
Role of the Aryl Hydrocarbon Receptor in Colon Neoplasia

Division of Gastroenterology and Hepatology, Veterans Administration Maryland Health Care System, University of Maryland School of Medicine, Baltimore, MD 21201, USA
*
Author to whom correspondence should be addressed.
Cancers 2015, 7(3), 1436-1446; https://doi.org/10.3390/cancers7030847
Submission received: 8 June 2015
/
Revised: 27 July 2015
/
Accepted: 29 July 2015
/
Published: 31 July 2015
{kind=link}
Abstract
:For both men and women, colorectal cancer (CRC) is the second leading cause of cancer death in the United States, primarily as a consequence of limited therapies for metastatic disease. The aryl hydrocarbon receptor (AhR) is a ligand-dependent transcription factor with diverse functions in detoxification of xenobiotics, inflammatory responses, and tissue homeostasis. Emerging evidence indicates that AhR also plays an important role in regulating intestinal cell proliferation and tumorigenesis. Here, we review both the pro- and anti-carcinogenic properties of AhR signaling and its potential role as a therapeutic target in CRC.
1. Introduction
1.1. Colorectal Cancer is a Major Health Concern
In the U.S., Colorectal cancer (CRC) is the second leading cause of cancer death in men and women combined [1,2,3,4]. The American Cancer Society estimates that in 2015, 137,000 people will be diagnosed with CRC, 50,000 of whom will die from this disease [5]. Although colonoscopy and stool-based screening tests are effective at preventing and reducing mortality from CRC, many people fail to undergo such tests [6]. Also, the efficacy of colon cancer screening is limited by the sensitivity of stool-based tests and “miss” rates on colonoscopy [7,8]. Non-steroidal anti-inflammatory drugs are marginally effective chemo-preventive agents [9,10] and limited by gastrointestinal [11] and cardiovascular [12] toxicity that led to the withdrawal of rofecoxib [13]. Chemotherapy and radiation are not very effective for patients with advanced disease. Recently, the use of biologicals targeting vascular endothelial growth factor (VEGF) and the epidermal growth factor receptor (EGFR; i.e., Bevacizumab, Cetuximab and Panitumumab) were shown to increase survival in patients with advanced CRC, but only by a few months with limited impact on five-year survival—which remains on the order of 5–10% [14,15,16]. Furthermore, the use of these agents is limited by off-target toxicity that includes gastrointestinal bleeding and skin rashes that often lead to discontinuation of therapy [17].
1.2. The Aryl Hydrocarbon Receptor
The aryl hydrocarbon receptor (AhR) is a cytosolic ligand-dependent transcription factor that belongs to the basic helix-loop-helix/Per-Arnt-Sim (bHLH/PAS) family [18]. AhR has diverse physiological functions including detoxification of environmental pollutants, mucosal inflammation and tissue regeneration [19]. When inactive, AhR remains bound to several chaperon proteins including heat-shock protein 90 and the AhR-interacting protein [20,21,22,23]. Upon ligand binding, cytosolic AhR translocates into the nucleus and forms a heterodimer with the AhR nuclear translocator (ARNT) protein, which then interacts with dioxin responsive elements (DREs) on the promoters of AhR target genes including cytochrome P450 enzymes CYP1A1/A2/B1, thereby activating transcription [20].
1.3. AhR Ligands
Numerous AhR ligands have been identified, including synthetic and environmental chemicals, naturally-occurring dietary and endogenous compounds [24,25,26,27]. Environmental pollutants include planar halogenated aromatics [e.g., 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)], synthetic polynuclear aromatic hydrocarbons and dioxin-like compounds. TCDD is the prototype of many chlorinated aromatic compounds that are abundant contaminants in the environment. With low-dose administration to rodents, TCDD acts as a carcinogen and causes tumors in multiple sites including the liver, lung, nasal turbinates, hard palate, thyroid, tongue, and skin, but not the intestines [28,29], indicating tissue-specific carcinogenic properties. Other common toxicants include halogenated dibenzo-p-dioxins, dibenzofurans, azo(xy)benzenes, and naphthalenes. Manufactured drugs include proton pump inhibitors, Sulindac and 4-hydroxytamoxifen. Candidates for endogenous ligands include indigoids and metabolites of arachidonic acid, heme and tryptophan. AhR antagonists include CH223191, GNF351, and 3ʹ-4ʹ-Dimethoxy-α-Naphthoflavone [25].
Colorectal cancer is often linked to a “so-called” Western diet, rich in carbohydrates, red meats and saturated fats, but low in vegetables and fresh fruits [30,31]. Recent data have shown that certain dietary components from vegetables have protective effects against CRC [32]. Several natural dietary AhR ligands may play important roles in mediating this protective effect. The tryptophan metabolites indole derivatives, including indole-3-acetic acid (IAA) and indole-3-carbinol (I3C) are natural AhR ligands generated by conversion from dietary tryptophan. Another indole derivative, 3,3ʹ-diindolylmethane (DIM) is converted from glucosinolates by commensal intestinal microbiota [33]. Glucosinolates are natural components of many pungent plants and cruciferous vegetables such as mustard, cabbage, cauliflower, broccoli and horseradish. An I3C derivative indole[3,2-b]carbazole (ICZ) has strong receptor binding affinity for AhR. It is reported that glucosinolates and tryptophan metabolites have chemo-preventive effects on CRC [34,35,36], perhaps by enhancing apoptosis in cancer cells [34] or modulating gut immune homeostasis [37,38].
