
Characterization of Adenomatous Polyposis Coli Protein Dynamics and Localization at the Centrosome


Abstract
:1. Introduction
2. Results
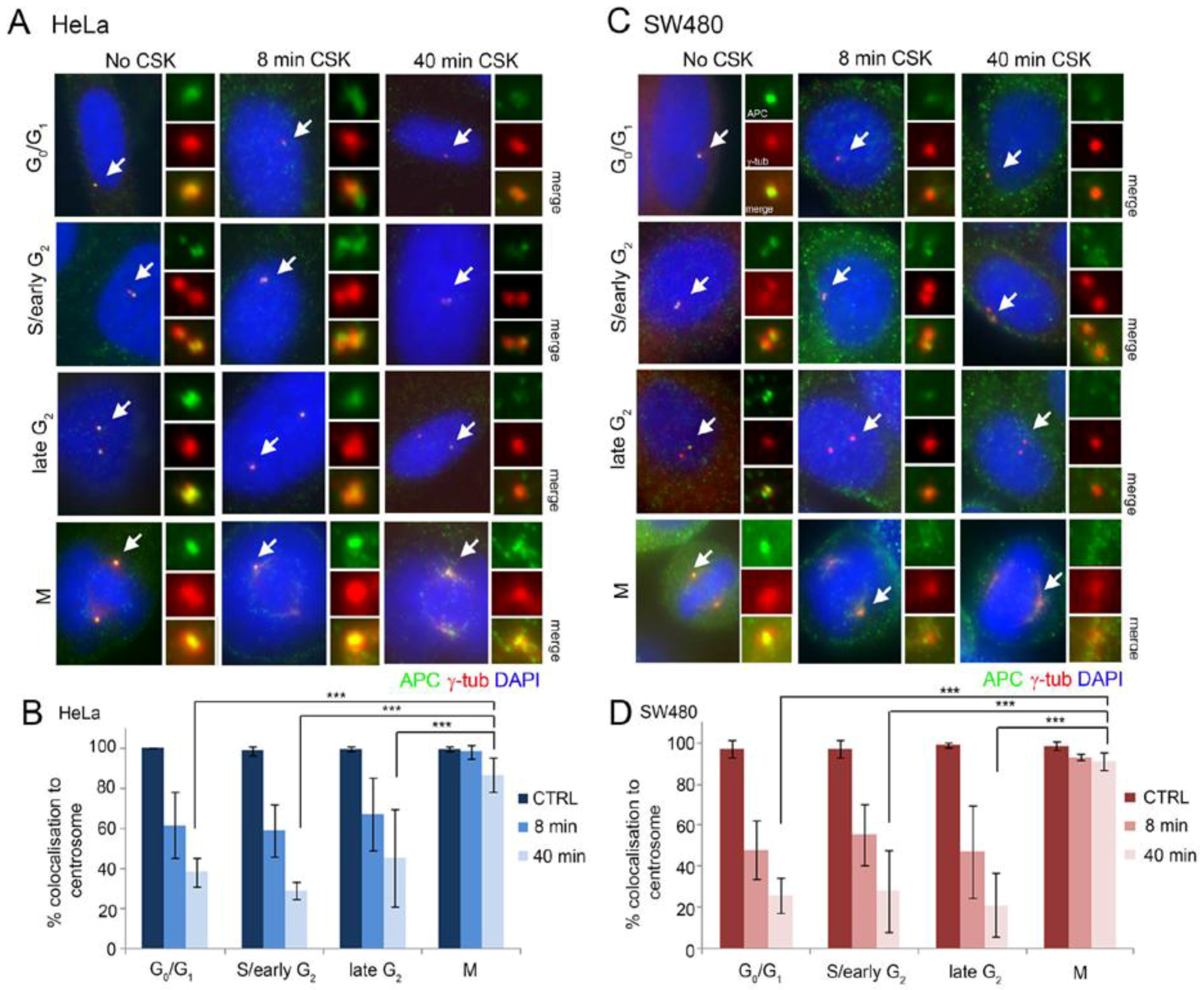
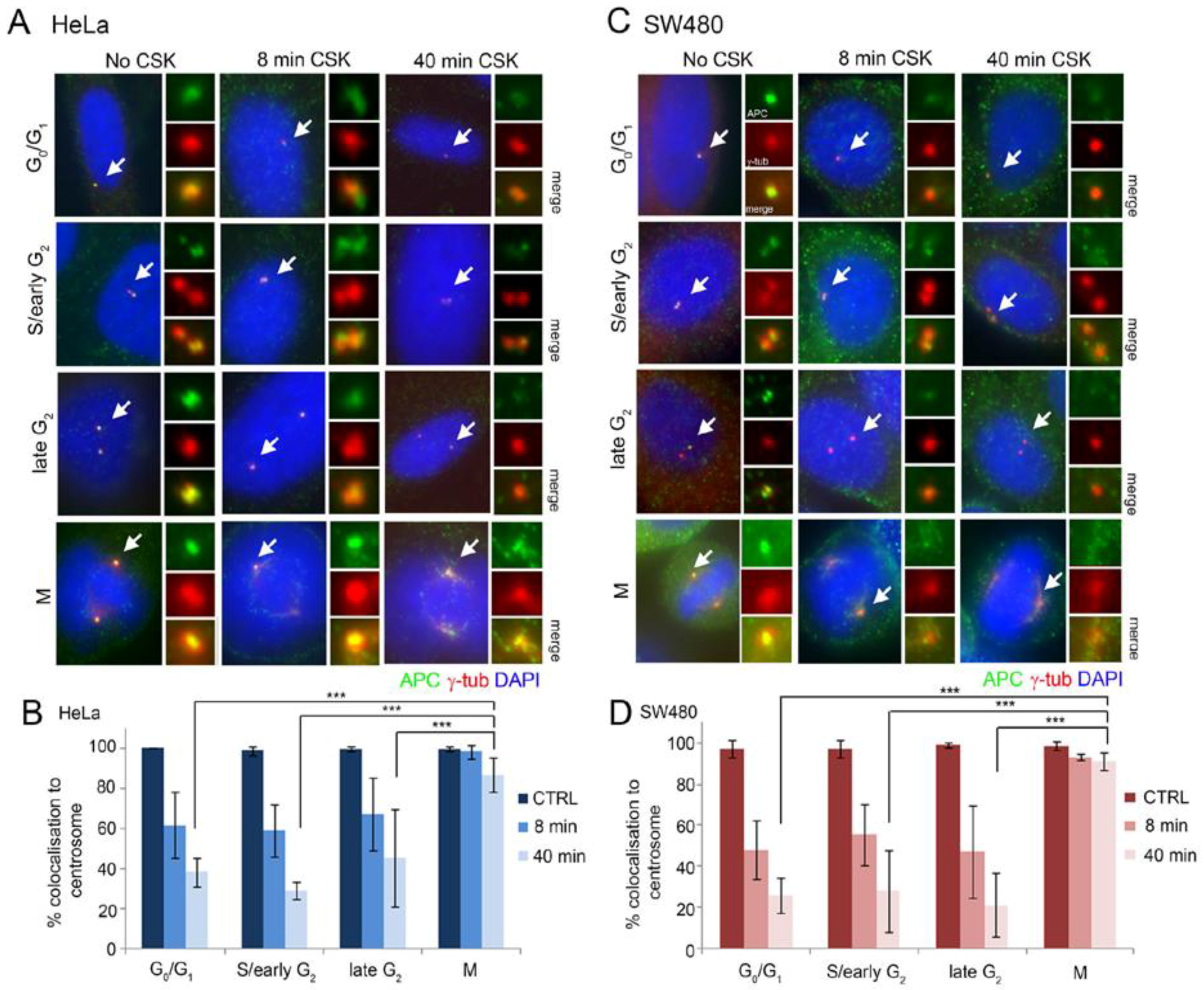
2.1. Full-Length and Truncated APC Localize to the Centrosome Throughout the Cell Cycle and Display High Dynamic Exchange during Interphase
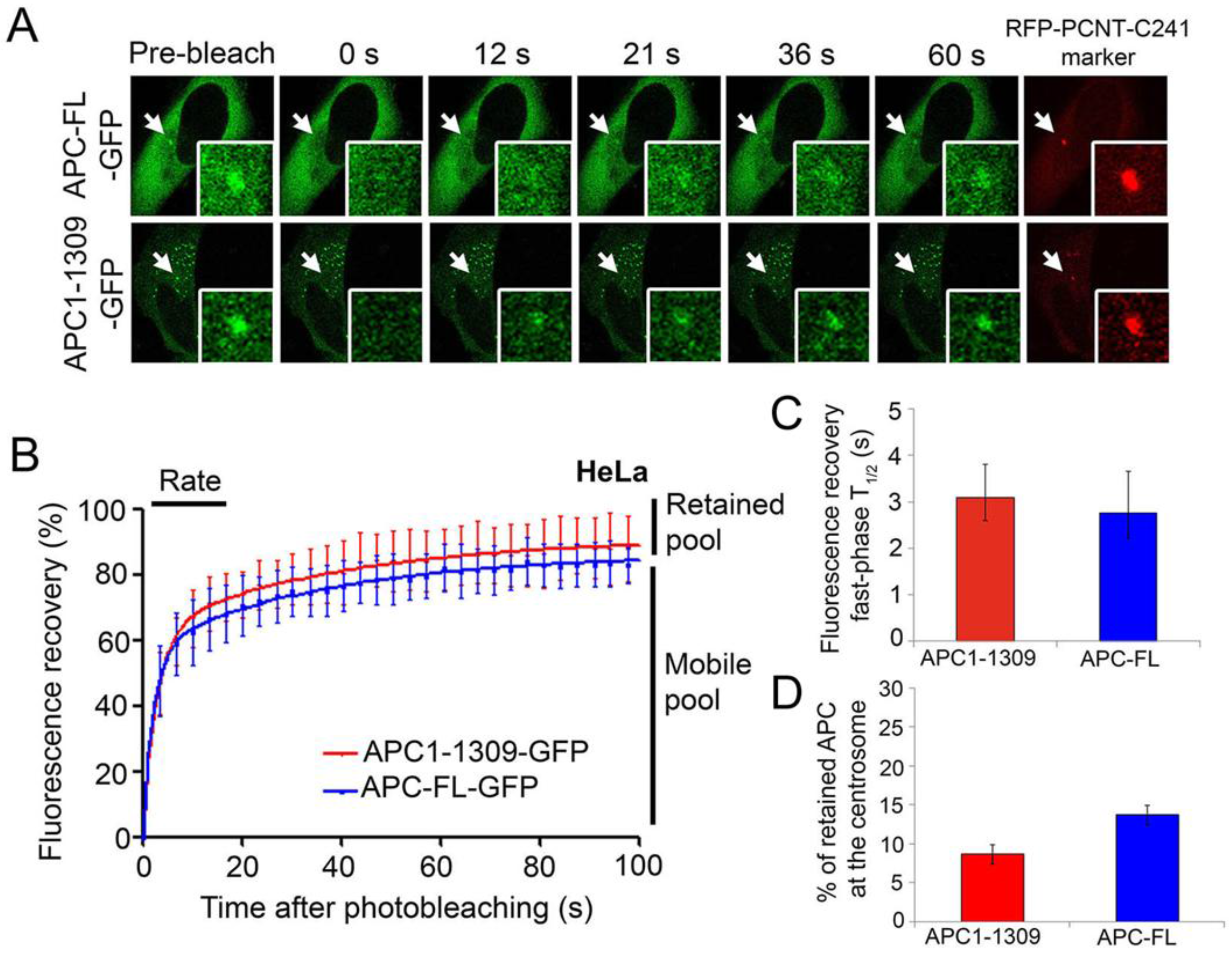
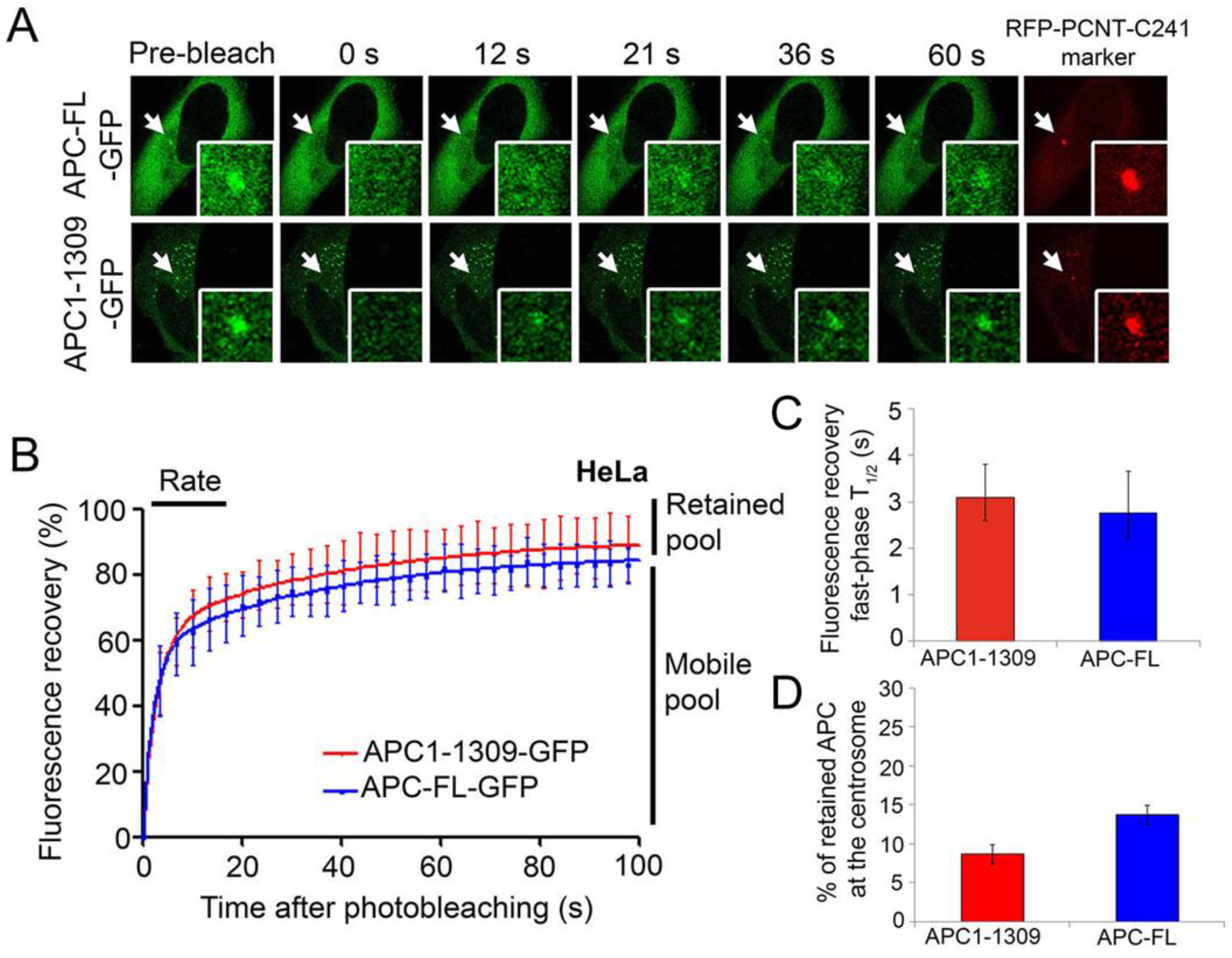
2.2. Wild-Type and Cancer Mutant APC Display Similar Dynamics at the Centrosome: Identification of Mobile and Immobile Pools of APC
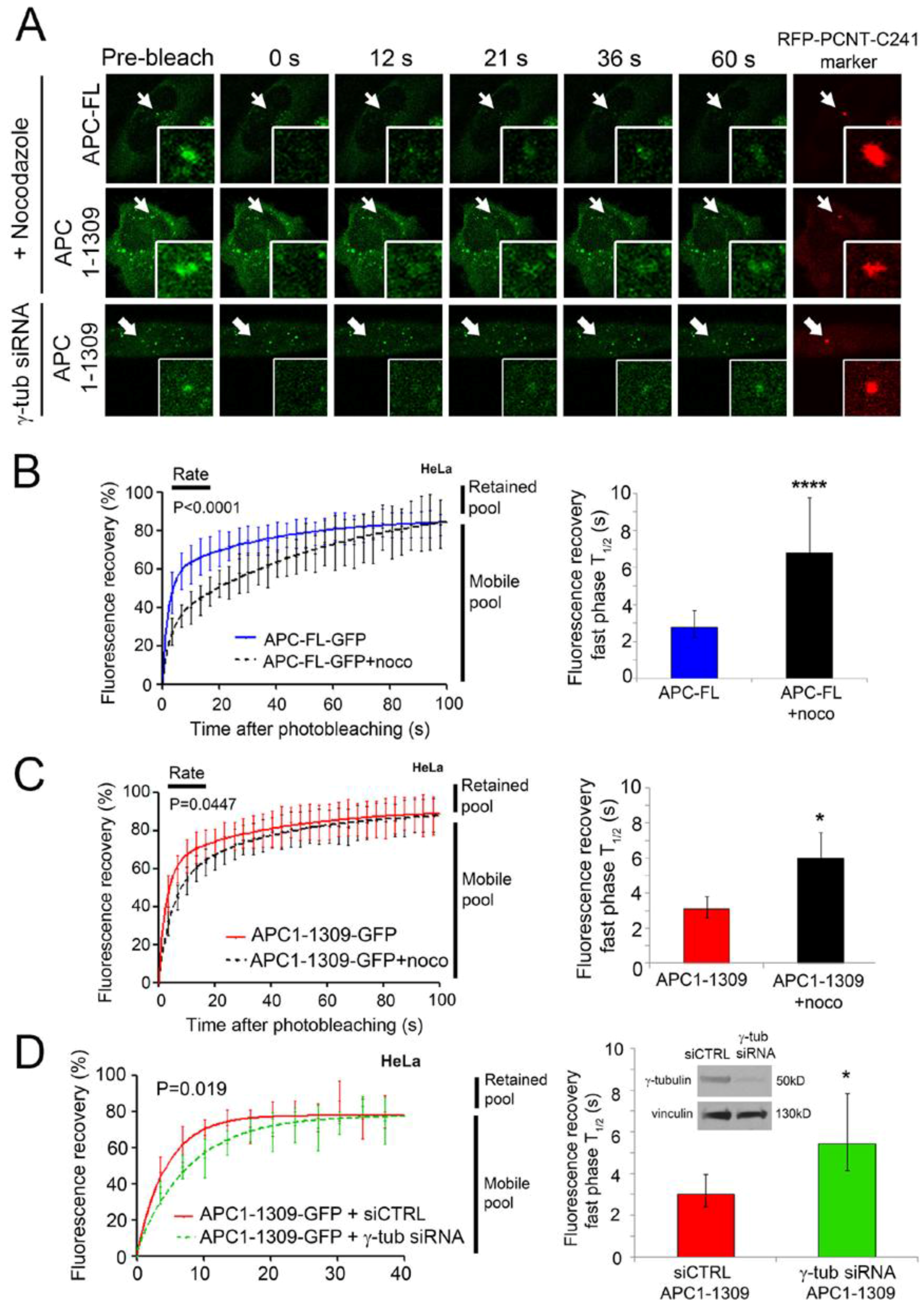
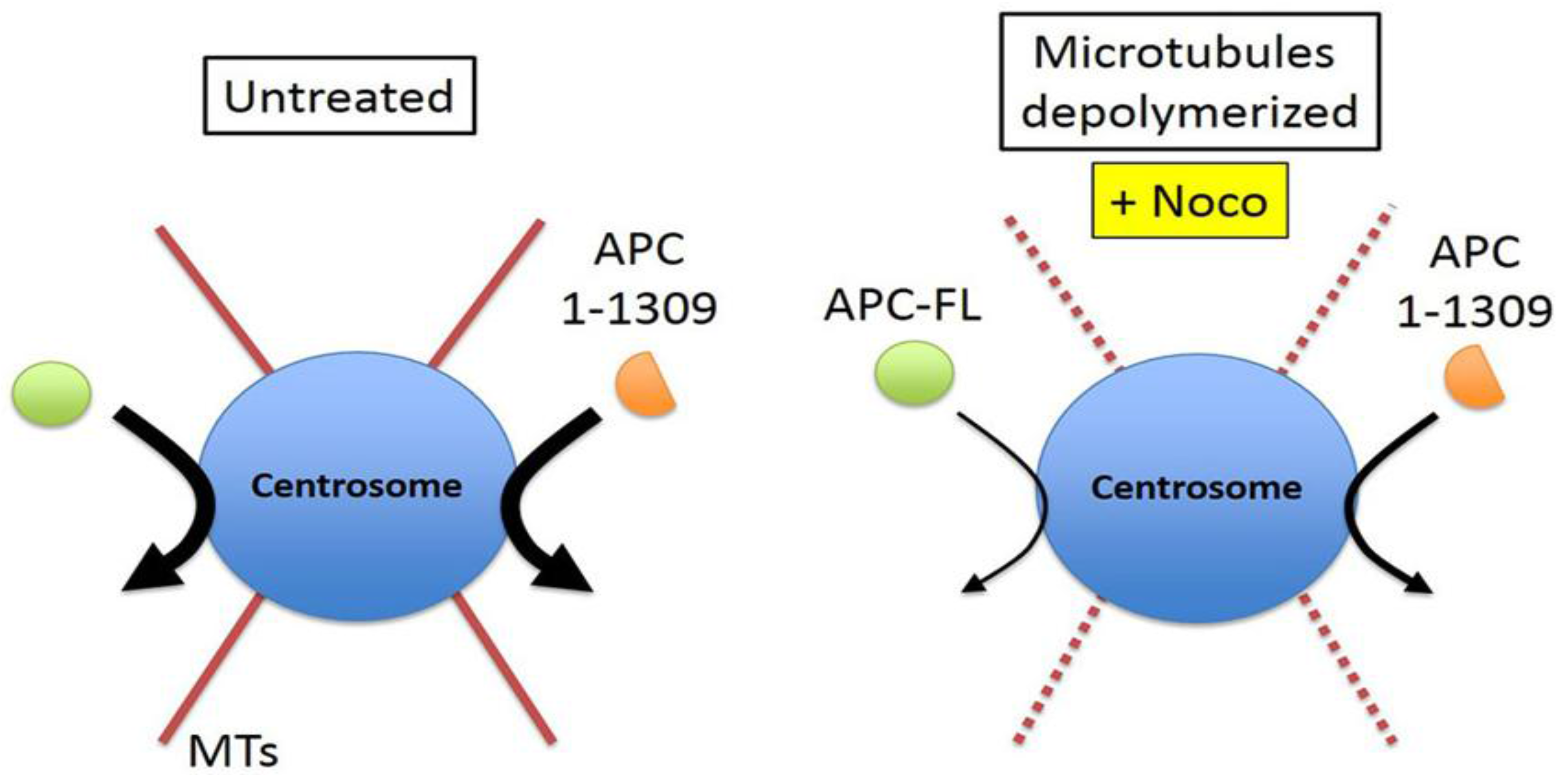
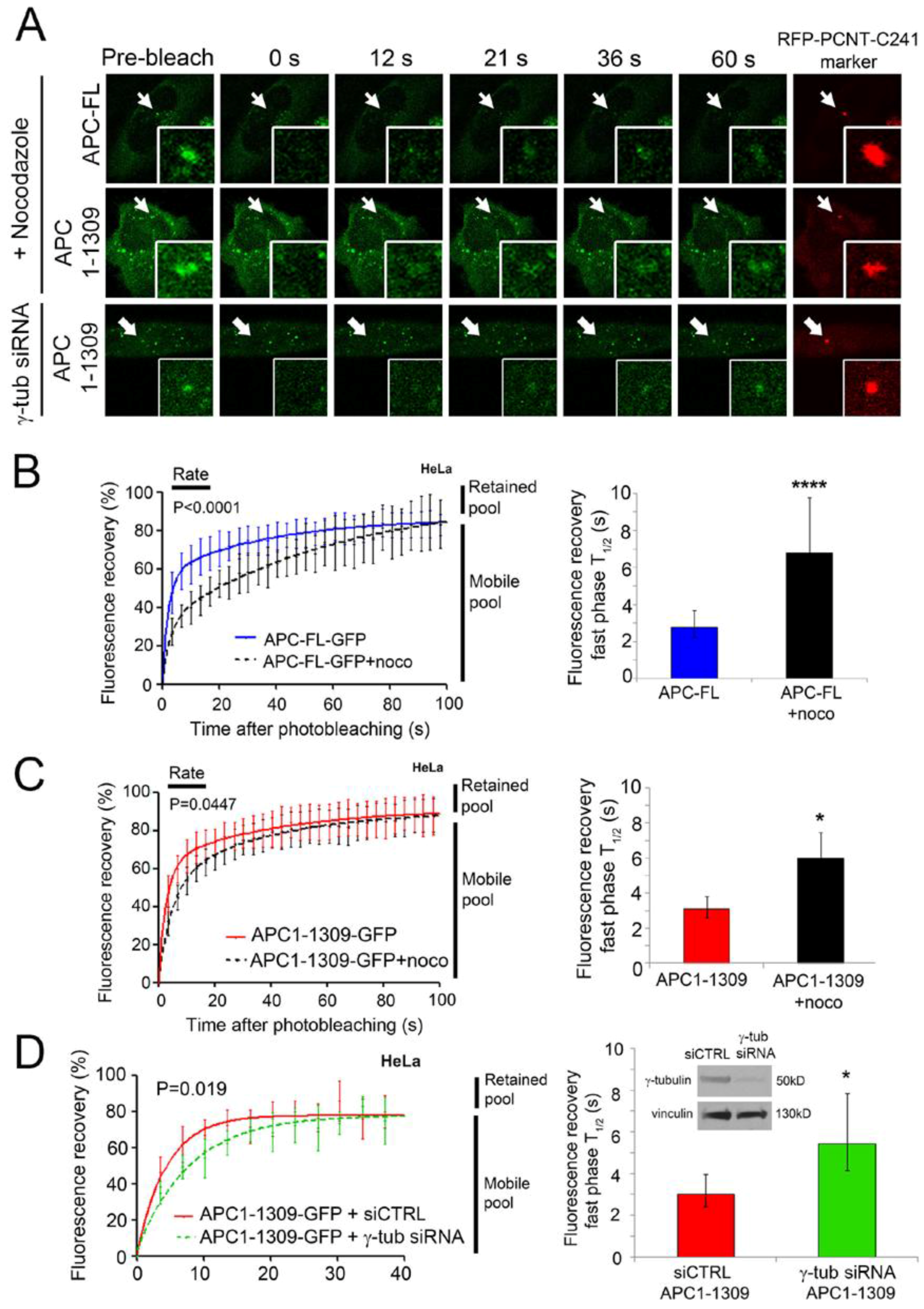
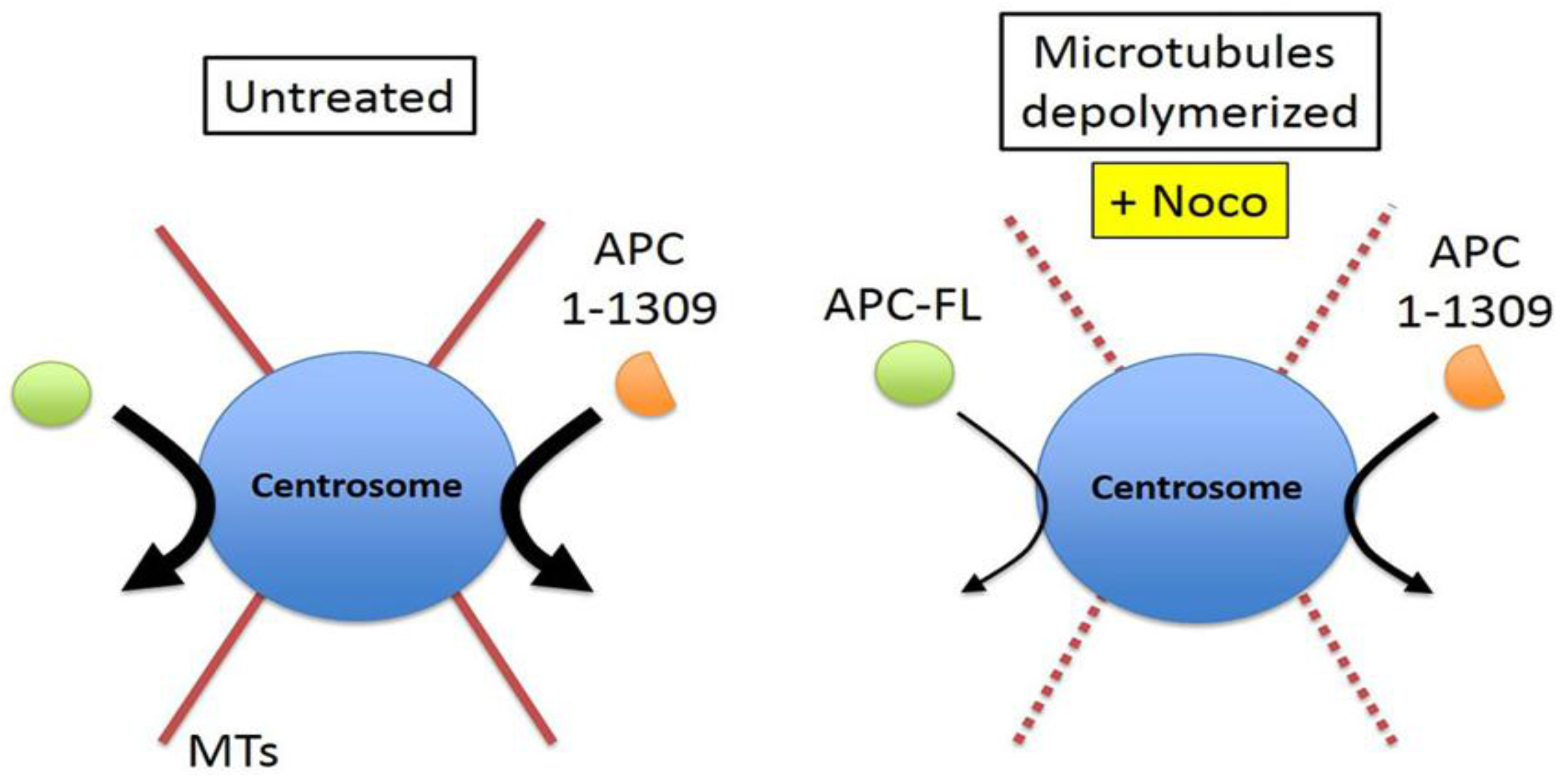
2.3. Intact Microtubules Stimulate Transport of APC to the Interphase Centrosome
2.4. γ-Tubulin Stimulates the Rate of Recruitment of APC to the Centrosome
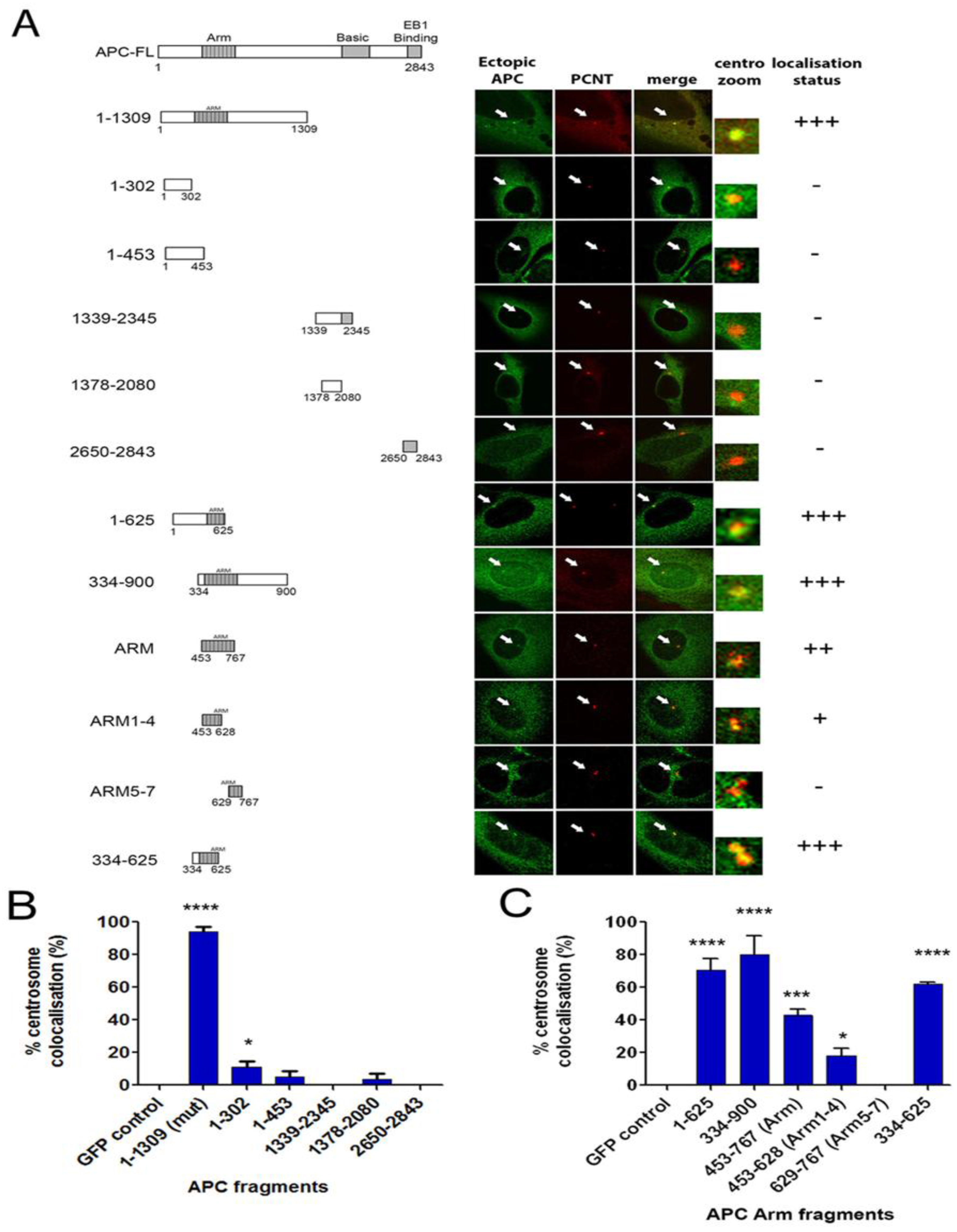
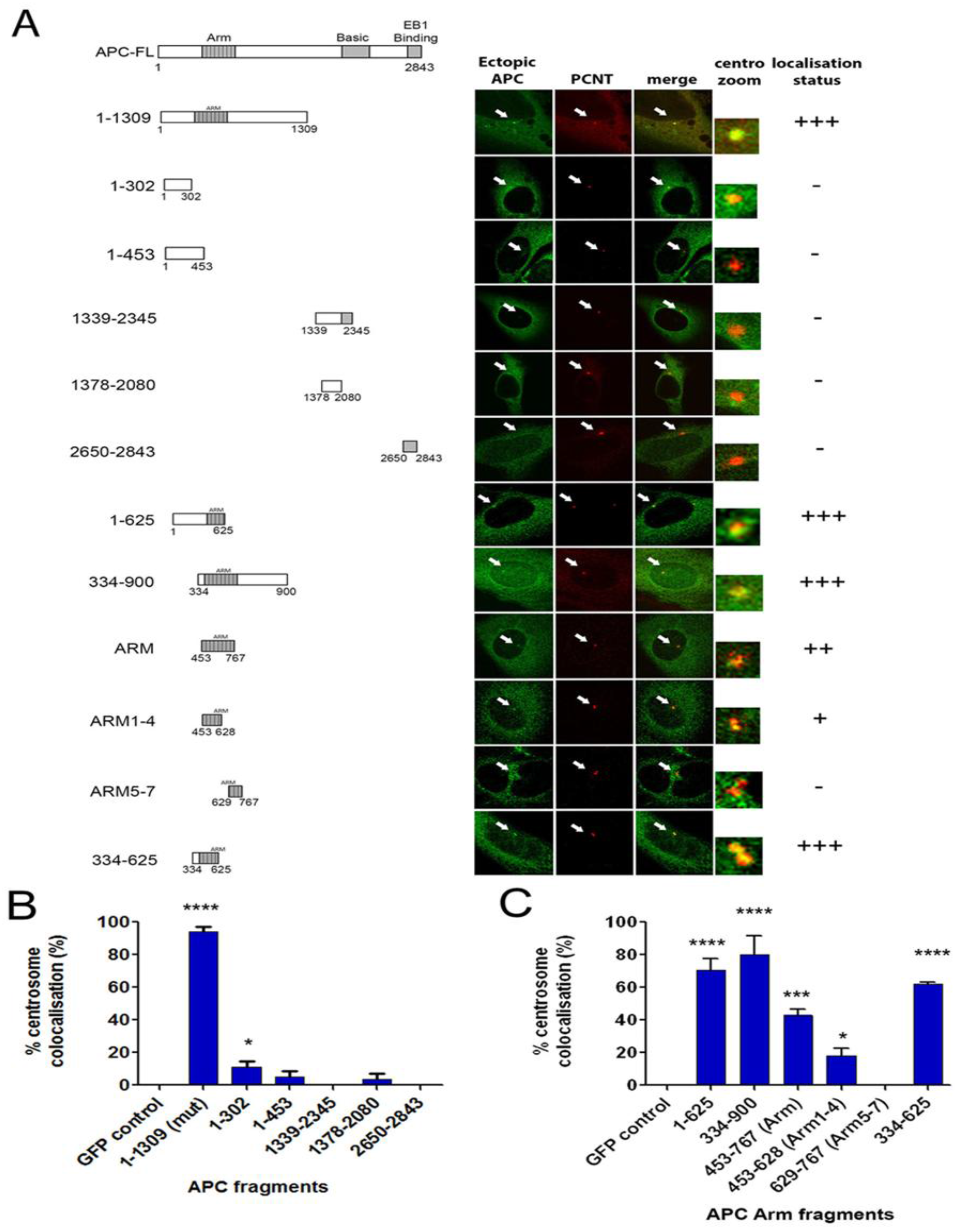
2.5. The Centrosome Targeting Sequence of APC Maps to the N-terminal ARM Domain Sequence 334-625
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Drug Treatment
4.2. Plasmids and Transfection
4.3. Centrosome Targeting Sequence Mapping Using Immunofluorescence Staining and Microscopy
4.4. Cell Retention Assay
4.5. Fluorescence Recovery after Photobleaching (FRAP)
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| APC | adenomatous polyposis coli |
| ARM | armadillo repeat |
| CRC | colorectal cancer |
| CSK | cytoskeletal extraction buffer |
| FRAP | fluorescence recovery after photobleaching |
| GFP | green fluorescent protein |
| RFP | red fluorescent protein |
| γ-tuRC | γ-tubulin ring complex |
| MT | microtubule |
| MTOC | microtubule organizing center |
| PCNT | pericentrin |
References
