Arabidopsis thaliana Immunity-Related Compounds Modulate Disease Susceptibility in Barley


Helmholtz Zentrum München, Department of Environmental Science, Institute of Biochemical Plant Pathology, Ingolstädter Landstr. 1, 85764 Neuherberg, Germany
*
Author to whom correspondence should be addressed.
Agronomy 2018, 8(8), 142; https://doi.org/10.3390/agronomy8080142
Submission received: 6 July 2018
/
Revised: 31 July 2018
/
Accepted: 4 August 2018
/
Published: 7 August 2018
(This article belongs to the Special Issue Biotic and Abiotic Stress Responses in Crop Plants)
Abstract
:Plants are exposed to numerous pathogens and fend off many of these with different phytohormone signalling pathways. Much is known about defence signalling in the dicotyledonous model plant Arabidopsis thaliana, but it is unclear to which extent knowledge from model systems can be transferred to monocotyledonous plants, including cereal crops. Here, we investigated the defence-inducing potential of Arabidopsis resistance-inducing compounds in the cereal crop barley. Salicylic acid (SA), folic acid (Fol), and azelaic acid (AzA), each inducing defence against (hemi-)biotrophic pathogens in Arabidopsis, were applied to barley leaves and the treated and systemic leaves were subsequently inoculated with Xanthomonas translucens pv. cerealis (Xtc), Blumeria graminis f. sp. hordei (powdery mildew, Bgh), or Pyrenophora teres. Fol and SA reduced Bgh propagation locally and/or systemically, whereas Fol enhanced Xtc growth in barley. AzA reduced Bgh propagation systemically and enhanced Xtc growth locally. Neither SA, Fol, nor AzA influenced lesion sizes caused by the necrotrophic fungus P. teres, suggesting that the tested compounds exclusively affected growth of (hemi-)biotrophic pathogens in barley. In addition to SA, Fol and AzA might thus act as resistance-inducing compounds in barley against Bgh, although adverse effects on the growth of pathogenic bacteria, such as Xtc, are possible.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Plants are constantly challenged with a plethora of pathogens, including herbivorous insects, fungi, oomycetes, bacteria and viruses [1]. To cope with these attacks, plants have evolved an effective immune system of pre-formed and inducible defence mechanisms. The former include morphological barriers, for instance the plant cell wall, cuticle, phytoanticipins, and antimicrobial proteins. Inducible defence mechanisms include processes like cell wall reinforcement by lignin and callose or the synthesis of phytoalexins and defence-related proteins or enzymes [1,2,3,4].
After the first contact between plant and pathogen, conserved structures such as bacterial flagellin or fungal chitin—so-called pathogen-associated molecular patterns (PAMPs)—encounter pattern recognition receptors on the cell surface. This triggers a first immune response termed PAMP-triggered immunity (PTI) [2,4,5]. During long-term evolution of plant–pathogen interactions, some pathogens developed effector proteins to suppress PTI. In response to this development, plants evolved resistance (R) genes, which encode proteins that directly or indirectly recognise effectors, and on recognition elicit so called effector-triggered immunity (ETI) [2,3,4,6].
Both PTI and ETI are associated with the induction of various cellular responses. These include the synthesis of antimicrobial compounds, the generation of reactive oxygen species, the activation of mitogen-activated protein kinases, transcriptional reprogramming, and the accumulation of phytohormones such as salicylic acid (SA) [2,3,4,7,8]. SA is known to affect the redox status of plants, thereby leading to transcriptional reprogramming and enhanced transcription of PATHOGENESIS-RELATED (PR) genes, which are thought to promote resistance [7,8,9,10,11]. Furthermore, SA not only helps the plant to defend itself against a present infection, it is also involved in the induction of a process protecting the plant in case of future pathogen challenge [4,6,8,12]. This response is termed systemic acquired resistance (SAR) and is usually triggered by a local, foliar infection. It elicits long-lasting, broad-spectrum resistance in systemic plant tissues (reviewed in [6,7,8,9,12]). Many signals and genes involved in SAR have been discovered in Arabidopsis thaliana. During SAR, SA levels in the infected and systemic tissues rise, and transcripts of PR as well as other defence-related genes accumulate. This establishes a status of heightened alert; the plant is “primed” to respond more quickly to a secondary infection [13,14]. SAR is most effective against pathogens fended off via SA-dependent responses [3,4,8].
During SAR, long-distance signals are generated in the infected leaves and travel to the systemic leaves, presumably via the phloem [12]. Recent evidence suggests that signal transmission also occurs via volatile compounds, in particular monoterpenes, which appear to be transmitted via the air [15]. In Arabidopsis, candidate long-distance SAR signals further include methyl salicylate [16], glycerol-3-phosphate (G3P) [17,18], the C9 dicarboxylic acid azelaic acid (AzA) [19], the diterpene dehydroabietinal [20], N-hydroxy-pipecolic acid [21,22], and the lipid-transfer proteins DIR1 (DEFECTIVE IN INDUCED RESISTANCE 1) and DIR1-like [23,24] (reviewed in [12,25,26]). Some of these compounds accumulate in the phloem after pathogen attack, among them SA, AzA, G3P, and pipecolic acid [19,27,28,29]. AzA and G3P were proposed to be symplastically loaded onto the phloem via plasmodesmata while SA appeared to be transported via an apoplastic route [29]. Only small proportions of SA and AzA are transported to the systemic tissue, and the majority of accumulation in systemic leaves thus appears to come from de novo synthesis [30,31,32]. Importantly, biosynthesis of the SAR-related compounds SA and pipecolic acid in systemic leaves is crucial for SAR establishment, questioning the biological relevance of the systemic mobility of these signals [18,33,34].
Most of the mobile metabolites induce SAR when exogenously applied to Arabidopsis plants. Local application of AzA, for example, induces a SAR-like state in the treated plants, protecting the systemic tissue against a subsequent infection with the hemi-biotrophic bacterium Pseudomonas syringae [19]. Additional SAR-inducing compounds include folate precursors or folic acid (Fol), which induces SA-mediated immunity in Arabidopsis, both locally and systemically, and in pepper (Capsicum annuum) [35,36].
In monocotyledonous plants, less is known about SAR and the signalling mechanisms involved. Key players in SA signalling, including NPR1 (NONEXPRESSOR OF PR GENES 1), the master regulator of SA signalling [7,9,10], several PR genes and SA-associated transcription factors are conserved between dicots and monocots, (reviewed in [37,38]). Most studies in this field focus on agronomically important plant species such as banana (Musa acuminata) [39], wheat (Triticum aestivum) [40,41,42], maize (Zea mays) [43], and rice (Oryza sativa) [38], in which important roles for SA signalling have been uncovered in resistance against pathogens including Fusarium oxysporum, Puccinia striiformis, Xanthomonas oryzae pv. oryzae and Magnaporthe oryzae. Treatments with the SA analogue benzothiadiazole (BTH) induce resistance in maize [44], wheat [45], and barley [46,47,48].
SA-associated immune responses in barley (Hordeum vulgare) often are studied in interaction with Blumeria graminis f. sp. hordei (Bgh), commonly named powdery mildew, an obligate biotrophic fungal pathogen that thrives on living host cells [49]. Since it causes reduced yield and serves as a model to study other mildews and obligate biotrophic pathogens [50], Bgh appeared on the list of the top 10 fungal pathogens by Dean et al. in 2012 [51]. In contrast to biotrophic pathogens of rice, Bgh inoculation does not result in SA accumulation in infected barley leaves [52,53,54]. Nevertheless, SA soil drench treatment of barley seedlings had a weak effect on Bgh infectivity [46,55], whereas soil drench treatment with BTH strongly enhanced barley resistance to Bgh [46].
Recently, Dey et al. [47] showed that in barley systemic resistance to the hemi-biotrophic bacterium Xanthomonas translucens pv. cerealis (Xtc) can be triggered by prior infiltration of a single barley leaf with the hemi-biotrophic bacterium Pseudomonas syringae pv. japonica. Unlike SAR in Arabidopsis, systemic immunity in barley was neither associated with SA nor with NPR1. In addition, local infiltration of SA or its functional analogue BTH did not induce systemic resistance to Xtc. Rather, local methyl jasmonate (MeJA) and abscisic acid (ABA) treatments, which in Arabidopsis induce systemic susceptibility to P. syringae [56] or antagonize SAR [57], respectively, triggered systemic resistance in barley to Xtc [47].
In plant immunity, cross talk between phytohormones is important and is believed to help achieve the best possible (defence) outcome. In phytohormone cross talk, an interaction is never defined by a single hormone, but rather by a complex network of interdependent positive and negative interactions [58,59,60]. The result of these interactions leads to responses in the plant, which allow it to appropriately respond to an invading pathogen. Pathogens of different lifestyles have different demands on their hosts and, therefore, must be combatted using different mechanisms; while biotrophs feed on living cells, necrotrophs acquire their nutrients from degraded and dead tissue. Several phytohormones are involved in plant defence responses against pathogens. The traditional three main players are SA, jasmonic acid (JA) and ethylene (ET), but recent evidence hints at the additional contribution of other hormones [58,59,60]. Mostly synergistic interactions are reported for JA and ET, which mainly promote defence against necrotrophic pathogens and insects [3,59,60,61]. SA and JA signalling pathways are interdependent; there is substantial cross talk between the two, comprising synergistic as well as antagonistic interactions, depending on the defence situation [3,59,62]. Biotrophic pathogens are mostly opposed using SA signalling, whereas necrotrophic pathogens are combatted using JA/ET-dependent pathways [3,59,60,62]. Because SA–JA cross talk is often antagonistic, SA-induced resistance against biotrophs can enhance susceptibility against necrotrophs and vice versa [60,63]. In Arabidopsis, for example, SA and Fol induce resistance against hemi-biotrophic bacteria and at the same time enhance susceptibility to the necrotrophic fungal pathogen Alternaria brassicicola [35,63]. Here, we query if such antagonistic cross talk between responses to biotrophic and necrotrophic pathogens also occurs in barley by using a barley pathogen from the necrotrophic side of the spectrum, the fungus Pyrenophora teres. P. teres is the causal agent of net and spot form net blotch, a major disease in many barley-growing areas, which can lead to severe yield loss, underlining the fungus’ economic importance [64].
We want to study the possible effects of Arabidopsis SAR-associated compounds on barley defence responses. Therefore, we will investigate the influence of different chemical compounds, including SA, on the propagation of barley-specific pathogens. We will employ pathogens of different lifestyles, namely the hemi-biotrophic bacterium Xtc, the biotrophic fungus Bgh and the necrotrophic fungus P. teres. In addition to SA, we will include Fol, which induces an SA-dependent defence response in Arabidopsis [35]. Also, we will include AzA, a SAR signal, which in Arabidopsis primes SA accumulation, inducing a faster and stronger response after pathogen attack compared to that in unprimed plants [19]. The data shed light upon the possible transferability of signalling intermediates related to SAR from Arabidopsis to barley. As we detected differences and similarities in the responses of these plants interacting with (hemi-)biotrophic and necrotrophic pathogens, the data reinforce the necessity of studying metabolite-induced resistance in cereal crops.
2. Materials and Methods
2.1. Plants and Growth Conditions
Barley (Hordeum vulgare L. cultivar ‘Golden Promise’ (GP)) seeds were sterilized in 1.2% sodium hypochlorite for 3 min with 25 inversions per minute. Subsequently, seeds were rinsed 3 times with water for 10 min with 25 inversions per minute and then sown (Einheitserde classic CL-T, Bayerische Gärtnereigenossenschaft). Plants for Xtc infections were either grown in a greenhouse with additional lights HQI-TS 400W/D (Osram) using a day-night cycle of 12 h with 24 °C during the day and 20 °C during the night or in a climate chamber with 16 h light and 8 h darkness at a temperature of 20 °C (day)/16 °C (night). Chamber-grown plants inoculated with Xtc were transferred to a climate chamber with 14 h light and 10 h darkness at a temperature of 29 °C (day)/19 °C (night). Plants for Bgh infections were grown in climate chambers with 16 h light and 8 h darkness at a temperature of 20 °C (day)/16 °C (night). Plants for P. teres infections were grown in the green house as described above.