Naturally-occurring flavonoids, ubiquitous in many fruits and vegetables, are another class of dietary AhR ligands that have chemo-preventive effects for CRC [39]. Statistically significant reductions in CRC risk were associated with the five classes of flavonoids-flavonols, quercetin, catechin, epicatechin and procyanisins. Risk reductions were greater in non-smokers. Many of these flavonoids activate AhR in vitro [40,41]. Veeriah et al. reported that flavonoids extracted from apples inhibited growth (cell proliferation) of HT29 human CRC cells [42].
2. Role of AhR in Colon Tumorigenesis
2.1. AhR Is a Tumor Suppressor in Mouse Models of CRC
The role of AhR in carcinogenesis remains controversial. Recent evidence supports both pro- and anti-carcinogenic properties of AhR signaling, perhaps in a tissue-selective manner. The Wnt/β-catenin signaling is a major signal transduction pathway involved in colon carcinogenesis. In quiescent cells β-catenin is sequestered in a multi-protein complex, including axin, adenomatous polyposis coli (APC) and glycogen synthase kinase-3β (GSK-3β), that targets β-catenin for phosphorylation, ubiquitination and proteosomal degradation [43,44]. Wnt ligands activate a cascade that inhibits GSK-3β-induced β-catenin phosphorylation, frees β-catenin from the destruction complex, and allows its nuclear translocation and subsequent activation of target genes leading to increased cell proliferation and tumorigenesis [43,44,45]. Kawajiri et al. showed that AhR-deficient mice spontaneously develop cecal adenocarcinomas by the age of 30 to 40 weeks [46]. These investigators demonstrated dual roles for AhR in regulating intracellular protein levels, both as a ligand-activated transcription factor and as a ligand-dependent E3 ubiquitin ligase [18,46]. AhR suppresses intestinal carcinogenesis by a ligand-dependent β-catenin degradation pathway that functions independently of and cooperatively with the canonical APC-dependent system. Natural AhR ligands converted from dietary tryptophan and glucoinolates in the intestines are as efficient as exogenous xenobiotic ligands in suppressing tumor formation in Apcmin/+ mice [46]. In addition, AhR also functions as a tumor suppressor for liver carcinogenesis by inhibiting cell proliferation through G0-G1 cell cycle arrest [47].
2.2. AhR Expression in Colon Tumors
AhR is ubiquitously expressed in mouse and human tissues, including the gastrointestinal tract [48,49]. The AhR mRNA expression profile of 967 human cancer cell lines showed that moderate levels of AhR are expressed in colon cancer cells [50]. It is not clear how this level of AhR expression compares to that of normal intestinal epithelial cells. Interestingly, reduced AhR expression was observed in specimens of human cecal cancers and adjacent tissues [46].
2.3. AhR Target Genes and Their Roles in CRC
AhR may regulate intestinal tumorigenesis through its target genes, including the Phase I drug metabolizing enzymes CYP1A1, CYP1A2, and CYP1B1. CYP1A1 and CYP1B1 are extrahepatic enzymes that catalyze conversion of polycyclic aromatic hydrocarbons including benzo[a]pyrene to active genotoxic metabolites, thereby contributing to carcinogenesis [51]. Androutsopoulos et al. showed that CYP1A1 and CYP1B1 are overexpressed in 80% and 60% of human colon tumors, respectively, suggesting an important role for these enzymes in colon neoplasia [52]. In addition, it was shown that AhR activation in colon cancer cells induces expression of multiple target genes including matrix metalloproteinase (MMP)-9, calcium ion flux, pro-inflammatory IL-1β and the drug transporter BCRP/ABCG2 [53,54,55].
