
Specific Depletion of Leukemic Stem Cells: Can MicroRNAs Make the Difference?
by
Tania Martiáñez Canales
†, 
David C. De Leeuw
†,
Eline Vermue
,
Gert J. Ossenkoppele
and
Linda Smit
* Department of Hematology, VU University Medical Center, Cancer Center Amsterdam, Boelelaan 1117, 1081 HV Amsterdam, The Netherlands
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cancers 2017, 9(7), 74; https://doi.org/10.3390/cancers9070074
Submission received: 2 May 2017
/
Revised: 15 June 2017
/
Accepted: 20 June 2017
/
Published: 30 June 2017
(This article belongs to the Special Issue The Roles of microRNA in Tumor Initiation and Development: Diagnostic and Therapeutic Potential)
Abstract
:For over 40 years the standard treatment for acute myeloid leukemia (AML) patients has been a combination of chemotherapy consisting of cytarabine and an anthracycline such as daunorubicin. This standard treatment results in complete remission (CR) in the majority of AML patients. However, despite these high CR rates, only 30–40% (<60 years) and 10–20% (>60 years) of patients survive five years after diagnosis. The main cause of this treatment failure is insufficient eradication of a subpopulation of chemotherapy resistant leukemic cells with stem cell-like properties, often referred to as “leukemic stem cells” (LSCs). LSCs co-exist in the bone marrow of the AML patient with residual healthy hematopoietic stem cells (HSCs), which are needed to reconstitute the blood after therapy. To prevent relapse, development of additional therapies targeting LSCs, while sparing HSCs, is essential. As LSCs are rare, heterogeneous and dynamic, these cells are extremely difficult to target by single gene therapies. Modulation of miRNAs and consequently the regulation of hundreds of their targets may be the key to successful elimination of resistant LSCs, either by inducing apoptosis or by sensitizing them for chemotherapy. To address the need for specific targeting of LSCs, miRNA expression patterns in highly enriched HSCs, LSCs, and leukemic progenitors, all derived from the same patients’ bone marrow, were determined and differentially expressed miRNAs between LSCs and HSCs and between LSCs and leukemic progenitors were identified. Several of these miRNAs are specifically expressed in LSCs and/or HSCs and associated with AML prognosis and treatment outcome. In this review, we will focus on the expression and function of miRNAs expressed in normal and leukemic stem cells that are residing within the AML bone marrow. Moreover, we will review their possible prospective as specific targets for anti-LSC therapy.
1. Introduction
The treatment outcome of acute myeloid leukemia (AML) patients depends on several factors, including karyotype and molecular alterations present in the leukemic cell bulk. Combination chemotherapy leads to complete remission (CR) in the majority of patients [1]. However, 50% of patients that have been in CR develop a relapse within 5 years after their initial diagnosis. This recurrence of the disease is thought to be caused by chemotherapy resistant leukemic cells with stem cell-like properties, named “leukemic stem cells” (LSCs) [2,3,4]. To improve the treatment outcome of AML patients it will be crucial to eradicate LSCs to finally prevent relapse. LSCs are functionally defined by their ability to initiate AML in immunodeficient mice [5], and were initially identified as a population of leukemic cells with a CD34+CD38− immunophenotype, similar to normal hematopoietic stem cells (HSCs) [4,5]. However, LSCs showed to be more heterogeneous than the CD34+CD38− phenotype and to reside also in other cell compartments [6,7,8,9]. Moreover, at AML relapse, LSC frequency and phenotypic diversity showed to much greater than at diagnosis, indicating that chemotherapy promotes changes in the LSC compartment [10]. In contrast to what is observed in the patient at relapse, cytarabine resistant cells generated in an AML xenograft mouse model are not enriched for the CD34+CD38− phenotype or for cells containing enhanced functional leukemia-initiating potential, neither were these cells enriched for stem cell genes [8]. The clinical importance of LSCs was shown by a study of Ng et al., in where it was demonstrated that the presence of a 17 gene LSC expression signature derived from functionally defined LSCs could predict the risk for relapse [11]. Altogether, to improve treatment outcome for AML patients it will be crucial to eradicate the dynamic LSC compartment during the disease course.
LSCs co-exist with residual normal CD34+CD38− HSCs in the bone marrow of the AML patient. Increasing the chemotherapy dose might eliminate LSCs, nevertheless will inevitably result in the non-specific elimination of HSCs, leading to prolonged or permanent marrow aplasia and other toxicities. Therefore, it will be crucial to develop additional therapies that specifically eradicate LSCs but that will spare HSCs (Figure 1). Several cell properties enabling discrimination of LSCs from HSCs within AML bone marrows were identified, including expression of CLEC12A (CLL-1), CD123, TIM-3, CD34 and CD45, scatter properties and activity of aldehyde dehydrogenases [12,13,14,15,16].
Previously, gene expression profiling (GEP) has been performed to uncover transcriptional programs present in normal bone marrow and AML cells and to discriminate between AML subtypes and responding and non-responding patients. Moreover, the presence of a gene expression profile GEP can also predict patient survival [5,6,7,8]. However, GEP has not often succeeded in the uncovering of genes that upon targeting specifically eradicate leukemia cells, leaving the HSCs untouched. The reason for this might be that a complex phenomenon like therapy resistance and cancer cell maintenance is not easily overruled by targeting a single gene. To unravel chemotherapy resistance mechanisms present in LSCs, several studies compared the gene expression signature of CD34+CD38− LSCs or functionally defined LSCs with that of non-LSC compartments, AML blasts or leukemic progenitors [11,17,18,19]. To identify whether potential anti-LSC targets might be specific and sparing healthy HSCs, GEPs of LSCs were compared with those of HSCs [20,21,22]. All these past studies compared the gene expression of AML LSCs with that of HSCs derived from healthy donors. However, as AML induce changes in HSCs and in the healthy bone marrow microenvironment, thereby suppressing HSC function and supporting LSC survival, self-renewal and chemotherapy resistance [23,24,25,26] these studies did not take into account the changes induced in HSCs by the leukemia itself. As LSCs utilize a variety of mechanisms to resist chemotherapy and to drive relapse, the major challenge in targeting all the leukemic cells that contain stem cell properties is their heterogeneity. MiRNA-based therapeutic strategies might successfully eliminate a large population of AML cells with stem cell features as they target hundreds of genes at the same time.
2. MicroRNAs
MicroRNAs (MiRNAs) are a class of small, non-coding RNAs of 18–25 nucleotides that post-transcriptionally control the translation and stability of mRNAs [27]. A miRNA is synthesized as a long RNA transcript known as pri-miRNA, which is cleaved by the RNAse III endoribonuclease Drosha to a pre-miRNA. This pre-miRNA is further processed in the cytoplasm by the protein Dicer to a mature functional miRNA [27]. The mature miRNA is silencing its target genes via mRNA degradation or via prevention of translation of the mRNA (Figure 2). By targeting tens to hundreds of genes at the same time, miRNAs can control basic biological functions and pathways such as HSC differentiation. Distinct miRNAs fine-tune each step of haematopoiesis, including the number and repopulation potential of HSCs [28,29,30]. Dicer showed to be essential for the persistence of HSCs and specifically miR-125a is able to control the number of HSCs by regulating hematopoietic stem/progenitor cell apoptosis [31] and long-term repopulating stem cell potential of mouse and human progenitors [30]. Deregulation of miRNA expression and function in hematopoietic cells can result in the development of cancer and cancer progression [32,33].
AML is characterized by founder mutations in an HSC or a more differentiated progenitor, disrupting the differentiation pathway originating from these cell populations and leading to abnormal miRNA expression patterns. A aberrant miRNA signatures strongly correlate with tumour classification, cytogenetics, molecular abnormalities, prognosis and therapy response [34,35,36]. Strikingly, miRNA expression profiles can provide prognostic information that is complementing cytogenetics, mutation analysis, and gene expression data [34] and can even be more potent in disease classification and prediction of therapy response and outcome than a GEP. For example, a miRNA expression profile could better classify acute leukemia of ambiguous lineage as either lymphoid or myeloid [37] than a GEP [38]. The outperformance of miRNA GEPs as compared to mRNA GEPs might be due to the enhanced stability of miRNAs as compared to mRNAs. Recently, it was also shown that specific serum exosome miRNAs can function as biomarkers for AML, circumventing the use of invasive bone-marrow aspirations and dependence on available leukemic blasts [39].
The aberrant expression of miRNAs contributes to the character of the tumor and can be either oncogenic or tumor suppressive depending on the cellular context and available expressed miRNA targets. In view of heterogeneity of LSCs and AML as being a heterogeneous disease, not successfully treated by targeting a single gene, miRNA modulation may hold the key to successful elimination of therapy resistant leukemic (stem) cells either by inducing apoptosis or by sensitizing for chemotherapy. Moreover, since there is differential expression of miRNAs in AML and normal bone marrow as well as in LSCs and HSCs and LSCs and leukemic progenitors, agents that modulate activity might potentially lead to leukemia and/or LSC-specific effects.
By using aldehyde dehydrogenases (ALDH) activity miRNA expression profiles of LSCs, leukemic progenitors and HSCs all obtained from the same AML sample were obtained [16,40]. This approach resulted in identification of miRNAs differentially expressed between LSC and HSC populations and between LSCs and leukemic progenitors [40]. Several of those miRNAs showed to be specifically expressed in LSCs and/or HSCs, and to have prognostic value in AML and/or to function as targets for miRNA-based anti-LSC therapy [40,41].
3. MicroRNAs in Healthy HSCs
HSCs can undergo self-renewal and give rise to all the cells of the hematopoietic system during life. To do so, there is a balance between self-renewal and differentiation that is strictly controlled by several molecular factors, including the activity of miRNAs. Over the last years, the expression and function of miRNAs during hematopoiesis have been intensively studied however studies about the expression of miRNAs in highly enriched human stem and progenitor cell populations are scarce. This is partly due to a lack of consensus on which markers to use for isolation of pure HSCs but also due to the difficulty in isolating sufficient numbers of HSCs for profiling. In general, human HSCs reside within the immuno-phenotypical defined compartment of lineage negative (Lin−) CD34+CD38−CD90+CD49f+CD45RA− cells, which differentiate into Lin−CD34+CD38−CD90−CD49f−CD45RA−(Thy−1neg-lo) [42] multipotent progenitors (MPP) containing both lymphoid and myeloid potential [43,44] The more committed Lin−CD34+CD38+ common myeloid progenitors (CMP), granulocyte-macrophage progenitors (GMP) and megakaryocyte-erythroid progenitors (MEP) that develop from the MPP, can be separated using differential expression of CD123, CD110 and CD45RA [45,46]. HSCs and immature progenitors are also characterized by expression of CD133 [47,48].
The expression of miRNAs has been mostly studied in fractions containing murine HSCs [49,50], in human CD34+ and CD133+ cell fractions [51,52,53,54] and in human CD34+CD38− [49] and CD90+CD45RA− fractions [55,56]. The group of Georgantas performed the first large scale miRNA profiling of human CD34+ peripheral blood and bone marrow cells and identified 33 miRNA [51]. Since then, other groups have performed similar analysis in more HSC-enriched fractions. Both miR-29a and miR-125a/b consistently showed higher expression in HSCs (Lin−CD34+CD38− CD90+CD45RA−) and multipotent progenitors (Lin−CD34+CD38−CD90−CD45RA−) than in committed and differentiated progenitors [55,56]. Depletion of miR-29a resulted in decreased numbers of HSCs and progenitors, decreased HSC self-renewal, increased HSC cell cycling and apoptosis, which is partly due to the enhanced expression of DNMT3A [57]. Ectopic expression of miR-29a in mouse HSC/progenitors resulted in acquisition of self-renewal capacity, a bias to myeloid differentiation, and induction of a myeloproliferative disorder that can develop into AML [55]. Comparing the expression of miRNAs in Lin−Sca−1+c−Kit+ (LSK), Lin−Sca−1−c−Kit+ (LS−K+), erythroid and myeloid mouse cells identified 131 miRNAs differentially expressed between these cell types. MiR-99b, let-7e and miR-125a showed to be highly expressed in LSKs and down-regulated upon differentiation. Overexpression of miR-125a increases the number of cobblestone-area forming cells and overexpression of miR-99b/let-7e/miR125a or miR-125a alone keeps the mouse HSCs in a primitive state [58] (Table 1). Ectopic expression of miR-125a in murine and human multipotent progenitors resulted also in increased self-renewal and robust long-term multi-lineage repopulation in transplanted recipient mice [30]. Besides enhancing HSC self-renewal potential, the size of the HSC population is modulated by miR-125a by regulating apoptosis [31] (Table 1).
An expression profiling and functional study by O’Connell et al identified 11 miRNAs enriched in HSCs. Ectopic expression of these miRNAs in normal bone marrow identified miR-125b as the miR that induces the greatest increase in repopulation potential [50]. Moreover, Ooi et al showed that miR-125b overexpression led to a reduction in apoptosis in HSCs [56]. Thus, miR-125b promotes self-renewal and inhibits apoptosis in HSCs [50,56,58].
MiR-126 and miR-130a are expressed in HSCs and early progenitors from both mice and human, but not in differentiated progenitors [59]. Downregulation of miR-126 in HSCs results in enhanced hematopoietic stem/progenitor cell proliferation without inducing exhaustion, resulting in expansion of mouse and human long-term repopulating HSCs. Decreased miR-126 increases also cell cycle progression and the number of HSCs (CD34+CD38-CD90+CD45RA−) [60].
Among the numerous miRNA expression studies in HSCs and progenitors there is considerable variation in results. This is partly due to the use of different profiling methods but also due to profiling of different immune-phenotypically defined HSCs that were derived from different sources; e.g., whole bone marrow, total CD34+ population or progenitor populations. Several studies investigating the functional role of specific miRNAs in normal hematopoiesis have been published and many of the identified miRNAs showed to affect progenitor lineage commitment and functions of mature hematopoietic cells (Table 1). Leukemia-inducing mutations cause aberrant miRNA expression in HSCs and or progenitors, resulting in impaired differentiation, apoptosis and/or self-renewal [128].
4. Differential Expression of MicroRNAs between LSCs and HSCs Residing within the AML Bone Marrow
The CD34+CD38− cell compartment residing within the AML bone marrow includes both leukemic and normal stem cells [4]. Both stem cell compartments have many features in common and the extent to which they differ is important for development of therapies targeting relapse-initiating cells while sparing HSCs. The properties of normal HSCs are influenced by the leukemic microenvironment but also by the AML cells themselves [24,25,26]. It has even been shown that AML cells can suppress hematopoiesis by the miRNAs in exosomes released from the AML cells [24]. Since LSCs and HSCs are influenced by both the leukemic microenvironment and the leukemic cells, searching for differences in miRNA expression between HSCs and LSCs that are both obtained from the AML patient’s bone marrow will enhance the chance of finding genuine anti-LSC targets.
For the purification of HSCs and LSCs from an AML bone marrow the unequivocal separation of both compartments is necessary. LSCs often have aberrant protein expression, i.e., cell surface markers that do not fit to their lineage or maturation state [12,14,15]. Frequently observed non-myeloid lineage markers that are often used to distinguish between leukemic and normal myeloid cells are for example CD7, CD19, CD11b and CD56 [129]. These lineage markers are generally absent on normal HSCs while expressed in a subset of AML cases on leukemic stem and progenitor cells [14]. Other markers that are specifically expressed on LSCs and lacking on HSCs are for example CLL-1 and CD123 [15,130]. Generally, the expression of an aberrant immune-phenotypic marker is not absolute; i.e., not expressed on all leukemic cells within one patient, but also not present in all AML patients, which makes it difficult to use one particular biomarker for the isolation of LSCs and HSCs from all the AML patients [reviewed in [131]. Since HSCs have high ALDH activity and CD34+CD38− LSCs are in general characterized by lower ALDH activity, this problem can be circumvented by using ALDH activity as a functional biomarker. Using the difference in ALDH activity in combination with detection of aberrant leukemia-associated marker expression, CD34+CD38− HSCs and CD34+CD38− LSCs from AML bone marrows have been purified. After determining the expression of miRNAs in these cell fractions the comparison of the miRNA profiles of LSCs with those of residual HSCs revealed that MiR-551b, miR-10a, miR-151-5p, miR-29b and miR-125b are higher expressed in HSCs than in LSCs while miR-181b, miR-221, miR-21 and miR-22 are higher expressed in LSCs than in HSCs (Figure 3) [40].