- Miyoshi, Y.; Nagase, H.; Ando, H.; Horii, A.; Ichii, S.; Nakatsuru, S.; Aoki, T.; Miki, Y.; Mori, T.; Nakamura, Y. Somatic mutations of the apc gene in colorectal tumors: Mutation cluster region in the apc gene. Hum. Mol. Genet. 1992, 1, 229–233. [Google Scholar] [PubMed]
- Nagase, H.; Miyoshi, Y.; Horii, A.; Aoki, T.; Ogawa, M.; Utsunomiya, J.; Baba, S.; Sasazuki, T.; Nakamura, Y. Correlation between the location of germ-line mutations in the apc gene and the number of colorectal polyps in familial adenomatous polyposis patients. Cancer Res. 1992, 52, 4055–4057. [Google Scholar] [PubMed]
- Powell, S.M.; Zilz, N.; Beazer-Barclay, Y.; Bryan, T.M.; Hamilton, S.R.; Thibodeau, S.N.; Vogelstein, B.; Kinzler, K.W. Apc mutations occur early during colorectal tumorigenesis. Nature 1992, 359, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Anastas, J.N.; Moon, R.T. Wnt signalling pathways as therapeutic targets in cancer. Nat. Rev. Cancer 2013, 13, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.H.; Wacker, I.; Appelt, U.K.; Behrens, J.; Schneikert, J. A common role for various human truncated adenomatous polyposis coli isoforms in the control of beta-catenin activity and cell proliferation. PLoS ONE 2012, 7, e34479. [Google Scholar] [CrossRef] [PubMed]
- Yamulla, R.J.; Kane, E.G.; Moody, A.E.; Politi, K.A.; Lock, N.E.; Foley, A.V.; Roberts, D.M. Testing models of the apc tumor suppressor/beta-catenin interaction reshapes our view of the destruction complex in wnt signaling. Genetics 2014, 197, 1285–1302. [Google Scholar] [CrossRef] [PubMed]
- Heinimann, K.; Mullhaupt, B.; Weber, W.; Attenhofer, M.; Scott, R.J.; Fried, M.; Martinoli, S.; Muller, H.; Dobbie, Z. Phenotypic differences in familial adenomatous polyposis based on apc gene mutation status. Gut 1998, 43, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Kinzler, K.W.; Vogelstein, B. Lessons from hereditary colorectal cancer. Cell 1996, 87, 159–170. [Google Scholar] [CrossRef]