2.4. AhR Cross-Talks with Multiple Signaling Pathways
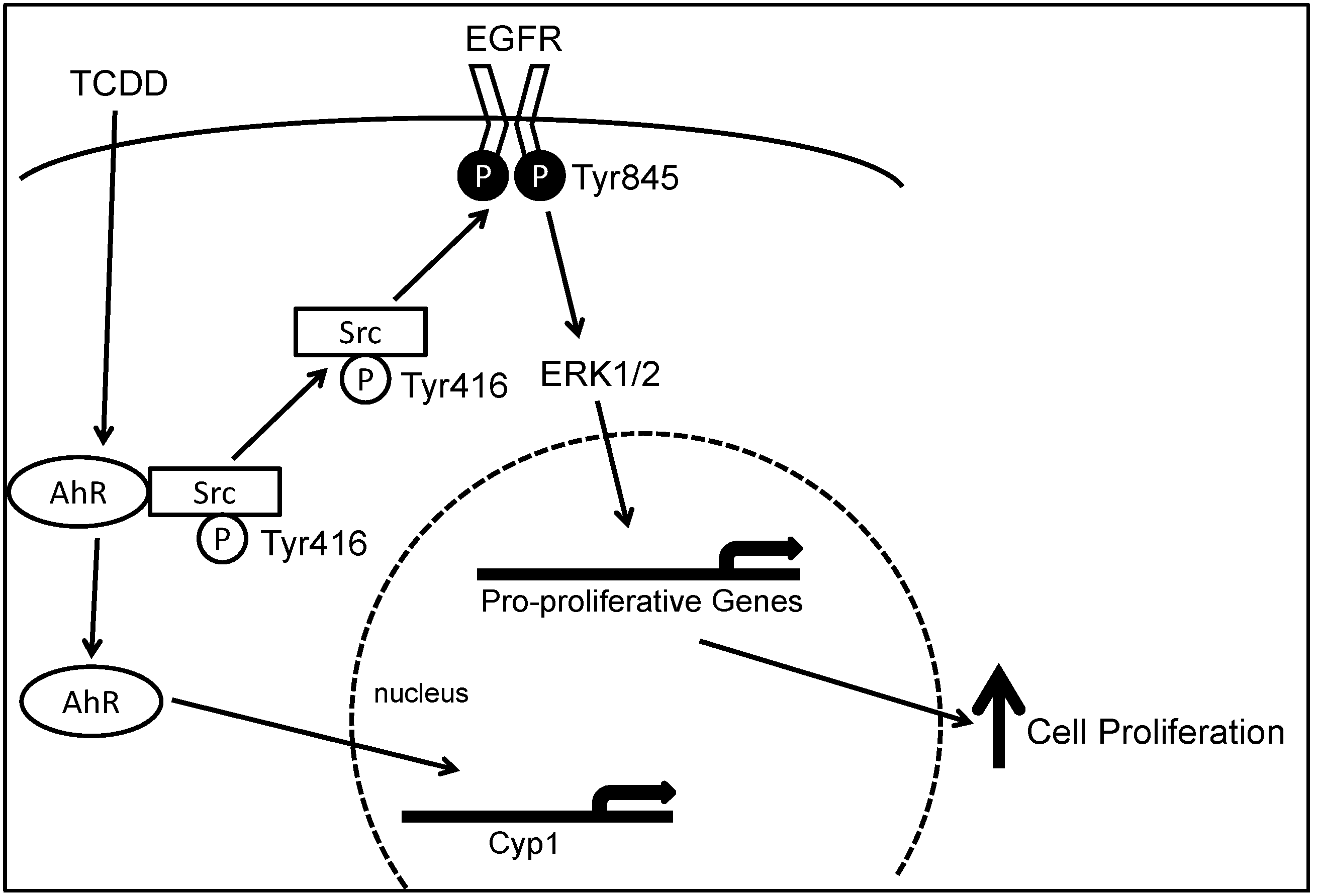
AhR cross-talks with multiple growth factor-mediated signal transduction pathways including transforming growth factor-β, tumor necrosis factor-α, EGFR and Src pathways [56,57,58,59,60]. As shown in Figure 1, we demonstrated that in human colon cancer cell lines, upon AhR activation by TCDD, Src-mediated cross-talk between AhR and EGFR results in ERK1/2 activation and enhanced cell proliferation [61]. Also, AhR interacts with the retinoblastoma protein (pRB) protein to inhibit G1 to S phase cell cycle transition via protein kinase C and p38 MAPK [62].
Figure 1.
Model depicting molecular mechanisms underlying 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-induced, Src-mediated activation of the epidermal growth factor receptor (EGFR) signaling in colon epithelial cells. Binding of aryl hydrocarbon receptor (AhR) by ligand, e.g., TCDD, results in phosphorylation of Src kinase at Tyr416 and de-phosphorylation of Src kinase at Tyr527. Activated Src either directly phosphorylates EGFR at Tyr 485 or indirectly activate EGFR through matrix metalloproteinase (MMP) and heparin-binding (HB)EGF, leading to activation of EGFR and downstream ERK1/2 signaling, which stimulates gene transcription and cell proliferation. Binding of AhR by TCDD also causes translocation of AhR from the cytosol to the nucleus where it acts as a transcription factor for CYP1A1 and CYP1B1. Reproduced with permission from G. Xie, Z. Peng and J.P. Raufman. Am. J. Physiol. Gastrointest. Liver Physiol.; published by the American Physiological Society, 2012.