Mir551b is the top differentially expressed miRNA between residual HSCs and LSCs, showing high expression in HSCs [40]. MiR-551b is not only highly expressed in residual HSCs in AML but also in HSCs residing in healthy bone marrow [41], suggesting a link between “stemness” and presence of miR-551b. Not only genuine normal stem cells have high mir-551b expression but also AML cases with an undifferentiated stem cell-like phenotype have [41]. Importantly, AML cases with enhanced miR-551b expression are associated with a poorer clinical outcome than those with lower miR-551b expression [41], potentially reflecting the influence of “stemness” on therapy sensitivity. Indeed, in ovarian cancer the expression of miR-551b is enhanced in the side population, the cell population that is enriched for cancer stem cells [132]. Mir-551b is located at the chromosome 3q26 locus, which is translocated and leading to overexpression of EVI-1 in a subset of AML patients. In ovarian cancer, amplification of 3q26 leads to increased expression of miR-551b, subsequently contributing to apoptosis resistance and increased survival and proliferation of the cancer cells. The mechanisms whereby miR-551b increases proliferation in ovarian cancer cells is not by decreasing the levels of mRNA targets but by binding to the STAT3 promoter and by recruitment of RNA polymerase II and TWIST1 [133]. In ovarian cancer and lung cancer cell lines, enhanced miR551b expression showed to be linked with therapy resistance [132,134]. Other miRNAs highly expressed in residual HSCs and potentially associated with HSC functions are miR-29b and miR-125b [40]. In general, both miRNAs are downregulated in AML patients as compared to HSCs [50,56]. However, it might be possible that miR-29b and miR-125b are higher expressed in a small subset of AML cases or in a small subpopulation of leukemic cells within the AML bulk. The research group of Marcucci supported a tumor suppressor role for miR-29b and used synthetic anti-miR-29b oligonucleotides as a novel strategy to eliminate AML cells [135,136]. MiR-29b overexpression had similar effects as the hypomethylating agents 5-azacytidine and decitabine [56] and its downregulation has been linked to promotion of DNA hypermethylation in AML cells by directly targeting DNMT3A, DNMT3B and SP1 [137,138]. MiR-29b upregulation has also demonstrated to inhibit cell proliferation, promote myeloid differentiation and induce apoptosis [135]. Together these data suggested that the consequences of overexpression of miR-29b is cell-type specific and may depend on the differentiation and/or transformation state of the cancer cell. The effect of enhanced miR-29b on the quiescence state of LSCs and thereby their chemotherapy sensitivity and survival has not been extensively investigated yet.
In AML, miR-125b is strongly upregulated as compared to whole healthy bone marrow, particularly in patients with a t(2;11)(p21;q23) [139]. Enhanced expression of miR-125b in myelodysplastic syndrome (MDS) and in AML with a (2;11)(p21;q23) resulted in a differentiation arrest, indicating a connection between high miR-125b and an immature leukemic phenotype [139,140]. Moreover, miR-125b overexpression causes a myeloproliferative disorder that progressed to an aggressive form of AML within 3–4 months [50,58,65]. Mice transplanted with hematopoietic progenitors overexpressing miR-125b led to various types of leukemia, including B-cell acute lymphoblastic leukemia, T-cell acute lymphoblastic leukemia or a myeloproliferative neoplasm depending on the degree of miR-125b expression [65].
MiR-181b is one of the upregulated miRNAs in LSCs as compared to HSCs [40]. MiR-181b is part of a miRNA signature expressed in cytogenetically normal (CN) AML containing high-risk molecular characteristics (e.g., NPM1 negative, FLT3-ITD positive) and is associated with a good prognosis [141]. Moreover, a 15 miRNA signature, including miR-181b, showed an association with the presence of the CEBPα mutation, possibly partly explaining the good prognostic characteristics of AML with high expression of the miR-181 family [142]. The overexpression of miR-181b promotes apoptosis and inhibits the viability of MLL-rearranged AML cells [143].
Lastly, miR-21 and miR-221, are higher expressed in LSCs and AML blasts than in HSCs and healthy bone marrow cells [34,40,144]. MiR-221 showed to be a biomarker distinguishing AML from acute lymphoid leukemia (ALL) [34,37]. An association between “stemness”, DNMT3A expression and miR-221 has been shown in breast cancer [145]. MiR-21 and miR-221 are both higher expressed in the pancreatic cancer cells residing in the side population and modulation of miR-21 and miR-221 in initiating stem-like cells affects tumorigenesis, metastasis, and chemotherapy resistance in pancreatic cancer [146]. MiR-21 showed to be overexpressed in AML with a NPM1 mutation [147]. Inhibition of miR-21 in the myeloid cell lines HL60 and K562 reduced cell growth, induced apoptosis and a G1 cell cycle arrest [148]. Moreover, several studies showed that downregulation of miR-21 in myeloid leukemia cell lines increased the sensitivity to various chemotherapeutic agents [149,150,151], making targeting of miR-21 a potential successful selective approach to sensitize LSCs for chemotherapy.
5. MicroRNAs Differentially Expressed between LSCs and Leukemic Progenitors
MiRNAs that are functionally involved in chemotherapy resistance and/or leukemia-initiating potential might be differentially expressed between LSCs and the chemotherapy sensitive AML bulk. By comparing miRNA expression in purified LSCs (CD34+CD38−) and leukemic progenitors (CD34+CD38+) 12 differentially expressed miRNAs were identified (Figure 3) [40]. The top three lower expressed miRNAs in LSCs as compared to leukemic progenitors are miR-1274a, miR-886 and miR-1305. Although there is not much information on the function of miR-1274a, it is suggested to be derived from tRNA processing rather than being a miRNA [152]. In gastric cancer cells, miR-1274a has been described as an oncogene involved in cell proliferation and migration by targeting FOXO4 [153].
Also miR-886 (vtRNA2-1, pre-miR-886, or CBL3) showed not to be a miRNA but a newly identified non-coding RNA (ncRNA) [154] that acts as a tumor suppressor targeting Protein Kinase R (PKR) [155,156]. PKR is a sensor that recognizes viruses and induces apoptosis to eliminate infected cells. Therefore, the nc886 signaling pathway in cancer cells is suggested to function in sensing and eliminating pre-malignant cells, analogous to PKR’s role in cellular innate immunity [154]. The nc886 is transcribed by RNA polymerase III (Pol III) [157] and is the first case of a Pol III gene whose expression is silenced by CpG DNA hypermethylation in several types of cancer. Low expression of nc886 has been associated with poor prognosis in AML, low risk MDS and small cell lung cancer [158,159,160], suggesting that its lower expression in LSCs might also be involved in a decreased response to chemotherapy in these cells. Thus, increasing nc886 expression might be a strategy to enhance chemotherapy sensitivity.
MiR-1305 showed to have a change in expression during the cell cycle in embryonic stem cells. Downregulation of miR-1305 facilitates the maintenance of pluripotency and increased cell survival, while its overexpression induced differentiation of pluripotent stem cells, increased cell apoptosis and sped up G1/S transition [161].
MiRNAs that are higher expressed in LSCs than in leukemic progenitors are miR-126-5p, miR-126-3p, miR-22, miR-335 and mir-150. MiR-126 is the top miRNA differentially expressed between CD34+CD38- LSCs and CD34+CD38+ leukemic progenitors. MiR-126 is high expressed in LSCs and even more enhanced in HSCs [40] (Figure 3). Moreover, miR-126 is part of an LSC-associated miRNA signature that was derived from functionally validated AML LSCs and its expression has been linked to LSC activity [60]. Patients with high miR-126 levels co-express genes that are present in stem cell gene signatures [19,40], implicating that miR-126 influences stem cell properties and maintenance of “stemness” in both normal and leukemic stem cells. In AML, high miR-126 expression is associated with poor survival and a high chance of relapse [40,60,162], reflecting the association of enhanced miR-126 activity with a decrease in chemotherapy sensitivity [60]. Both the overexpression and knockout of miR-126 result in enhanced leukemogenesis in cooperation with the t(8;21) fusion gene. MiR-126 overexpression has a stronger effect on long-term survival and progression of AML1-ETO9a-mediated LSCs in mice than does miR-126 knock-out [163]. Knockdown of miR-126 led to differentiation, apoptosis and reduction of AML growth [40,60] but also enhances responsiveness to chemotherapy [163]. Most importantly, targeting of miR-126 specifically reduced the clonogenic capacity of LSCs and leukemic progenitors without affecting normal HSCs [40,60] (Figure 4). Moreover, miR-126 targeting using nanoparticles resulted in the depletion of LSCs in an AML xenotransplantation model [162]. This specific effect of miR-126 is due to the opposite function of miR-126 in LSCs and in HSCs [40,60]. In LSCs, decreased miR-126 attenuates LSC quiescence and overrules chemotherapy resistance [60]. In HSCs, knockdown of miR-126 results in enhanced hematopoietic stem/progenitor cell proliferation, increases cell cycle progression and increases the number of stem cells (CD34+CD38-CD90+CD45RA-) [72] (Figure 4). MiR-126 targets multiple genes affecting the PI3K/AKT/mTOR pathway [60,72] and its discriminative function in LSCs and HSCs might reflect the opposite function of the PI3K/PTEN signaling pathway in LSCs and HSCs [164]. The distinct function of miR-126 in HSCs and LSCs makes it an ideal therapeutic target for depletion of LSCs without harming HSCs and potentially even enhancing hematopoietic recovery.
MiR-22 is upregulated in myelodysplastic syndrome (MDS) and leukemia and its aberrant expression correlates with poor survival [85]. MiR-22 was identified as being higher in LSCs than in leukemic progenitors but decreased in residual HSCs as compared to LSCs [40]. In contrast to these results, Jiang et al., showed that AML cells have lower miR-22 than CD34+ normal healthy bone marrow cells [104]. Mice that conditionally express miR-22 in the hematopoietic cell compartment showed decreased levels of 5-hydroxymethylcytosine (5-hmC), enhanced HSC self-renewal and developed MDS and hematological malignancies [85], indicating the oncogenic function of miR-22. MiR-22 targets TET2, a member of the TET methylcytosine dioxygenase family, and ectopic expression of TET2 suppressed the miR-22-induced phenotypes [85]. In contrast to this study, Jiang et al. [104] showed that miR-22 also has a tumor suppressive function. Forced expression of miR-22 inhibited leukemia development and maintenance in a xenograft AML mouse model [104]. Enhanced miR-22 expression, regulated by PU.1, resulted in monocyte/macrophage differentiation. In AML, there is downregulation of PU.1 and Mir-22 as well as upregulation of EVI-1. Reintroduction of miR-22, and the associated downregulation of its target EVI-1, relieved the differentiation block and inhibited the growth of AML bone marrow blasts [105].
6. Therapeutic Approaches to Specifically Eliminate LSCs: Sensitization to Chemotherapy
As miRNAs can regulate many genes and are also controlled by more than one gene, miRNAs are very appealing targets for specific anti-LSC therapies. Depending on the expression and function of miRNAs in AML, there are two approaches for developing miRNA-based therapies: antagonists and mimics. Antagonists can inhibit miRNAs and are usually single-stranded oligonucleotides [165]. Efficient silencing of miRNA activity in vivo requires the chemical modification of these oligonucleotides to improve their binding affinity, bio stability and pharmacokinetic properties. The most common modifications to increase the duplex melting temperature and to improve their resistance to nucleases include 2′-O-methyl- (2′-O-Me-), 2′-methoxyethyl- (2′-MOE-), 2′-fluoro- and the bicyclic locked nucleic acid (LNA) modifications [166,167,168]. Among these modifications, LNA exhibits the highest affinity toward complementary RNA [169]. Moreover, increased nuclease resistance is also achieved by substituting the phosphodiester (PO) backbone linkages with phosphorothioate (PS) which, apart from nuclease resistance, enhances the binding to plasma proteins leading to reduced clearance by glomerular filtration and urinary excretion. Moreover, inhibition of miRNAs by ultra-short 8-mer LNAs, which enable antagonism of a complete miRNA family, can result in therapeutic benefit in mouse disease models [170,171]. Currently, the most advanced therapeutic miRNA antagonists are directed against miR-122. One anti-miR-122 LNA has already been successfully tested in Phase II clinical trials for patients with hepatitis C [172]. Moreover a phase I clinical trial using a synthetic microRNA antagonist of microRNA-155 (MRG-106) is conducted in patients suffering from cutaneous T-cell lymphoma (CTCL).
Mimics, double stranded oligonucleotides, are used to restore miRNA function and have been designed to restore the function of various tumor-suppressive miRNAs [173]. For example miR-29b is known to be downregulated in AML [135] and a nanoparticle-based delivery system of miR-29b mimics in AML blasts showed decreased AML cell growth and impaired colony formation in a mice model of AML [136]. The use of lenti-, adeno- or adeno-associated viruses to drive the expression of a miRNA has been successfully applied to reduce tumor growth in mouse models [174,175]. The clinical application of a miRNA mimic of miR-34 (MRX34) is at the moment tested in a phase-I trial including patients with hematological malignancies [176].
The specific expression of miRNAs during haematopoiesis can also be used to specifically express a gene in differentiated hematopoietic cells. In a lysosomal storage disorder, there is a need for specific expression of the enzyme galactocerebrosidase in differentiated hematopoietic cells since its expression in HSCs results in toxicity. The enhanced expression of miR-126 in HSCs can inhibit the expression of galactocerebrosidase from a lentiviral construct containing miR-126 binding sites in HSCs, while miR-126 expression decreases during myeloid differentiation thereby causing expression of galactocerebrosidase and restoring lysosomal storage [59].
MiRNA-based therapy might not only function as a single agent but also holds great potential in complementing currently used chemotherapeutics. Since a single miRNA can induce global changes in overall gene expression, modulation of miRNA expression might be very effective in targeting a multi-factorial phenomenon like drug resistance. MiRNA modulation has been shown to have the capacity to enhance the response and suppress the resistance to cytotoxic therapies [177,178].
7. The Delivery of MiRNA Modulators to AML LSCs within the Leukemic Bone Marrow
Successful delivery of therapeutic miRNA(s) to the leukemic cells in the AML bone marrow, without inducing toxicity, is the final challenge. The charged miRNAs have a small size and low molecular weight making it possible to formulate them into effective delivery systems which reduce their clearance and degradation in the blood [179]. Examples of delivery systems for mimics and anti-miRs are lipids, polyethylenimine, dendrimers, poly (lactide-co-glycolide) particles but also naturally occurring polymers, such as chitosan, protamine and atelocollagen [136,179,180,181]. Importantly, the first liposome-formulated mimic is currently being tested in a Phase I clinical trial in patients with unresectable primary liver cancer. Beside the delivery of mimics and anti-miRs by formulation, viral constructs can be used [174,175].
Marcucci et al. developed a transferrin-conjugated nanoparticle delivery system conjugating transferrin (Tf) to PEGylated lipopolyplex nanoparticles (Tf-LPs) which incorporates protamine as a DNA condensing agent, pH-sensitive fusogenic lipids to improve cytoplasmic delivery, and Tf as the targeting ligand specific for cellular delivery (commonly overexpressed on cancer cells and also in AML) [182]. In a study that tested the delivery of miR-29b loaded transferrin-conjugated nanoparticles (Tf-NP-miR-29b) to leukemic cells in a xenograft mouse model, high uptake and strong downregulation of miR-29b targets in the leukemic cells was observed [136]. Tf-NP-miR-29b suppressed AML growth, impaired colony formation, and reduced cell viability in AML patient samples. In addition, Tf-NP-miR-29b also reduced spleen weight and increased overall survival in NSG mice transplanted with AML cell lines [136]. Next to miR-29b, transferrin conjugated nanoparticles containing miR-126 have recently been used. As previously mentioned, treatment with Tf-NP-miR-126 specifically targets the LSC leading to diminished engraftment of both human and mouse AML in secondary recipient transplantations [162]. Before these works, in vivo targeting of miR-196b was also reported to successfully eradicate LSCs from AML blasts harboring MLL translocations [183]. A mimic Tf-NP-miR181a treatment downregulated KRAS, NRAS and MAPK1 and decreased AML growth in mice, resulting in a longer survival compared to the controls [184]. The use of nanoparticles to force expression of miR-22, which is often downregulated in AML, significantly inhibited AML progression in vivo [104]. Altogether, these studies show the great potential for future miRNA based treatment and the use of nanoparticles to deliver them to AML cells.
