scholar] [CrossRef] [PubMed]
- Tymms, M.; Gaff, D. Recovery of protein synthesis and RNA synthesis after rewetting of desiccated leaves of the angiosperm Xerophyta villosa. Biochem. Physiol. Pflanz. 1984, 179, 211–217. [Google Scholar] [CrossRef]
Figure 1.
Summary and flow chart detailing the hypothesis for the regulation of desiccation tolerance in flowering-plants, with a focus on S. stapfianus.
Figure 1.
Summary and flow chart detailing the hypothesis for the regulation of desiccation tolerance in flowering-plants, with a focus on S. stapfianus.
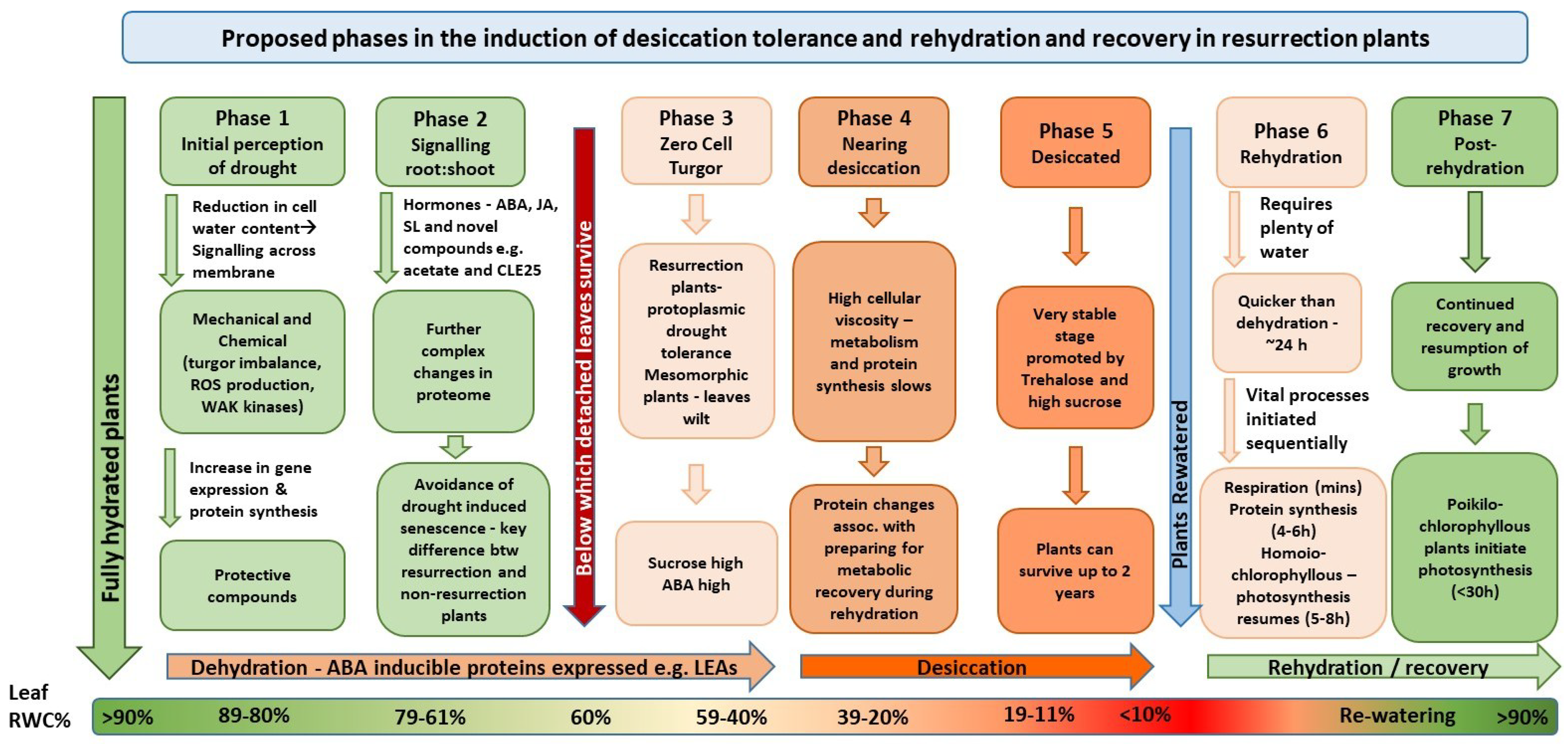

{kind=link}
Table 1.
Grass species (Poaceae) with desiccation-tolerant foliage. The number of desiccation tolerant species is given in relation to the total number of species in each genus. Additional details on tolerance limits are noted for each species. Data from [2,3,4,5,6,7,8,9].