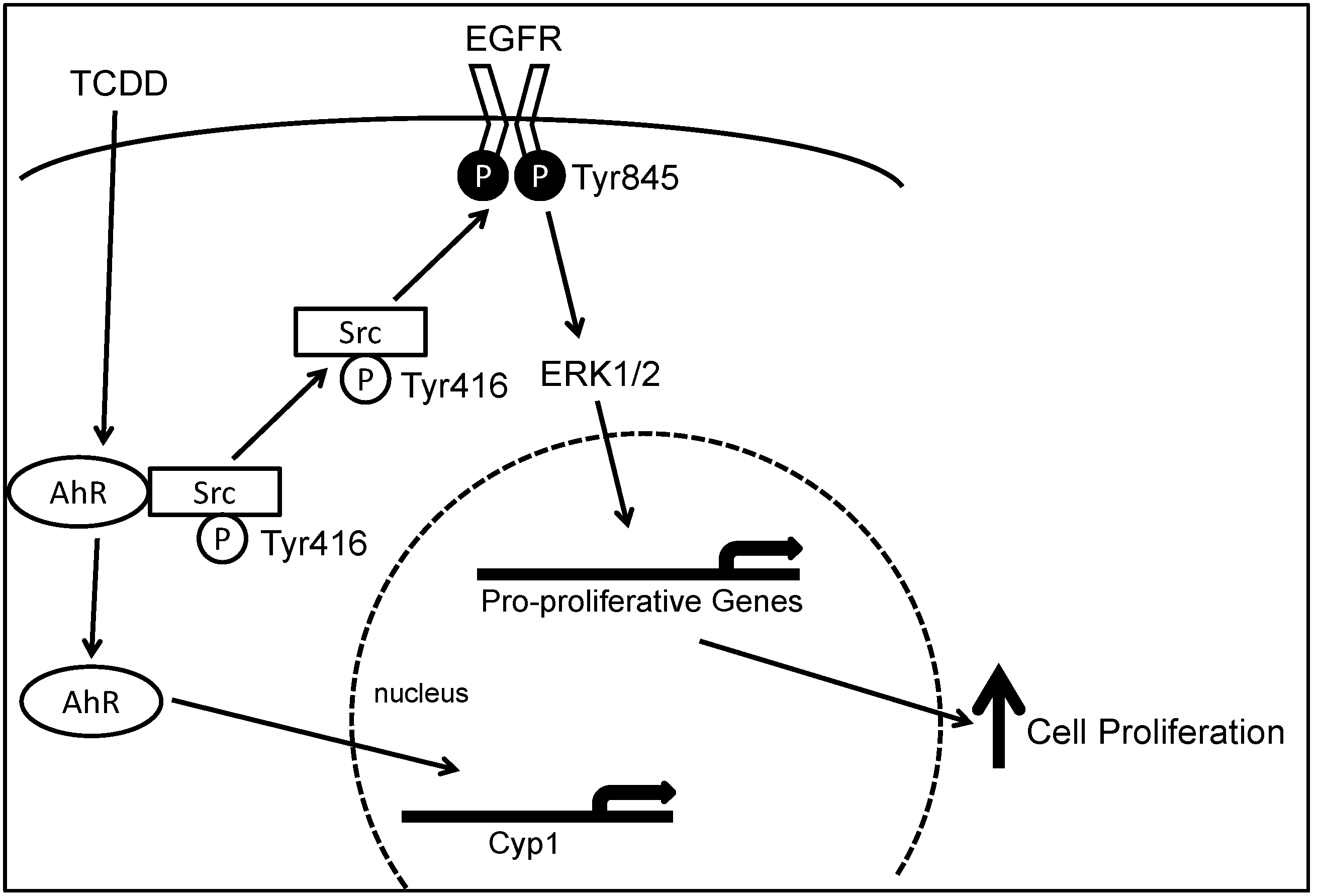
Figure 1.
Model depicting molecular mechanisms underlying 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD)-induced, Src-mediated activation of the epidermal growth factor receptor (EGFR) signaling in colon epithelial cells. Binding of aryl hydrocarbon receptor (AhR) by ligand, e.g., TCDD, results in phosphorylation of Src kinase at Tyr416 and de-phosphorylation of Src kinase at Tyr527. Activated Src either directly phosphorylates EGFR at Tyr 485 or indirectly activate EGFR through matrix metalloproteinase (MMP) and heparin-binding (HB)EGF, leading to activation of EGFR and downstream ERK1/2 signaling, which stimulates gene transcription and cell proliferation. Binding of AhR by TCDD also causes translocation of AhR from the cytosol to the nucleus where it acts as a transcription factor for CYP1A1 and CYP1B1. Reproduced with permission from G. Xie, Z. Peng and J.P. Raufman. Am. J. Physiol. Gastrointest. Liver Physiol.; published by the American Physiological Society, 2012.
2.5. Role of AhR in Chronic Intestinal Inflammation
AhR activation by dietary ligands is required for the maintenance of intestinal immune homeostasis including innate immune cells such as intraepithelial lymphocytes (IELs) and interleukin (IL)-22-producing lymphoid cells [63]. AhR-deficient mice lack IELs and are more susceptible to bacterial infections and experimental colitis. In patients with Crohn’s disease, intestinal T cells and natural killer cells have reduced AhR expression levels [64]. Furumatsu et al. showed that dextran sodium sulfate (DSS)-induced colitis was more severe in AhR-knockout mice than in wild-type mice [65]. Administration of DSS increased AhR expression in the colonic epithelium [65]. Also, oral administration of the AhR agonist β-naphthoflavone attenuated DSS-evoked colitis [65]. In addition, AhR activation by 6-formylindolo (3,2-b) carbazole (Ficz) down-regulates IL-7 and reduces inflammation in DSS-induced colitis [66].