8. Concluding Remarks
Two decades ago miRNA research started with the expression profiling of various hematopoietic cell populations and types of leukemia which provided us with an enormous number of miRNAs that could potentially play a regulatory role in normal and malignant hematopoiesis. Indeed, many of these miRNAs now have established involvement in controlling differentiation, apoptosis, proliferation and self-renewal in hematopoiesis and leukemia. The most extensively studied miRNAs include miR-125b, miR-29b and miR-126 which are all involved in stem cell regulation and leukemogenesis. MiRNA-based therapy that modulates these miRNAs to prevent leukemogenesis or treat frank leukemia is now possible and holds great potential. Although recent reports on in vivo miRNA treatment are promising, still many issues in optimizing delivery methods and unknown factors like toxicity, due to off-target effects, should be evaluated and solved. It is therefore wanted that future miRNA research focusses on the efficient in vivo delivery and specific targeting of leukemia (stem cells) to really bring miRNAs from bench to bedside.
Acknowledgments
This work is financed by a grant from the Dutch Cancer Society (#7223).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rowe, J.M.; Kim, H.T.; Cassileth, P.A.; Lazarus, H.M.; Litzow, M.R.; Wiernik, P.H.; Tallman, M.S. Adult patients with acute myeloid leukemia who achieve complete remission after 1 or 2 cycles of induction have a similar prognosis: A report on 1980 patients registered to 6 studies conducted by the Eastern Cooperative Oncology Group. Cancer 2010, 116, 5012–5021. [Google Scholar] [CrossRef] [PubMed]
- Van Rhenen, A.; Feller, N.; Kelder, A.; Westra, A.H.; Rombouts, E.; Zweegman, S.; van der Pol, M.A.; Waisfisz, Q.; Ossenkoppele, G.J.; Schuurhuis, G.J. High stem cell frequency in acute myeloid leukemia at diagnosis predicts high minimal residual disease and poor survival. Clin. Cancer Res. 2005, 11, 6520–6527. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, F.; Yoshida, S.; Saito, Y.; Hijikata, A.; Kitamura, H.; Tanaka, S.; Nakamura, R.; Tanaka, T.; Tomiyama, H.; Saito, N.; et al. Chemotherapy-resistant human AML stem cells home to and engraft within the bone-marrow endosteal region. Nat. Biotechnol. 2007, 25, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, D.; Dick, J.E. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat. Med. 1997, 3, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, T.; Sirard, C.; Vormoor, J.; Murdoch, B.; Hoang, T.; Caceres-Cortes, J.; Minden, M.; Paterson, B.; Caligiuri, M.A.; Dick, J.E. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature 1994, 367, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Taussig, D.C.; Miraki-Moud, F.; Anjos-Afonso, F.; Pearce, D.J.; Allen, K.; Ridler, C.; Lillington, D.; Oakervee, H.; Cavenagh, J.; Agrawal, S.G.; et al. Anti-CD38 antibody-mediated clearance of human repopulating cells masks the heterogeneity of leukemia-initiating cells. Blood 2008, 112, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Taussig, D.C.; Vargaftig, J.; Miraki-Moud, F.; Griessinger, E.; Sharrock, K.; Luke, T.; Lillington, D.; Oakervee, H.; Cavenagh, J.; Agrawal, S.G.; et al. Leukemia-initiating cells from some acute myeloid leukemia patients with mutated nucleophosmin reside in the CD34(-) fraction. Blood 2010, 115, 1976–1984. [Google Scholar] [CrossRef] [PubMed]
- Farge, T.; Saland, E.; de Toni, F.; Aroua, N.; Hosseini, M.; Perry, R.; Bosc, C.; Sugita, M.; Stuani, L.; Fraisse, M.; et al. Chemotherapy Resistant Human Acute Myeloid Leukemia Cells are Not Enriched for Leukemic Stem Cells but Require Oxidative Metabolism. Cancer Discov. 2017. [Google Scholar] [CrossRef] [PubMed]
- Pabst, C.; Bergeron, A.; Lavallee, V.-P.; Yeh, J.; Gendron, P.; Norddahl, G.L.; Krosl, J.; Boivin, I.; Deneault, E.; Simard, J.; et al. GPR56 identifies primary human acute myeloid leukemia cells with high repopulating potential in vivo. Blood 2016, 127, 2018–2127. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-C.; LaMere, M.; Stevens, B.M.; Ashton, J.M.; Myers, J.R.; O’Dwyer, K.M.; Liesveld, J.L.; Mendler, J.H.; Guzman, M.; Morrissette, J.D.; et al. Evolution of acute myelogenous leukemia stem cell properties after treatment and progression. Blood 2016, 128, 1671–1678. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.W.K.; Mitchell, A.; Kennedy, J.A.; Chen, W.C.; McLeod, J.; Ibrahimova, N.; Arruda, A.; Popescu, A.; Gupta, V.; Schimmer, A.D.; et al. A 17-gene stemness score for rapid determination of risk in acute leukaemia. Nature 2016, 540, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Terwijn, M.; Zeijlemaker, W.; Kelder, A.; Rutten, A.P.; Snel, A.N.; Scholten, W.J.; Pabst, T.; Verhoef, G.; Löwenberg, B.; Zweegman, S.; et al. Leukemic stem cell frequency: A strong biomarker for clinical outcome in acute myeloid leukemia. PLoS ONE 2014, 9, e107587. [Google Scholar] [CrossRef] [PubMed]
- Kikushige, Y.; Shima, T.; Takayanagi, S.; Urata, S.; Miyamoto, T.; Iwasaki, H.; Takenaka, K.; Teshima, T.; Tanaka, T.; Inagaki, Y.; et al. TIM-3 Is a Promising Target to Selectively Kill Acute Myeloid Leukemia Stem Cells. Cell Stem Cell 2010, 7, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Van Rhenen, A.; Moshaver, B.; Kelder, A.; Feller, N.; Nieuwint, A.W.M.; Zweegman, S.; Ossenkoppele, G.J.; Schuurhuis, G.J. Aberrant marker expression patterns on the CD34+CD38− stem cell compartment in acute myeloid leukemia allows to distinguish the malignant from the normal stem cell compartment both at diagnosis and in remission. Leukemia 2007, 21, 1700–1707. [Google Scholar] [CrossRef] [PubMed]
- Van Rhenen, A.; van Dongen, G.A.M.S.; Kelder, A.; Rombouts, E.J.; Feller, N.; Moshaver, B.; Stigter-van Walsum, M.; Zweegman, S.; Ossenkoppele, G.J.; Jan Schuurhuis, G. The novel AML stem cell associated antigen CLL-1 aids in discrimination between normal and leukemic stem cells. Blood 2007, 110, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Schuurhuis, G.J.; Meel, M.H.; Wouters, F.; Min, L.A.; Terwijn, M.; de Jonge, N.A.; Kelder, A.; Snel, A.N.; Zweegman, S.; Ossenkoppele, G.J.; et al. Normal hematopoietic stem cells within the AML bone marrow have a distinct and higher ALDH activity level than co-existing leukemic stem cells. PLoS ONE 2013, 8, e78897. [Google Scholar] [CrossRef] [PubMed]
- Gentles, A.J.; Plevritis, S.K.; Majeti, R.; Alizadeh, A.A. Association of a leukemic stem cell gene expression signature with clinical outcomes in acute myeloid leukemia. JAMA 2010, 304, 2706–2715. [Google Scholar] [CrossRef] [PubMed]
- Gal, H.; Amariglio, N.; Trakhtenbrot, L.; Jacob-Hirsh, J.; Margalit, O.; Avigdor, A.; Nagler, A.; Tavor, S.; Ein-Dor, L.; Lapidot, T.; et al. Gene expression profiles of AML derived stem cells; similarity to hematopoietic stem cells. Leukemia 2006, 20, 2147–2154. [Google Scholar] [CrossRef] [PubMed]
- Eppert, K.; Takenaka, K.; Lechman, E.R.; Waldron, L.; Nilsson, B.; van Galen, P.; Metzeler, K.H.; Poeppl, A.; Ling, V.; Beyene, J.; et al. Stem cell gene expression programs influence clinical outcome in human leukemia. Nat. Med. 2011, 17, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Kitamura, H.; Hijikata, A.; Tomizawa-Murasawa, M.; Tanaka, S.; Takagi, S.; Uchida, N.; Suzuki, N.; Sone, A.; Najima, Y.; et al. Identification of therapeutic targets for quiescent, chemotherapy-resistant human leukemia stem cells. Sci. Transl. Med. 2010, 2, 17ra9. [Google Scholar] [CrossRef] [PubMed]
- Majeti, R.; Becker, M.W.; Tian, Q.; Lee, T.-L.M.; Yan, X.; Liu, R.; Chiang, J.-H.; Hood, L.; Clarke, M.F.; Weissman, I.L. Dysregulated gene expression networks in human acute myelogenous leukemia stem cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3396–3401. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, H.J.M.; Woolthuis, C.M.; Vos, A.Z.; Mulder, A.; van den Berg, E.; Kluin, P.M.; van der Weide, K.; de Bont, E.S.J.M.; Huls, G.; Vellenga, E.; et al. Gene expression profiling in the leukemic stem cell-enriched CD34+ fraction identifies target genes that predict prognosis in normal karyotype AML. Leukemia 2011, 25, 1825–1833. [Google Scholar] [CrossRef] [PubMed]
- Huan, J.; Hornick, N.I.; Goloviznina, N.A.; Kamimae- Lanning, A.N.; David, L.L.; Wilmarth, P.A.; Mori, T.; Chevillet, J.R.; Narla, A.; Roberts, C.T.; et al. Coordinate regulation of residual bone marrow function by paracrine trafficking of AML exosomes. Leukemia 2015, 29, 2285–2295. [Google Scholar] [CrossRef] [PubMed]
- Hornick, N.I.; Doron, B.; Abdelhamed, S.; Huan, J.; Harrington, C.A.; Shen, R.; Cambronne, X.A.; Chakkaramakkil Verghese, S.; Kurre, P. AML suppresses hematopoiesis by releasing exosomes that contain microRNAs targeting c-MYB. Sci. Signal 2016, 9, ra88. [Google Scholar] [CrossRef] [PubMed]
- Konopleva, M.Y.; Jordan, C.T. Leukemia stem cells and microenvironment: Biology and therapeutic targeting. J. Clin. Oncol. 2011, 29, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-A.; Shim, J.-S.; Lee, G.-Y.; Yim, H.W.; Kim, T.-M.; Kim, M.; Leem, S.-H.; Lee, J.-W.; Min, C.-K.; Oh, I.-H. Microenvironmental remodeling as a parameter and prognostic factor of heterogeneous leukemogenesis in acute myelogenous leukemia. Cancer Res. 2015, 75, 2222–2231. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C.P.; Tan, R.; Zhou, B.; Yue, S.-B.; Schaffert, S.; Biggs, J.R.; Doyonnas, R.; Lo, M.-C.; Perry, J.M.; Renault, V.M.; et al. MicroRNA programs in normal and aberrant stem and progenitor cells. Genome Res. 2011, 21, 798–810. [Google Scholar] [CrossRef] [PubMed]
- Gangaraju, V.K.; Lin, H. MicroRNAs: Key regulators of stem cells. Nat. Rev. Mol. Cell Biol. 2009, 10, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Wojtowicz, E.E.; Lechman, E.R.; Hermans, K.G.; Schoof, E.M.; Wienholds, E.; Isserlin, R.; van Veelen, P.A.; Broekhuis, M.J.C.; Janssen, G.M.C.; Trotman-Grant, A.; et al. Ectopic miR-125a Expression Induces Long-Term Repopulating Stem Cell Capacity in Mouse and Human Hematopoietic Progenitors. Cell Stem Cell 2016, 19, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Lu, J.; Schlanger, R.; Zhang, H.; Wang, J.Y.; Fox, M.C.; Purton, L.E.; Fleming, H.H.; Cobb, B.; Merkenschlager, M.; et al. MicroRNA miR-125a controls hematopoietic stem cell number. Proc. Natl. Acad. Sci. USA 2010, 107, 14229–14234. [Google Scholar] [CrossRef] [PubMed]
- Felli, N.; Fontana, L.; Pelosi, E.; Botta, R.; Bonci, D.; Facchiano, F.; Liuzzi, F.; Lulli, V.; Morsilli, O.; Santoro, S.; et al. MicroRNAs 221 and 222 inhibit normal erythropoiesis and erythroleukemic cell growth via kit receptor down-modulation. Proc. Natl. Acad. Sci. USA 2005, 102, 18081–18086. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Cammarata, G.; Augugliaro, L.; Salemi, D.; Agueli, C.; La Rosa, M.; Dagnino, L.; Civiletto, G.; Messana, F.; Marfia, A.; Bica, M.G.; et al. Differential expression of specific microRNA and their targets in acute myeloid leukemia. Am. J. Hematol. 2010, 85, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lu, J.; Sun, M.; Mi, S.; Zhang, H.; Luo, R.T.; Chen, P.; Wang, Y.; Yan, M.; Qian, Z.; et al. Distinct microRNA expression profiles in acute myeloid leukemia with common translocations. Proc. Natl. Acad. Sci. USA 2008, 105, 15535–15540. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Garofalo, M.; Martelli, M.P.; Briesewitz, R.; Wang, L.; Fernandez-Cymering, C.; Volinia, S.; Liu, C.-G.; Schnittger, S.; Haferlach, T.; et al. Distinctive microRNA signature of acute myeloid leukemia bearing cytoplasmic mutated nucleophosmin. Proc. Natl. Acad. Sci. USA 2008, 105, 3945–3950. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, D.C.; van den Ancker, W.; Denkers, F.; de Menezes, R.X.; Westers, T.M.; Ossenkoppele, G.J.; van de Loosdrecht, A.A.; Smit, L. MicroRNA profiling can classify acute leukemias of ambiguous lineage as either acute myeloid leukemia or acute lymphoid leukemia. Clin. Cancer Res. 2013, 19, 2187–2196. [Google Scholar] [CrossRef] [PubMed]
- Rubnitz, J.E.; Onciu, M.; Pounds, S.; Shurtleff, S.; Cao, X.; Raimondi, S.C.; Behm, F.G.; Campana, D.; Razzouk, B.I.; Ribeiro, R.C.; et al. Acute mixed lineage leukemia in children: The experience of St Jude Children’s Research Hospital. Blood 2009, 113, 5083–5089. [Google Scholar] [CrossRef] [PubMed]
- Hornick, N.I.; Huan, J.; Doron, B.; Goloviznina, N.A.; Lapidus, J.; Chang, B.H.; Kurre, P. Serum Exosome MicroRNA as a Minimally-Invasive Early Biomarker of AML. Sci. Rep. 2015, 5, 11295. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, D.C.; Denkers, F.; Olthof, M.C.; Rutten, A.P.; Pouwels, W.; Schuurhuis, G.J.; Ossenkoppele, G.J.; Smit, L. Attenuation of microRNA-126 expression that drives CD34+38− stem/progenitor cells in acute myeloid leukemia leads to tumor eradication. Cancer Res. 2014, 74, 2094–2105. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, D.C.; Verhagen, H.J.M.P.; Denkers, F.; Kavelaars, F.G.; Valk, P.J.M.; Schuurhuis, G.J.; Ossenkoppele, G.J.; Smit, L. MicroRNA-551b is highly expressed in hematopoietic stem cells and a biomarker for relapse and poor prognosis in acute myeloid leukemia. Leukemia 2016, 30, 742–746. [Google Scholar] [CrossRef] [PubMed]
