Repositioning of Tyrosine Kinase Inhibitors as Antagonists of ATP-Binding Cassette Transporters in Anticancer Drug Resistance


Abstract
:1. Introduction
2. MDR and ABC Transporters
2.1. MDR
2.2. ABC Transporters
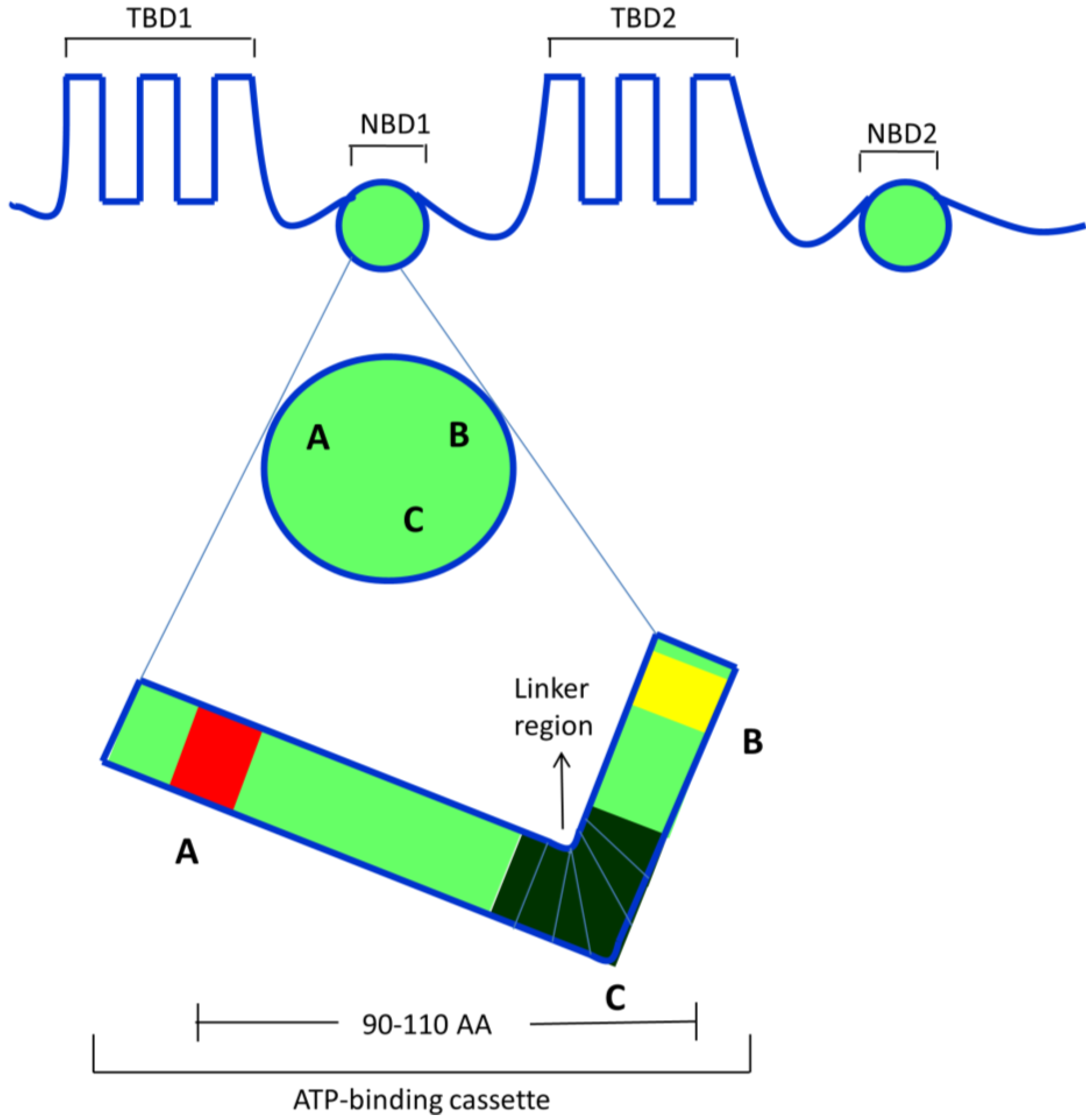
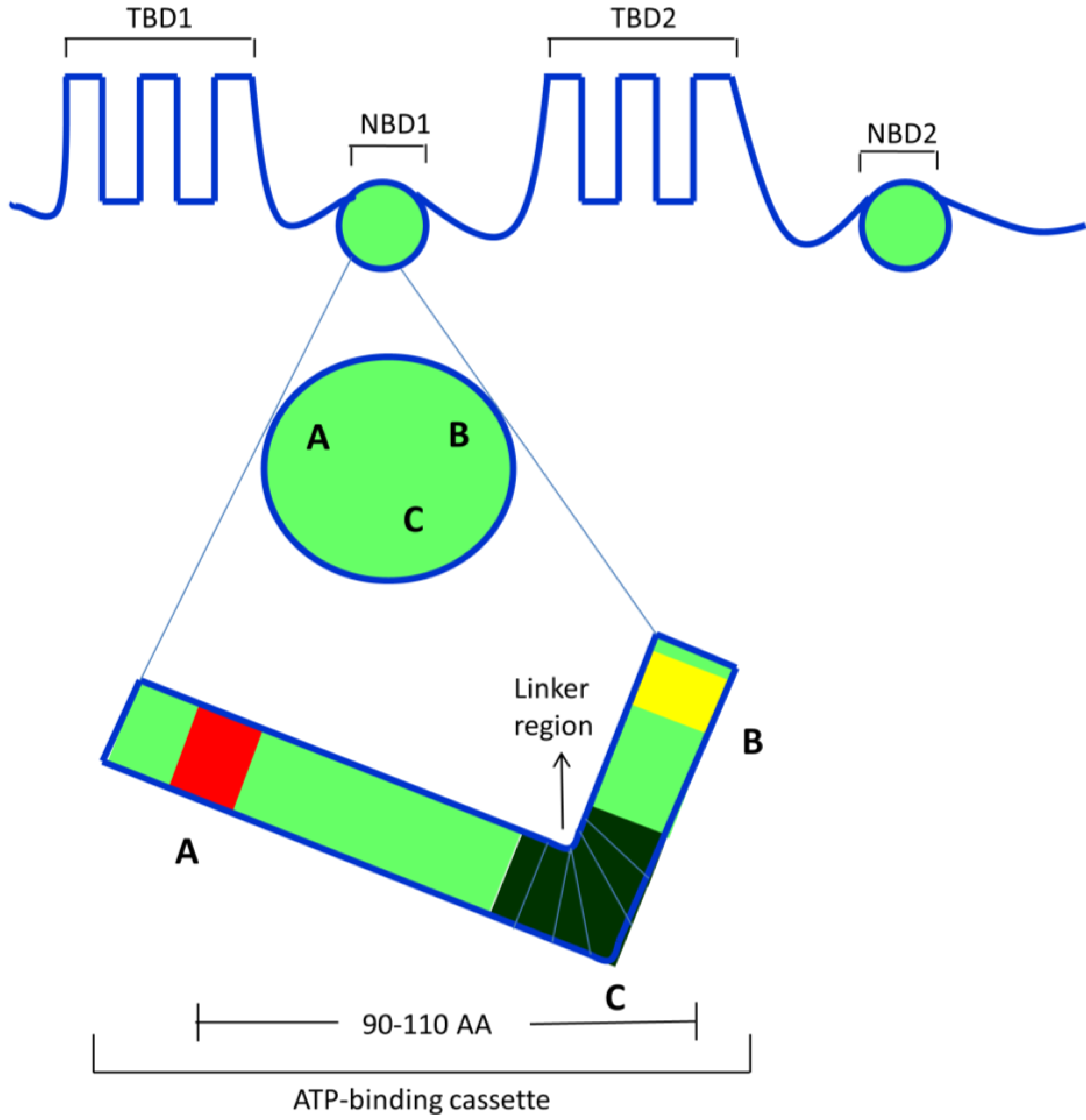
2.3. ABCB1/P-Glycoprotein (P-gp/MDR1)
2.4. ABCC/Multidrug Resistance Proteins (MRPs)
2.5. ABCG2/Breast Cancer Resistance Protein (BCRP)/Mitoxantrone Resistant Protein (MXR)
3. Reversal of MDR in Cancers
3.1. Strategies for Overcoming MDR in Cancer Cells

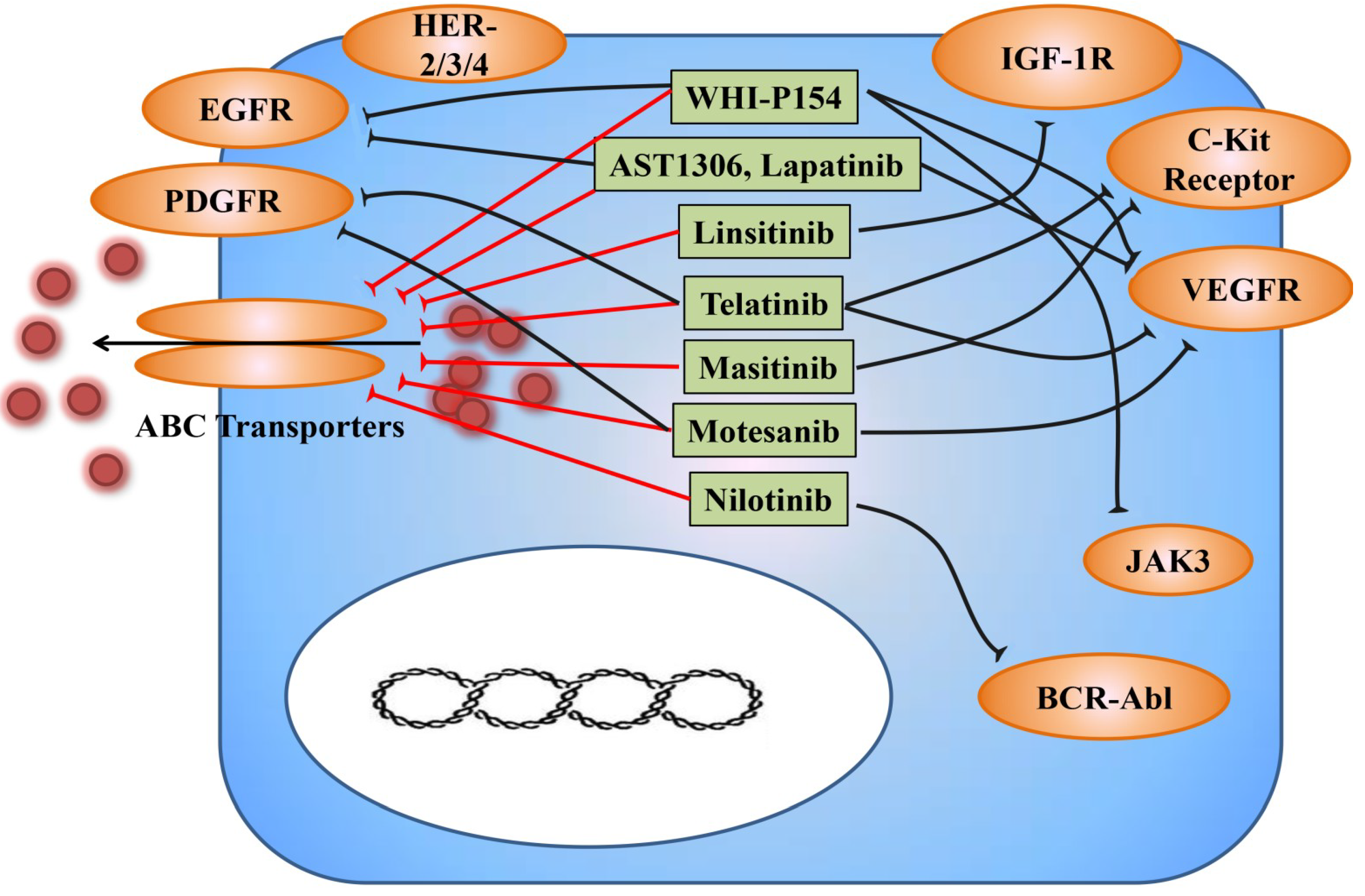
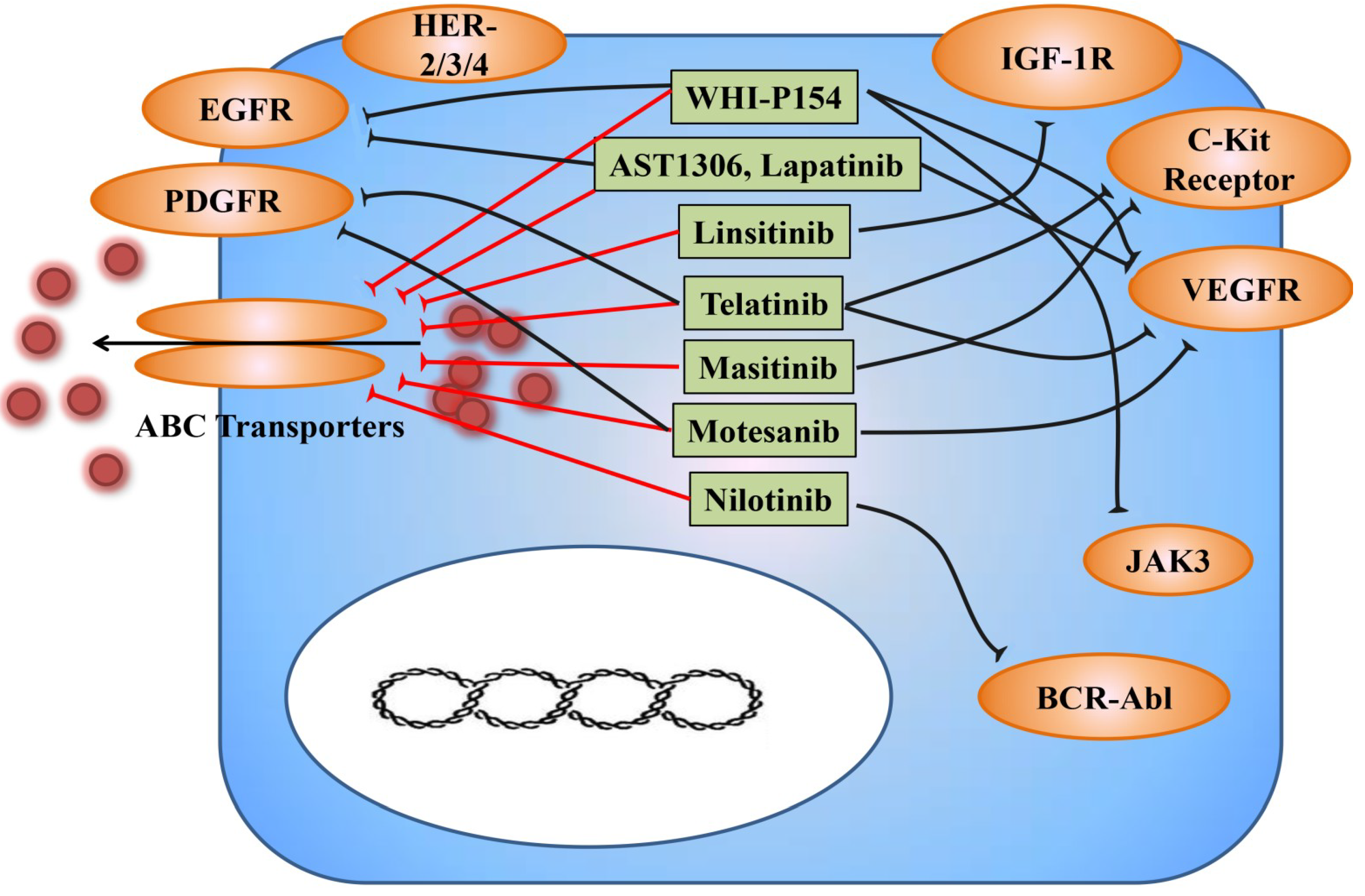
3.2. TKIs as Modulators for Anticancer Drug Resistance
3.2.1. AST1306

{kind=link}
{kind=link}
{kind=link}
| Compound | Concentrations (μM) | Targets | Targeted ABC Transporters | Effects on Targeted ABC Transporters Substrate | Effects on Targeted ABC Transporters Protein Expression |
|---|---|---|---|---|---|
| AST1306 | 0.25, 1 | EGFR and ErbB2 | ABCG2 | + | ↔ |
| Lapatinib | 0.625, 1.25, 2.5 | EGFR, HER-2, HER-3 and HER-4 | ABCB1 and ABCG2 | + | ↔ |
| Linsitinib (OSI-906) | 1, 2 | IGF-1R | ABCG2 and ABCC10 | + | ↔ |
| Masitinib | 1.25, 2.5 | c-Kit | ABCG2 and ABCC10 | + | ↔ |
| Motesanib (AMG-706) | 1, 3 | VEGFR-1/2/3, PDGFR and c-Kit | ABCB1 and ABCG2 | + | ↔ |
| Nilotinib (AMN-107) | 1, 2.5 | BCR-Abl | ABCB1, ABCG2 and ABCC10 | + | ↔ |
| Telatinib | 0.25, 0.5, 1 | VEGFR-2/3,PDGFR-β | ABCG2 | + | ↔ |
| WHI-P154 | 1, 4 | JAK3, EGFR, VEGFR | ABCG2 | + | ↔ |
3.2.2. Lapatinib
3.2.3. Linsitinib (OSI-906)
3.2.4. Masitinib
3.2.5. Motesanib (AMG-706)
3.2.6. Nilotinib (AMN-107)
3.2.7. Telatinib
3.2.8. WHI-P154
3.3. Effect of ABC Transporters Inhibition by TKIs on the Pharmacokinetics and Toxicity of Concomitantly Administered Anticancer Drugs
4. Clinical Relevance of ABC Transporters
4.1. ABCB1
4.2. ABCG2
4.3. ABCCs
5. Pharmacogenomics of ABC Transporters
6. Conclusions
Acknowledgements
Conflicts of Interest
References
- Cockerill, G.S.; Lackey, K.E. Small molecule inhibitors of the class 1 receptor tyrosine kinase family. Curr. Top. Med. Chem. 2002, 2, 1001–1010. [Google Scholar] [PubMed]
- Gottesman, M.M. Mechanisms of cancer drug resistance. Annu. Rev. Med. 2002, 53, 615–627. [Google Scholar] [PubMed]
- Quintieri, L.; Fantin, M.; Vizler, C. Identification of molecular determinants of tumor sensitivity and resistance to anticancer drugs. Adv. Exp. Med. Biol. 2007, 593, 95–104. [Google Scholar] [PubMed]