2.6. Role of AhR in Inflammation-Associated Colon Neoplasia
The risk of CRC is increased with chronic intestinal inflammation as is observed with inflammatory bowel disease, including both Crohn’s and ulcerative colitis [67]. Ikuta et al. showed that AhR-deficient mice develop cecal tumors with severe inflammation which is dependent on the apoptosis-associated speck-like protein containing a caspase recruitment protein (ASC) [68]. In AhR-deficient mice, blocking interleukin (IL)-1β signaling with a caspase-1 inhibitor attenuated cecal tumorigenesis in AhR-deficient mice [68]. Also, germ-free AhR (−/−) and AhR (−/−)/ASC (−/−) mice had reduced tumor formation compared with AhR (−/−) mice [68]. These observations suggest that AhR also acts as tumor suppressor in inflammation-associated intestinal neoplasia.
2.7. Role of AhR in Circadian Clock Circuitry and CRC
Growing evidence indicates that there is physiological cross-talk between AhR and the circadian system [69]. The AhR complex is involved in sensing and transforming environmental xenobiotics and naturally occurring AhR ligands. Epidemiological studies showed an increased incidences of CRC in shift workers, suggesting disruption of circadian timing may contribute to CRC carcinogenesis, possibly by interrupting intestinal homeostatic control of cell proliferation, differentiation and apoptosis [70].
3. Conclusions
Emerging evidence suggests that AhR and its ligands play important roles in intestinal tumorigenesis, identifying AhR as a potential therapeutic target for CRC. Although most previously-known AhR ligands are toxic HAs and genotoxic PAHs, many newly-identified AhR agonists are non-toxic and have therapeutic potential [24,25,26]. However, there remain inconsistencies regarding the role of AhR in colon neoplasia. Studies in animal models of CRC suggest that in the basal state without xenobiotic ligands AhR acts as a tumor suppressor, particularly in cecal tumors. This tumor-suppressive action of AhR is radically different from the pro-oncogenic role that AhR plays as a receptor for pro-carcinogenic ligands such as TCDD. Over-expression of CYP1A1 and CYP1B1 in human colon tumors implies a pro-carcinogenic role for AhR target genes. In addition, studies using human colon cancer cell lines showed that AhR activation in vitro increases cell proliferation, migration, and invasion via transactivation of the EGFR/ERK signaling pathway. Interestingly, most in vivo studies demonstrated protective effects of AhR in CRC; in contrast, many in vitro studies showed pro-carcinogenic effects of AhR ligands. This disconnect suggests that colon cancer cell lines may not always be reliable for predicting the in vivo role of AhR in colon carcinogenesis. Further translational studies are needed to clarify the role of AhR in different stages of CRC progression, including initiation, growth and dissemination. Such information will be useful to guide potential AhR-based therapies for CRC.
Acknowledgments
This work was supported by American Cancer Society Institutional Research Grant (G. Xie), National Institute of Health grants K08 DK-080843 (G. Xie), T32 DK067872 (J.P. Raufman) and R01 CA-120407 (J.P. Raufman).
Author Contributions
G.X. wrote the article. J.P. edited and revised the article. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siegel, R.; Desantis, C.; Jemal, A. Colorectal cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Peery, A.F.; Dellon, E.S.; Lund, J.; Crockett, S.D.; McGowan, C.E.; Bulsiewicz, W.J.; Gangarosa, L.M.; Thiny, M.T.; Stizenberg, K.; Morgan, D.R.; et al. Burden of gastrointestinal disease in the united states: 2012 update. Gastroenterology 2012, 143. [Google Scholar] [CrossRef] [PubMed]
- Edwards, B.K.; Ward, E.; Kohler, B.A.; Eheman, C.; Zauber, A.G.; Anderson, R.N.; Jemal, A.; Schymura, M.J.; Lansdorp-Vogelaar, I.; Seeff, L.C.; et al. Annual report to the nation on the status of cancer, 1975–2006, featuring colorectal cancer trends and impact of interventions (risk factors, screening, and treatment) to reduce future rates. Cancer 2010, 116, 544–573. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Xu, J.; Ward, E. Cancer statistics, 2010. CA Cancer J. Clin. 2010, 60, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Hewett, D.G.; Rex, D.K. The big picture: Does colonoscopy work? Gastrointest. Endosc. Clin. N. Am. 2015, 25, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Pignone, M.; Saha, S.; Hoerger, T.; Mandelblatt, J. Cost-effectiveness analyses of colorectal cancer screening: A systematic review for the U.S. Preventive services task force. Ann. Intern. Med. 2002, 137, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Corley, D.A. The future of colon cancer screening: What do we recommend and will it be too much, too little, or just right? Gastroenterology 2011, 141, 1956–1958. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.A.; Sandler, R.S.; Bresalier, R.S.; Quan, H.; Riddell, R.; Lanas, A.; Bolognese, J.A.; Oxenius, B.; Horgan, K.; Loftus, S.; et al. A randomized trial of rofecoxib for the chemoprevention of colorectal adenomas. Gastroenterology 2006, 131, 1674–1682. [Google Scholar] [CrossRef] [PubMed]