- Doulatov, S.; Notta, F.; Eppert, K.; Nguyen, L.T.; Ohashi, P.S.; Dick, J.E. Revised map of the human progenitor hierarchy shows the origin of macrophages and dendritic cells in early lymphoid development. Nat. Immunol. 2010, 11, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Majeti, R.; Park, C.Y.; Weissman, I.L. Identification of a hierarchy of multipotent hematopoietic progenitors in human cord blood. Cell Stem Cell 2007, 1, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Edvardsson, L.; Dykes, J.; Olofsson, T. Isolation and characterization of human myeloid progenitor populations—TpoR as discriminator between common myeloid and megakaryocyte/erythroid progenitors. Exp. Hematol. 2006, 34, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Manz, M.G.; Miyamoto, T.; Akashi, K.; Weissman, I.L. Prospective isolation of human clonogenic common myeloid progenitors. Proc. Natl. Acad. Sci. USA 2002, 99, 11872–11877. [Google Scholar] [CrossRef] [PubMed]
- Summers, Y.J.; Heyworth, C.M.; de Wynter, E.A.; Hart, C.A.; Chang, J.; Testa, N.G. AC133+ G0 Cells from Cord Blood Show a High Incidence of Long-Term Culture-Initiating Cells and a Capacity for More Than 100 Million-Fold Amplification of Colony-Forming Cells In Vitro. Stem Cells 2004, 22, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Yasui, K.; Yamashita, N.; Horie, Y.; Yamada, T.; Tani, Y.; Shibata, H.; Nakano, T. In Vitro Proliferation Potential of AC133 Positive Cells in Peripheral Blood. Stem Cells 2000, 18, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Liao, R.; Sun, J.; Zhang, L.; Lou, G.; Chen, M.; Zhou, D.; Chen, Z.; Zhang, S. MicroRNAs play a role in the development of human hematopoietic stem cells. J. Cell Biochem. 2008, 104, 805–817. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Chaudhuri, A.A.; Rao, D.S.; Gibson, W.S.J.; Balazs, A.B.; Baltimore, D. MicroRNAs enriched in hematopoietic stem cells differentially regulate long-term hematopoietic output. Proc. Natl. Acad. Sci. USA 2010, 107, 14235–14240. [Google Scholar] [CrossRef] [PubMed]
- Georgantas, R.W.; Hildreth, R.; Morisot, S.; Alder, J.; Liu, C.; Heimfeld, S.; Calin, G.A.; Croce, C.M.; Civin, C.I. CD34+ hematopoietic stem-progenitor cell microRNA expression and function: A circuit diagram of differentiation control. Proc. Natl. Acad. Sci. USA 2007, 104, 2750–2755. [Google Scholar] [CrossRef] [PubMed]
- Merkerova, M.; Vasikova, A.; Belickova, M.; Bruchova, H. MicroRNA expression profiles in umbilical cord blood cell lineages. Stem Cells Dev. 2010, 19, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Jin, P.; Wang, E.; Ren, J.; Childs, R.; Shin, J.W.; Khuu, H.; Marincola, F.M.; Stroncek, D.F. Differentiation of two types of mobilized peripheral blood stem cells by microRNA and cDNA expression analysis. J. Transl. Med. 2008, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Bissels, U.; Wild, S.; Tomiuk, S.; Hafner, M.; Scheel, H.; Mihailovic, A.; Choi, Y.-H.; Tuschl, T.; Bosio, A. Combined characterization of microRNA and mRNA profiles delineates early differentiation pathways of CD133+ and CD34+ hematopoietic stem and progenitor cells. Stem Cells 2011, 29, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.-C.; Park, C.Y.; Bhagat, G.; Zhang, J.; Wang, Y.; Fan, J.-B.; Liu, M.; Zou, Y.; Weissman, I.L.; Gu, H. microRNA-29a induces aberrant self-renewal capacity in hematopoietic progenitors, biased myeloid development, and acute myeloid leukemia. J. Exp. Med. 2010, 207, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Ooi, A.G.L.; Sahoo, D.; Adorno, M.; Wang, Y.; Weissman, I.L.; Park, C.Y. MicroRNA-125b expands hematopoietic stem cells and enriches for the lymphoid-balanced and lymphoid-biased subsets. Proc. Natl. Acad. Sci. USA 2010, 107, 21505–21510. [Google Scholar]
- Hu, W.; Dooley, J.; Chung, S.S.; Chandramohan, D.; Cimmino, L.; Mukherjee, S.; Mason, C.E.; de Strooper, B.; Liston, A.; Park, C.Y. miR-29a maintains mouse hematopoietic stem cell self-renewal by regulating Dnmt3a. Blood 2015, 125, 2206–2216. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, A.; Walasek, M.A.; Olthof, S.; Weersing, E.; Ritsema, M.; Zwart, E.; van Os, R.; Bystrykh, L.V.; de Haan, G. Genetic screen identifies microRNA cluster 99b/let-7e/125a as a regulator of primitive hematopoietic cells. Blood 2012, 119, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Gentner, B.; Visigalli, I.; Hiramatsu, H.; Lechman, E.; Ungari, S.; Giustacchini, A.; Schira, G.; Amendola, M.; Quattrini, A.; Martino, S.; et al. Identification of hematopoietic stem cell-specific miRNAs enables gene therapy of globoid cell leukodystrophy. Sci. Transl. Med. 2010, 2, 58ra84. [Google Scholar] [CrossRef] [PubMed]
- Lechman, E.R.; Gentner, B.; Ng, S.W.K; Schoof, E.M.; van Galen, P.; Kennedy, J.A.; Nucera, S.; Ciceri, F.; Kaufmann, K.B.; Takayama, N.; et al. miR-126 Regulates Distinct Self-Renewal Outcomes in Normal and Malignant Hematopoietic Stem Cells. Cancer Cell 2016, 29, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Copley, M.R.; Babovic, S.; Benz, C.; Knapp, D.J.H.F.; Beer, P.A.; Kent, D.G.; Wohrer, S.; Treloar, D.Q.; Day, C.; Rowe, K.; et al. The Lin28b-let-7-Hmga2 axis determines the higher self-renewal potential of fetal haematopoietic stem cells. Nat. Cell Biol. 2013, 15, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, X.; Timani, K.A.; Broxmeyer, H.E.; He, J.J. MicroRNA-124 Targets Tip110 Expression and Regulates Hematopoiesis. Stem Cells Dev. 2015, 24, 2009–2017. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.; Nguyen, D.; Chen, C.; Shields, L.; Lodish, H.F. MicroRNA-125b transforms myeloid cell lines by repressing multiple mRNA. Haematologica 2012, 97, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Le, M.T.N.; Shyh-Chang, N.; Khaw, S.L.; Chin, L.; Teh, C.; Tay, J.; O’Day, E.; Korzh, V.; Yang, H.; Lal, A.; et al. Conserved regulation of p53 network dosage by microRNA-125b occurs through evolving miRNA-target gene pairs. PLoS Genet. 2011, 7, e1002242. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.; Harris, M.H.; Zhou, B.; Lodish, H.F. MicroRNA miR-125b causes leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 21558–21563. [Google Scholar] [CrossRef] [PubMed]
- Surdziel, E.; Cabanski, M.; Dallmann, I.; Lyszkiewicz, M.; Krueger, A.; Ganser, A.; Scherr, M.; Eder, M. Enforced expression of miR-125b affects myelopoiesis by targeting multiple signaling pathways. Blood 2011, 117, 4338–4348. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.A.; So, A.Y.-L.; Mehta, A.; Minisandram, A.; Sinha, N.; Jonsson, V.D.; Rao, D.S.; O’Connell, R.M.; Baltimore, D. Oncomir miR-125b regulates hematopoiesis by targeting the gene Lin28A. Proc. Natl. Acad. Sci. USA 2012, 109, 4233–4238. [Google Scholar] [CrossRef] [PubMed]
- Jinlong, S.; Lin, F.; Yonghui, L.; Li, Y.; Weidong, W. Identification of let-7a-2-3p or/and miR-188-5p as Prognostic Biomarkers in Cytogenetically Normal Acute Myeloid Leukemia. PLoS ONE 2015, 10, e0118099. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-Y.; Zhang, X.-J.; Feng, D.-D.; Zhang, H.; Zeng, C.-W.; Han, B.-W.; Zhou, A.-D.; Qu, L.-H.; Xu, L.; Chen, Y.-Q. miR-125b, a target of CDX2, regulates cell differentiation through repression of the core binding factor in hematopoietic malignancies. J. Biol. Chem. 2011, 286, 38253–38263. [Google Scholar] [CrossRef] [PubMed]
- Emmrich, S.; Rasche, M.; Schöning, J.; Reimer, C.; Keihani, S.; Maroz, A.; Xie, Y.; Li, Z.; Schambach, A.; Reinhardt, D.; et al. miR-99a/100~125b tricistrons regulate hematopoietic stem and progenitor cell homeostasis by shifting the balance between TGFβ and Wnt signaling. Genes Dev. 2014, 28, 858–874. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.-F.; Hu, Y.-L.; Uttarwar, L.; Passegue, E.; Largman, C. MicroRNA-126 regulates HOXA9 by binding to the homeobox. Mol. Cell Biol. 2008, 28, 4609–4619. [Google Scholar] [CrossRef] [PubMed]
- Lechman, E.R.; Gentner, B.; van Galen, P.; Giustacchini, A.; Saini, M.; Boccalatte, F.E.; Hiramatsu, H.; Restuccia, U.; Bachi, A.; Voisin, V.; et al. Attenuation of miR-126 Activity Expands HSC In Vivo without Exhaustion. Cell Stem Cell 2012, 11, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Zhao, J.L.; Sinha, N.; Marinov, G.K.; Mann, M.; Kowalczyk, M.S.; alimidi, R.P.; Du, X.; Erikci, E.; Regev, A.; et al. The MicroRNA-132 and MicroRNA-212 Cluster Regulates Hematopoietic Stem Cell Maintenance and Survival with Age by Buffering FOXO3 Expression. Immunity 2015, 42, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Starczynowski, D.T.; Kuchenbauer, F.; Wegrzyn, J.; Rouhi, A.; Petriv, O.; Hansen, C.L.; Humphries, R.K.; Karsan, A. MicroRNA-146a disrupts hematopoietic differentiation and survival. Exp. Hematol. 2011, 39, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Starczynowski, D.T.; Kuchenbauer, F.; Argiropoulos, B.; Sung, S.; Morin, R.; Muranyi, A.; Humphries, R.K.; Karsan, A. Identification of miR-145 and miR-146a as mediators of the 5q- syndrome phenotype. Nat. Med. 2010, 16, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.L.; Rao, D.S.; O’Connell, R.M.; Garcia-Flores, Y.; Baltimore, D. MicroRNA-146a acts as a guardian of the quality and longevity of hematopoietic stem cells in mice. Elife 2013, 2, e00537. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Li, Z.; Chen, P.; He, C.; Cao, D.; Elkahloun, A.; Lu, J.; Pelloso, L.A.; Wunderlich, M.; Huang, H.; et al. Aberrant overexpression and function of the miR-17-92 cluster in MLL-rearranged acute leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 3710–3715. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. c-Myc-regulated microRNAs modulate E2F1 expression. Nature 2005, 435, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Vecchiarelli-Federico, L.M.; Li, Y.-J.; Egan, S.E.; Spaner, D.; Hough, M.R.; Ben-David, Y. The miR-17-92 cluster expands multipotent hematopoietic progenitors whereas imbalanced expression of its individual oncogenic miRNAs promotes leukemia in mice. Blood 2012, 119, 4486–4498. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Srinivasan, L.; Calado, D.P.; Patterson, H.C.; Zhang, B.; Wang, J.; Henderson, J.M.; Kutok, J.L.; Rajewsky, K. Lymphoproliferative disease and autoimmunity in mice with increased miR-17-92 expression in lymphocytes. Nat. Immunol. 2008, 9, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, H.; Chen, P.; He, M.; Li, Y.; Arnovitz, S.; Jiang, X.; He, C.; Hyjek, E.; Zhang, J.; et al. miR-196b directly targets both HOXA9/MEIS1 oncogenes and FAS tumour suppressor in MLL-rearranged leukaemia. Nat. Commun. 2012, 3, 688. [Google Scholar] [CrossRef] [PubMed]
- Yekta, S.; Shih, I.-H.; Bartel, D.P. MicroRNA-directed cleavage of HOXB8 mRNA. Science 2004, 304, 594–596. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Rich, A.; Dahl, R. MiR-24 promotes the survival of hematopoietic cells. PLoS ONE 2013, 8, e55406. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Merchan, A.; Cerrato, C.; Luengo, G.; Dominguez, O.; Piris, M.A.; Serrano, M.; Gonzalez, S. miR-33-mediated downregulation of p53 controls hematopoietic stem cell self-renewal. Cell Cycle. 2010, 9, 3277–3285. [Google Scholar] [CrossRef] [PubMed]
- Song, S.J.; Ito, K.; Ala, U.; Kats, L.; Webster, K.; Sun, S.M.; Jongen-Lavrencic, M.; Manova-Todorova, K.; Teruya-Feldstein, J.; Avigan, D.E.; et al. The oncogenic microRNA miR-22 targets the TET2 tumor suppressor to promote hematopoietic stem cell self-renewal and transformation. Cell Stem Cell 2013, 13, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Meenhuis, A.; van Veelen, P.A.; de Looper, H.; van Boxtel, N.; van den Berge, I.J.; Sun, S.M.; Taskesen, E.; Stern, P.; de Ru, A.H.; van Adrichem, A.J.; et al. MiR-17/20/93/106 promote hematopoietic cell expansion by targeting sequestosome 1-regulated pathways in mice. Blood 2011, 118, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Neilson, J.R.; Zheng, G.X.Y.; Burge, C.B.; Sharp, P.A. Dynamic regulation of miRNA expression in ordered stages of cellular development. Genes Dev. 2007, 21, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Z.; Li, L.; Lodish, H.F.; Bartel, D.P. MicroRNAs Modulate Hematopoietic Lineage Differentiation. Science 2004, 303, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.-J.; Chau, J.; Ebert, P.J.R.; Sylvester, G.; Min, H.; Liu, G.; Braich, R.; Manoharan, M.; Soutschek, J.; Skare, P.; et al. miR-181a Is an Intrinsic Modulator of T Cell Sensitivity and Selection. Cell 2007, 129, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-S.; Gong, J.-N.; Yu, J.; Wang, F.; Zhang, X.-H.; Yin, X.-L.; Tan, Z.-Q.; Luo, Z.-M.; Yang, G.-H.; Shen, C.; et al. MicroRNA-29a and microRNA-142-3p are regulators of myeloid differentiation and acute myeloid leukemia. Blood 2012, 119, 4992–5004. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; Boldin, M.P.; Taganov, K.D.; Nicoll, J.; Paquette, R.L.; Baltimore, D. Sustained expression of microRNA-155 in hematopoietic stem cells causes a myeloproliferative disorder. J. Exp. Med. 2008, 205, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.T.; Häger, M.; Glenthøj, A.; Asmar, F.; Clemmensen, S.N.; Mora-Jensen, H.; Borregaard, N.; Cowland, J.B. miRNA-130a regulates C/EBP-ε expression during granulopoiesis. Blood 2014, 123, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Pelosi, E.; Greco, P.; Racanicchi, S.; Testa, U.; Liuzzi, F.; Croce, C.M.; Brunetti, E.; Grignani, F.; Peschle, C. MicroRNAs 17-5p-20a-106a control monocytopoiesis through AML1 targeting and M-CSF receptor upregulation. Nat. Cell Biol. 2007, 9, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Johnnidis, J.B.; Harris, M.H.; Wheeler, R.T.; Stehling-Sun, S.; Lam, M.H.; Kirak, O.; Brummelkamp, T.R.; Fleming, M.D.; Camargo, F.D. Regulation of progenitor cell proliferation and granulocyte function by microRNA-223. Nature 2008, 451, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Zardo, G.; Ciolfi, A.; Vian, L.; Starnes, L.M.; Billi, M.; Racanicchi, S.; Maresca, C.; Fazi, F.; Travaglini, L.; Noguera, N.; et al. Polycombs and microRNA-223 regulate human granulopoiesis by transcriptional control of target gene expression. Blood 2012, 119, 4034–4046. [Google Scholar] [CrossRef] [PubMed]