- Aretz, S.; Uhlhaas, S.; Caspari, R.; Mangold, E.; Pagenstecher, C.; Propping, P.; Friedl, W. Frequency and parental origin of de novo apc mutations in familial adenomatous polyposis. Eur. J. Hum. Genet. 2004, 12, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Nathke, I. Cytoskeleton out of the cupboard: Colon cancer and cytoskeletal changes induced by loss of APC. Nat. Rev. Cancer 2006, 6, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Dikovskaya, D.; Zumbrunn, J.; Penman, G.A.; Nathke, I.S. The adenomatous polyposis coli protein: In the limelight out at the edge. Trends Cell Biol. 2001, 11, 378–384. [Google Scholar] [CrossRef]
- Fodde, R.; Kuipers, J.; Rosenberg, C.; Smits, R.; Kielman, M.; Gaspar, C.; van Es, J.H.; Breukel, C.; Wiegant, J.; Giles, R.H.; et al. Mutations in the apc tumour suppressor gene cause chromosomal instability. Nat. Cell Biol. 2001, 3, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Fodde, R.; Smits, R.; Clevers, H. APC, signal transduction and genetic instability in colorectal cancer. Nat. Rev. Cancer 2001, 1, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Tighe, A.; Johnson, V.L.; Taylor, S.S. Truncating apc mutations have dominant effects on proliferation, spindle checkpoint control, survival and chromosome stability. J. Cell. Sci. 2004, 117, 6339–6353. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, C.M.; Green, R.A.; Kaplan, K.B. Apc mutations lead to cytokinetic failures in vitro and tetraploid genotypes in min mice. J. Cell Biol. 2007, 178, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Field, J.J.; Kanakkanthara, A.; Miller, J.H. Microtubule-targeting agents are clinically successful due to both mitotic and interphase impairment of microtubule function. Bioorg. Med. Chem. 2014, 22, 5050–5059. [Google Scholar] [CrossRef] [PubMed]