2.2. Chemicals and Treatments
Stock solutions of each chemical compound were freshly prepared for each experiment. SA (Roth, Karlsruhe, Germany) was dissolved at 4 M in 100% methanol (MeOH; Merck, Darmstadt, Germany), Fol (Roth, Karlsruhe, Germany) was dissolved at 1 M and AzA (Sigma-Aldrich, St. Louis, MO, USA) at 2 M in 50% MeOH. For plant treatments, the substances were diluted to 1 mM SA [35,45], 50 or 500 µM Fol [35], and 1 mM AzA [19,31] in 10 mM MgCl2 (Roth, Karlsruhe, Germany). 0.025% MeOH in 10 mM MgCl2 served as the mock treatment. To monitor systemically induced resistance, the first true leaves of 3-week-old barley plants (6 plants per treatment) were infiltrated with the different compounds or the mock solution using a needleless syringe. Five days after the primary treatment, the second leaves of the treated plants were infected with either Xtc, Bgh, or P. teres (see below). To monitor local induced resistance against Xtc, 4-week-old barley plants were sprayed with 1 mM SA, 500 µM Fol, 1 mM Aza, or 0.05% MeOH (as mock treatment) in 0.01% Tween-20 (Calbiochem, San Diego, CA, USA). To monitor local induced resistance against Bgh, 1 mM SA, 500 µM Fol, and 1 mM AzA in 10 mM MgCl2 were syringe-infiltrated into the first true leaves of 3-week-old barley plants. 0.025% MeOH in 10 mM MgCl2 served as the mock treatment. To monitor local induced resistance against P. teres, the second true leaves of 3-week-old barley plants were syringe-infiltrated with the compounds. The treated leaves were inoculated with Xtc 1 day after treatment, with Bgh 5 days after treatment, or with P. teres 1 day after treatment.
2.3. Xanthomonas Translucens pv. Cerealis (Xtc) Infection
Xanthomonas translucens pv. cerealis (Xtc) strain LMG7393 was obtained from the Laboratory of Microbiology UGent (LMG) collection of the Belgian Coordinated Collections of Microorganisms. For infection, Xtc was grown on LMG medium (15 g/L tryptone, 5 g/L soya peptone, 5 g/L NaCl, and 18 g/L agar-agar (Roth, Karlsruhe, Germany); pH adjusted to 7.3) over night at 28 °C. Bacteria were subsequently resuspended in 1 mL 10 mM MgCl2 (Roth, Karlsruhe, Germany). The concentration of the bacterial suspension was adjusted to ~105 colony forming units (cfu)/mL in 10 mM MgCl2 using a photometer (assuming the formula: OD600 of 0.2 equals ~108 cfu/mL). Leaves of 3–5 barley plants were subsequently inoculated with the resulting Xtc suspension by infiltration using a needleless syringe. The infected plants were covered with a plastic hood and kept in the green house for 4 days. The resulting in planta Xtc titres were determined as previously described [47,65].
2.4. Blumeria Graminis f. sp. Hordei (Bgh) Infection
Blumeria graminis f. sp. hordei (Bgh) Swiss field isolate CH4.8 was obtained from Dr. Patrick Schweizer (Leibniz-Institut für Pflanzengenetik und Kulturpflanzenforschung, Gatersleben, Germany). Bgh propagation and inoculation was performed essentially as described in [66]. In short, a pot containing 12 10-day-old seedlings was infected with Bgh one week prior to each experiment. Six hours prior to the start of an experiment, these plants were shaken in order to remove old conidia and provide a uniform inoculum for the experiment [67]. Compound- or mock-treated plants were subsequently inoculated with spores from the prepared Bgh-infected plants in an inoculation tower [66] at an inoculation density of ~30 spores/mm2. The inoculated plants were placed back in the climate chamber for 6 days. Subsequently, 4 leaf discs (6 mm) were cut out of the distal halves of each first (local/compound-treated) and second (systemic) true leaf of the treated plants. The discs were incubated with 5 µM DAF-FM-DA (4-amino-5-methylamino-2’,7’-difluorofluorescein diacetate; Sigma-Aldrich, St. Louis, MO, USA or Santa Cruz Biotechnology, Dallas, TX, USA) in MES buffer (2-(N-morpholino)ethanesulfonic acid, 50 mM MES-KOH pH 5.7 (Roth, Karlsruhe, Germany), 1 mM CaCl2 (Merck, Darmstadt, Germany), 0.25 mM KOH) for 45 min in the dark and then vacuum-infiltrated. Subsequently, the leaf discs were placed in light (45 V lamp) for 1 h and 45 min and afterwards distributed on 96-well plates (1 leaf disc per well, 20–24 discs from 6 plants per treatment) with the wells filled evenly to the rim with 1% phytoagar. Fluorescence of appropriate z-stacks was visualized using the 5× objective of an inverse spinning disc confocal microscope (Zeiss Axio Observer. Z1, Zeiss, Oberkochen, Germany). Chlorophyll was excited using a laser with 561 nm and detected using a bandpass 629/62 filter; DAF-FM DA was excited using a laser with 488 nm and detected using a bandpass 525/50 filter [68]. The 96-well plates were transferred to the microscope with a KiNDx Robot (Model KX-300-660-SSU, Peak Analysis and Automation, Inc., Farnborough, UK; assembled and set up by Analytik Jena, Jena, Germany). The robot and the visualization as well as evaluation were controlled by the softwares Microscope AppStudio (Analytik Jena, Jena, Germany) and ZEN2 (Zeiss, Oberkochen, Germany). Fluorescence intensities were normalized to those of uninfected barley plants (background fluorescence) of the same age.
2.5. P. Teres Infection
A field isolate of Pyrenophora teres was donated by Günther Bahnweg (Helmholtz Zentrum München, Neuherberg, Germany) and grown on oat plates (10 g rolled oats (Alnatura, Germany), 7.5 g agar-agar (Roth, Karlsruhe, Germany), 500 mL H2O) for ~1 week at room temperature in the dark and then transferred to light for at least 2 weeks. Two mL of infection solution for fungi (0.85 g KH2PO4 (Merck, Darmstadt, Germany), 0.1 g glucose (Roth, Karlsruhe, Germany), and 1 µL Tween 20 in 100 mL H2O, pH 6.0) were pipetted onto the P. teres plates and spores were scratched off the agar using an inoculation loop. The spore suspension was subsequently pipetted into a 5 mL tube and vortexed. After determining the spore concentration under a binocular, the spore suspension was diluted to 65–110 spores per µL. Infections were performed on 6 cm-long segments of a leaf, at a distance of 1.5 centimetres from the leaf base. Five 3 µL droplets of spore suspension were pipetted alternatingly on each side of the leaf midrib. The drops were left to dry for ~1 h and the plants (6 plants per treatment) were then covered with a plastic hood. Necrotic lesions caused by P. teres were measured 4 days after infection using the ImageJ macro PIDIQ [69]. The macro was modified to measure brown necrotic lesions caused by P. teres. These modifications were restricted to the values used for colour characterisation; the values for lesion measurements were as follows: hue 0–52, saturation 150–255, brightness 0–150.
2.6. Statistics
Results of biologically independent replicate experiments with two groups were tested together for homogeneity of variance using the F-test in Microsoft Excel. Depending on the outcome of the F-test, a two-sided t-test either for equal (homoscedastic) or unequal variances (heteroscedastic) was conducted in Microsoft Excel. Results of biologically independent replicate experiments with more than two groups were analysed in GraphPad Prism 7 for Windows (version 7.04). Data with only positive values were log2 transformed and data with negative and positive values were transformed according to the formula: Y = log2[Y + 1 − min(Y)], where min(Y) denotes the lowest measured value within the experiment. Subsequently, the data were analysed for outliers using Grubbs’ test with α = 0.05. The D’Agostino–Pearson normality test was performed with α = 0.01 in order to test for normal distribution. If necessary the Grubbs’ outlier test was repeated to assure normal data distribution (a maximum of 2 Grubbs’ outlier tests were performed per data set). The data were subsequently analysed using one-way analysis of variance (ANOVA) with the Geisser-Greenhouse correction (p < 0.05) and a subsequent Dunnett’s post hoc test with α = 0.05.
3. Results
3.1. Folic Acid and Azelaic Acid Enhance the Susceptibility of Barley to Xtc
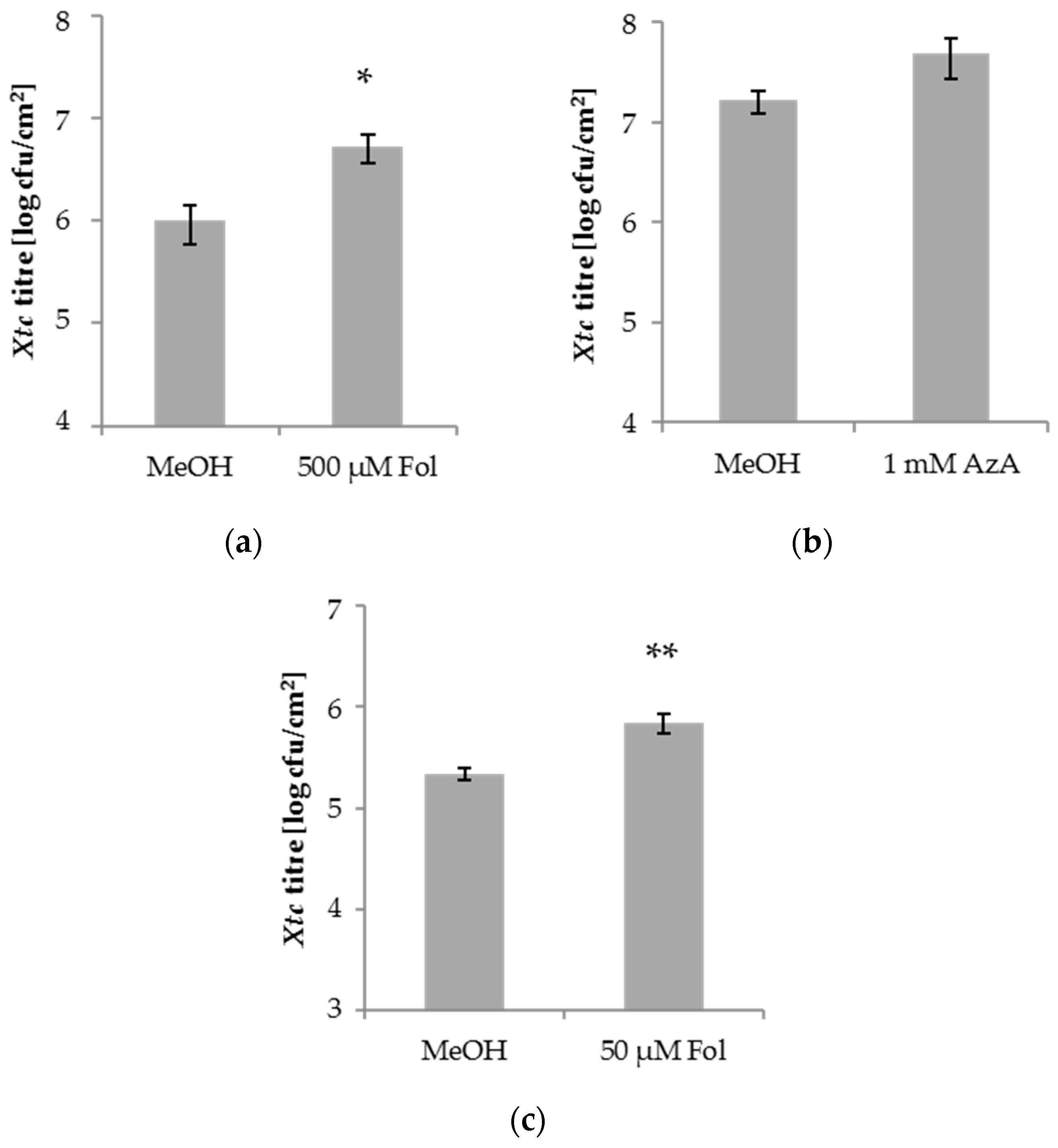
Exogenous SA or BTH application enhances the resistance of barley to the biotrophic fungus Bgh [46], but not to the hemi-biotrophic bacterium Xtc [47]. Here, we examined Xtc growth in barley after application of Fol and AzA, which each trigger SA-mediated resistance and/or SAR against the hemi-biotrophic bacterium P. syringae pv. tomato in A. thaliana [19,35]. In 3-week-old barley plants, the first true leaves were syringe-infiltrated with 50 or 500 µM Fol, 1 mM AzA, or the corresponding 0.025% MeOH mock control. Five days later, the second true leaves of the treated plants were syringe-infiltrated with Xtc and the resulting in planta Xtc titres determined at 4 days post-inoculation (dpi). Similar to SA, Fol and AzA did not reduce Xtc growth in the systemic tissue of barley (Figure 1). Whereas AzA application did not influence Xtc growth in the systemic tissue (Figure 1b), growth of the systemic Xtc inoculum was enhanced by a local Fol application (Figure 1a), suggesting the induction of enhanced susceptibility in barley against Xtc by Fol.