- Bertagnolli, M.M.; Eagle, C.J.; Zauber, A.G.; Redston, M.; Solomon, S.D.; Kim, K.; Tang, J.; Rosenstein, R.B.; Wittes, J.; Corle, D.; et al. Celecoxib for the prevention of sporadic colorectal adenomas. N. Engl. J. Med. 2006, 355, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Helin-Salmivaara, A.; Saarelainen, S.; Gronroos, J.M.; Vesalainen, R.; Klaukka, T.; Huupponen, R. Risk of upper gastrointestinal events with the use of various nsaids: A case-control study in a general population. Scand. J. Gastroenterol. 2007, 42, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.J.; Dunn, J.A.; Langman, M.J.; Smith, J.L.; Midgley, R.S.; Stanley, A.; Stokes, J.C.; Julier, P.; Iveson, C.; Duvvuri, R.; et al. Rofecoxib and cardiovascular adverse events in adjuvant treatment of colorectal cancer. N. Engl. J. Med. 2007, 357, 360–369. [Google Scholar] [CrossRef] [PubMed]
- McGettigan, P.; Henry, D. Cardiovascular risk and inhibition of cyclooxygenase: A systematic review of the observational studies of selective and nonselective inhibitors of cyclooxygenase 2. JAMA 2006, 296, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Modjtahedi, H.; Essapen, S. Epidermal growth factor receptor inhibitors in cancer treatment: Advances, challenges and opportunities. Anticancer Drugs 2009, 20, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Overman, M.J.; Hoff, P.M. Egfr-targeted therapies in colorectal cancer. Dis. Colon Rectum 2007, 50, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Venook, A.P. Epidermal growth factor receptor-targeted treatment for advanced colorectal carcinoma. Cancer 2005, 103, 2435–2446. [Google Scholar] [CrossRef] [PubMed]
- Sipples, R. Common side effects of anti-egfr therapy: Acneform rash. Semin. Oncol. Nurs. 2006, 22, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Ohtake, F.; Fujii-Kuriyama, Y.; Kato, S. AhR acts as an E3 ubiquitin ligase to modulate steroid receptor functions. Biochem. Pharmacol. 2009, 77, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Yamamoto, H.; Itoh, F.; Hinoda, Y.; Okada, Y.; Imai, K. Contribution of matrilysin (MMP-7) to the metastatic pathway of human colorectal cancers. Gut 1999, 45, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Hankinson, O. The aryl hydrocarbon receptor complex. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 307–340. [Google Scholar] [CrossRef] [PubMed]
- Carver, L.A.; Bradfield, C.A. Ligand-dependent interaction of the aryl hydrocarbon receptor with a novel immunophilin homolog in vivo. J. Biol. Chem. 1997, 272, 11452–11456. [Google Scholar] [PubMed]
- Chen, H.S.; Perdew, G.H. Subunit composition of the heteromeric cytosolic aryl hydrocarbon receptor complex. J. Biol. Chem. 1994, 269, 27554–27558. [Google Scholar] [PubMed]
- Ma, Q.; Whitlock, J.P., Jr. A novel cytoplasmic protein that interacts with the ah receptor, contains tetratricopeptide repeat motifs, and augments the transcriptional response to 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 1997, 272, 8878–8884. [Google Scholar] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the aryl hydrocarbon receptor by structurally diverse exogenous and endogenous chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.P.; Bradfield, C.A. The search for endogenous activators of the aryl hydrocarbon receptor. Chem. Res. Toxicol. 2008, 21, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Lee, S.O.; Jin, U.H. Role of the aryl hydrocarbon receptor in carcinogenesis and potential as a drug target. Toxicol. Sci. 2013, 135, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Murray, I.A.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat. Rev. Cancer 2014, 14, 801–814. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W.; Kohle, C. Ah receptor- and TCDD-mediated liver tumor promotion: Clonal selection and expansion of cells evading growth arrest and apoptosis. Biochem. Pharmacol. 2005, 69, 1403–1408. [Google Scholar] [CrossRef] [PubMed]
- Knerr, S.; Schrenk, D. Carcinogenicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in experimental models. Mol. Nutr. Food Res. 2006, 50, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Thun, M.J.; Connell, C.J.; McCullough, M.L.; Jacobs, E.J.; Flanders, W.D.; Rodriguez, C.; Sinha, R.; Calle, E.E. Meat consumption and risk of colorectal cancer. JAMA 2005, 293, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, H.; Johnsen, N.F.; Christensen, J.; Frederiksen, K.; Overvad, K.; Tjonneland, A. Association of adherence to lifestyle recommendations and risk of colorectal cancer: A prospective danish cohort study. BMJ 2010, 341. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Bjeldanes, L.F.; Kim, J.Y.; Grose, K.R.; Bartholomew, J.C.; Bradfield, C.A. Aromatic hydrocarbon responsiveness-receptor agonists generated from indole-3-carbinol in vitro and in vivo: Comparisons with 2,3,7,8-tetrachlorodibenzo-p-dioxin. Proc. Natl. Acad. Sci. USA 1991, 88, 9543–9547. [Google Scholar] [CrossRef] [PubMed]