- Deeley, R.G.; Westlake, C.; Cole, S.P. Transmembrane transport of endo- and xenobiotics by mammalian ATP-binding cassette multidrug resistance proteins. Physiol. Rev. 2006, 86, 849–899. [Google Scholar] [PubMed]
- Jemal, A.; Murray, T.; Ward, E.; Samuels, A.; Tiwari, R.C.; Ghafoor, A.; Feuer, E.J.; Thun, M.J. Cancer statistics, 2005. CA Cancer. J. Clin. 2005, 55, 10–30. [Google Scholar] [PubMed]
- Mao, Q.; Unadkat, J.D. Role of the breast cancer resistance protein (ABCG2) in drug transport. AAPS J. 2005, 7, E118–E133. [Google Scholar]
- Ambudkar, S.V.; Kim, I.W.; Xia, D.; Sauna, Z.E. The A-Loop, a novel conserved aromatic acid subdomain upstream of the walker a motif in ABC transporters, is critical for ATP binding. FEBS Lett. 2006, 580, 1049–1055. [Google Scholar] [PubMed]
- Wu, C.P.; Hsieh, C.H.; Wu, Y.S. The emergence of drug transporter-mediated multidrug resistance to cancer chemotherapy. Mol. Pharm. 2011, 8, 1996–2011. [Google Scholar] [PubMed]
- Locher, K.P.; Borths, E. ABC Transporter architecture and mechanism: Implications from the crystal structures of BtuCD and BtuF. FEBS Lett. 2004, 564, 264–268. [Google Scholar] [PubMed]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar]
- Hyde, S.C.; Emsley, P.; Hartshorn, M.J.; Mimmack, M.M.; Gileadi, U.; Pearce, S.R.; Gallagher, M.P.; Gill, D.R.; Hubbard, R.E.; Higgins, C.F. Structural model of ATP-binding proteins associated with cystic fibrosis, multidrug resistance and bacterial transport. Nature 1990, 346, 362–365. [Google Scholar] [PubMed]
- Tiwari, A.K.; Sodani, K.; Dai, C.L.; Ashby, C.R., Jr.; Chen, Z.S. Revisiting the ABCs of multidrug resistance in cancer chemotherapy. Curr. Pharm. Biotechnol. 2011, 12, 570–594. [Google Scholar] [PubMed]
- Dean, M. ABC transporters, drug resistance, and cancer stem cells. J. Mammary Gland Biol. Neoplasia 2009, 14, 3–9. [Google Scholar] [PubMed]
- Dean, M.; Annilo, T. Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu. Rev. Genomics Hum. Genet. 2005, 6, 123–142. [Google Scholar] [PubMed]
- Juliano, R.L.; Ling, V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochim. Biophys. Acta 1976, 455, 152–162. [Google Scholar] [PubMed]
- Schinkel, A.H.; Mol, C.A.; Wagenaar, E.; van Deemter, L.; Smit, J.J.; Borst, P. Multidrug resistance and the role of P-Glycoprotein knockout mice. Eur. J. Cancer 1995, 31A, 1295–1298. [Google Scholar]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [PubMed]