Fol enhances A. thaliana resistance to P. syringae when applied at concentrations of 50 to 100 µM [35]. If we applied 50 µM Fol to the first leaves of 3-week-old barley plants, the growth of subsequently applied Xtc bacteria in the systemic leaves was enhanced in three out of 10 experiments and a strong tendency in the same direction was observed in a further two experiments. If we analyse all experiments together, the data suggest that application of 50 µM Fol can, similarly to 500 µM Fol, enhance barley systemic susceptibility to Xtc (Figure 1c), albeit perhaps less robustly.
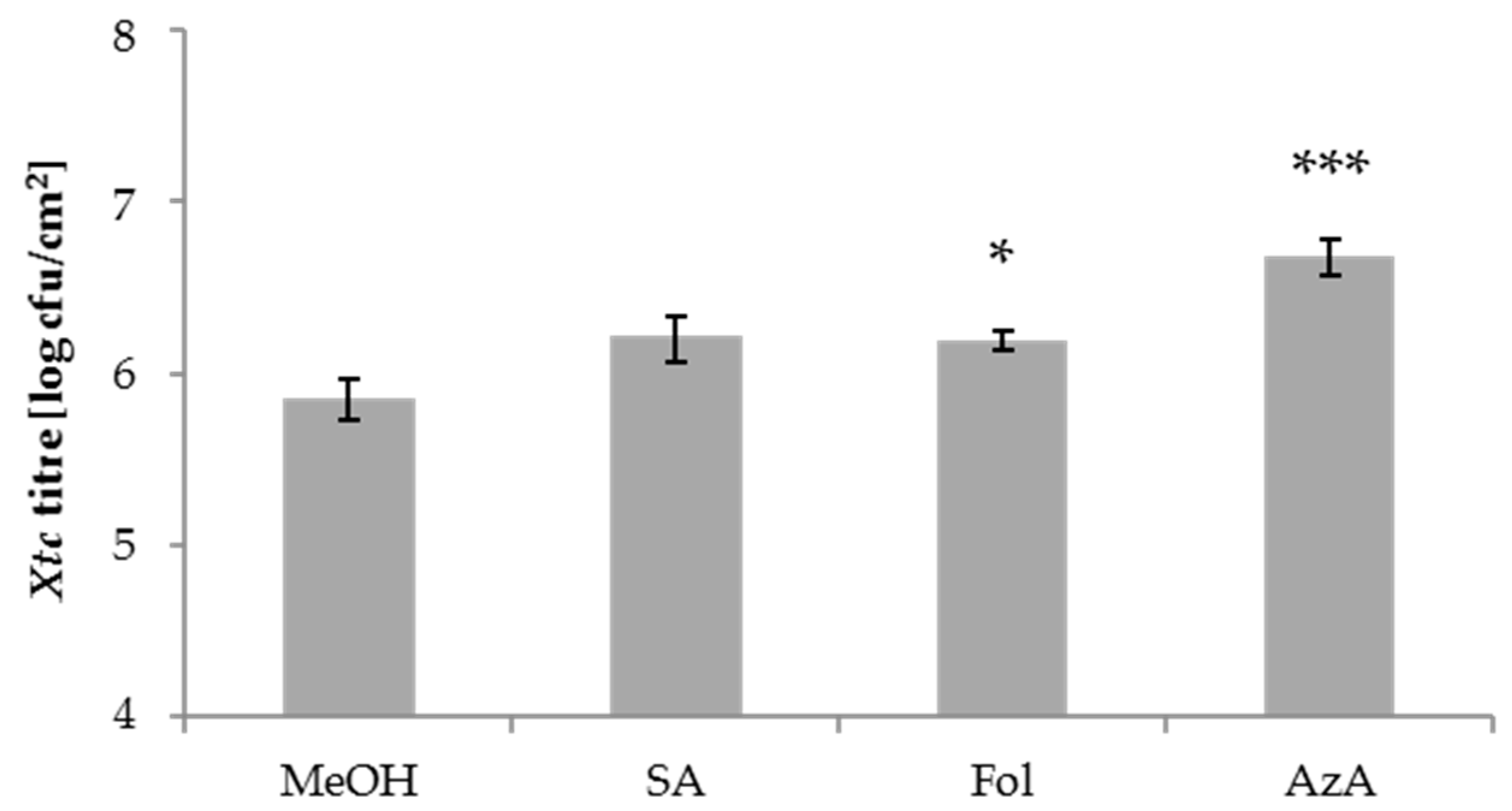
We next tested the local barley responses to the compounds used above. To this end, 1 mM SA, 500 µM Fol, 1 mM AzA, or the appropriate mock control were applied onto the leaves of 4-week-old barley plants by spray treatment. One day later, treated leaves were syringe-infiltrated with Xtc and the resulting in planta Xtc titres determined at 4 dpi. SA and Fol appeared to moderately elevate local Xtc titres, with a significant trend observed only after Fol treatment. AzA application caused an increase of the Xtc titre (Figure 2). Thus, in addition to systemic susceptibility, Fol might also moderately enhance the local susceptibility of barley to the hemi-biotrophic bacterium Xtc. The effect appeared more robust in the systemic compared to local treated tissues (Figure 1a,c and Figure 2). AzA enhanced local, but not systemic susceptibility to Xtc.
3.2. Folic Acid Enhances the Resistance of Barley to Powdery Mildew (Bgh)
In Arabidopsis Fol triggers SA-mediated immunity against hemi-biotrophic bacteria [35]. In barley, SA and in particular its functional analogue BTH enhances immunity against the biotrophic fungus Bgh while Fol enhances barley susceptibility to Xtc (Figure 1 and Figure 2; [46]). Here, we compared the effects of SA, Fol, and AzA on propagation of Bgh in barley. To this end, the first true leaves of 3-week-old barley plants were syringe-infiltrated with 1 mM SA, 500 µM Fol, 1 mM AzA, or 0.025% MeOH as control treatment. Five days later, the plants were inoculated with Bgh. The resulting Bgh infection was quantified on a fluorescence microscope 6 days later. We used the fluorescent dye DAF-FM DA which usually is used to detect and quantify NO [70] and, as shown in Figure 3, also has a high affinity for Bgh. If leaf discs from Bgh-infected leaves were stained with DAF-FM DA, the resulting fluorescence intensities reflected visual differences in Bgh infection levels and were quantified as a measure for fungal growth (Supplemental Figure S1). To this end, the stained leaf discs were loaded onto 96-well plates and scanned by a spinning disc (confocal) microscope (Figure 3).
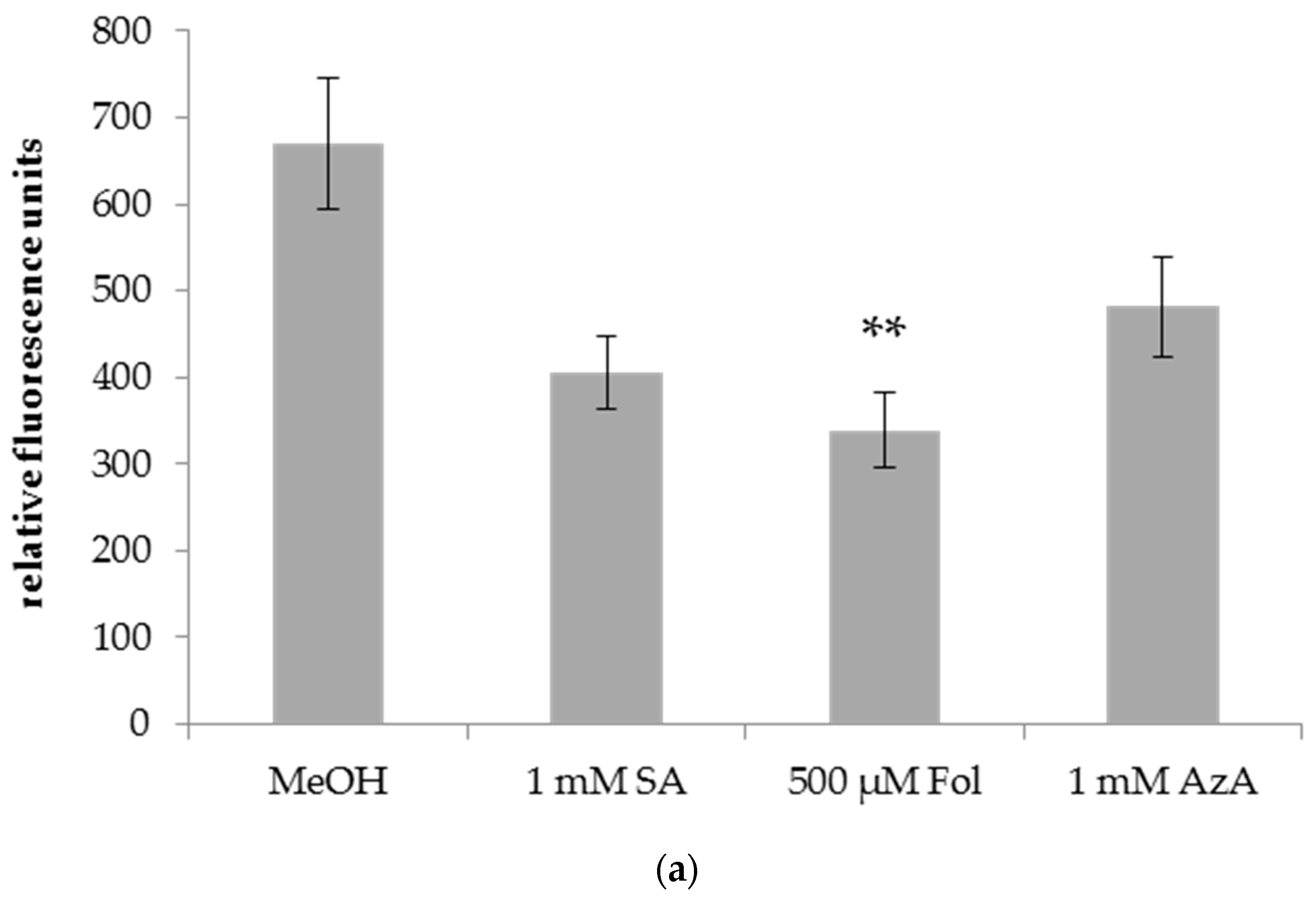
A local infiltration of Fol in leaf 1 of barley reduced Bgh growth on the Fol-treated leaf (Figure 4a) as well as on systemic leaf 2 of the treated plants (Figure 4b) as evidenced by a ~50% decrease in DAF-FM-DA fluorescence on the leaves of Bgh-infected Fol-treated compared to mock-treated plants. In contrast, SA and AzA infiltration did not induce significant changes in the DAF-FM-DA fluorescence of the treated leaves and thus had no local effect on Bgh growth (Figure 4a). Nevertheless, SA and AzA appeared to reduce Bgh propagation, although the differences to the control were not significant (p = 0.0518 in the case of SA, Figure 4a). In the systemic leaves, Bgh-associated DAF-FM-DA fluorescence was decreased by SA and AzA to a similar extent as by Fol (Figure 4b). Thus, Fol might induce local and systemic resistance to Bgh, while SA and AzA appear to trigger systemic resistance.