- Bonnesen, C.; Eggleston, I.M.; Hayes, J.D. Dietary indoles and isothiocyanates that are generated from cruciferous vegetables can both stimulate apoptosis and confer protection against DNA damage in human colon cell lines. Cancer Res. 2001, 61, 6120–6130. [Google Scholar] [PubMed]
- Kim, Y.S.; Milner, J.A. Targets for indole-3-carbinol in cancer prevention. J. Nutr. Biochem. 2005, 16, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Potter, J.D.; Steinmetz, K. Vegetables, fruit and phytoestrogens as preventive agents. IARC Sci. Publ. 1996, 139, 61–90. [Google Scholar] [PubMed]
- Jin, U.H.; Lee, S.O.; Sridharan, G.; Lee, K.; Davidson, L.A.; Jayaraman, A.; Chapkin, R.S.; Alaniz, R.; Safe, S. Microbiome-derived tryptophan metabolites and their aryl hydrocarbon receptor-dependent agonist and antagonist activities. Mol. Pharmacol. 2014, 85, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; de Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Theodoratou, E.; Kyle, J.; Cetnarskyj, R.; Farrington, S.M.; Tenesa, A.; Barnetson, R.; Porteous, M.; Dunlop, M.; Campbell, H. Dietary flavonoids and the risk of colorectal cancer. Cancer Epidemiol. Biomarkers Prev. 2007, 16, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Amakura, Y.; Tsutsumi, T.; Nakamura, M.; Kitagawa, H.; Fujino, J.; Sasaki, K.; Toyoda, M.; Yoshida, T.; Maitani, T. Activation of the aryl hydrocarbon receptor by some vegetable constituents determined using in vitro reporter gene assay. Biol. Pharm. Bull. 2003, 26, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Qin, C.; Safe, S.H. Flavonoids as aryl hydrocarbon receptor agonists/antagonists: Effects of structure and cell context. Environ. Health Perspect. 2003, 111, 1877–1882. [Google Scholar] [CrossRef] [PubMed]
- Veeriah, S.; Kautenburger, T.; Habermann, N.; Sauer, J.; Dietrich, H.; Will, F.; Pool-Zobel, B.L. Apple flavonoids inhibit growth of HT29 human colon cancer cells and modulate expression of genes involved in the biotransformation of xenobiotics. Mol. Carcinog. 2006, 45, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Moon, R.T.; Kohn, A.D.; de Ferrari, G.V.; Kaykas, A. Wnt and beta-catenin signalling: Diseases and therapies. Nat. Rev. Genet. 2004, 5, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, S.D.; Bertagnolli, M.M. Molecular origins of cancer: Molecular basis of colorectal cancer. N. Engl. J. Med. 2009, 361, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Kawajiri, K.; Kobayashi, Y.; Ohtake, F.; Ikuta, T.; Matsushima, Y.; Mimura, J.; Pettersson, S.; Pollenz, R.S.; Sakaki, T.; Hirokawa, T.; et al. Aryl hydrocarbon receptor suppresses intestinal carcinogenesis in apcmin/+ mice with natural ligands. Proc. Natl. Acad. Sci. USA 2009, 106, 13481–13486. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Boivin, G.P.; Knudsen, E.S.; Nebert, D.W.; Xia, Y.; Puga, A. The aryl hydrocarbon receptor functions as a tumor suppressor of liver carcinogenesis. Cancer Res. 2010, 70, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Dolwick, K.M.; Schmidt, J.V.; Carver, L.A.; Swanson, H.I.; Bradfield, C.A. Cloning and expression of a human ah receptor cdna. Mol. Pharmacol. 1993, 44, 911–917. [Google Scholar] [PubMed]
- Hayashi, S.; Watanabe, J.; Nakachi, K.; Eguchi, H.; Gotoh, O.; Kawajiri, K. Interindividual difference in expression of human ah receptor and related p450 genes. Carcinogenesis 1994, 15, 801–806. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, E.F.; Kopparapu, P.R.; Koch, D.C.; Jang, H.S.; Phillips, J.L.; Tanguay, R.L.; Kerkvliet, N.I.; Kolluri, S.K. The aryl hydrocarbon receptor mediates leflunomide-induced growth inhibition of melanoma cells. PLoS ONE 2012, 7, e40926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Cytochrome p450 cyp1a1: Wider roles in cancer progression and prevention. BMC Cancer 2009, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Androutsopoulos, V.P.; Spyrou, I.; Ploumidis, A.; Papalampros, A.E.; Kyriakakis, M.; Delakas, D.; Spandidos, D.A.; Tsatsakis, A.M. Expression profile of cyp1a1 and cyp1b1 enzymes in colon and bladder tumors. PLoS ONE 2013, 8, e82487. [Google Scholar] [CrossRef] [PubMed]