- Sarkadi, B.; Homolya, L.; Szakacs, G.; Varadi, A. Human multidrug resistance ABCB and ABCG transporters: Participation in a chemoimmunity defense system. Physiol. Rev. 2006, 86, 1179–1236. [Google Scholar] [PubMed]
- Matsuo, K.; Eno, M.L.; Ahn, E.H.; Shahzad, M.M.; Im, D.D.; Rosenshein, N.B.; Sood, A.K. Multidrug resistance gene (MDR-1) and risk of brain metastasis in epithelial ovarian, fallopian tube, and peritoneal cancer. Am. J. Clin. Oncol. 2011, 34, 488–493. [Google Scholar] [PubMed]
- Eechoute, K.; Sparreboom, A.; Burger, H.; Franke, R.M.; Schiavon, G.; Verweij, J.; Loos, W.J.; Wiemer, E.A.; Mathijssen, R.H. Drug transporters and imatinib treatment: Implications for clinical practice. Clin. Cancer Res. 2011, 17, 406–415. [Google Scholar] [PubMed]
- Gao, Y.J.; Li, B.; Wu, X.Y.; Cui, J.; Han, J.K. Thyroid tumor-initiating cells: Increasing evidence and opportunities for anticancer therapy (Review). Oncol. Rep. 2014, 31, 1035–1042. [Google Scholar] [PubMed]
- Gao, B.; Russell, A.; Beesley, J.; Chen, X.Q.; Healey, S.; Henderson, M.; Wong, M.; Emmanuel, C.; Galletta, L.; Johnatty, S.E.; et al. Paclitaxel sensitivity in relation to ABCB1 expression, efflux and single nucleotide polymorphisms in ovarian cancer. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Lim, H.S.; Yoo, Y.K.; Shin, E.S.; Park, Y.H.; Lee, S.Y.; Lee, J.E.; Lee, D.H.; Kim, H.T.; Lee, J.S. Associations of ABCB1, ABCC2, and ABCG2 polymorphisms with irinotecan-pharmacokinetics and clinical outcome in patients with advanced non-small cell lung cancer. Cancer 2007, 110, 138–147. [Google Scholar] [PubMed]
- Leonard, G.D.; Fojo, T.; Bates, S.E. The role of ABC transporters in clinical practice. Oncologist 2003, 8, 411–424. [Google Scholar] [PubMed]
- Sodani, K.; Patel, A.; Kathawala, R.J.; Chen, Z.S. Multidrug resistance associated proteins in multidrug resistance. Chin. J. Cancer 2012, 31, 58–72. [Google Scholar] [PubMed]
- Kruh, G.D.; Belinsky, M.G. The MRP family of drug efflux pumps. Oncogene 2003, 22, 7537–7552. [Google Scholar] [PubMed]
- Norris, M.D.; Smith, J.; Tanabe, K.; Tobin, P.; Flemming, C.; Scheffer, G.L.; Wielinga, P.; Cohn, S.L.; London, W.B.; Marshall, G.M.; et al. Expression of multidrug transporter MRP4/ABCC4 is a marker of poor prognosis in neuroblastoma and confers resistance to irinotecan in vitro. Mol. Cancer. Ther. 2005, 4, 547–553. [Google Scholar] [PubMed]