3.3. Arabidopsis Immunity-Related Compounds Do Not Alter Barley Susceptibility to P. Teres
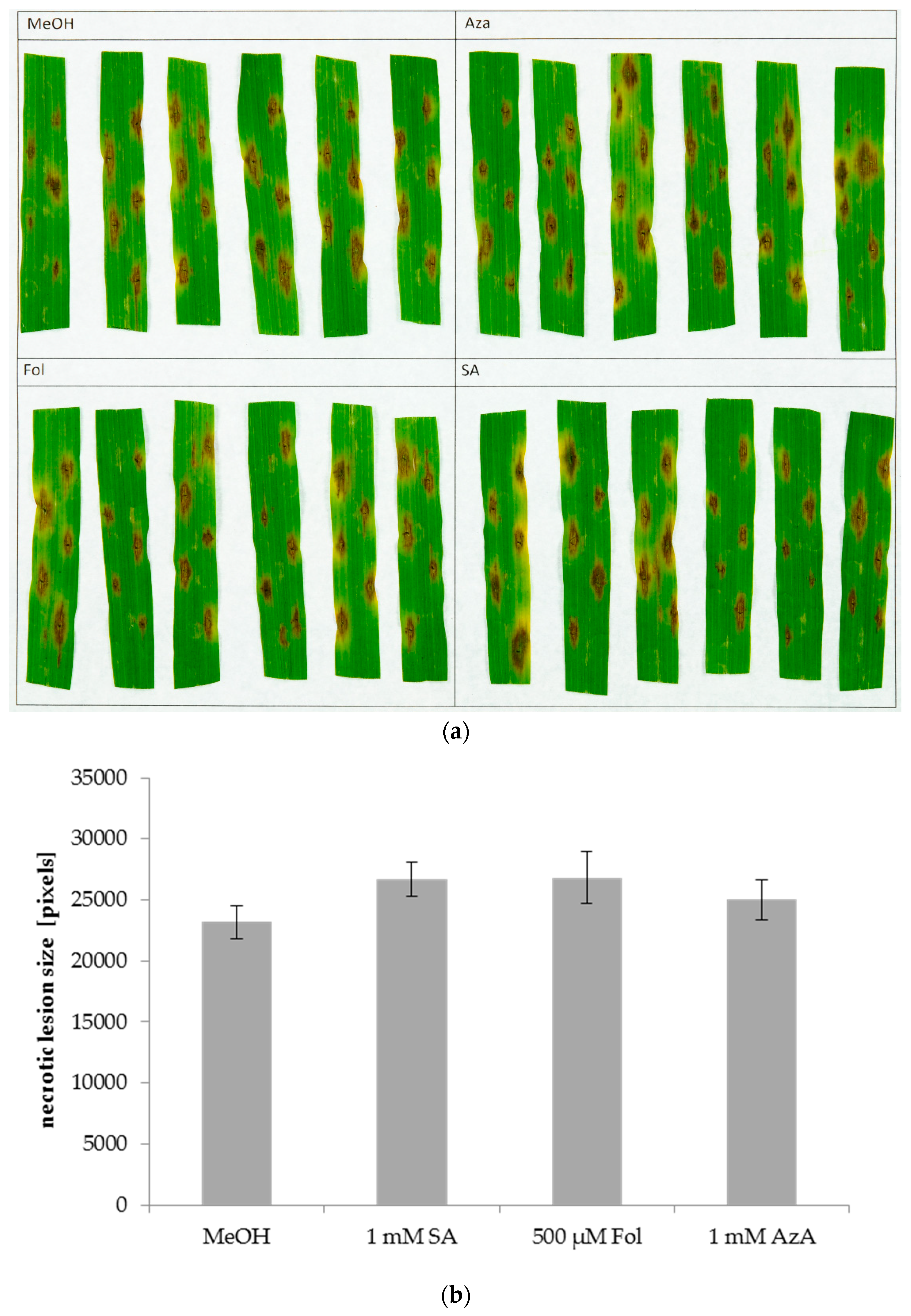
In Arabidopsis, SA induces local, but not systemic susceptibility to the necrotrophic fungal pathogen Alternaria brassicicola [35,63]. This most likely happens due to negative crosstalk between the SA and JA pathways, with SA inhibiting JA-mediated defence against A. brassicicola. A similar effect was observed for Fol, as its application increased the size of lesions caused by A. brassicicola on the treated, but not on systemic leaves [35]. Here, we examined the effects of SA, Fol, and AzA on barley local and systemic defence responses to the necrotrophic fungus P. teres. To this end, leaves of 3-week-old barley plants were syringe-infiltrated with 1 mM SA, 50 or 500 µM Fol, 1 mM AzA, or 0.025% MeOH as control treatment. The same or systemic leaves were inoculated with P. teres 1 or 5 days later, respectively, and necrotic lesions were measured at 4 dpi using the ImageJ macro PIDIQ [69], which was modified to recognize the brown lesion caused by P. teres. The outcome varied strongly between different replicate experiments. Strikingly, SA application caused increases in P. teres lesion sizes in 4 out of 8 experiments if leaves systemic to the site of SA treatment were inoculated. However, taking all data together SA, Fol, and AzA did not significantly influence P. teres lesion sizes either locally (Figure 5) or systemically (Supplemental Figure S2), suggesting that these compounds do not affect the susceptibility of barley to P. teres.
4. Discussion
4.1. Salicylic Acid Has Differential Effects on Pathogens with Different Lifestyles
Here, we report on the role of SA, Fol, and AzA on barley defence against pathogens with different lifestyles. The three tested compounds are involved in Arabidopsis SAR and known for inducing resistance to (hemi-)biotrophic pathogens in the model plant. In contrast, it has been reported that a local application of SA does not induce systemic resistance to the hemi-biotrophic bacterium Xtc in barley [47]. In order to find out if SA is involved in (systemic) defence responses in barley, we performed inoculations with two additional pathogens.
Although a local application of SA did not affect growth of the hemi-biotrophic bacterium Xtc in the systemic tissue, the same treatment reduced the disease burden of the biotrophic fungal pathogen Bgh in the systemic leaves by more than 50% (Figure 4). There might also be a local effect of SA on Bgh propagation, for which we observed a clear, but statistically insignificant trend. Thus, SA might enhance the resistance of barley to the powdery mildew pathogen Bgh. Similar to SA, its functional analogue BTH does not affect barley resistance to Xtc [47], but it induces resistance to Bgh [46]. Additionally, NPR1, the master regulator of SA responses in Arabidopsis [7,9,10], is important for barley defence responses against Bgh, but not against Xtc [47], further supporting a possible role of SA in barley defence against Bgh but not Xtc. Previous studies had reported only a minor effect on Bgh, if any, after SA treatment of barley plants [46,55]. The relatively robust effect of SA on systemic Bgh propagation in barley that we observe in this study can have one or more of three reasons. First, the apparent difference in Bgh burden between SA- and mock-treated plants might be exaggerated by the method used for evaluation of the Bgh infections. While other studies rely on pustule counts, we used the fluorescent dye DAF-FM-DA to quantify fungal material. It is known that barley produces NO as part of the plants early defence responses against Bgh [71]. Additionally, at certain stages of its life cycle the fungus itself produces NO [72]. However, it seems unlikely that this NO interferes with our quantification. The production of NO is very short-lived and happens mostly in early defence responses and early life stages of Bgh, whereas we stain with DAF-FM-DA at 6 dpi, a relatively late stage of the infection, at which time Bgh displays significant hyphal growth (Figure 3). Second, previous studies used soil-drench treatment for SA application [46,55] while in this study, syringe-infiltration of leaves was used. Finally, and perhaps most importantly, plant age differed between both prior and the current studies. In both of the cited publications, seedlings of 5 or 7 days of age were used while we worked with 3-week-old plants. It is known that plant age can positively affect plant resistance against Bgh [73]. In support of this hypothesis, we observed robust effects of SA and the other tested compounds on Bgh propagation and Xtc growth in 3-week-old plants, but did not reproducibly observe the same effects in 2-week-old plants. Comparing our data to those presented in [46,55], it is possible that SA more effectively enhances barley resistance to Bgh if applied directly to the leaves rather than the soil and/or if applied to 3-week-old rather than younger plants.
Although SA locally enhances the susceptibility of Arabidopsis plants to a necrotrophic fungal pathogen [35,63], it had no effect on P. teres lesion sizes either locally or systemically in barley (Figure 5 and Figure S2). Thus, the trade-off between plant defence responses to biotrophic and necrotrophic pathogens that is observed in Arabidopsis does not appear to influence growth of the necrotrophic fungus in SA-treated barley plants.
In summary, SA appears to induce systemic resistance against Bgh in barley but likely does not contribute to resistance against Xtc or P. teres.
4.2. Folic Acid Has Differential Effects against Bacteria and Fungi with Similar Lifestyles
Fol application is known to induce local and systemic resistance to hemi-biotrophic bacteria in Arabidopsis [35]. This effect is dependent on SA biosynthesis and signalling and on the SAR-associated compound glycerol-3-phosphate. Similar to SA, Fol application triggers local susceptibility to necrotrophic A. brassicicola. Here, we infiltrated or sprayed barley with Fol and monitored the effects on local and systemic propagation of bacterial and fungal barley pathogens.
In contrast to SA, Fol application enhanced barley susceptibility to Xtc both systemically and to a lesser extent also in the local treated tissue (Figure 1 and Figure 2). In Arabidopsis, Fol enhances resistance, probably through the SA pathway [35]. Here, SA and Fol differentially affected Xtc growth in barley, which was not affected by SA [47] and was enhanced (rather than reduced) by Fol. It is conceivable that Xtc can take up Fol, which is needed for nucleotide biosynthesis. Such supplementation could directly enhance bacterial growth, mimicking the induction of plant susceptibility. However, because the effect of Fol application on Xtc growth was stronger in the systemic compared to the local, treated tissue (Figure 1 and Figure 2), the data argue in favour of a Fol-induced effect on plant immunity.
Similar to SA, the application of Fol reduced Bgh propagation on barley both locally and systemically but did not affect P. teres lesion formation (Figure 4 and Figure 5). Again, the effects of SA and Fol on Bgh propagation were more pronounced in tissues that were systemic to the site of SA/Fol treatment than in the treated leaves themselves. Although we cannot exclude direct effects of the compounds on fungal growth, the data argue in favour of SA- and Fol-induced plant defence mechanisms affecting Bgh propagation in barley. Importantly, Fol-related compounds can promote plant yield [36], whereas SA causes cell death when applied at high concentrations. Thus, although adverse effects on barley susceptibility to hemi-biotrophic bacteria such as Xtc should be considered, Fol could be used as an alternative to SA or BTH to enhance the resistance of barley to the economically relevant powdery mildew pathogen Bgh.
4.3. Azelaic Acid Moderately Affects Barley Defence Responses
Application of AzA to Arabidopsis confers local and systemic resistance to hemi-biotrophic bacteria [19,30,31]. AzA primes Arabidopsis to accumulate higher SA levels more quickly after a subsequent infection [19]. Similarly to Fol-induced responses in Arabidopsis [35], AzA-mediated SAR depends on SA [19]. Here, we found that AzA influences pathogen propagation in barley very similarly to Fol. Whereas Fol locally and systemically enhanced Xtc growth, AzA did the same only in the local treated tissue and not systemically (Figure 1b and Figure 2). Nevertheless, this similarity in the effects of Fol and AzA on barley susceptibility to Xtc argues for a possible interference of these compounds with the barley defence response to Xtc rather than for direct effects of either compound on bacterial growth. Furthermore, in contrast to its effect on Xtc growth, AzA reduced Bgh propagation systemically but not locally (Figure 4). In this case, the systemic response induced by AzA is similar to the responses induced by SA and Fol, which also appear to induce systemic resistance to Bgh. Similar to SA and Fol, AzA application did not have an influence on fungal growth of P. teres (Figure 5 and Figure S2), neither in local treated nor in systemic tissues.
In Arabidopsis, SAR appears to be regulated by two parallel signalling pathways that are inter-dependent. One of these pathways depends on SA, the other one on AzA, G3P, reactive oxygen species, and nitric oxide [26,74]. While the SA pathway seems to be effective in barley at least against Bgh ([46] and Figure 4), the function of the other pathway, if existent in barley, is still unclear. Since SA did not affect barley resistance to Xtc while both Fol and to a minor extent AzA enhanced susceptibility rather than immunity to this hemi-biotrophic pathogen, it is possible that Fol and AzA influenced a SA-independent immune pathway. Such a pathway might rely on JA and/or ABA which are positively associated with barley defence against Xtc [47].