- Ogden, A.; Rida, P.C.; Reid, M.D.; Aneja, R. Interphase microtubules: Chief casualties in the war on cancer? Drug Discov. Today 2014, 19, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Leung, L.; Brocardo, M.; Henderson, J.; Flegg, C.; Henderson, B.R. Membrane localization of adenomatous polyposis coli protein at cellular protrusions: Targeting sequences and regulation by beta-catenin. J. Biol. Chem. 2006, 281, 17140–17149. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Prosperi, J.R.; Goss, K.H. APC/beta-catenin-rich complexes at membrane protrusions regulate mammary tumor cell migration and mesenchymal morphology. BMC Cancer 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Lui, C.; Ashton, C.; Sharma, M.; Brocardo, M.G.; Henderson, B.R. APC functions at the centrosome to stimulate microtubule growth. Internat. J. Biochem. Cell Biol. 2015, 70, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Louie, R.K.; Bahmanyar, S.; Siemers, K.A.; Votin, V.; Chang, P.; Stearns, T.; Nelson, W.J.; Barth, A.I. Adenomatous polyposis coli and eb1 localize in close proximity of the mother centriole and EB1 is a functional component of centrosomes. J. Cell Sci. 2004, 117, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Dikovskaya, D.; Newton, I.P.; Nathke, I.S. The adenomatous polyposis coli protein is required for the formation of robust spindles formed in csf xenopus extracts. Mol. Biol. Cell 2004, 15, 2978–2991. [Google Scholar] [CrossRef] [PubMed]