- Toh, S.; Wada, M.; Uchiumi, T.; Inokuchi, A.; Makino, Y.; Horie, Y.; Adachi, Y.; Sakisaka, S.; Kuwano, M. Genomic structure of the canalicular multispecific organic anion-transporter gene (MRP2/cMOAT) and mutations in the ATP-binding-cassette region in Dubin-Johnson syndrome. Am. J. Hum. Genet. 1999, 64, 739–746. [Google Scholar] [PubMed]
- Ringpfeil, F.; Lebwohl, M.G.; Christiano, A.M.; Uitto, J. Pseudoxanthoma elasticum: Mutations in the MRP6 gene encoding a transmembrane ATP-binding cassette (ABC) transporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6001–6006. [Google Scholar] [PubMed]
- Hopper, E.; Belinsky, M.G.; Zeng, H.; Tosolini, A.; Testa, J.R.; Kruh, G.D. Analysis of the structure and expression pattern of MRP7 (ABCC10), a new member of the MRP subfamily. Cancer Lett. 2001, 162, 181–191. [Google Scholar] [PubMed]
- Takayanagi, S.; Kataoka, T.; Ohara, O.; Oishi, M.; Kuo, M.T.; Ishikawa, T. Human ATP-binding cassette transporter ABCC10: Expression profile and p53-dependent upregulation. J. Exp. Ther. Oncol. 2004, 4, 239–246. [Google Scholar] [PubMed]
- Hopper-Borge, E.A.; Churchill, T.; Paulose, C.; Nicolas, E.; Jacobs, J.D.; Ngo, O.; Kuang, Y.; Grinberg, A.; Westphal, H.; Chen, Z.S.; et al. Contribution of Abcc10 (Mrp7) to in vivo paclitaxel resistance as assessed in Abcc10(−/−) Mice. Cancer Res. 2011, 71, 3649–3657. [Google Scholar] [PubMed]
- Chen, Z.S.; Hopper-Borge, E.; Belinsky, M.G.; Shchaveleva, I.; Kotova, E.; Kruh, G.D. Characterization of the transport properties of human multidrug resistance protein 7 (MRP7, ABCC10). Mol. Pharmacol. 2003, 63, 351–358. [Google Scholar] [PubMed]
- Bessho, Y.; Oguri, T.; Ozasa, H.; Uemura, T.; Sakamoto, H.; Miyazaki, M.; Maeno, K.; Sato, S.; Ueda, R. ABCC10/MRP7 is associated with vinorelbine resistance in Non-small cell lung cancer. Oncol. Rep. 2009, 21, 263–268. [Google Scholar] [PubMed]
- Naramoto, H.; Uematsu, T.; Uchihashi, T.; Doto, R.; Matsuura, T.; Usui, Y.; Uematsu, S.; Li, X.; Takahashi, M.; Yamaoka, M.; et al. Multidrug resistance-Associated protein 7 expression is involved in cross-resistance to docetaxel in salivary gland adenocarcinoma cell lines. Int. J. Oncol. 2007, 30, 393–401. [Google Scholar] [PubMed]