5. Conclusions
In this study, we investigated the role of resistance-inducing compounds from Arabidopsis, salicylic acid (SA), folic acid, and azelaic acid, on barley defence against the pathogens Xanthomonas translucens, Bgh (powdery mildew), and Pyrenophora teres. Azelaic acid appeared to induce local susceptibility to X. translucens and at the same time systemic resistance to powdery mildew. Also, we observed a possible activation of local and/or systemic resistance to powdery mildew after application of SA and folic acid. Because folic acid and azelaic acid, which both enhance SA-mediated immune responses in Arabidopsis, both enhance barley resistance to powdery mildew similarly to SA, the associated barley immune response might be related to SA. Importantly, the data show that folic acid- and azelaic acid-induced resistance is a double-edged sword that can at the same time induce resistance and susceptibility against different pathogens (powdery mildew and X. translucens). Also, folic acid and azelaic acid differentially influence the responses of Arabidopsis and barley to host-adapted hemi-biotrophic bacterial pathogens. Thus, investigating induced defence responses in barley and studying the signalling pathways used to achieve resistance in this cereal crop will be challenging topics for future research.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/8/8/142/s1, Figure S1: Fol induces resistance to Bgh, Figure S2: Size of P. teres lesions on the systemic leaves of barley after local application of SA, 50 µM Fol, 500 µM Fol, or AzA.
Author Contributions
Conceptualization, M.L., M.W. and A.C.V.; Data curation, M.L., M.W., F.M. and F.H.; Funding acquisition, A.C.V.; Investigation, M.L., M.W., F.M. and F.H.; Methodology, M.L., M.W. and F.M.; Supervision, A.C.V.; Writing—original draft, M.L.; Writing—review & editing, A.C.V.
Funding
This work was funded by the Deutsche Forschungsgemeinschaft (DFG) as part of SFB924 (Project B06 to A.C.V.) and by the German Federal Ministry of Education and Research as part of the German Plant Phenotyping Network (DPPN, no. 031A053C).
Acknowledgments
We thank Ralph Hückelhoven for helpful discussion, the Patrick Schweizer lab (IPK Gatersleben) for Bgh and advice on inoculation methods, Eva Rudolf for help with statistics, and Florian Hug, Claudia Knappe, and Sanjukta Dey for their help with and valuable input on the experiments in this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Panstruga, R.; Parker, J.E.; Schulze-Lefert, P. SnapShot: Plant Immune Response Pathways. Cell 2009, 136, 978.e1–978.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Felix, G. Plants and animals: A different taste for microbes? Curr. Opin. Plant Biol. 2005, 8, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.; Yadeta, K.A.; Coaker, G. Recognition of bacterial plant pathogens: Local, systemic and transgenerational immunity. New Phytol. 2013, 199, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Klessig, D.F.; Choi, H.W.; Dempsey, M.A. Systemic Acquired Resistance and Salicylic Acid: Past, Present and Future. Mol. Plant-Microbe Interact. 2018. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a Multifaceted Hormone to Combat Disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic Acquired Resistance: Turning Local Infection into Global Defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Pajerowska-Mukhtar, K.M.; Emerine, D.K.; Mukhtar, M.S. Tell me more: Roles of NPRs in plant immunity. Trends Plant Sci. 2013, 18, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, J.; Chaturvedi, R.; Chowdhury, Z.; Venables, B.; Petros, R.A. Signaling by small metabolites in systemic acquired resistance. Plant J. 2014, 79, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting Ready for Battle. Mol. Plant-Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourbal, B.; Pinaud, S.; Beckers, G.J.M.; Van Der Meer, J.W.M.; Conrath, U.; Netea, M.G. Innate immune memory: An evolutionary perspective. Immunol. Rev. 2018, 283, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Riedlmeier, M.; Ghirardo, A.; Wenig, M.; Knappe, C.; Koch, K.; Georgii, E.; Dey, S.; Parker, J.E.; Schnitzler, J.-P.; Vlot, A.C. Monoterpenes Support Systemic Acquired Resistance within and between Plants. Plant Cell 2017, 29, 1440–1459. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Chanda, B.; Xia, Y.; Mandal, M.K.; Yu, K.; Sekine, K.T.; Gao, Q.M.; Selote, D.; Hu, Y.; Stromberg, A.; Navarre, D.; et al. Glycerol-3-phosphate is a critical mobile inducer of systemic immunity in plants. Nat. Genet. 2011, 43, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, R.; Lim, G.-H.; De Lorenzo, L.; Yu, K.; Zhang, K.; Hunt, A.G.; Kachroo, A.; Kachroo, P. Pipecolic acid confers systemic immunity by regulating free radicals. Sci. Adv. 2018, 4, eaar4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.W.; Tschaplinski, T.J.; Wang, L.; Glazebrook, J.; Greenberg, J.T. Priming in Systemic Plant Immunity. Science 2009, 324, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Venables, B.; Petros, R.A.; Nalam, V.; Li, M.; Wang, X.; Takemoto, L.J.; Shah, J. An abietane diterpenoid is a potent activator of systemic acquired resistance. Plant J. 2012, 71, 161–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, M.; Zeier, T.; Bernsdorff, F.; Reichel-Deland, V.; Kim, D.; Hohmann, M.; Scholten, N.; Schuck, S.; Bräutigam, A.; Hölzel, T.; et al. Flavin Monooxygenase-Generated N-Hydroxypipecolic Acid Is a Critical Element of Plant Systemic Immunity. Cell 2018, 173, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Holmes, E.C.; Rajniak, J.; Kim, J.-G.; Tang, S.; Fischer, C.R.; Mudgett, M.B.; Sattely, E.S. N-hydroxy-pipecolic acid is a mobile metabolite that induces systemic disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E4920–E4929. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.M.; Doerner, P.; Dixon, R.A.; Lamb, C.J.; Cameron, R.K. A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 2002, 419, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Champigny, M.J.; Isaacs, M.; Carella, P.; Faubert, J.; Fobert, P.R.; Cameron, R.K. Long distance movement of DIR1 and investigation of the role of DIR1-like during systemic acquired resistance in Arabidopsis. Front. Plant Sci. 2013, 4, 230. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Pabst, E.; Riedlmeier, M. Systemic Signalling in Plant Defence. eLS 2017, 1–9. [Google Scholar] [CrossRef]
- Gao, Q.-M.; Zhu, S.; Kachroo, P.; Kachroo, A. Signal regulators of systemic acquired resistance. Front. Plant Sci. 2015, 6, 228. [Google Scholar] [CrossRef] [PubMed]
- Métraux, J.P.; Signer, H.; Ryals, J.; Ward, E.; Wyss-Benz, M.; Gaudin, J.; Raschdorf, K.; Schmid, E.; Blum, W.; Inverardi, B. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science 1990, 250, 1004–1006. [Google Scholar] [CrossRef] [PubMed]
- Navarova, H.; Bernsdorff, F.; Doring, A.-C.; Zeier, J. Pipecolic Acid, an Endogenous Mediator of Defense Amplification and Priming, Is a Critical Regulator of Inducible Plant Immunity. Plant Cell 2012, 24, 5123–5141. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.H.; Shine, M.B.; de Lorenzo, L.; Yu, K.; Cui, W.; Navarre, D.; Hunt, A.G.; Lee, J.Y.; Kachroo, A.; Kachroo, P. Plasmodesmata Localizing Proteins Regulate Transport and Signaling during Systemic Acquired Immunity in Plants. Cell Host Microbe 2016, 19, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Soares, J.; Mandal, M.K.; Wang, C.; Chanda, B.; Gifford, A.N.; Fowler, J.S.; Navarre, D.; Kachroo, A.; Kachroo, P. A feed-back regulatory loop between glycerol-3-phosphate and lipid transfer proteins DIR1 and AZI1 mediates azelaic acid-induced systemic immunity. Cell Rep. 2013, 3, 1266–1278. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, N.M.; Steffes, K.; Schläppi, M.R.; Gifford, A.N.; Greenberg, J.T. Arabidopsis AZI1 family proteins mediate signal mobilization for systemic defence priming. Nat. Commun. 2015, 6, 7658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meuwly, P.; Molders, W.; Buchala, A.; Metraux, J.-P. Local and Systemic Biosynthesis of Salicylic Acid in lnfected Cucumber Plants. Plant Phisiology 1995, 109, 1107–1114. [Google Scholar] [CrossRef]
- Vernooij, B.; Friedrichya, L.; Reist, R.; Kolditzjawhar, R.; Ward, E.; Uknes, S.; Kessmann, H.; Ryals, J. Salicylic Acid Is Not the Translocated Signal Responsible for lnducing Systemic Acquired Resistance but Is Required in Signal Transduction. Plant Cell 1994, 6, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Ding, P.; Rekhter, D.; Ding, Y.; Feussner, K.; Busta, L.; Haroth, S.; Xu, S.; Li, X.; Jetter, R.; Feussner, I.; et al. Characterization of a Pipecolic Acid Biosynthesis Pathway Required for Systemic Acquired Resistance. Plant Cell 2016, 28, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Wittek, F.; Kanawati, B.; Wenig, M.; Hoffmann, T.; Franz-Oberdorf, K.; Schwab, W.; Schmitt-Kopplin, P.; Vlot, A.C. Folic acid induces salicylic acid-dependent immunity in Arabidopsis and enhances susceptibility to Alternaria brassicicola. Mol. Plant Pathol. 2015, 16, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Song, G.C.; Choi, H.K.; Ryu, C.M. The folate precursor para-aminobenzoic acid elicits induced resistance against Cucumber mosaic virus and Xanthomonas axonopodis. Ann. Bot. 2013, 111, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Balmer, D.; Planchamp, C.; Mauch-Mani, B. On the move: Induced resistance in monocots. J. Exp. Bot. 2013, 64, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; De Vleesschauwer, D.; Sharma, M.K.; Ronald, P.C. Recent advances in dissecting stress-regulatory crosstalk in rice. Mol. Plant 2013, 6, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yi, G.; Peng, X.; Huang, B.; Liu, E.; Zhang, J. Systemic acquired resistance in Cavendish banana induced by infection with an incompatible strain of Fusarium oxysporum f. sp. cubense. J. Plant Physiol. 2013, 170, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, J.; Liu, P.; Xing, H.; Li, C.; Wei, G.; Kang, Z. Glycerol-3-phosphate metabolism in wheat contributes to systemic acquired resistance against Puccinia striiformis f. sp. tritici. PLoS ONE 2013, 8, e81756. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Liu, P.; Xue, Q.; Ji, C.; Qi, T.; Guo, J.; Guo, J.; Kang, Z. TaDIR1-2, a Wheat Ortholog of Lipid Transfer Protein AtDIR1 Contributes to Negative Regulation of Wheat Resistance against Puccinia striiformis f. sp. tritici. Front. Plant Sci. 2017, 8, 521. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Liu, P.; Ding, Y.; Mu, X.; Liu, X.; Wang, X.; Zhao, M.; Huai, B.; Huang, L.; et al. TaRar1 Is Involved in Wheat Defense against Stripe Rust Pathogen Mediated by YrSu. Front. Plant Sci. 2017, 8, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balmer, D.; De Papajewski, D.V.; Planchamp, C.; Glauser, G.; Mauch-Mani, B. Induced resistance in maize is based on organ-specific defence responses. Plant J. 2013, 74, 213–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, S.W.; Vernooij, B.; Titatarn, S.; Starrett, M.; Thomas, S.; Wiltse, C.C.; Frederiksen, R.A.; Bhandhufalck, A.; Hulbert, S.; Uknes, S. Induced Resistance Responses in Maize. Mol. Plant-Microbe Interact. 1998, 11, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Görlach, J.; Volrath, S.; Knauf-Beiter, G.; Hengy, G.; Beckhove, U.; Kogel, K.; Oostendorp, M.; Staub, T.; Ward, E.; Kessmann, H.; et al. Benzothiadiazole, a Novel Class of Inducers of Systemic Acquired Resistance, Activates Gene Expression and Disease Resistance in Wheat. Plant Cell 1996, 8, 629–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beßer, K.; Jarosch, B.; Langen, G.; Kogel, K.-H. Expression analysis of genes induced in barley after chemical activation reveals distinct disease resistance pathways. Mol. Plant Pathol. 2000, 1, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Wenig, M.; Langen, G.; Sharma, S.; Kugler, K.G.; Knappe, C.; Hause, B.; Bichlmeier, M.; Babaeizad, V.; Imani, J.; et al. Bacteria-Triggered Systemic Immunity in Barley Is Associated with WRKY and ETHYLENE RESPONSIVE FACTORs But Not with Salicylic Acid. Plant Physiol. 2014, 166, 2133–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, C.; Korell, M.; Eckey, C.; Biedenkopf, D.; Kogel, K.-H. Identification and transcriptional analysis of powdery mildew-induced barley genes. Plant Sci. 2005, 168, 373–380. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Gregersen, P.L.; Collinge, D.B. The Barley/Blumeria (Syn. Erysiphe) Graminis Interaction. In Mechanisms of Resistance to Plant Diseases; Springer: Dordrecht, The Netherlands, 2000; pp. 77–100. [Google Scholar]
- Hückelhoven, R.; Panstruga, R. Cell biology of the plant-powdery mildew interaction. Curr. Opin. Plant Biol. 2011, 14, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallelian-Bindschedler, L.; Mösinger, E.; Métraux, J.P.; Schweizer, P. Structure, expression and localization of a germin-like protein in barley (Hordeum vulgare L.) that is insolubilized in stressed leaves. Plant Mol. Biol. 1998, 37, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Hückelhoven, R.; Fodor, J.; Preis, C.; Kogel, K.-H. Hypersensitive Cell Death and Papilla Formation in Barley Attacked by the Powdery Mildew Fungus Are Associated with Hydrogen Peroxide but Not with Salicylic Acid Accumulation1. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Langen, G.; Hess, W.; Börner, T.; Hückelhoven, R.; Kogel, K.-H. The white barley mutant albostrians shows enhanced resistance to the biotroph Blumeria graminis f. sp. hordei. Mol. Plant-Microbe Interact. 2004, 17, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Kogel, K.H.; Ortel, B.; Jarosch, B.; Atzorn, R.; Schiffer, R.; Wasternack, C. Resistance in barley against the powdery mildew fungus (Erysiphe graminis f. sp. hordei) is not associated with enhanced levels of endogenous jasmonates. Eur. J. Plant Pathol. 1995, 101, 319–332. [Google Scholar] [CrossRef]
- Cui, J.; Bahrami, A.K.; Pringle, E.G.; Hernandez-Guzman, G.; Bender, C.L.; Pierce, N.E.; Ausubel, F.M. Pseudomonas syringae manipulates systemic plant defenses against pathogens and herbivores. Proc. Natl. Acad. Sci. USA 2005, 102, 1791–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, M.; Ishikawa, A.; Jikumaru, Y.; Seki, M.; Umezawa, T.; Asami, T.; Maruyama-Nakashita, A.; Kudo, T.; Shinozaki, K.; Yoshida, S.; et al. Antagonistic Interaction between Systemic Acquired Resistance and the Abscisic Acid-Mediated Abiotic Stress Response in Arabidopsis. Plant Cell 2008, 20, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Shigenaga, A.M.; Argueso, C.T. No hormone to rule them all: Interactions of plant hormones during the responses of plants to pathogens. Semin. Cell Dev. Biol. 2016, 56, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone Crosstalk in Plant Disease and Defense: More Than Just JASMONATE-SALICYLATE Antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; An, F.; Feng, Y.; Li, P.; Xue, L.; Mu, A.; Jiang, Z.; Kim, J.-M.; To, T.K.; Li, W.; et al. Derepression of ethylene-stabilized transcription factors (EIN3/EIL1) mediates jasmonate and ethylene signaling synergy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 12539–12544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckers, G.J.M.; Spoel, S.H. Fine-tuning plant defence signalling: Salicylate versus jasmonate. Plant Biol. 2006, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Johnson, J.S.; Dong, X. Regulation of tradeoffs between plant defenses against pathogens with different lifestyles. Proc. Natl. Acad. Sci. USA 2007, 104, 18842–18847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ellwood, S.R.; Oliver, R.P.; Friesen, T.L. Pyrenophora teres: Profile of an increasingly damaging barley pathogen. Mol. Plant Pathol. 2011, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Vlot, A.C.; Liu, P.P.; Cameron, R.K.; Park, S.W.; Yang, Y.; Kumar, D.; Zhou, F.; Padukkavidana, T.; Gustafsson, C.; Pichersky, E.; et al. Identification of likely orthologs of tobacco salicylic acid-binding protein 2 and their role in systemic acquired resistance in Arabidopsis thaliana. Plant J. 2008, 56, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Delventhal, R.; Rajaraman, J.; Stefanato, F.L.; Rehman, S.; Aghnoum, R.; McGrann, G.R.D.; Bolger, M.; Usadel, B.; Hedley, P.E.; Boyd, L.; et al. A comparative analysis of nonhost resistance across the two Triticeae crop species wheat and barley. BMC Plant Biol. 2017, 17, 232. [Google Scholar] [CrossRef] [PubMed]
- Nair, K.R.S.; Ellingboe, A.H. A method of controlled inoculations with conidiospores of Erysiphe graminis var. tritici. Phytopathology 1962, 52, 714. [Google Scholar]
- Foissner, I.; Wendehenne, D.; Langebartels, C.; Durner, J. In vivo imaging of elicitor-induced nitric oxide burst in tobacco. Plant J. 2000, 23, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, B.; Middleton, M.; Lo, T.; Desveaux, D.; Guttman, D.S. Image-Based Quantification of Plant Immunity and Disease. Mol. Plant-Microbe Interact. 2016, 29, MPMI-07-16-0129. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Hirata, Y.; Nagano, T. Fluorescent indicators for imaging nitric oxide production. Angew. Chem. Int. Ed. 1999, 38, 3209–3212. [Google Scholar] [CrossRef]
- Prats, E.; Mur, L.A.J.; Sanderson, R.; Carver, T.L.W. Nitric oxide contributes both to papilla-based resistance and the hypersensitive response in barley attacked by Blumeria graminis f. sp. hordei. Mol. Plant Pathol. 2005, 6, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Prats, E.; Carver, T.L.W.; Mur, L.A.J. Pathogen-derived nitric oxide influences formation of the appressorium infection structure in the phytopathogenic fungus Blumeria graminis. Res. Microbiol. 2008, 159, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-R.; Edwards, H.H. Primary Penetration Process in Powdery Mildewed Barley Related to Host Cell Age, Cell Type, and Occurrence of Basic Staining Material. New Phytol. 1974, 73, 131–137. [Google Scholar] [CrossRef]
- Wendehenne, D.; Gao, Q.; Kachroo, A.; Kachroo, P. Free radical-mediated systemic immunity in plants. Curr. Opin. Plant Biol. 2014, 20, 127–134. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Xanthomonas translucens pv. cerealis (Xtc) titres in the systemic leaves of barley after local application of folic acid (Fol) or azelaic acid (AzA). Barley cultivar Golden Promise (GP) plants were infiltrated in the first true leaf with 0.025% methanol (MeOH) as control, 500 µM Fol (a), 1 mM AzA (b), or 50 µM Fol (c) in 10 mM MgCl2 as indicated below the panels. Five days later, the second true leaves of the plants were inoculated with Xtc by syringe infiltration. The resulting Xtc titres in leaf 2 at 4 dpi are shown. Bars represent the average of 18–36 replicates from 5 ((a): 4× greenhouse, 1× growth chamber; (b): 3× greenhouse, 2× growth chamber) to 9 ((c): 8× greenhouse, 1× growth chamber) biologically independent experiments ± standard error; replicates were as follows: (a) MeOH: 19 (4 + 4 + 3 + 4 + 4), Fol: 18 (4 + 4 + 3 + 4 + 3), (b) MeOH: 19 (4 + 3 + 3 + 4 + 5), AzA: 20 (4 + 4 + 4 + 4 + 4), (c) MeOH: 34 (4 + 3 + 4 + 4 + 3 + 4 + 4 + 4 + 4), Fol: 36 (4 + 4 + 4 + 4 + 4 + 4 + 4 + 4 + 4). Asterisks above bars indicate statistically significant differences from the control treatments (t-test; * p < 0.05, ** p < 0.005).
Figure 1.
Xanthomonas translucens pv. cerealis (Xtc) titres in the systemic leaves of barley after local application of folic acid (Fol) or azelaic acid (AzA). Barley cultivar Golden Promise (GP) plants were infiltrated in the first true leaf with 0.025% methanol (MeOH) as control, 500 µM Fol (a), 1 mM AzA (b), or 50 µM Fol (c) in 10 mM MgCl2 as indicated below the panels. Five days later, the second true leaves of the plants were inoculated with Xtc by syringe infiltration. The resulting Xtc titres in leaf 2 at 4 dpi are shown. Bars represent the average of 18–36 replicates from 5 ((a): 4× greenhouse, 1× growth chamber; (b): 3× greenhouse, 2× growth chamber) to 9 ((c): 8× greenhouse, 1× growth chamber) biologically independent experiments ± standard error; replicates were as follows: (a) MeOH: 19 (4 + 4 + 3 + 4 + 4), Fol: 18 (4 + 4 + 3 + 4 + 3), (b) MeOH: 19 (4 + 3 + 3 + 4 + 5), AzA: 20 (4 + 4 + 4 + 4 + 4), (c) MeOH: 34 (4 + 3 + 4 + 4 + 3 + 4 + 4 + 4 + 4), Fol: 36 (4 + 4 + 4 + 4 + 4 + 4 + 4 + 4 + 4). Asterisks above bars indicate statistically significant differences from the control treatments (t-test; * p < 0.05, ** p < 0.005).
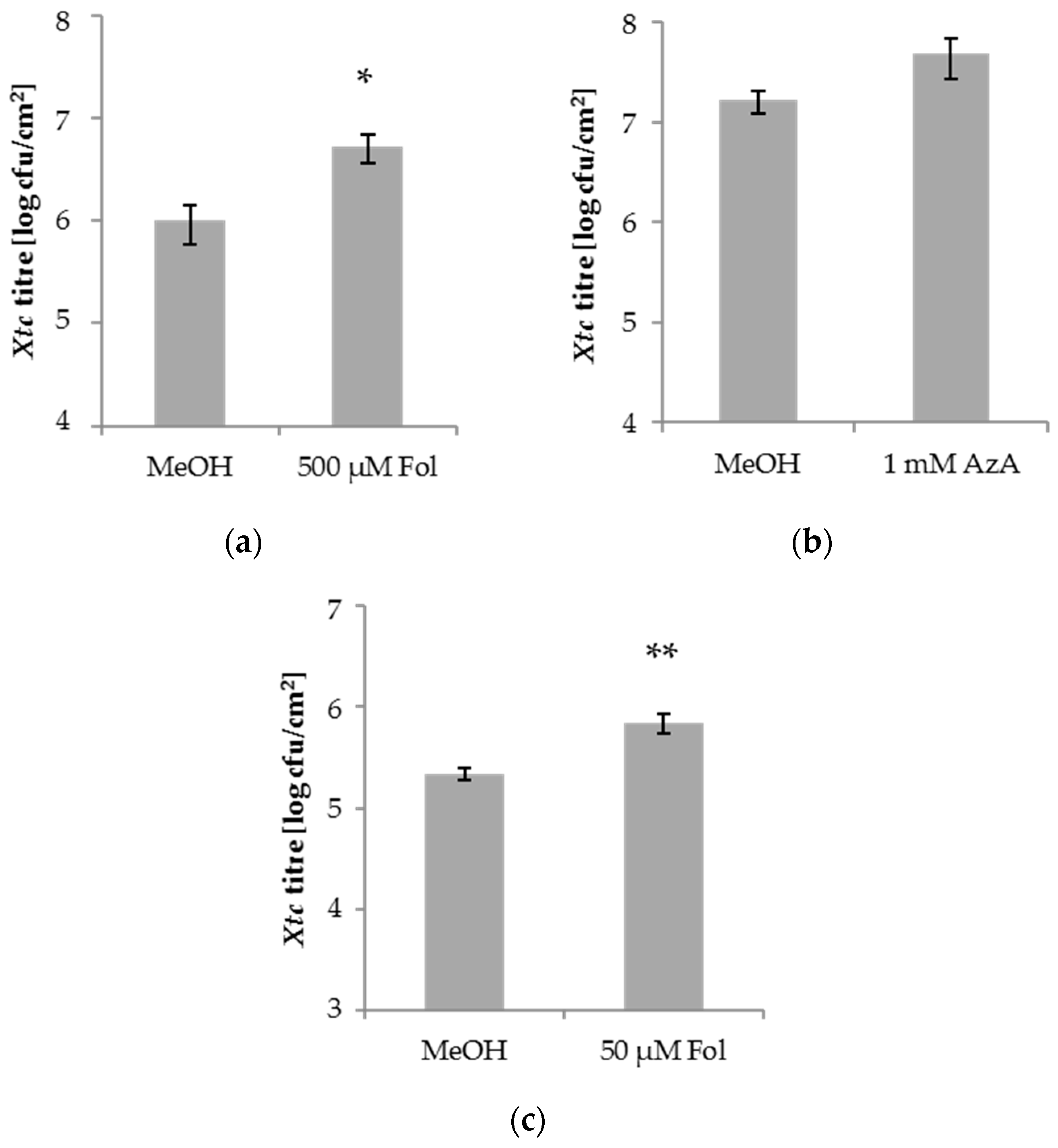
Figure 2.