- Le Ferrec, E.; Lagadic-Gossmann, D.; Rauch, C.; Bardiau, C.; Maheo, K.; Massiere, F.; le Vee, M.; Guillouzo, A.; Morel, F. Transcriptional induction of cyp1a1 by oltipraz in human caco-2 cells is aryl hydrocarbon receptor- and calcium-dependent. J. Biol. Chem. 2002, 277, 24780–24787. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, L.M.; Li, H.; Li, L.; Lynch, C.; Xie, Y.; Nakanishi, T.; Ross, D.D.; Wang, H. A novel xenobiotic responsive element regulated by aryl hydrocarbon receptor is involved in the induction of BCRP/ABCG2 in LS174T cells. Biochem. Pharmacol. 2010, 80, 1754–1761. [Google Scholar] [CrossRef] [PubMed]
- Villard, P.H.; Caverni, S.; Baanannou, A.; Khalil, A.; Martin, P.G.; Penel, C.; Pineau, T.; Seree, E.; Barra, Y. PPARalpha transcriptionally induces ahr expression in Caco-2, but represses AhR pro-inflammatory effects. Biochem. Biophys. Res. Commun. 2007, 364, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Haarmann-Stemmann, T.; Bothe, H.; Abel, J. Growth factors, cytokines and their receptors as downstream targets of arylhydrocarbon receptor (AhR) signaling pathways. Biochem. Pharmacol. 2009, 77, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Puga, A.; Ma, C.; Marlowe, J.L. The aryl hydrocarbon receptor cross-talks with multiple signal transduction pathways. Biochem. Pharmacol. 2009, 77, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Duran, A.; Carvajal-Gonzalez, J.M.; Mulero-Navarro, S.; Santiago-Josefat, B.; Puga, A.; Fernandez-Salguero, P.M. Fitting a xenobiotic receptor into cell homeostasis: How the dioxin receptor interacts with tgfbeta signaling. Biochem. Pharmacol. 2009, 77, 700–712. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, S.K.; Smolenski, A. Phosphodiesterases link the aryl hydrocarbon receptor complex to cyclic nucleotide signaling. Biochem. Pharmacol. 2009, 77, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, J.C.; Gustafsson, J.A. Aryl hydrocarbon receptor-mediated signal transduction. Crit. Rev. Toxicol. 1997, 27, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Peng, Z.; Raufman, J.P. Src-mediated aryl hydrocarbon and epidermal growth factor receptor cross talk stimulates colon cancer cell proliferation. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1006–G1015. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, C.; Kaina, B. The aryl hydrocarbon receptor (AhR) in the regulation of cell-cell contact and tumor growth. Carcinogenesis 2010, 31, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, I.; MacDonald, T.T.; Pallone, F.; Monteleone, G. The aryl hydrocarbon receptor in inflammatory bowel disease: Linking the environment to disease pathogenesis. Curr. Opin. Gastroenterol. 2012, 28, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, I.; Rizzo, A.; Sarra, M.; Sica, G.; Sileri, P.; Biancone, L.; MacDonald, T.T.; Pallone, F.; Monteleone, G. Aryl hydrocarbon receptor-induced signals up-regulate il-22 production and inhibit inflammation in the gastrointestinal tract. Gastroenterology 2011, 141. [Google Scholar] [CrossRef] [PubMed]
- Furumatsu, K.; Nishiumi, S.; Kawano, Y.; Ooi, M.; Yoshie, T.; Shiomi, Y.; Kutsumi, H.; Ashida, H.; Fujii-Kuriyama, Y.; Azuma, T.; et al. A role of the aryl hydrocarbon receptor in attenuation of colitis. Dig. Dis. Sci. 2011, 56, 2532–2544. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Xu, C.; Sun, L.; Yu, M.; Peng, K.; Qiu, Y.; Xiao, W.; Yang, H. Aryl hydrocarbon receptor activation down-regulates IL-7 and reduces inflammation in a mouse model of DSS-induced colitis. Dig. Dis. Sci. 2015, 60, 1958–1066. [Google Scholar] [CrossRef] [PubMed]
- Ullman, T.A.; Itzkowitz, S.H. Intestinal inflammation and cancer. Gastroenterology 2011, 140, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Ikuta, T.; Kobayashi, Y.; Kitazawa, M.; Shiizaki, K.; Itano, N.; Noda, T.; Pettersson, S.; Poellinger, L.; Fujii-Kuriyama, Y.; Taniguchi, S.; et al. Asc-associated inflammation promotes cecal tumorigenesis in aryl hydrocarbon receptor-deficient mice. Carcinogenesis 2013, 34, 1620–1627. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Beischlag, T.V.; Vinciguerra, M.; Mazzoccoli, G. The circadian clock circuitry and the AhR signaling pathway in physiology and pathology. Biochem. Pharmacol. 2013, 85, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Mazzoccoli, G.; Vinciguerra, M.; Papa, G.; Piepoli, A. Circadian clock circuitry in colorectal cancer. World J. Gastroenterol. 2014, 20, 4197–4207. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xie, G.; Raufman, J.-P. Role of the Aryl Hydrocarbon Receptor in Colon Neoplasia. Cancers 2015, 7, 1436-1446. https://doi.org/10.3390/cancers7030847
AMA Style
Xie G, Raufman J-P. Role of the Aryl Hydrocarbon Receptor in Colon Neoplasia. Cancers. 2015; 7(3):1436-1446. https://doi.org/10.3390/cancers7030847
Chicago/Turabian StyleXie, Guofeng, and Jean-Pierre Raufman. 2015. "Role of the Aryl Hydrocarbon Receptor in Colon Neoplasia" Cancers 7, no. 3: 1436-1446. https://doi.org/10.3390/cancers7030847