- Dabrowska, M.; Sirotnak, F.M. Regulation of transcription of the human MRP7 gene: Characteristics of the basal promoter and identification of tumor-derived transcripts encoding additional 5' end heterogeneity. Gene 2004, 341, 129–139. [Google Scholar] [PubMed]
- Robey, R.W.; Polgar, O.; Deeken, J.; To, K.W.; Bates, S.E. ABCG2: Determining its relevance in clinical drug resistance. Cancer Metastasis Rev. 2007, 26, 39–57. [Google Scholar]
- Doyle, L.A.; Yang, W.; Abruzzo, L.V.; Krogmann, T.; Gao, Y.; Rishi, A.K.; Ross, D.D. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc. Natl. Acad. Sci. USA 1998, 95, 15665–15670. [Google Scholar] [PubMed]
- Miyake, K.; Mickley, L.; Litman, T.; Zhan, Z.; Robey, R.; Cristensen, B.; Brangi, M.; Greenberger, L.; Dean, M.; Fojo, T.; et al. Molecular cloning of cDNAs which are highly overexpressed in mitoxantrone-resistant cells: Demonstration of homology to ABC transport genes. Cancer Res. 1999, 59, 8–13. [Google Scholar] [PubMed]
- Litman, T.; Brangi, M.; Hudson, E.; Fetsch, P.; Abati, A.; Ross, D.D.; Miyake, K.; Resau, J.H.; Bates, S.E. The multidrug-resistant phenotype associated with overexpression of the new ABC half-transporter, MXR (ABCG2). J. Cell. Sci. 2000, 113, 2011–2021. [Google Scholar] [PubMed]
- Maliepaard, M.; van Gastelen, M.A.; Tohgo, A.; Hausheer, F.H.; van Waardenburg, R.C.; de Jong, L.A.; Pluim, D.; Beijnen, J.H.; Schellens, J.H. Circumvention of breast cancer resistance protein (BCRP)-mediated resistance to camptothecins in vitro using non-substrate drugs or the BCRP inhibitor GF120918. Clin. Cancer Res. 2001, 7, 935–941. [Google Scholar] [PubMed]
- Schinkel, A.H.; Jonker, J.W. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: An overview. Adv. Drug Deliv. Rev. 2003, 55, 3–29. [Google Scholar] [PubMed]
- Merino, G.; Alvarez, A.I.; Pulido, M.M.; Molina, A.J.; Schinkel, A.H.; Prieto, J.G. Breast cancer resistance protein (BCRP/ABCG2) transports fluoroquinolone antibiotics and affects their oral availability, pharmacokinetics, and milk secretion. Drug Metab. Dispos. 2006, 34, 690–695. [Google Scholar] [PubMed]
- Ejendal, K.F.; Hrycyna, C.A. Multidrug resistance and cancer: The role of the human ABC transporter ABCG2. Curr. Protein Pept. Sci. 2002, 3, 503–511. [Google Scholar] [PubMed]
- Krishnamurthy, P.; Schuetz, J.D. Role of ABCG2/BCRP in biology and medicine. Annu. Rev. Pharmacol. Toxicol. 2006, 46, 381–410. [Google Scholar]