Xtc titres in salicylic acid (SA)-, Fol- and AzA-treated barley plants. Barley cultivar GP plants were sprayed with 0.05% MeOH as control, 1 mM SA, 500 µM Fol, or 1 mM AzA in 0.01% Tween-20 as indicated below the panel. One day later, the second true leaves of the plants were inoculated with Xtc by syringe infiltration. The resulting Xtc titres in leaf 2 at 4 dpi are shown. Bars represent the average of 8–9 replicates from 3 biologically independent greenhouse experiments ± standard error, replicates were as follows: MeOH: 9 (3 + 3 + 3), SA: 9 (3 + 3 + 3), Fol: 8 (3 + 3 + 2), AzA: 8 (3 + 3 + 2); asterisks above bars indicate statistically significant differences from the mock control treatment (one-way ANOVA and post hoc Dunnett’s test, * p < 0.05, *** p < 0.0005).
Figure 2.
Xtc titres in salicylic acid (SA)-, Fol- and AzA-treated barley plants. Barley cultivar GP plants were sprayed with 0.05% MeOH as control, 1 mM SA, 500 µM Fol, or 1 mM AzA in 0.01% Tween-20 as indicated below the panel. One day later, the second true leaves of the plants were inoculated with Xtc by syringe infiltration. The resulting Xtc titres in leaf 2 at 4 dpi are shown. Bars represent the average of 8–9 replicates from 3 biologically independent greenhouse experiments ± standard error, replicates were as follows: MeOH: 9 (3 + 3 + 3), SA: 9 (3 + 3 + 3), Fol: 8 (3 + 3 + 2), AzA: 8 (3 + 3 + 2); asterisks above bars indicate statistically significant differences from the mock control treatment (one-way ANOVA and post hoc Dunnett’s test, * p < 0.05, *** p < 0.0005).
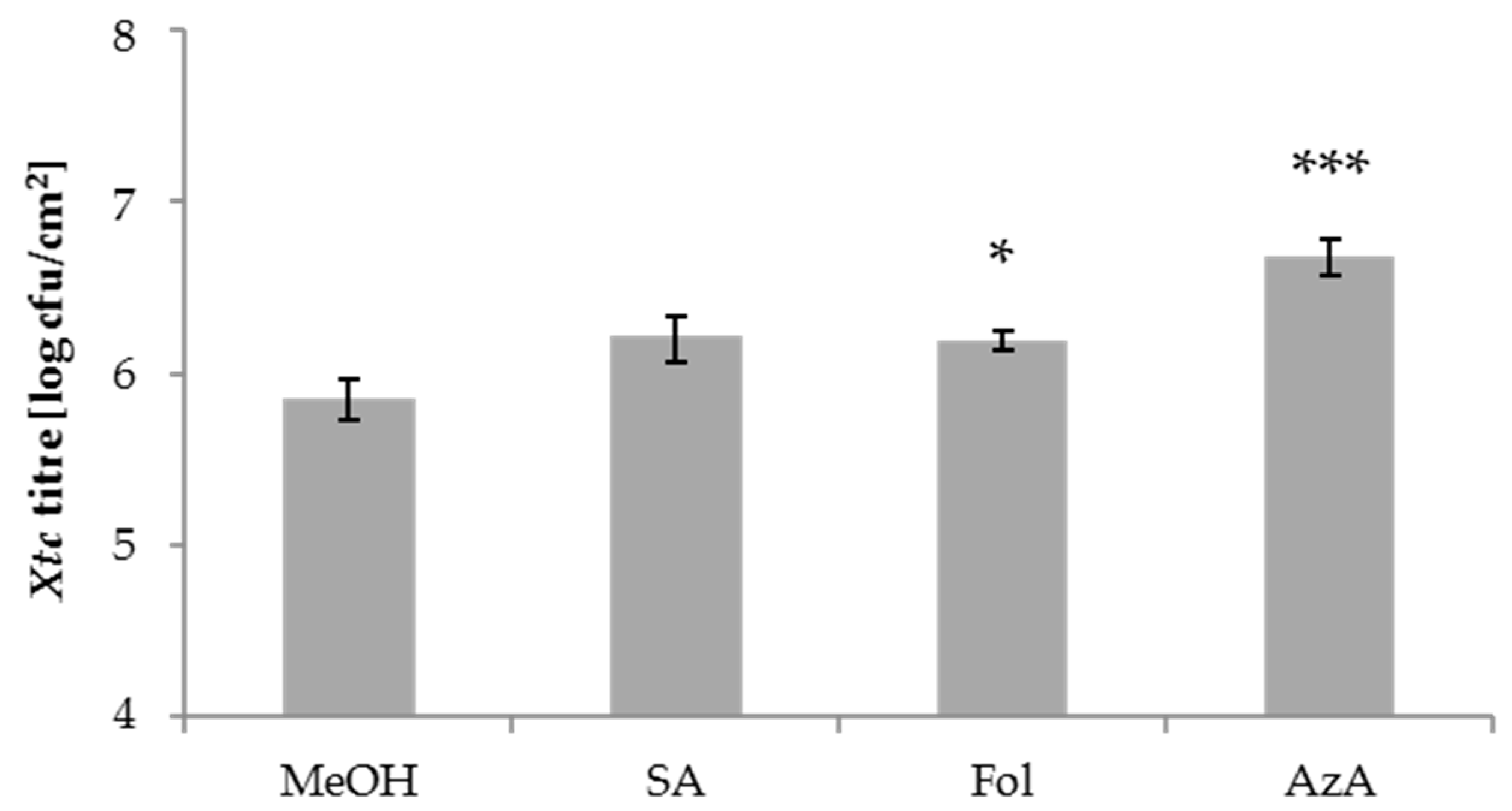
Figure 3.
Merged z-stack 3 × 3 tiled images of 4-amino-5-methylamino-2’,7’-difluorofluorescein diacetate (DAF-FM-DA) stained discs of Blumeria graminis f. sp. hordei (Bgh)-infected barley leaves in the first 3 columns of a 96-well plate. Enlarged: merged image of a single well. Chlorophyll fluorescence is shown in red, DAF-FM DA fluorescence in green.
Figure 3.
Merged z-stack 3 × 3 tiled images of 4-amino-5-methylamino-2’,7’-difluorofluorescein diacetate (DAF-FM-DA) stained discs of Blumeria graminis f. sp. hordei (Bgh)-infected barley leaves in the first 3 columns of a 96-well plate. Enlarged: merged image of a single well. Chlorophyll fluorescence is shown in red, DAF-FM DA fluorescence in green.

Figure 4.
Relative fluorescence of DAF-FM DA staining of Bgh in leaves of barley plants after application of SA, Fol, or AzA to leaf 1. Barley cultivar GP plants were infiltrated in the first true leaf with 0.025% MeOH as control, 1 mM SA, 500 µM Fol, or 1 mM AzA in 10 mM MgCl2 as indicated below the panel. Five days later, the plants were infected with Bgh spores. Leaf discs were cut out of the first (local) (a) and second (systemic) (b) true leaf and stained with DAF-FM DA. Fluorescence was recorded using a spinning disc (confocal) microscope. Bars represent the average of 37–64 replicates from 2 (SA treatment in b) to 3 (all other treatments) independent experiments ± standard error; replicates were as follows: (a) MeOH: 57 (22 + 14 + 21), SA: 62 (22 + 16 + 24), Fol: 55 (19 + 19 + 17), AzA: 58 (19 + 18 + 21); (b) MeOH: 55 (20 + 22 + 13), SA: 37 (17 + 20), Fol: 64 (21 + 23 + 20), AzA: 53 (6 + 24 + 23). Asterisks above the bars indicate statistically significant differences from the mock control treatment (one-way ANOVA and poct hoc Dunnett’s test, ** p < 0.005).
Figure 4.
Relative fluorescence of DAF-FM DA staining of Bgh in leaves of barley plants after application of SA, Fol, or AzA to leaf 1. Barley cultivar GP plants were infiltrated in the first true leaf with 0.025% MeOH as control, 1 mM SA, 500 µM Fol, or 1 mM AzA in 10 mM MgCl2 as indicated below the panel. Five days later, the plants were infected with Bgh spores. Leaf discs were cut out of the first (local) (a) and second (systemic) (b) true leaf and stained with DAF-FM DA. Fluorescence was recorded using a spinning disc (confocal) microscope. Bars represent the average of 37–64 replicates from 2 (SA treatment in b) to 3 (all other treatments) independent experiments ± standard error; replicates were as follows: (a) MeOH: 57 (22 + 14 + 21), SA: 62 (22 + 16 + 24), Fol: 55 (19 + 19 + 17), AzA: 58 (19 + 18 + 21); (b) MeOH: 55 (20 + 22 + 13), SA: 37 (17 + 20), Fol: 64 (21 + 23 + 20), AzA: 53 (6 + 24 + 23). Asterisks above the bars indicate statistically significant differences from the mock control treatment (one-way ANOVA and poct hoc Dunnett’s test, ** p < 0.005).