- Olmeda, D.; Castel, S.; Vilaro, S.; Cano, A. Beta-catenin regulation during the cell cycle: Implications in g2/m and apoptosis. Mol. Biol. Cell 2003, 14, 2844–2860. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.M.; Brocardo, M.G.; Henderson, B.R. APC binds the miro/milton motor complex to stimulate transport of mitochondria to the plasma membrane. Mol. Biol. Cell 2016, 27, 466–482. [Google Scholar] [CrossRef] [PubMed]
- Brodie, K.M.; Henderson, B.R. Characterization of BRCA1 protein targeting, dynamics, and function at the centrosome: A role for the nuclear export signal, CRM1, and aurora a kinase. J. Biol. Chem. 2012, 287, 7701–7716. [Google Scholar] [CrossRef] [PubMed]
- Brodie, K.M.; Mok, M.T.; Henderson, B.R. Characterization of BARD1 targeting and dynamics at the centrosome: The role of CRM1, BRCA1 and the Q564H mutation. Cell. Signal. 2012, 24, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Prosser, S.L.; Fry, A.M. Fluorescence imaging of the centrosome cycle in mammalian cells. Methods Mol. Biol. 2009, 545, 165–183. [Google Scholar] [PubMed]
- Stenoien, D.L.; Sen, S.; Mancini, M.A.; Brinkley, B.R. Dynamic association of a tumor amplified kinase, aurora-a, with the centrosome and mitotic spindle. Cell. Motil. Cytoskeleton 2003, 55, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Hames, R.S.; Crookes, R.E.; Straatman, K.R.; Merdes, A.; Hayes, M.J.; Faragher, A.J.; Fry, A.M. Dynamic recruitment of Nek2 kinase to the centrosome involves microtubules, PCM-1, and localized proteasomal degradation. Mol. Biol. Cell 2005, 16, 1711–1724. [Google Scholar] [CrossRef] [PubMed]
- Akhmanova, A.; Steinmetz, M.O. Microtubule +tips at a glance. J. Cell Sci. 2010, 123, 3415–3419. [Google Scholar] [CrossRef] [PubMed]
- Zumbrunn, J.; Kinoshita, K.; Hyman, A.A.; Nathke, I.S. Binding of the adenomatous polyposis coli protein to microtubules increases microtubule stability and is regulated by GSK3 beta phosphorylation. Curr. Biol. 2001, 11, 44–49. [Google Scholar] [CrossRef]