- Sharom, F.J.; Yu, X.; Lu, P.; Liu, R.; Chu, J.W.; Szabo, K.; Muller, M.; Hose, C.D.; Monks, A.; Varadi, A.; et al. Interaction of the P-Glycoprotein multidrug transporter (MDR1) with high affinity peptide chemosensitizers in isolated membranes, reconstituted systems, and intact cells. Biochem. Pharmacol. 1999, 58, 571–586. [Google Scholar] [PubMed]
- Nie, S.; Xing, Y.; Kim, G.J.; Simons, J.W. Nanotechnology applications in cancer. Annu. Rev. Biomed. Eng. 2007, 9, 257–288. [Google Scholar] [PubMed]
- Basu, S.; Chaudhuri, P.; Sengupta, S. Targeting oncogenic signaling pathways by exploiting nanotechnology. Cell Cycle 2009, 8, 3480–3487. [Google Scholar]
- Xue, X.; Liang, X.J. Overcoming drug efflux-based multidrug resistance in cancer with nanotechnology. Chin. J. Cancer 2012, 31, 100–109. [Google Scholar] [PubMed]
- Shi, Z.; Peng, X.X.; Kim, I.W.; Shukla, S.; Si, Q.S.; Robey, R.W.; Bates, S.E.; Shen, T.; Ashby, C.R., Jr.; Fu, L.W.; et al. Erlotinib (Tarceva, OSI-774) antagonizes ATP-binding cassette subfamily B member 1 and ATP-binding cassette subfamily G member 2-mediated drug resistance. Cancer Res. 2007, 67, 11012–11020. [Google Scholar] [PubMed]
- Ludwig, J.A.; Szakacs, G.; Martin, S.E.; Chu, B.F.; Cardarelli, C.; Sauna, Z.E.; Caplen, N.J.; Fales, H.M.; Ambudkar, S.V.; Weinstein, J.N.; et al. Selective toxicity of NSC73306 in MDR1-positive cells as a new strategy to circumvent multidrug resistance in cancer. Cancer Res. 2006, 66, 4808–4815. [Google Scholar] [PubMed]
- Krause, D.S.; van Etten, R.A. Tyrosine kinases as targets for cancer therapy. N. Engl. J. Med. 2005, 353, 172–187. [Google Scholar] [PubMed]
- Faivre, S.; Djelloul, S.; Raymond, E. New paradigms in anticancer therapy: Targeting multiple signaling pathways with kinase inhibitors. Semin. Oncol. 2006, 33, 407–420. [Google Scholar] [PubMed]
- Luke, J.J.; Hodi, F.S. Vemurafenib and BRAF inhibition: A new class of treatment for metastatic melanoma. Clin. Cancer Res. 2012, 18, 9–14. [Google Scholar] [PubMed]
- Vispute, S.G.; Chen, J.J.; Sun, Y.L.; Sodani, K.S.; Singh, S.; Pan, Y.; Talele, T.; Ashby, C.R., Jr.; Chen, Z.S. Vemurafenib (plx4032, zelboraf®), a BRAF inhibitor, modulates ABCB1-, ABCG2-, and ABCC10-mediated multidrug resistance. J. Cancer Res. Updates 2013, 2, 306–317. [Google Scholar]