- Fazi, F.; Rosa, A.; Fatica, A.; Gelmetti, V.; De Marchis, M.L.; Nervi, C.; Bozzoni, I. A Minicircuitry Comprised of MicroRNA-223 and Transcription Factors NFI-A and C/EBPα Regulates Human Granulopoiesis. Cell 2005, 123, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Pulikkan, J.A.; Dengler, V.; Peramangalam, P.S.; Peer Zada, A.A.; Müller-Tidow, C.; Bohlander, S.K.; Tenen, D.G.; Behre, G. Cell-cycle regulator E2F1 and microRNA-223 comprise an autoregulatory negative feedback loop in acute myeloid leukemia. Blood 2010, 115, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Iwama, A.; Satake, M.; Kohu, K. MicroRNA-27 enhances differentiation of myeloblasts into granulocytes by post-transcriptionally downregulating Runx1. Br. J. Haematol. 2009, 145, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Katzerke, C.; Madan, V.; Gerloff, D.; Bräuer-Hartmann, D.; Hartmann, J.-U.; Wurm, A.A.; Müller-Tidow, C.; Schnittger, S.; Tenen, D.G.; Niederwieser, D.; et al. Transcription factor C/EBPα-induced microRNA-30c inactivates Notch1 during granulopoiesis and is downregulated in acute myeloid leukemia. Blood 2013, 122, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Pulikkan, J.A.; Peramangalam, P.S.; Dengler, V.; Ho, P.A.; Preudhomme, C.; Meshinchi, S.; Christopeit, M.; Nibourel, O.; Müller-Tidow, C.; Bohlander, S.K.; et al. C/EBPα regulated microRNA-34a targets E2F3 during granulopoiesis and is down-regulated in AML with CEBPA mutations. Blood 2010, 116, 5638–5649. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.; Ballarino, M.; Sorrentino, A.; Sthandier, O.; De Angelis, F.G.; Marchioni, M.; Masella, B.; Guarini, A.; Fatica, A.; Peschle, C.; et al. The interplay between the master transcription factor PU.1 and miR-424 regulates human monocyte/macrophage differentiation. Proc. Natl. Acad. Sci. USA 2007, 104, 19849–19854. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Bulgarelli, J.; Ruberti, S.; Rontauroli, S.; Sacchi, G.; Norfo, R.; Pennucci, V.; Zini, R.; Salati, S.; Prudente, Z.; et al. MYB controls erythroid versus megakaryocyte lineage fate decision through the miR-486-3p-mediated downregulation of MAF. Cell Death Differ. 2015, 22, 1906–1921. [Google Scholar] [CrossRef] [PubMed]
- Kamat, V.; Paluru, P.; Myint, M.; French, D.L.; Gadue, P.; Diamond, S.L. MicroRNA screen of human embryonic stem cell differentiation reveals miR-105 as an enhancer of megakaryopoiesis from adult CD34+ cells. Stem Cells 2014, 32, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hu, C.; Arnovitz, S.; Bugno, J.; Yu, M.; Zuo, Z.; Chen, P.; Huang, H.; Ulrich, B.; Gurbuxani, S.; et al. miR-22 has a potent anti-tumour role with therapeutic potential in acute myeloid leukaemia. Nat. Commun. 2016, 7, 11452. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Chen, M.-T.; Zhang, X.-H.; Yin, X.-L.; Ning, H.-M.; Su, R.; Lin, H.-S.; Song, L.; Wang, F.; Ma, Y.-N.; et al. The PU.1-Modulated MicroRNA-22 Is a Regulator of Monocyte/Macrophage Differentiation and Acute Myeloid Leukemia. PLoS Genet. 2016, 12, e1006259. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes-Sternberg, C.; Meerson, A.; Shaked, I.; Soreq, H. MicroRNA modulation of megakaryoblast fate involves cholinergic signaling. Leuk. Res. 2006, 30, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, J.; Gao, L.; McClellan, S.; Finan, M.A.; Butler, T.W.; Owen, L.B.; Piazza, G.A.; Xi, Y. MiR-181 mediates cell differentiation by interrupting the Lin28 and let-7 feedback circuit. Cell Death Differ. 2012, 19, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Klusmann, J.-H.; Li, Z.; Böhmer, K.; Maroz, A.; Koch, M.L.; Emmrich, S.; Godinho, F.J.; Orkin, S.H.; Reinhardt, D. miR-125b-2 is a potential oncomiR on human chromosome 21 in megakaryoblastic leukemia. Genes Dev. 2010, 24, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Grabher, C.; Payne, E.M.; Johnston, A.B.; Bolli, N.; Lechman, E.; Dick, J.E.; Kanki, J.P.; Look, A.T. Zebrafish microRNA-126 determines hematopoietic cell fate through c-Myb. Leukemia 2011, 25, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.S.; Narla, A.; Nonami, A.; Mullally, A.; Dimitrova, N.; Ball, B.; McAuley, J.R.; Poveromo, L.; Kutok, J.L.; Galili, N.; et al. Coordinate loss of a microRNA and protein-coding gene cooperate in the pathogenesis of 5q- syndrome. Blood 2011, 118, 4666–4673. [Google Scholar] [CrossRef] [PubMed]
- Labbaye, C.; Spinello, I.; Quaranta, M.T.; Pelosi, E.; Pasquini, L.; Petrucci, E.; Biffoni, M.; Nuzzolo, E.R.; Billi, M.; Foà, R.; et al. A three-step pathway comprising PLZF/miR-146a/CXCR4 controls megakaryopoiesis. Nat. Cell Biol. 2008, 10, 788–801. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Kalota, A.; Jin, S.; Gewirtz, A.M. The c-myb proto-oncogene and microRNA-15a comprise an active autoregulatory feedback loop in human hematopoietic cells. Blood 2009, 113, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Guo, S.; Ebert, B.L.; Zhang, H.; Peng, X.; Bosco, J.; Pretz, J.; Schlanger, R.; Wang, J.Y.; Mak, R.H.; et al. MicroRNA-mediated control of cell fate in megakaryocyte-erythrocyte progenitors. Dev. Cell 2008, 14, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Barroga, C.F.; Pham, H.; Kaushansky, K. Thrombopoietin regulates c-Myb expression by modulating micro RNA 150 expression. Exp. Hematol. 2008, 36, 1585–1592. [Google Scholar] [CrossRef] [PubMed]
- Romania, P.; Lulli, V.; Pelosi, E.; Biffoni, M.; Peschle, C.; Marziali, G. MicroRNA 155 modulates megakaryopoiesis at progenitor and precursor level by targeting Ets-1 and Meis1 transcription factors. Br. J. Haematol. 2008, 143, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bai, H.; Zhang, Z.; Li, W.; Dong, L.; Wei, X.; Ma, Y.; Zhang, J.; Yu, J.; Sun, G.; et al. The up-regulation of miR-199b-5p in erythroid differentiation is associated with GATA-1 and NF-E2. Mol. Cells 2014, 37, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.-Y.; Wang, F.; Yu, J.; Yang, G.-H.; Liu, X.-L.; Zhang, J.-W. MicroRNA-223 reversibly regulates erythroid and megakaryocytic differentiation of K562 cells. J. Cell. Mol. Med. 2009, 13, 4551–4559. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, D.; Wang, F.; Li, T.; Dong, L.; Liu, H.; Ma, Y.; Jiang, F.; Yin, H.; Yan, W.; et al. A comprehensive analysis of GATA-1-regulated miRNAs reveals miR-23a to be a positive modulator of erythropoiesis. Nucleic Acids Res. 2013, 41, 4129–4143. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhu, Y.; Guo, L.; Dong, L.; Liu, H.; Yin, H.; Zhang, Z.; Li, Y.; Liu, C.; Ma, Y.; et al. A regulatory circuit comprising GATA1/2 switch and microRNA-27a/24 promotes erythropoiesis. Nucleic Acids Res. 2014, 42, 442–457. [Google Scholar] [CrossRef] [PubMed]
- Tenedini, E.; Roncaglia, E.; Ferrari, F.; Orlandi, C.; Bianchi, E.; Bicciato, S.; Tagliafico, E.; Ferrari, S. Integrated analysis of microRNA and mRNA expression profiles in physiological myelopoiesis: Role of hsa-mir-299-5p in CD34+ progenitor cells commitment. Cell Death Dis. 2010, 1, e28. [Google Scholar] [CrossRef] [PubMed]
- Navarro, F.; Gutman, D.; Meire, E.; Cáceres, M.; Rigoutsos, I.; Bentwich, Z.; Lieberman, J. miR-34a contributes to megakaryocytic differentiation of K562 cells independently of p53. Blood 2009, 114, 2181–2192. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yu, J.; Yang, G.-H.; Wang, X.-S.; Zhang, J.-W. Regulation of erythroid differentiation by miR-376a and its targets. Cell Res. 2011, 21, 1196–1209. [Google Scholar] [CrossRef] [PubMed]
- Dore, L.C.; Amigo, J.D.; Dos Santos, C.O.; Zhang, Z.; Gai, X.; Tobias, J.W.; Yu, D.; Klein, A.M.; Dorman, C.; Wu, W.; et al. A GATA-1-regulated microRNA locus essential for erythropoiesis. Proc. Natl. Acad. Sci. USA 2008, 105, 3333–3338. [Google Scholar] [CrossRef] [PubMed]
- Pase, L.; Layton, J.E.; Kloosterman, W.P.; Carradice, D.; Waterhouse, P.M.; Lieschke, G.J. miR-451 regulates zebrafish erythroid maturation in vivo via its target gata2. Blood 2009, 113, 1794–1804. [Google Scholar] [CrossRef] [PubMed]
- Kamesaki, H.; Michaud, G.Y.; Irving, S.G.; Suwabe, N.; Kamesaki, S.; Okuma, M.; Cossman, J. TPA-induced arrest of erythroid differentiation is coupled with downregulation of GATA-1 and upregulation of GATA-2 in an erythroid cell line SAM-1. Blood 1996, 87, 999–1005. [Google Scholar] [PubMed]
- Lulli, V.; Romania, P.; Morsilli, O.; Cianciulli, P.; Gabbianelli, M.; Testa, U.; Giuliani, A.; Marziali, G. MicroRNA-486-3p regulates γ-globin expression in human erythroid cells by directly modulating BCL11A. PLoS ONE 2013, 8, e60436. [Google Scholar] [CrossRef] [PubMed]
- Zhai, P.-F.; Wang, F.; Su, R.; Lin, H.-S.; Jiang, C.-L.; Yang, G.-H.; Yu, J.; Zhang, J.-W. The regulatory roles of microRNA-146b-5p and its target platelet-derived growth factor receptor α (PDGFRA) in erythropoiesis and megakaryocytopoiesis. J. Biol. Chem. 2014, 289, 22600–22613. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Volinia, S.; Liu, C.-G.; Fernandez-Cymering, C.; Palumbo, T.; Pichiorri, F.; Fabbri, M.; Coombes, K.; Alder, H.; Nakamura, T.; et al. MicroRNA signatures associated with cytogenetics and prognosis in acute myeloid leukemia. Blood 2008, 111, 3183–3189. [Google Scholar] [CrossRef] [PubMed]
- Feller, N.; van der Velden, V.H.J.; Brooimans, R.A.; Boeckx, N.; Preijers, F.; Kelder, A.; de Greef, I.; Westra, G.; te Marvelde, J.G.; Aerts, P.; et al. Defining consensus leukemia-associated immunophenotypes for detection of minimal residual disease in acute myeloid leukemia in a multicenter setting. Blood Cancer J. 2013, 3, e129. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Upchurch, D.; Szilvassy, S.J.; Guzman, M.L.; Howard, D.S.; Pettigrew, A.L.; Meyerrose, T.; Rossi, R.; Grimes, B.; Rizzieri, D.A.; et al. The interleukin-3 receptor alpha chain is a unique marker for human acute myelogenous leukemia stem cells. Leukemia 2000, 14, 1777–1784. [Google Scholar] [CrossRef] [PubMed]
- Wouters, R.; Cucchi, D.; Kaspers, G.J.L.; Schuurhuis, G.J.; Cloos, J. Relevance of leukemic stem cells in acute myeloid leukemia: Heterogeneity and influence on disease monitoring, prognosis and treatment design. Expert Rev. Hematol. 2014, 7, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Liu, Y.; Wang, Y.; Zhang, Y.; Luo, Q.; Man, X.; Wei, F.; Yu, X. Downregulation of Foxo3 and TRIM31 by miR-551b in side population promotes cell proliferation, invasion, and drug resistance of ovarian cancer. Med. Oncol. 2016, 33, 126. [Google Scholar] [CrossRef] [PubMed]
- Chaluvally-Raghavan, P.; Jeong, K.J.; Pradeep, S.; Silva, A.M.; Yu, S.; Liu, W.; Moss, T.; Rodriguez-Aguayo, C.; Zhang, D.; Ram, P.; et al. Direct Upregulation of STAT3 by MicroRNA-551b-3p Deregulates Growth and Metastasis of Ovarian Cancer. Cell Rep. 2016, 15, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wells, A.; Padilla, M.T.; Kato, K.; Kim, K.C.; Lin, Y. A signaling pathway consisting of miR-551b, catalase and MUC1 contributes to acquired apoptosis resistance and chemoresistance. Carcinogenesis 2014, 35, 2457–2466. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Heaphy, C.E.A.; Havelange, V.; Fabbri, M.; Volinia, S.; Tsao, T.; Zanesi, N.; Kornblau, S.M.; Marcucci, G.; Calin, G.A.; et al. MicroRNA 29b functions in acute myeloid leukemia. Blood 2009, 114, 5331–5341. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Schwind, S.; Yu, B.; Santhanam, R.; Wang, H.; Hoellerbauer, P.; Mims, A.; Klisovic, R.; Walker, A.R.; Chan, K.K.; et al. Targeted delivery of microRNA-29b by transferrin-conjugated anionic lipopolyplex nanoparticles: A novel therapeutic strategy in acute myeloid leukemia. Clin. Cancer Res. 2013, 19, 2355–2367. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Liu, S.; Fabbri, M.; Liu, Z.; Heaphy, C.E.A.; Callegari, E.; Schwind, S.; Pang, J.; Yu, J.; Muthusamy, N.; et al. MicroRNA-29b induces global DNA hypomethylation and tumor suppressor gene reexpression in acute myeloid leukemia by targeting directly DNMT3A and 3B and indirectly DNMT1. Blood 2009, 113, 6411–6418. [Google Scholar] [CrossRef] [PubMed]
- Blum, W.; Garzon, R.; Klisovic, R.B.; Schwind, S.; Walker, A.; Geyer, S.; Liu, S.; Havelange, V.; Becker, H.; Schaaf, L.; et al. Clinical response and miR-29b predictive significance in older AML patients treated with a 10-day schedule of decitabine. Proc. Natl. Acad. Sci. USA 2010, 107, 7473–7478. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.R.; Higgins, R.R.; Grutkoski, P.S.; Bousquet, M.; Quelen, C.; Bartholomaus, L.M.; Brousset, P. Myeloid neoplasm with translocation t(2;11)(p21;q23-24), elevated microRNA 125b-1, and JAK2 exon 12 mutation. Br. J. Haematol. 2015, 169, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, M.; Quelen, C.; Rosati, R.; Mansat-De Mas, V.; La Starza, R.; Bastard, C.; Lippert, E.; Talmant, P.; Lafage-Pochitaloff, M.; Leroux, D.; et al. Myeloid cell differentiation arrest by miR-125b-1 in myelodysplastic syndrome and acute myeloid leukemia with the t(2;11)(p21;q23) translocation. J. Exp. Med. 2008, 205, 2499–2506. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, G.; Radmacher, M.D.; Maharry, K.; Mrózek, K.; Ruppert, A.S.; Paschka, P.; Vukosavljevic, T.; Whitman, S.P.; Baldus, C.D.; Langer, C.; et al. MicroRNA expression in cytogenetically normal acute myeloid leukemia. N. Engl. J. Med. 2008, 358, 1919–1928. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, G.; Maharry, K.; Radmacher, M.D.; Mrózek, K.; Vukosavljevic, T.; Paschka, P.; Whitman, S.P.; Langer, C.; Baldus, C.D.; Liu, C.-G.; et al. Prognostic significance of, and gene and microRNA expression signatures associated with, CEBPA mutations in cytogenetically normal acute myeloid leukemia with high-risk molecular features: A Cancer and Leukemia Group B Study. J. Clin. Oncol. 2008, 26, 5078–5087. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, H.; Li, Y.; Jiang, X.; Chen, P.; Arnovitz, S.; Radmacher, M.D.; Maharry, K.; Elkahloun, A.; Yang, X.; et al. Up-regulation of a HOXA-PBX3 homeobox-gene signature following down-regulation of miR-181 is associated with adverse prognosis in patients with cytogenetically abnormal AML. Blood 2012, 119, 2314–2324. [Google Scholar] [CrossRef] [PubMed]