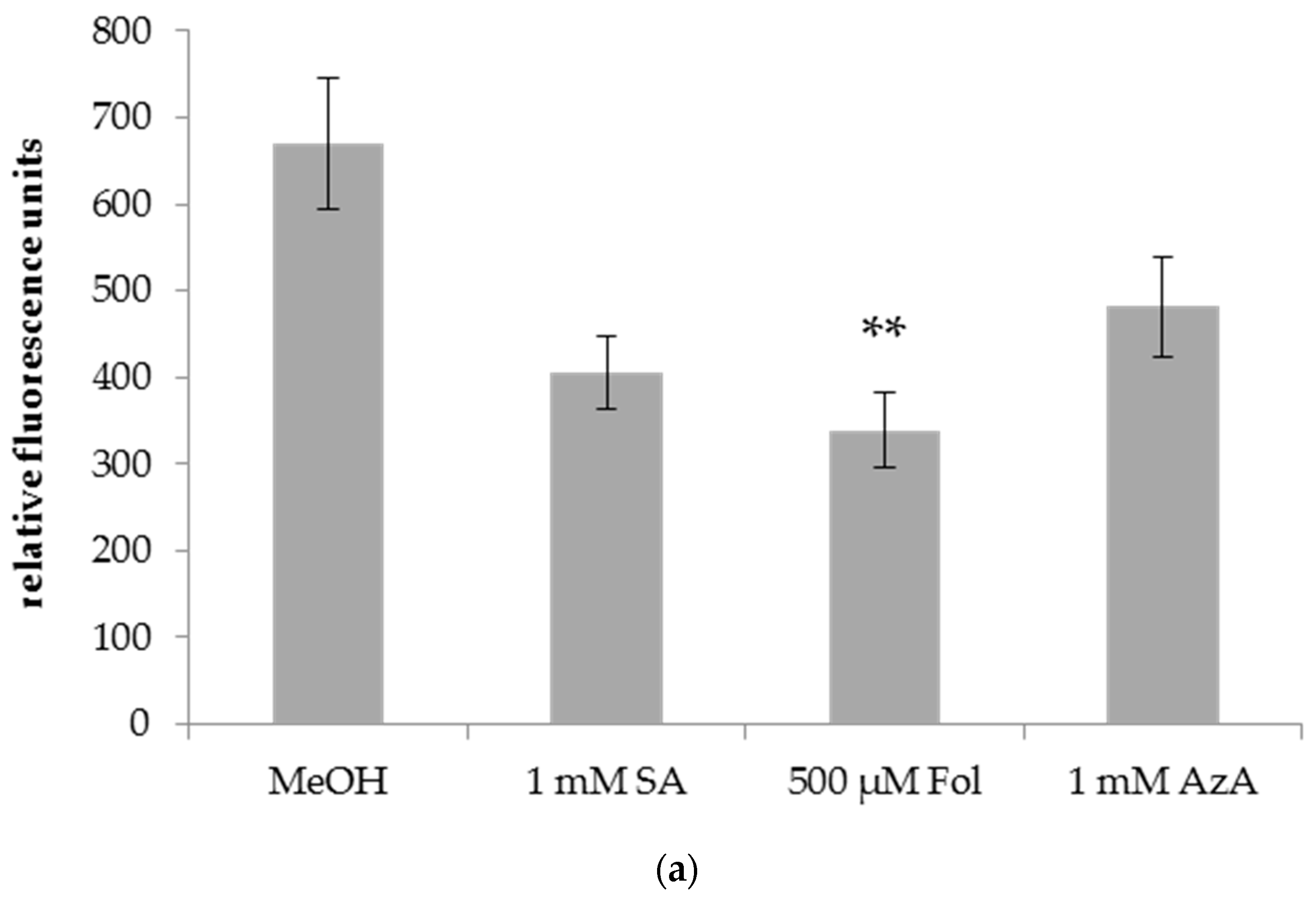
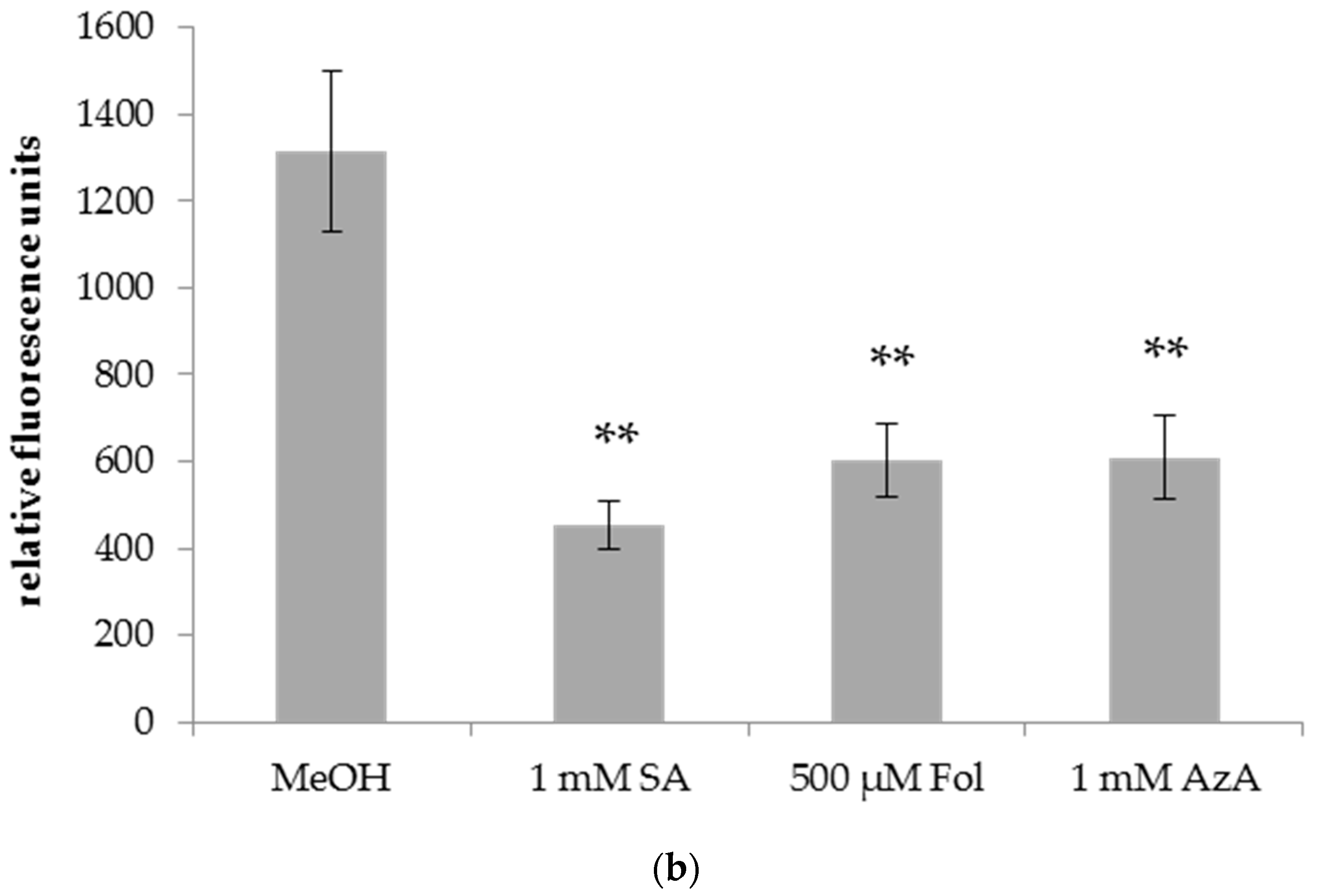
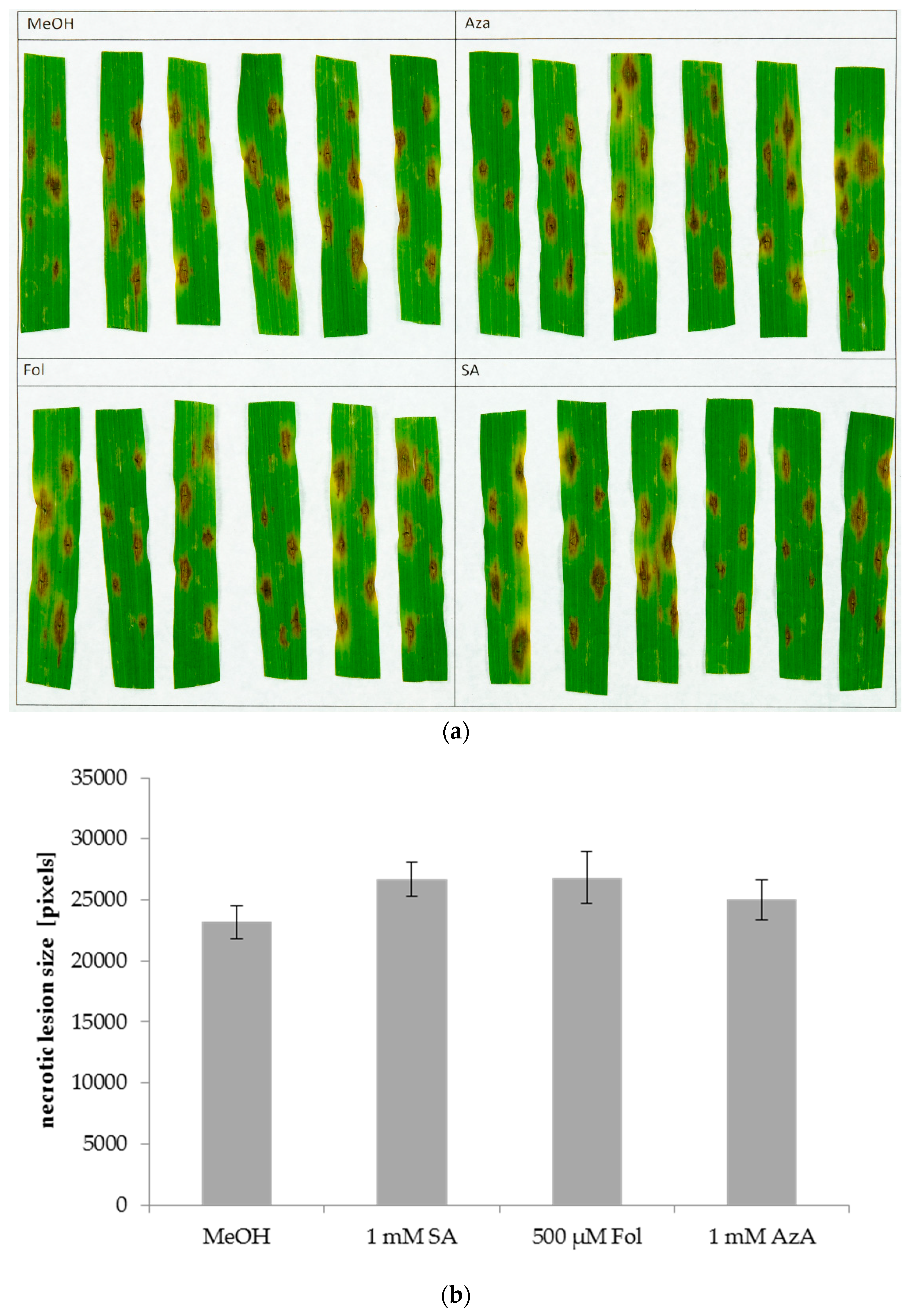
Figure 5.
Lesions caused by Pyrenophora teres on the second leaves of barley after application of SA, Fol, or AzA on leaf 2. Barley cultivar GP plants were infiltrated in the second true leaf with 0.025% MeOH as control, 1 mM SA, 500 µM Fol, or 1 mM AzA in 10 mM MgCl2 as indicated. One day later, the same leaves were inoculated with P. teres by pipetting droplets of a solution containing P. teres spores onto the leaf surface. The resulting necrotic lesions were photographed at 4 dpi (a) and measured using ImageJ (b). Bars in (b) represent the average of 35 replicates from 6 (Fol) or 42 replicates from 7 (all other treatments) independent experiments (each experiment with 6 replicates per treatment, except one experiment with Fol comprising 5 replicates) ± standard error.
Figure 5.
Lesions caused by Pyrenophora teres on the second leaves of barley after application of SA, Fol, or AzA on leaf 2. Barley cultivar GP plants were infiltrated in the second true leaf with 0.025% MeOH as control, 1 mM SA, 500 µM Fol, or 1 mM AzA in 10 mM MgCl2 as indicated. One day later, the same leaves were inoculated with P. teres by pipetting droplets of a solution containing P. teres spores onto the leaf surface. The resulting necrotic lesions were photographed at 4 dpi (a) and measured using ImageJ (b). Bars in (b) represent the average of 35 replicates from 6 (Fol) or 42 replicates from 7 (all other treatments) independent experiments (each experiment with 6 replicates per treatment, except one experiment with Fol comprising 5 replicates) ± standard error.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lenk, M.; Wenig, M.; Mengel, F.; Häußler, F.; Vlot, A.C. Arabidopsis thaliana Immunity-Related Compounds Modulate Disease Susceptibility in Barley. Agronomy 2018, 8, 142. https://doi.org/10.3390/agronomy8080142
AMA Style
Lenk M, Wenig M, Mengel F, Häußler F, Vlot AC. Arabidopsis thaliana Immunity-Related Compounds Modulate Disease Susceptibility in Barley. Agronomy. 2018; 8(8):142. https://doi.org/10.3390/agronomy8080142
Chicago/Turabian StyleLenk, Miriam, Marion Wenig, Felicitas Mengel, Finni Häußler, and A. Corina Vlot. 2018. "Arabidopsis thaliana Immunity-Related Compounds Modulate Disease Susceptibility in Barley" Agronomy 8, no. 8: 142. https://doi.org/10.3390/agronomy8080142
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.