- Dai, C.L.; Tiwari, A.K.; Wu, C.P.; Su, X.D.; Wang, S.R.; Liu, D.G.; Ashby, C.R., Jr.; Huang, Y.; Robey, R.W.; Liang, Y.J.; et al. Lapatinib (Tykerb, GW572016) reverses multidrug resistance in cancer cells by inhibiting the activity of ATP-binding cassette subfamily B member 1 and G member 2. Cancer Res. 2008, 68, 7905–7914. [Google Scholar] [PubMed]
- Shen, T.; Kuang, Y.H.; Ashby, C.R.; Lei, Y.; Chen, A.; Zhou, Y.; Chen, X.; Tiwari, A.K.; Hopper-Borge, E.; Ouyang, J.; et al. Imatinib and nilotinib reverse multidrug resistance in cancer cells by inhibiting the efflux activity of the MRP7 (ABCC10). PLoS One 2009, 4, e7520. [Google Scholar]
- Tiwari, A.K.; Sodani, K.; Wang, S.R.; Kuang, Y.H.; Ashby, C.R., Jr.; Chen, X.; Chen, Z.S. Nilotinib (AMN107, Tasigna) reverses multidrug resistance by inhibiting the activity of the ABCB1/Pgp and ABCG2/BCRP/MXR transporters. Biochem. Pharmacol. 2009, 78, 153–161. [Google Scholar]
- Tiwari, A.K.; Sodani, K.; Dai, C.L.; Abuznait, A.H.; Singh, S.; Xiao, Z.J.; Patel, A.; Talele, T.T.; Fu, L.; Kaddoumi, A.; et al. Nilotinib potentiates anticancer drug sensitivity in murine ABCB1-, ABCG2-, and ABCC10-multidrug resistance xenograft models. Cancer Lett. 2013, 328, 307–317. [Google Scholar] [PubMed]
- Kathawala, R.J.; Sodani, K.; Chen, K.; Patel, A.; Abuznait, A.H.; Anreddy, N.; Sun, Y.L.; Kaddoumi, A.; Ashby, C.R., Jr.; Chen, Z.S. Masitinib antagonizes ATP-binding cassette subfamily C member 10-mediated paclitaxel resistance: A preclinical study. Mol. Cancer. Ther. 2014, 13, 714–723. [Google Scholar] [PubMed]
- Sodani, K.; Patel, A.; Anreddy, N.; Singh, S.; Yang, D.H.; Kathawala, R.J.; Kumar, P.; Talele, T.T.; Chen, Z.S. Telatinib reverses chemotherapeutic multidrug resistance mediated by ABCG2 efflux transporter in vitro and in vivo. Biochem. Pharmacol. 2014, 89, 52–61. [Google Scholar] [PubMed]
- Zhang, H.; Kathawala, R.J.; Wang, Y.J.; Zhang, Y.K.; Patel, A.; Shukla, S.; Robey, R.W.; Talele, T.T.; Ashby, C.R., Jr.; Ambudkar, S.V.; et al. Linsitinib (OSI-906) antagonizes ATP-binding cassette subfamily G member 2 and subfamily C member 10-mediated drug resistance. Int. J. Biochem. Cell Biol. 2014, 51, 111–119. [Google Scholar] [PubMed]
- Zhang, H.; Wang, Y.J.; Zhang, Y.K.; Wang, D.S.; Kathawala, R.J.; Patel, A.; Talele, T.T.; Chen, Z.S.; Fu, L.W. AST1306, a potent EGFR inhibitor, antagonizes ATP-binding cassette subfamily G member 2-mediated multidrug resistance. Cancer Lett. 2014, 350, 61–68. [Google Scholar] [PubMed]