- Moritz, M.; Agard, D.A. Gamma-tubulin complexes and microtubule nucleation. Curr. Opin. Struct. Biol. 2001, 11, 174–181. [Google Scholar] [CrossRef]
- Teixido-Travesa, N.; Roig, J.; Luders, J. The where, when and how of microtubule nucleation—One ring to rule them all. J. Cell Sci. 2012, 125, 4445–4456. [Google Scholar] [CrossRef] [PubMed]
- Moss, D.K.; Bellett, G.; Carter, J.M.; Liovic, M.; Keynton, J.; Prescott, A.R.; Lane, E.B.; Mogensen, M.M. Ninein is released from the centrosome and moves bi-directionally along microtubules. J. Cell Sci. 2007, 120, 3064–3074. [Google Scholar] [CrossRef] [PubMed]
- Khodjakov, A.; Rieder, C.L. The sudden recruitment of gamma-tubulin to the centrosome at the onset of mitosis and its dynamic exchange throughout the cell cycle, do not require microtubules. J. Cell Biol. 1999, 146, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Krieghoff, E.; Behrens, J.; Mayr, B. Nucleo-cytoplasmic distribution of beta-catenin is regulated by retention. J. Cell Sci. 2006, 119, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Bahmanyar, S.; Kaplan, D.D.; Deluca, J.G.; Giddings, T.H., Jr.; O’Toole, E.T.; Winey, M.; Salmon, E.D.; Casey, P.J.; Nelson, W.J.; Barth, A.I. Beta-catenin is a Nek2 substrate involved in centrosome separation. Genes Dev. 2008, 22, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Flegg, C.P.; Sharma, M.; Medina-Palazon, C.; Jamieson, C.; Galea, M.; Brocardo, M.G.; Mills, K.; Henderson, B.R. Nuclear export and centrosome targeting of the protein phosphatase 2a subunit b56alpha: Role of b56alpha in nuclear export of the catalytic subunit. J. Biol. Chem. 2010, 285, 18144–18154. [Google Scholar] [CrossRef] [PubMed]