| Genus & Species No. of Desiccation Tolerant Species/of Total spp. in Genus | Notes on Tolerance Limits | Genus & Species No. of Desiccation Tolerant Species/of Total spp. in Genus | Notes on Tolerance Limits | ||
|---|---|---|---|---|---|
| Brachyachne 1/10 | Oropetium 3/4 | ||||
| B. patentifolia | 0% RH | >15 cm | O. capense | 0% RH | |
| Eragrostiella 3/5 | O. roxburghianum | 0% RH | |||
| E. bifaria var. bifaria | 15% RH | G | O. thomaeum | 0% RH | |
| E. brachyphylla | 0% RH, | Poa 1/~500 | |||
| E. nardoides | 11% RH | Poa bulbosa | 5% RH | Bs V | |
| Eragrostis 4/350 | Sporobolus 7/160 | ||||
| Er. hispida | 0–5% RH | Bs, >15 cm | S. atrovirens | <4% RWC | |
| Er. invalida | 0% RH | Bs, >30 cm | S. blakei | 0% RH | Bs |
| Er. nindensis | 0–2% RH | G, >15 cm | S. elongatus | <10% RWC | Bs > 30 cm |
| Er. paradoxa | 0% RH | >15 cm | S. festivus | 0% RH | >15 cm |
| Micraira 7/14 | S. lampranthus | 0% RH | Bs | ||
| M. adamsii | 0–2% RH | S. pellucidus | 0–5% RH | >15 cm | |
| M. lazaridis | - | S. stapfianus | 2% RH | >15 cm | |
| M. multinervia | >5% RH | Tripogon 10/30 | |||
| M. spinifera | <7.5% RWC | T. capillatus | 0% RH | >15 cm | |
| M. subulifolia | 5% RH | T. curvatus | 0% RH | ||
| M. tenuis | <8% RWC | T. filiformis | 2% RH | >25 cm | |
| M. viscidula | <7.5% RWC | T. jaquemontii | 0% RH | G | |
| Microchloa 3/4 | T. lisboae | 0% RH | >25 cm | ||
| Mi. caffra | 0–5% RH | >15 cm | T. loliiformis | 0% RH | G |
| Mi. indica | 0% RH | T. major | 0% RH | Bs | |
| Mi. kunthii | 0% RH | T. minimus | 0–2% RH | ||
| T. polyanthus | 2% RH | >15 cm | |||
| T. spicatus | 0% RH | ||||
Abbreviations: ‘PDT’ is leaf Protoplasmic Drought Tolerance = ‘RH’ the lowest relative humidity of air survived at equilibrium or ‘RWC’ the lowest relative water content survived; ‘>numerical value’ e.g., >15 = vegetative foliage 15 cm or more tall; ‘Bs’ = only the basal 1cm of leaf is desiccation tolerant; ‘G’ = plants recorded as grazed by animals; V = vivaporous bulbils are desiccation-tolerant.
Table 2.
Species with desiccation tolerance confined to immature leaf tissue. Data from the references cited in Table 1 and [23].
| DICOTYLEDONS | Tissue Type |
|---|---|
| Gesneriaceae | |
| Conandron ramondioides | Young leaf to 1 cm size |
| Opithandra primuloides | To 2nd youngest leaf (1 cm long) |
| Stylidiaceae | |
| Stylidium sp. | Buds only |
| MONOCOTYLEDONS | |
| Anthericaceae | |
| Borya scirpioides | High in 3 youngest leaves |
| Cyperaceae | |
| Carex physodes | Bs = ~1 cm at base of the leaf lamina, including the leaf meristem |
| Carex pachystilis | Bs |
| Cyperus bellis | Bs |
| Fimbristylis dichotoma | Bs |
| Kyllinga alba Nees | Bs |
| Kyllinga alata | Bs |
| Kyllinga nervosa | Bs |
| Mariscus capensis | Bs |
| Poaceae: see Table 1 |
Table 3.
Beginning of induction of desiccation tolerance in leaves on drying S. stapfianus plants. Leaves, detached from plants at different stages of drying, were equilibrated to air of 76% Relative Humidity, then rehydrated 24 h in water and their survival scored as the % of their leaf area that survived. Data based on [35].
Table 3.
Beginning of induction of desiccation tolerance in leaves on drying S. stapfianus plants. Leaves, detached from plants at different stages of drying, were equilibrated to air of 76% Relative Humidity, then rehydrated 24 h in water and their survival scored as the % of their leaf area that survived. Data based on [35].
| Leaf RWC (%) | 94 | 82 | 66 | 61 | 58 | 49 | 48 | 46 |
| Survival % of total leaf area | 6 | 2 | 0 | 48 | 67 | 59 | 85 | 85 |
Table 4.
Protein synthesis indicated by polyribosome content in leaves of the resurrection plant Xerophyta villosa on intact plants drying to a viable airdry condition. Data based on [132].
Table 4.
Protein synthesis indicated by polyribosome content in leaves of the resurrection plant Xerophyta villosa on intact plants drying to a viable airdry condition. Data based on [132].
| RWC (%) | 96 | 90–75 | 75–60 | 45–40 | 30 | 20 | 4 |
| % RNA as Polysomes | ~30 | ~25 | ~45 | ~55 | ~32 | ~1 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Blomstedt, C.K.; Griffiths, C.A.; Gaff, D.F.; Hamill, J.D.; Neale, A.D. Plant Desiccation Tolerance and its Regulation in the Foliage of Resurrection “Flowering-Plant” Species. Agronomy 2018, 8, 146. https://doi.org/10.3390/agronomy8080146
AMA Style
Blomstedt CK, Griffiths CA, Gaff DF, Hamill JD, Neale AD. Plant Desiccation Tolerance and its Regulation in the Foliage of Resurrection “Flowering-Plant” Species. Agronomy. 2018; 8(8):146. https://doi.org/10.3390/agronomy8080146
Chicago/Turabian StyleBlomstedt, Cecilia K., Cara A. Griffiths, Donald F. Gaff, John D. Hamill, and Alan D. Neale. 2018. "Plant Desiccation Tolerance and its Regulation in the Foliage of Resurrection “Flowering-Plant” Species" Agronomy 8, no. 8: 146. https://doi.org/10.3390/agronomy8080146
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.