- Isken, F.; Steffen, B.; Merk, S.; Dugas, M.; Markus, B.; Tidow, N.; Zühlsdorf, M.; Illmer, T.; Thiede, C.; Berdel, W.E.; et al. Identification of acute myeloid leukaemia associated microRNA expression patterns. Br. J. Haematol. 2008, 140, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Roscigno, G.; Quintavalle, C.; Donnarumma, E.; Puoti, I.; Diaz-Lagares, A.; Iaboni, M.; Fiore, D.; Russo, V.; Todaro, M.; Romano, G.; et al. MiR-221 promotes stemness of breast cancer cells by targeting DNMT3b. Oncotarget 2010, 1, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, L.; Ischenko, I.; Bao, Q.; Schwarz, B.; Nieß, H.; Wang, Y.; Renner, A.; Mysliwietz, J.; Jauch, K.-W.; et al. Antisense inhibition of microRNA-21 and microRNA-221 in tumor-initiating stem-like cells modulates tumorigenesis, metastasis, and chemotherapy resistance in pancreatic cancer. Target Oncol. 2015, 10, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Riccioni, R.; Lulli, V.; Castelli, G.; Biffoni, M.; Tiberio, R.; Pelosi, E.; Lo-Coco, F.; Testa, U. miR-21 is overexpressed in NPM1-mutant acute myeloid leukemias. Leuk. Res. 2015, 39, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Zhu, X.; Li, Y.; Dong, D.; Yao, J.; Lin, C.; Huang, K.; Hu, H.; Fei, J. miRNA-21 regulates arsenic-induced anti-leukemia activity in myelogenous cell lines. Med. Oncol. 2011, 28, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, X.; Gu, J.; Hu, H.; Dong, D.; Yao, J.; Lin, C.; Fei, J. Anti-miR-21 oligonucleotide enhances chemosensitivity of leukemic HL60 cells to arabinosylcytosine by inducing apoptosis. Hematology 2010, 15, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; Xu, R.; Cao, Z.; Wei, D.; Wang, C. Involvement of miR-21 in resistance to daunorubicin by regulating PTEN expression in the leukaemia K562 cell line. FEBS Lett. 2011, 585, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, Y.; Gu, J.; Zhu, X.; Dong, D.; Yao, J.; Lin, C.; Fei, J. Antisense oligonucleotide against miR-21 inhibits migration and induces apoptosis in leukemic K562 cells. Leuk. Lymphoma 2010, 51, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Schopman, N.C.T.; Heynen, S.; Haasnoot, J.; Berkhout, B. A miRNA-tRNA mix-up: tRNA origin of proposed miRNA. RNA Biol. 2010, 7, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-J.; Liu, G.-H.; Ye, Y.-W.; Fu, Y.; Zhang, X.-F. The role of microRNA-1274a in the tumorigenesis of gastric cancer: Accelerating cancer cell proliferation and migration via directly targeting FOXO4. Biochem. Biophys. Res. Commun. 2015, 459, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S. A Novel Type of Non-coding RNA, nc886, Implicated in Tumor Sensing and Suppression. Genom. Inform. 2015, 13, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Kunkeaw, N.; Jeon, S.H.; Lee, I.; Johnson, B.H.; Kang, G.-Y.; Bang, J.Y.; Park, H.S.; Leelayuwat, C.; Lee, Y.S. Precursor miR-886, a novel noncoding RNA repressed in cancer, associates with PKR and modulates its activity. RNA 2011, 17, 1076–1089. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Lee, K.; Jang, H.-J.; Lee, G.K.; Park, J.-L.; Kim, S.-Y.; Kim, S.-B.; Johnson, B.H.; Zo, J.I.; Lee, J.-S.; et al. Epigenetic silencing of the non-coding RNA nc886 provokes oncogenes during human esophageal tumorigenesis. Oncotarget 2014, 5, 3472–3481. [Google Scholar] [CrossRef] [PubMed]
- Oler, A.J.; Alla, R.K.; Roberts, D.N.; Wong, A.; Hollenhorst, P.C.; Chandler, K.J.; Cassiday, P.A.; Nelson, C.A.; Hagedorn, C.H.; Graves, B.J.; et al. Human RNA polymerase III transcriptomes and relationships to Pol II promoter chromatin and enhancer-binding factors. Nat. Struct. Mol. Biol. 2010, 17, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Treppendahl, M.B.; Qiu, X.; Søgaard, A.; Yang, X.; Nandrup-Bus, C.; Hother, C.; Andersen, M.K.; Kjeldsen, L.; Möllgaard, L.; Hellström-Lindberg, E.; et al. Allelic methylation levels of the noncoding VTRNA2-1 located on chromosome 5q31.1 predict outcome in AML. Blood 2012, 119, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Helbo, A.S.; Treppendahl, M.; Aslan, D.; Dimopoulos, K.; Nandrup-Bus, C.; Holm, M.S.; Andersen, M.K.; Liang, G.; Kristensen, L.S.; Grønbæk, K. Hypermethylation of the VTRNA1-3 Promoter is Associated with Poor Outcome in Lower Risk Myelodysplastic Syndrome Patients. Genes 2015, 6, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Song, Y.; Bi, N.; Shen, J.; Liu, W.; Fan, J.; Sun, G.; Tong, T.; He, J.; Shi, Y.; et al. DNA methylation-mediated repression of miR-886–3p predicts poor outcome of human small cell lung cancer. Cancer Res. 2013, 73, 3326–3335. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Collin, J.; Zhu, L.; Montaner, D.; Armstrong, L.; Neganova, I.; Lako, M. A Novel Role for miR-1305 in Regulation of Pluripotency-Differentiation Balance, Cell Cycle, and Apoptosis in Human Pluripotent Stem Cells. Stem Cells 2016, 34, 2306–2317. [Google Scholar] [CrossRef] [PubMed]
- Dorrance, A.M.; Neviani, P.; Ferenchak, G.J.; Huang, X.; Nicolet, D.; Maharry, K.S.; Ozer, H.G.; Hoellarbauer, P.; Khalife, J.; Hill, E.B.; et al. Targeting leukemia stem cells in vivo with antagomiR-126 nanoparticles in acute myeloid leukemia. Leukemia 2015, 29, 2143–2153. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, P.; Su, R.; Li, Y.; Hu, C.; Wang, Y.; Arnovitz, S.; He, M.; Gurbuxani, S.; Zuo, Z.; et al. Overexpression and knockout of miR-126 both promote leukemogenesis. Blood Am. Soc. Hematol. 2015, 126, 2005–2015. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.H.; Valdez, R.; Theisen, B.K.; Guo, W.; Ferguson, D.O.; Wu, H.; Arnovitz, S.; He, M.; Gurbuxani, S.; Zuo, Z.; et al. Pten dependence distinguishes haematopoietic stem cells from leukaemia-initiating cells. Nature 2006, 441, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Krützfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of microRNAs in vivo with “antagomirs”. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Esau, C.C. Inhibition of microRNA with antisense oligonucleotides. Methods 2008, 44, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Vester, B.; Wengel, J. LNA (locked nucleic acid): High-affinity targeting of complementary RNA and DNA. Biochemistry 2004, 43, 13233–13241. [Google Scholar] [CrossRef] [PubMed]
- Elmén, J.; Lindow, M.; Schütz, S.; Lawrence, M.; Petri, A.; Obad, S.; Lindholm, M.; Hedtjärn, M.; Hansen, H.F.; Berger, U.; et al. LNA-mediated microRNA silencing in non-human primates. Nature 2008, 452, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.; Wengel, J. LNA: A versatile tool for therapeutics and genomics. Trends Biotechnol. 2003, 21, 74–81. [Google Scholar] [CrossRef]
- Bernardo, B.C.; Gao, X.-M.; Winbanks, C.E.; Boey, E.J.H.; Tham, Y.K.; Kiriazis, H.; Gregorevic, P.; Obad, S.; Kauppinen, S.; Du, X.-J.; et al. Therapeutic inhibition of the miR-34 family attenuates pathological cardiac remodeling and improves heart function. Proc. Natl. Acad. Sci. USA 2012, 109, 17615–17620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Roccaro, A.M.; Rombaoa, C.; Flores, L.; Obad, S.; Fernandes, S.M.; Sacco, A.; Liu, Y.; Ngo, H.; Quang, P.; et al. LNA-mediated anti-miR-155 silencing in low-grade B-cell lymphomas. Blood 2012, 120, 1678–1686. [Google Scholar] [CrossRef] [PubMed]
- Janssen, H.L.A.; Reesink, H.W.; Lawitz, E.J.; Zeuzem, S.; Rodriguez-Torres, M.; Patel, K.; van der Meer, A.J.; Patick, A.K.; Chen, A.; Zhou, Y.; et al. Treatment of HCV infection by targeting microRNA. N. Engl. J. Med. 2013, 368, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Bader, A.G.; Brown, D.; Winkler, M. The promise of microRNA replacement therapy. Cancer Res. 2010, 70, 7027–7030. [Google Scholar] [CrossRef] [PubMed]
- Trang, P.; Medina, P.P.; Wiggins, J.F.; Ruffino, L.; Kelnar, K.; Omotola, M.; Homer, R.; Brown, D.; Bader, A.G.; Weidhaas, J.B.; et al. Regression of murine lung tumors by the let-7 microRNA. Oncogene 2010, 29, 1580–1587. [Google Scholar] [CrossRef] [PubMed]
- Kota, J.; Chivukula, R.R.; O’Donnell, K.A.; Wentzel, E.A.; Montgomery, C.L.; Hwang, H.-W.; Chang, T.-C.; Vivekanandan, P.; Torbenson, M.; Clark, K.R.; et al. Therapeutic microRNA delivery suppresses tumorigenesis in a murine liver cancer model. Cell 2009, 137, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Fayyaz, S.; Shatynska-Mytsyk, I.; Javed, Z.; Jabeen, S.; Yaylim, I.; Javed, Z.; Jabeen, S.; Yaylim, I.; Gasparri, M.L.; et al. Is miR-34a a Well-equipped Swordsman to Conquer Temple of Molecular Oncology? Chem. Biol. Drug Des. 2016, 87, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Blower, P.E.; Chung, J.-H.; Verducci, J.S.; Lin, S.; Park, J.-K.; Dai, Z.; Liu, C.-G.; Schmittgen, T.D.; Reinhold, W.C.; Croce, C.M.; et al. MicroRNAs modulate the chemosensitivity of tumor cells. Mol. Cancer Ther. 2008, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Sarmiento, C.; Tan, T.; Zhu, H. Regulation of multidrug resistance by microRNAs in anti-cancer therapy. Acta Pharm. Sin. B 2017, 7, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Gemeinhart, R.A. Progress in microRNA delivery. J. Control. Release 2013, 172, 962–974. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, F.; Minakuchi, Y.; Nagahara, S.; Honma, K.; Sasaki, H.; Hirai, K.; Teratani, T.; Namatame, N.; Yamamoto, Y.; Hanai, K.; et al. Efficient delivery of small interfering RNA to bone-metastatic tumors by using atelocollagen in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 12177–12182. [Google Scholar] [CrossRef] [PubMed]
- Pramanik, D.; Campbell, N.R.; Karikari, C.; Chivukula, R.; Kent, O.A.; Mendell, J.T.; Maitra, A. Restitution of tumor suppressor microRNAs using a systemic nanovector inhibits pancreatic cancer growth in mice. Mol. Cancer Ther. 2011, 10, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, S.; Yu, B.; Golan, S.; Koh, C.-G.; Yang, J.; Huynh, L.; Yang, X.; Pang, J.; Muthusamy, N.; et al. Targeted Delivery of Antisense Oligodeoxynucleotide by Transferrin Conjugated pH-Sensitive Lipopolyplex Nanoparticles: A Novel Oligonucleotide-Based Therapeutic Strategy in Acute Myeloid Leukemia. Mol. Pharm. 2010, 7, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Velu, C.S.; Chaubey, A.; Phelan, J.D.; Horman, S.R.; Wunderlich, M.; Guzman, M.L.; Jegga, A.G.; Zeleznik-Le, N.J.; Chen, J.; Mulloy, J.C.; et al. Therapeutic antagonists of microRNAs deplete leukemia-initiating cell activity. J. Clin. Investig. 2014, 124, 222–236. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Schwind, S.; Santhanam, R.; Eisfeld, A.K; Chiang, C.L.; Lankenau, M.; Yu, B.; Hoellerbauer, P.; Jin, Y.; Tarighat, S.S.; et al. Targeting the RAS/MAPK pathway with miR-181a in acute myeloid leukemia. Oncotarget 2016, 7, 59273–59286. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Role of LSCs in relapse development. At diagnosis, AML consist of a heterogeneous population of leukemic (stem) cells and residual normal hematopoietic (stem) cells. (A) Treatment with chemotherapy often results in complete remission. However, small numbers of leukemic cells survive the treatment (minimal residual disease, MRD). MRD contains chemotherapy resistant LSCs, which have the capacity to re-initiate leukemia and form a relapse. (B) MicroRNA-based therapy in combination with chemotherapy could eradicate LSCs and leukemic progenitors (LP) while sparing, or stimulating, HSCs.
Figure 1.
Role of LSCs in relapse development. At diagnosis, AML consist of a heterogeneous population of leukemic (stem) cells and residual normal hematopoietic (stem) cells. (A) Treatment with chemotherapy often results in complete remission. However, small numbers of leukemic cells survive the treatment (minimal residual disease, MRD). MRD contains chemotherapy resistant LSCs, which have the capacity to re-initiate leukemia and form a relapse. (B) MicroRNA-based therapy in combination with chemotherapy could eradicate LSCs and leukemic progenitors (LP) while sparing, or stimulating, HSCs.
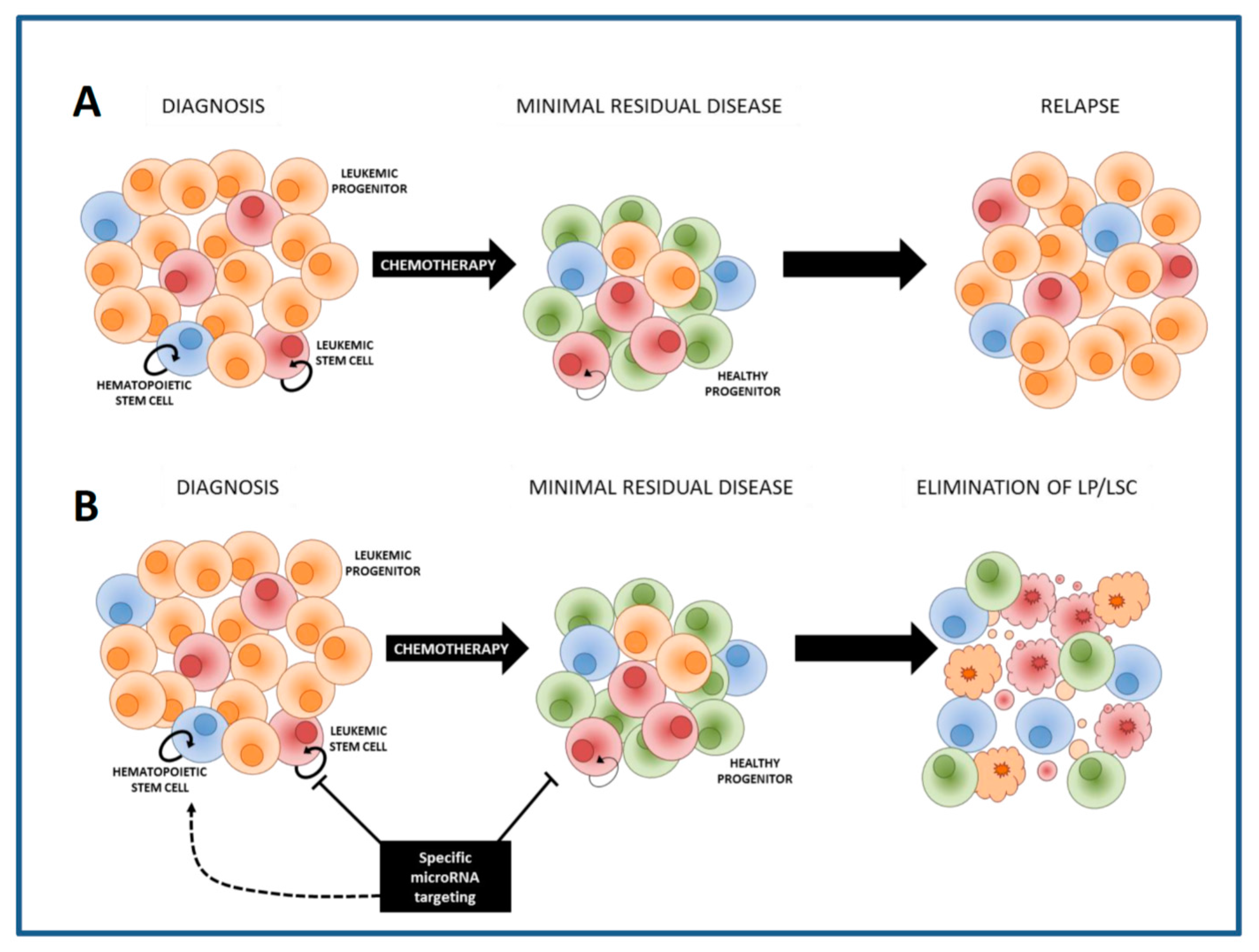
Figure 2.
MicroRNA biogenesis and function. Primary miRNAs (pri-miRNA) are transcribed from miRNA genes by RNA polymerase II (RNA-Pol II). In the nucleus, the endonuclease Drosha together with a double-stranded RNA (dsRNA)-binding protein (DGCR8) cleave the stem–loop structure of the pri-miRNA. This results in a precursor miRNA (pre-miRNA) which is exported from the nucleus by exportin 5 (XPO5). In the cytoplasm, the pre-miRNA is then further cleaved by the endonuclease Dicer leading to a miRNA–miRNA* duplex. This duplex is loaded into an Argonaute (AGO) protein. The mature RNA-induced-silencing complex (RISC) is formed when miRNA*-strand is expelled from the AGO protein. The RISC complex can inhibit initiation of translation by affecting recruitment of 40S small ribosomal subunit and/or by inhibiting the 60S subunit. Alternatively, RISC may obstruct translation by inhibiting the elongation of ribosomes. RISC binding can also lead to recruitment of RNA decapping and/or deadenylating enzymes leading to mRNA destabilization. Some of the target mRNAs bound by the RISC are transported into cytoplasmic processing bodies (P-bodies) for degradation or storage.