- Bouissou, A.; Verollet, C.; Sousa, A.; Sampaio, P.; Wright, M.; Sunkel, C.E.; Merdes, A.; Raynaud-Messina, B. {gamma}-tubulin ring complexes regulate microtubule plus end dynamics. J. Cell Biol. 2009, 187, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.H.; Whipple, R.A.; Matrone, M.A.; Balzer, E.M.; Martin, S.S. Delocalization of gamma-tubulin due to increased solubility in human breast cancer cell lines. Cancer Biol. Ther. 2010, 9, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Haren, L.; Remy, M.H.; Bazin, I.; Callebaut, I.; Wright, M.; Merdes, A. Nedd1-dependent recruitment of the gamma-tubulin ring complex to the centrosome is necessary for centriole duplication and spindle assembly. J. Cell Biol. 2006, 172, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Haren, L.; Stearns, T.; Luders, J. Plk1-dependent recruitment of gamma-tubulin complexes to mitotic centrosomes involves multiple PCM components. PLoS ONE 2009, 4, e5976. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Koch, B.; Walczak, R.; Ciray-Duygu, F.; Gonzalez-Sanchez, J.C.; Devos, D.P.; Mattaj, I.W.; Gruss, O.J. The nucleoporin MEL-28 promotes RanGTP-dependent gamma-tubulin recruitment and microtubule nucleation in mitotic spindle formation. Nat. Commun. 2014, 5, 3270. [Google Scholar] [CrossRef] [PubMed]
- Munemitsu, S.; Souza, B.; Muller, O.; Albert, I.; Rubinfeld, B.; Polakis, P. The APC gene product associates with microtubules in vivo and promotes their assembly in vitro. Cancer Res. 1994, 54, 3676–3681. [Google Scholar] [PubMed]
- Jimbo, T.; Kawasaki, Y.; Koyama, R.; Sato, R.; Takada, S.; Haraguchi, K.; Akiyama, T. Identification of a link between the tumour suppressor APC and the kinesin superfamily. Nat. Cell Biol. 2002, 4, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Lui, C.; Mills, K.; Brocardo, M.G.; Sharma, M.; Henderson, B.R. APC as