Figure 2.
MicroRNA biogenesis and function. Primary miRNAs (pri-miRNA) are transcribed from miRNA genes by RNA polymerase II (RNA-Pol II). In the nucleus, the endonuclease Drosha together with a double-stranded RNA (dsRNA)-binding protein (DGCR8) cleave the stem–loop structure of the pri-miRNA. This results in a precursor miRNA (pre-miRNA) which is exported from the nucleus by exportin 5 (XPO5). In the cytoplasm, the pre-miRNA is then further cleaved by the endonuclease Dicer leading to a miRNA–miRNA* duplex. This duplex is loaded into an Argonaute (AGO) protein. The mature RNA-induced-silencing complex (RISC) is formed when miRNA*-strand is expelled from the AGO protein. The RISC complex can inhibit initiation of translation by affecting recruitment of 40S small ribosomal subunit and/or by inhibiting the 60S subunit. Alternatively, RISC may obstruct translation by inhibiting the elongation of ribosomes. RISC binding can also lead to recruitment of RNA decapping and/or deadenylating enzymes leading to mRNA destabilization. Some of the target mRNAs bound by the RISC are transported into cytoplasmic processing bodies (P-bodies) for degradation or storage.
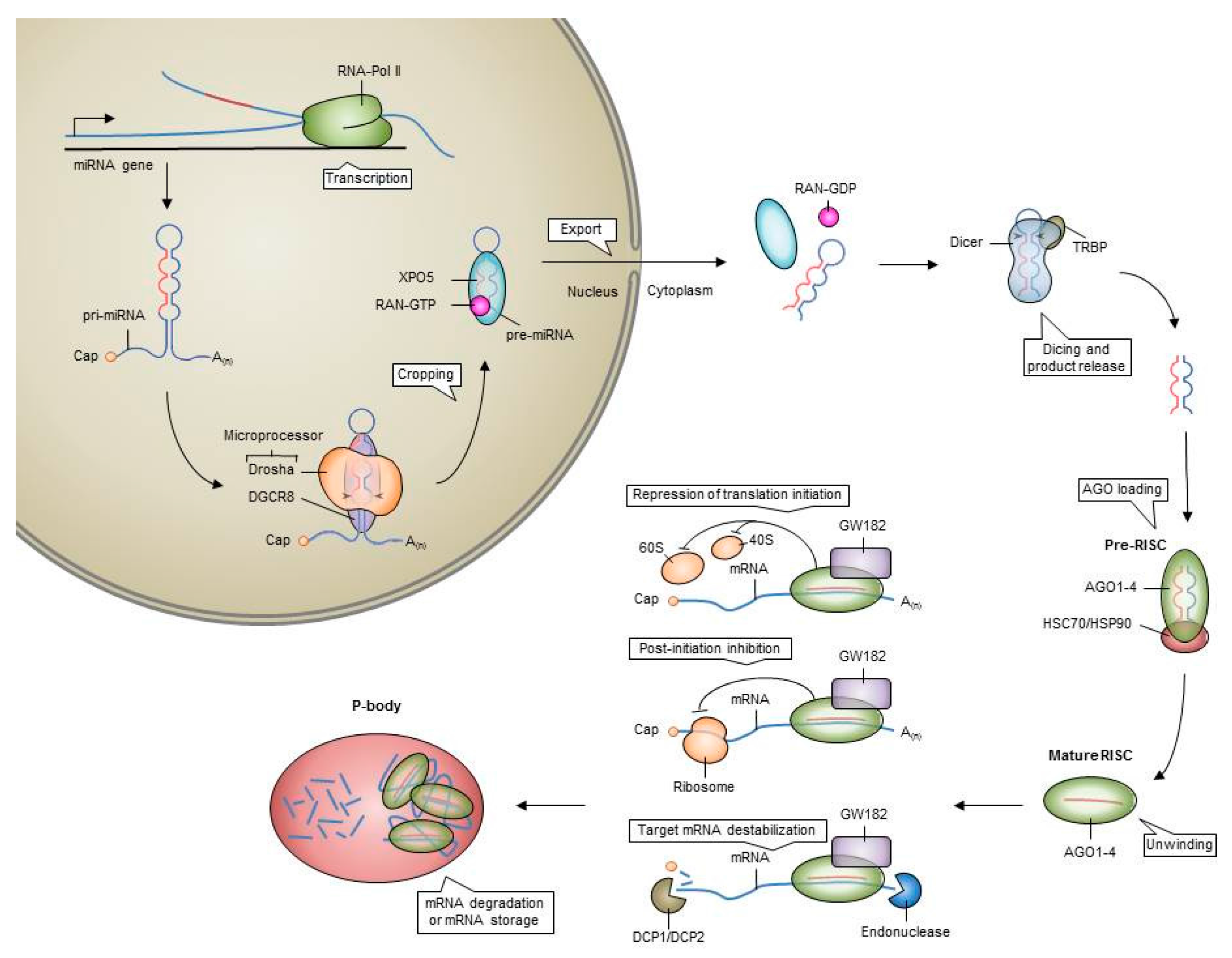
Figure 3.
Differential expressed miRNAs between normal and leukemic stem cells and leukemic stem and progenitor cells obtained from AML bone marrow.
Figure 3.
Differential expressed miRNAs between normal and leukemic stem cells and leukemic stem and progenitor cells obtained from AML bone marrow.
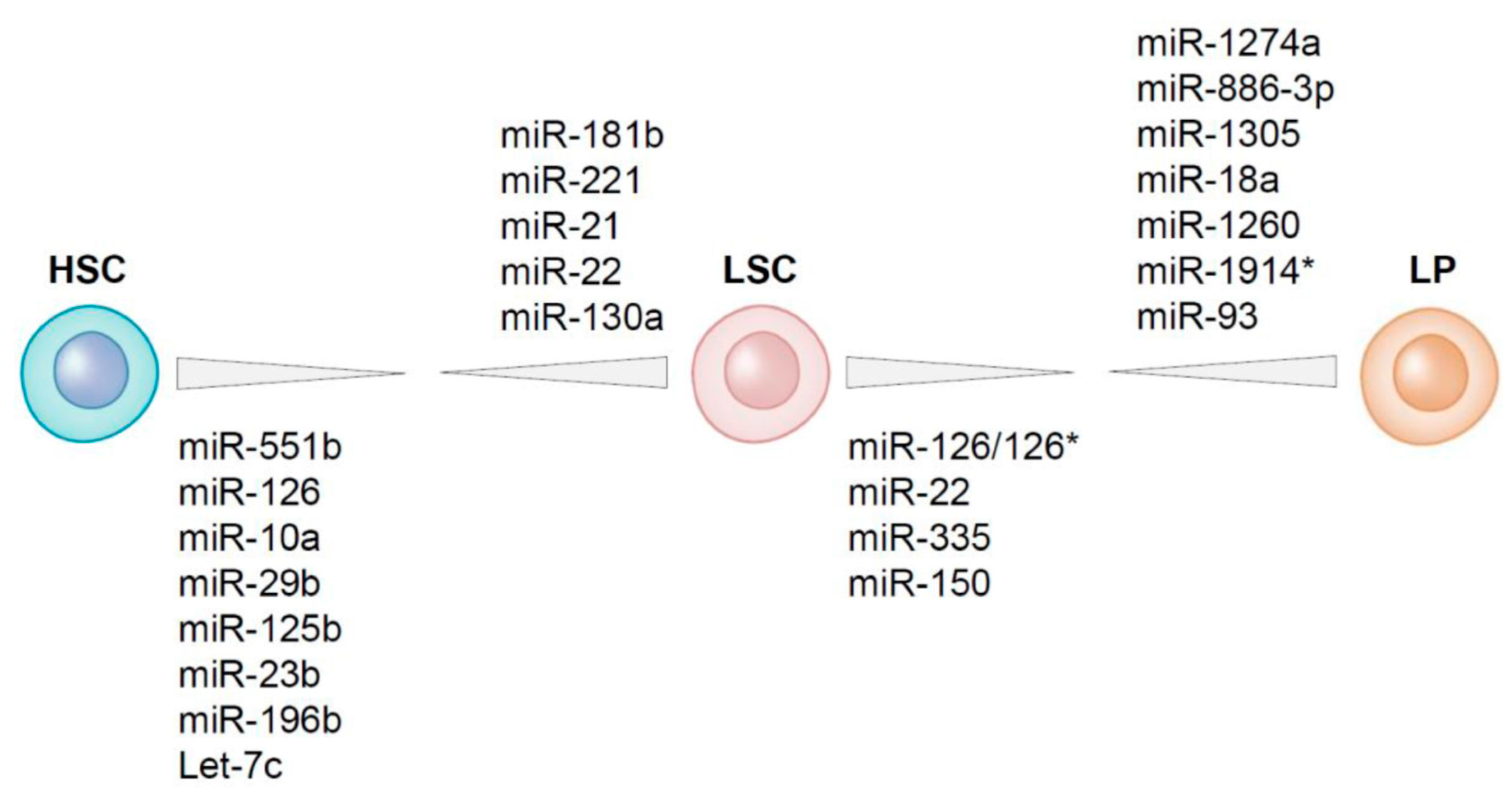
Figure 4.
MiR-126 expression and modulation in normal hematopoiesis and leukemia. MiR-126 is highly expressed in normal HSCs. A reduction in miR-126 expression in HSCs, for example by lentiviral sponges, increases AKT signaling thereby inducing cell-cycle entry via CDK3 that leads to enhanced self-renewal and expansion of long-term HSCs. Overexpression of miR-126 in HSCs, by mimics or internal and external stimuli (cytokines), results in a reduction in AKT signaling impairing cell-cycle entry resulting in increased quiescence and a gradual loss of hematopoietic output. In leukemia, LSCs highly express miR-126 as compared to leukemic progenitors. Knockdown of miR-126 in LSCs increases AKT signaling, de-repressing CKD3, thereby inducing differentiation and proliferation leading to chemo-sensitivity and apoptosis. Overexpression of miR-126 in LSCs lowers AKT signaling and inhibits cell cycle entry leading to increased quiescence and self-renewal via the suppression of CDK3 resulting in chemotherapy resistance.
Figure 4.
MiR-126 expression and modulation in normal hematopoiesis and leukemia. MiR-126 is highly expressed in normal HSCs. A reduction in miR-126 expression in HSCs, for example by lentiviral sponges, increases AKT signaling thereby inducing cell-cycle entry via CDK3 that leads to enhanced self-renewal and expansion of long-term HSCs. Overexpression of miR-126 in HSCs, by mimics or internal and external stimuli (cytokines), results in a reduction in AKT signaling impairing cell-cycle entry resulting in increased quiescence and a gradual loss of hematopoietic output. In leukemia, LSCs highly express miR-126 as compared to leukemic progenitors. Knockdown of miR-126 in LSCs increases AKT signaling, de-repressing CKD3, thereby inducing differentiation and proliferation leading to chemo-sensitivity and apoptosis. Overexpression of miR-126 in LSCs lowers AKT signaling and inhibits cell cycle entry leading to increased quiescence and self-renewal via the suppression of CDK3 resulting in chemotherapy resistance.
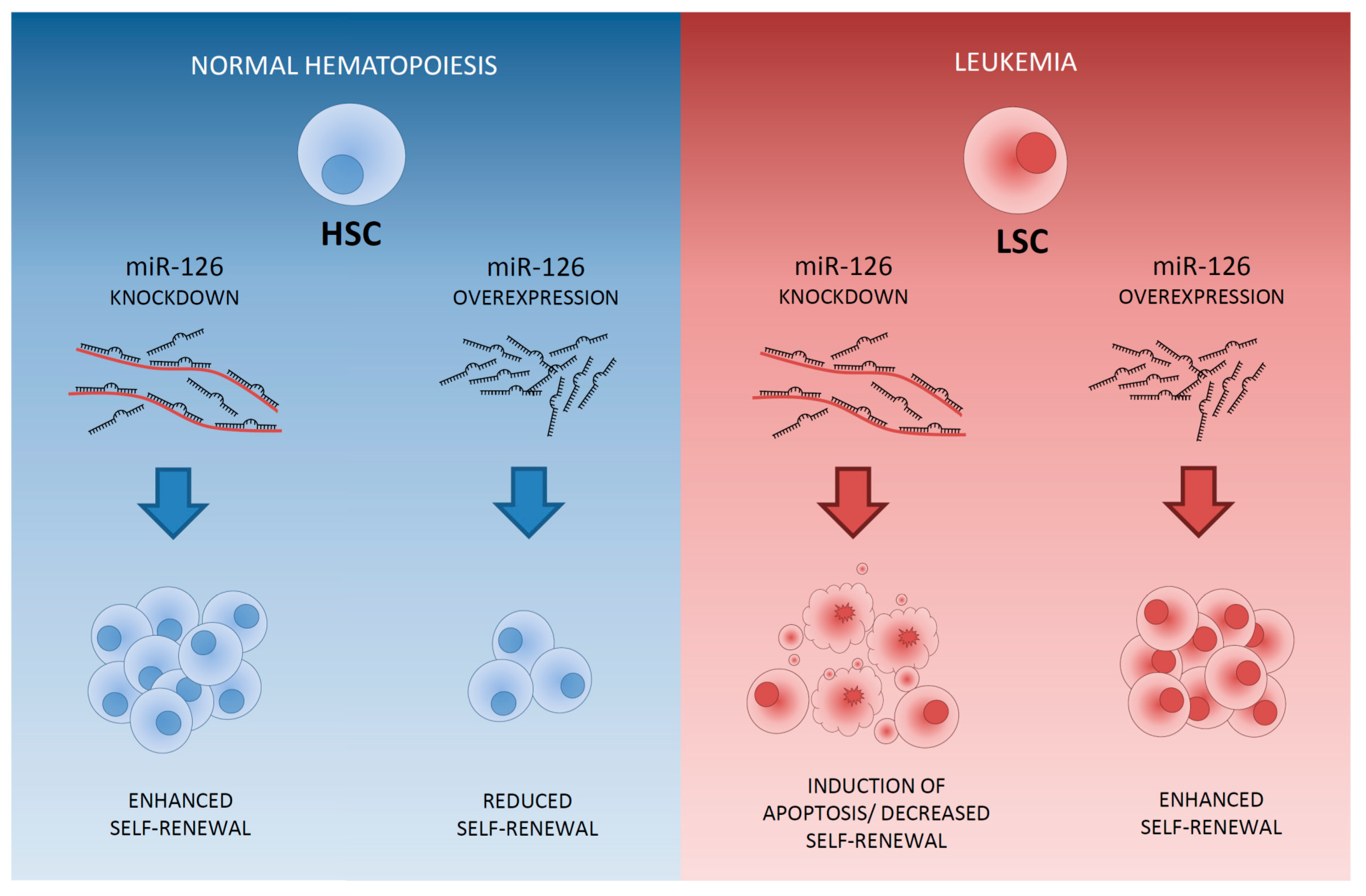

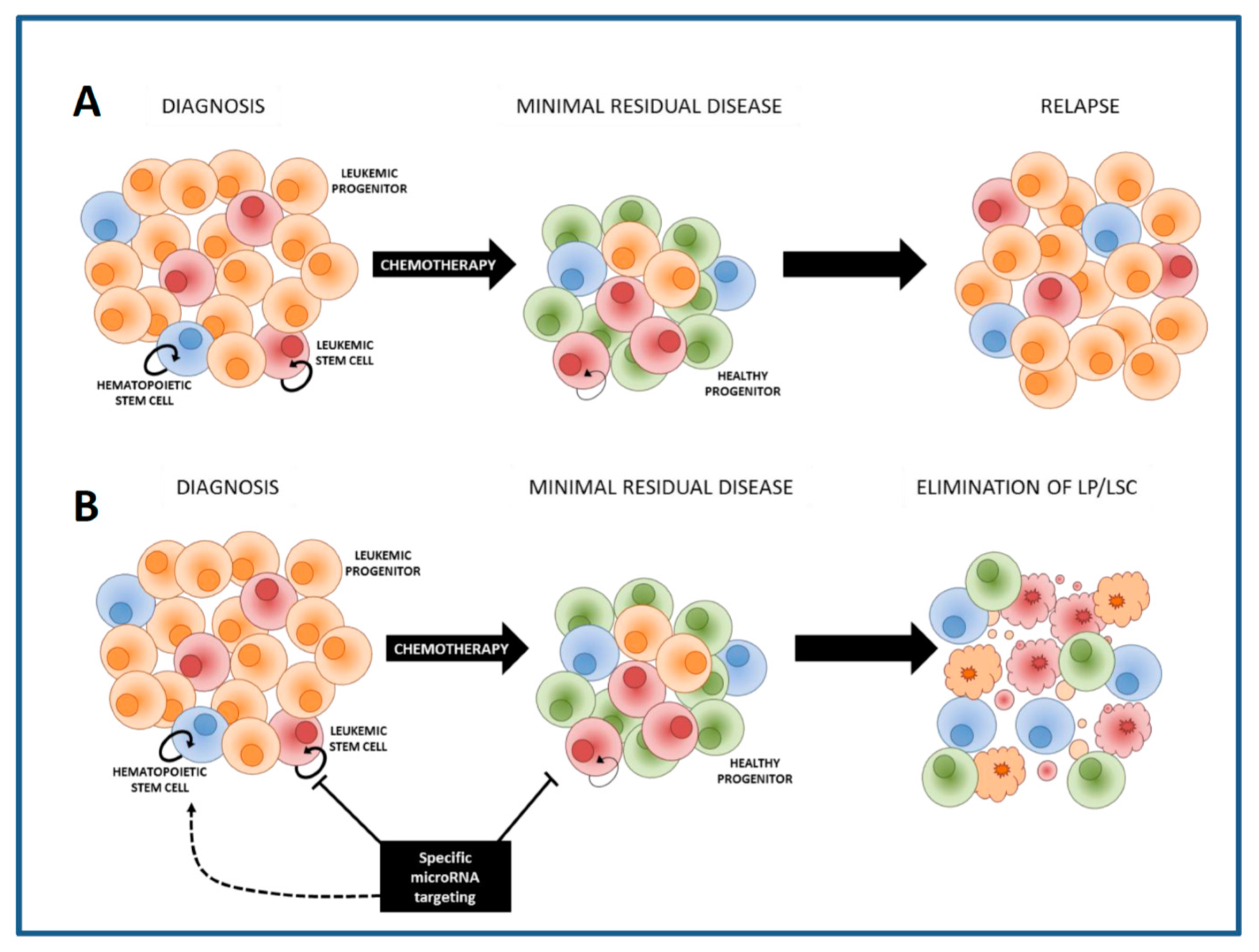{kind=link}
{kind=link}
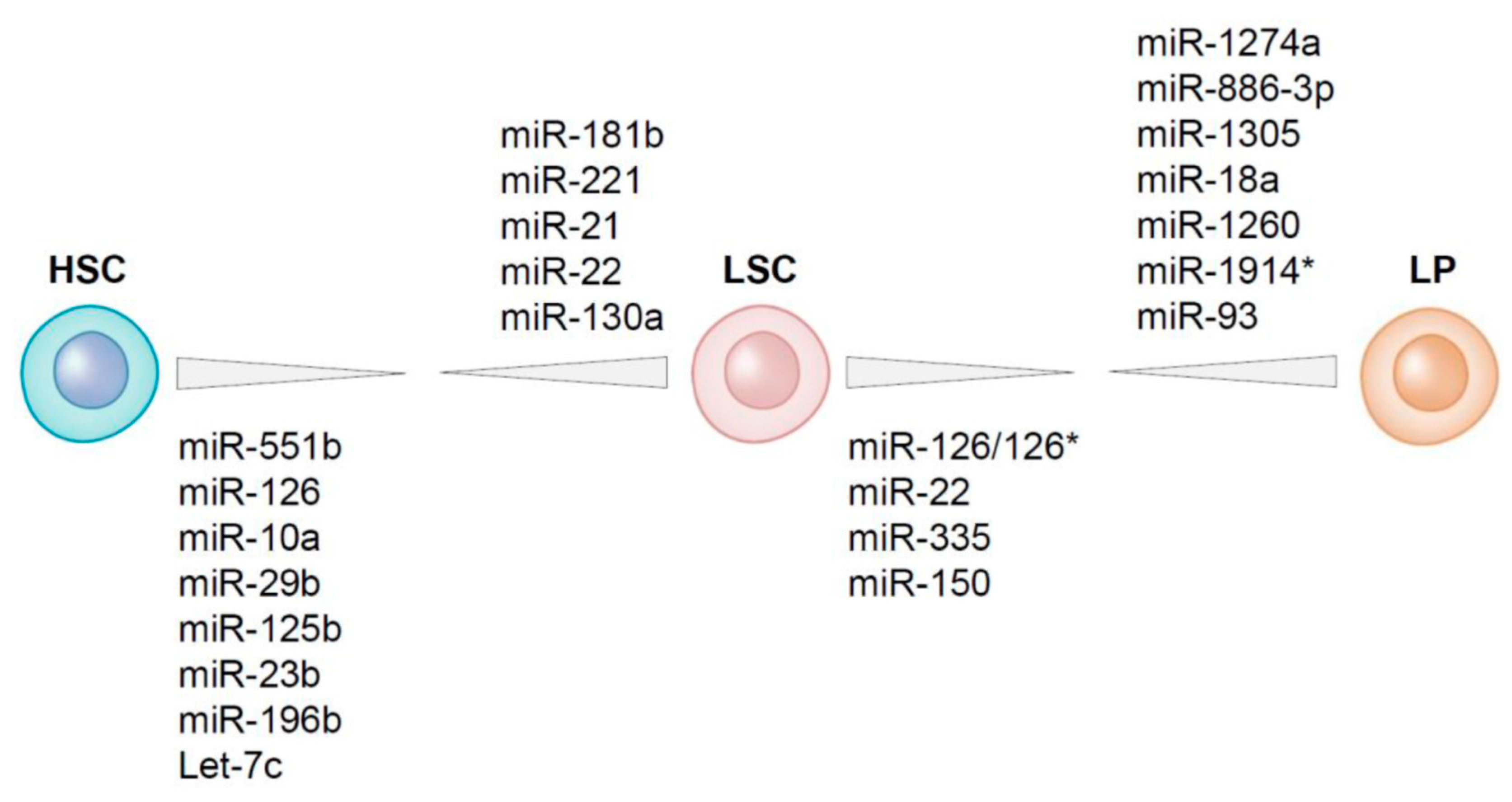{kind=link}
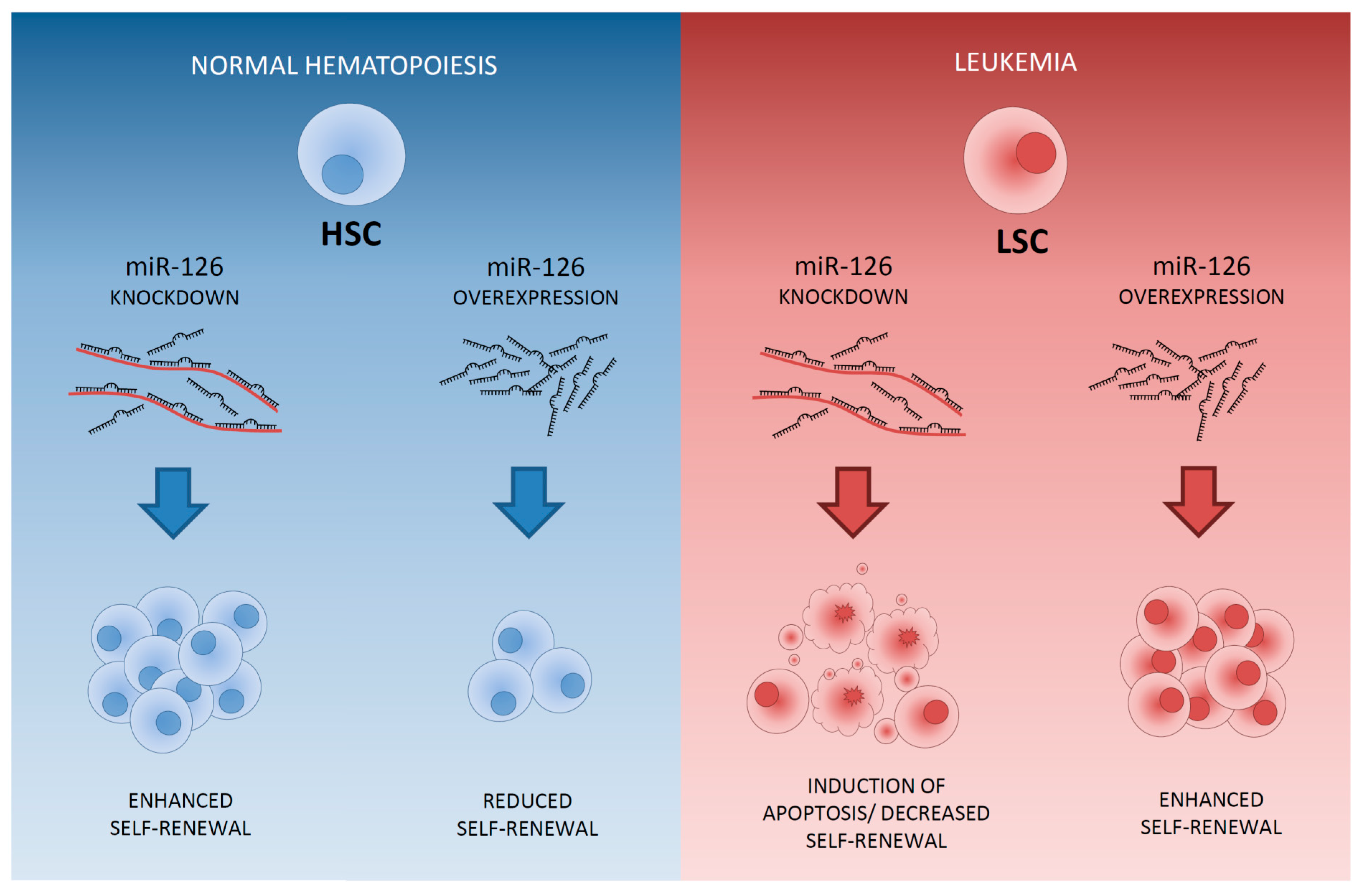{kind=link}
Table 1.
miRNAs involved in myelopoiesis which are involved in key (stem) cell processes (differentiation, self-renewal, apoptosis and proliferation) and their identified targets.
Table 1.
miRNAs involved in myelopoiesis which are involved in key (stem) cell processes (differentiation, self-renewal, apoptosis and proliferation) and their identified targets.
| Cell Stage | microRNA | Target | Function | References | |
|---|---|---|---|---|---|
| HSC | Let-7 | Hmg2a |  | self-renewal | [61] |
| miR-12 | Tip110 |  | differentiation | [62] | |
| miR-125a | BAK1 | | apoptosis | [30,31,58] | |
| miR-125b | ABTB1/CDC25C/PPP1CA |  | proliferation | [63,64] | |
| Bmf/KLF13/p53 | | apoptosis | [56,58,65] | ||