a mobile scaffold: Regulation and function at the nucleus, centrosomes, and mitochondria. IUBMB Life 2012, 64, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Kunttas-Tatli, E.; Roberts, D.M.; McCartney, B.M. Self-association of the APC tumor suppressor is required for the assembly, stability, and activity of the wnt signaling destruction complex. Mol. Biol Cell. 2014, 25, 3424–3436. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kroboth, K.; Newton, I.P.; Nathke, I.S. Novel self-association of the APC molecule affects apc clusters and cell migration. J. Cell Sci. 2008, 121, 1916–1925. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APC +/− Treatments | Mobile Pool 2 (%) | Retained Pool 2 (%) | T1/2 Fast Pool 3 (s) | T1/2 Slow Pool (s) |
|---|---|---|---|---|
| APC-FL | 86.23 | 13.77 | 2.77 | 25.58 |
| APC-FL+ noco | 97.30 | 2.70 | 6.79 (p < 0.0001) | 42.67 |
| APC1-1309 | 91.31 | 8.69 | 3.09 | 27.82 |
| APC1-1309+noco | 90.39 | 9.61 | 5.99 (p = 0.045) | 25.79 |
| APC1309 +control siRNA | 87.81 | 12.19 | 3.02 | 23.34 |
| APC1309 + γ-tub siRNA | 83.77 | 16.23 | 5.44 (p = 0.019) | 10.52 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lui, C.; Mok, M.T.S.; Henderson, B.R. Characterization of Adenomatous Polyposis Coli Protein Dynamics and Localization at the Centrosome. Cancers 2016, 8, 47. https://doi.org/10.3390/cancers8050047
Lui C, Mok MTS, Henderson BR. Characterization of Adenomatous Polyposis Coli Protein Dynamics and Localization at the Centrosome. Cancers. 2016; 8(5):47. https://doi.org/10.3390/cancers8050047
Chicago/Turabian StyleLui, Christina, Myth T. S. Mok, and Beric R. Henderson. 2016. "Characterization of Adenomatous Polyposis Coli Protein Dynamics and Localization at the Centrosome" Cancers 8, no. 5: 47. https://doi.org/10.3390/cancers8050047