| STAT3/c-JUN/JUND/LIN28A/CBFB | | differentiation | [63,66,67,68,69,70] | ||
| miR-126 | HOXA9/PI3K/AKT2/CRKII | | self-renewal | [71,72] | |
| miR-132 | FOXO3 | | proliferation | [73] | |
| miR-146a | TRAF6/IRAK1/STAT1 | | self-renewal | [74,75,76] | |
| miR-17-92 cluster | E2F1/E2F2 | | proliferation and block differentiation | [77,78] | |
| PTEN/Bim | | apoptosis | [79,80] | ||
| miR-196b | HOXA9/MEIS1/FAS/HOXB8 | | differentiation | [81,82] | |
| miR-24 | Bim/CASP9 | | apoptosis | [83] | |
| miR-29a | Dnmt3a | | self-renewal | [57] | |
| miR-33 | p53 | self-renewal | [84] | ||
| miR-22 | Tet2 | | self-renewal | [85] | |
| MPP | miR-17/20/93/106 | SQSTM1 | | differentiation towards myeloid progenitors | [54,86] |
| miR-24 | Unknown | | differentiation towards myeloid progenitors | [83] | |
| miR-29a | HBP1, FZD5, TPM1 | | differentiation towards myeloid progenitors | [55] | |
| miR-520h | ABCG2 | differentiation towards myeloid progenitors | [49] | ||
| miR-181a | Bcl2, CD69 | | differentiation towards lymphoid progenitors | [87,88,89] | |
| CMP | miR-142-3p | CCNT2/TAB2 | | granulocytic-macrophage differentiation | [90] |
| miR-155 | PU.1 | | granulocytic-macrophage differentiation | [91] | |
| miR-29a | CCNT2/CDK6 | granulocytic-macrophage differentiation | [90] | ||
| miR-130a | C/EBPɛ | | granulocytic differentiation | [92] | |
| GMP | miR-17-5p/20a/106a | RUNX1 | | monocytic differentiation and maturation | [93] |
| miR-223 | MEF2C | | progenitor proliferation and granulocyte differentiation | [94] | |
| miR-223 miR-27 | NFI-A/E2F1 | | granulocytic differentiation | [95,96,97] | |
| RUNX1 | | granulocytic differentiation | [98] | ||
| miR-30c | NOTCH1 | | granulocytic differentiation | [99] | |
| miR-34a | E2F3 | | granulocytic differentiation | [100] | |
| miR-424 | NFI-A | | monocytic differentiation | [101] | |
| miR-486-3p | MAF | Skews from monocytopoiesis towards granulopoiesis | [102] | ||
| miR-105 | MYB | megakaryopoiesis | [103] | ||
| miR-22 | PU.1, MECOM | | monocytic differentiation | [104,105] | |
| miR-181a | Unknown | | megakaryocytic differentiation | [106] | |
| Lin28, let7 | megakaryocytic differentiation | [107] | |||
| MEP | miR-125b | Unknown | | proliferation and self-renewal | [108] |
| miR-126 | MYB | Skews from erythropoiesis towards megakaryopoiesis | [109] | ||
| miR-145 | Fli-1 | Skews from megakaryopoiesis towards erythropoiesis | [110] | ||
| miR-146a | CXCR4 | Impairs megakaryocytic proliferation, differentiation and maturation | [111] | ||
| miR-15 | MYB | | erythropoiesis | [112] | |
| miR-150 | MYB | Skews from erythropoiesis towards megakaryopoiesis | [109,113,114] | ||
| miR-155 | ETS-1/MEIS1 | | megakaryocytic proliferation and differentiation | [115] | |
| miR-199b-5p | c-Kit | | erythroid differentiation | [116] | |
| miR-221/222 | c-Kit | Impairs proliferation and accelerates differentiation of erythroid cells | [32] | ||
| miR-223 | LMO2 | Skews from erythroid towards megakaryocytic differentiation | [117] | ||
| miR-23 | SHP2 | | erythroid differentiation | [118] | |
| miR-27a/24 | GATA2 | | erythroid differentiation | [119] | |
| miR-299-5p | Unknown | Skews from erythroid-monocytic towards megakaryocytic-granulocytic differentiation | [120] | ||
| miR-34a | MYB/CDK4/CDK6 | | megakaryocytic differentiation and inhibit cell cycle | [121] | |
| miR-376a | CDK2 | | erythroid differentiation | [122] | |
| miR-451/144 | GATA2 | | erythroid differentiation | [123,124,125] | |
| miR-486-3p | MAF/BCL11A | Skews from megakaryopoiesis towards erythropoiesis | [102,126] | ||
| miR-146b | PDGFRA | | erythrocytic-megakaryocytic differentiation | [127] | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Martiáñez Canales, T.; De Leeuw, D.C.; Vermue, E.; Ossenkoppele, G.J.; Smit, L. Specific Depletion of Leukemic Stem Cells: Can MicroRNAs Make the Difference? Cancers 2017, 9, 74. https://doi.org/10.3390/cancers9070074
AMA Style
Martiáñez Canales T, De Leeuw DC, Vermue E, Ossenkoppele GJ, Smit L. Specific Depletion of Leukemic Stem Cells: Can MicroRNAs Make the Difference? Cancers. 2017; 9(7):74. https://doi.org/10.3390/cancers9070074
Chicago/Turabian StyleMartiáñez Canales, Tania, David C. De Leeuw, Eline Vermue, Gert J. Ossenkoppele, and Linda Smit. 2017. "Specific Depletion of Leukemic Stem Cells: Can MicroRNAs Make the Difference?" Cancers 9, no. 7: 74. https://doi.org/10.3390/cancers9070074
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.