- Zhang, H.; Zhang, Y.K.; Wang, Y.J.; Kathawala, R.J.; Patel, A.; Zhu, H.; Sodani, K.; Talele, T.T.; Ambudkar, S.V.; Chen, Z.S.; et al. WHI-P154 enhances the chemotherapeutic effect of anticancer agents in ABCG2-overexpressing cells. Cancer Sci. 2014, 105, 1071–1078. [Google Scholar] [PubMed]
- Wang, Y.J.; Kathawala, R.J.; Zhang, Y.K.; Patel, A.; Kumar, P.; Shukla, S.; Fung, K.L.; Ambudkar, S.V.; Talele, T.T.; Chen, Z.S. Motesanib (AMG706), a potent multikinase inhibitor, antagonizes multidrug resistance by inhibiting the efflux activity of the ABCB1. Biochem. Pharmacol. 2014, 90, 367–378. [Google Scholar] [PubMed]
- Aller, S.G.; Yu, J.; Ward, A.; Weng, Y.; Chittaboina, S.; Zhuo, R.; Harrell, P.M.; Trinh, Y.T.; Zhang, Q.; Urbatsch, I.L.; et al. Structure of P-Glycoprotein reveals a molecular basis for poly-specific drug binding. Science 2009, 323, 1718–1722. [Google Scholar] [PubMed]
- Nicolle, E.; Boumendjel, A.; Macalou, S.; Genoux, E.; Ahmed-Belkacem, A.; Carrupt, P.A.; di Pietro, A. QSAR analysis and molecular modeling of ABCG2-specific inhibitors. Adv. Drug Deliv. Rev. 2009, 61, 34–46. [Google Scholar] [PubMed]
- Crivori, P.; Reinach, B.; Pezzetta, D.; Poggesi, I. Computational models for identifying potential P-Glycoprotein substrates and inhibitors. Mol. Pharm. 2006, 3, 33–44. [Google Scholar] [PubMed]
- Pajeva, I.K.; Globisch, C.; Wiese, M. Combined pharmacophore modeling, docking, and 3D QSAR studies of ABCB1 and ABCC1 transporter inhibitors. ChemMedChem 2009, 4, 1883–1896. [Google Scholar] [PubMed]
- Xie, H.; Lin, L.; Tong, L.; Jiang, Y.; Zheng, M.; Chen, Z.; Jiang, X.; Zhang, X.; Ren, X.; Qu, W.; et al. AST1306, a novel irreversible inhibitor of the epidermal growth factor receptor 1 and 2, exhibits antitumor activity both in vitro and in vivo. PLoS One 2011, 6, e21487. [Google Scholar]
- Zhang, J.; Cao, J.; Li, J.; Zhang, Y.; Chen, Z.; Peng, W.; Sun, S.; Zhao, N.; Wang, J.; Zhong, D.; et al. A phase I study of AST1306, a novel irreversible EGFR and HER2 kinase inhibitor, in patients with advanced solid tumors. J. Hematol. Oncol. 2014, 7, 22. [Google Scholar] [PubMed]
- Normanno, N.; de Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; de Feo, G.; Caponigro, F.; Salomon, D.S. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene 2006, 366, 2–16. [Google Scholar] [PubMed]
- Mulvihill, M.J.; Cooke, A.; Rosenfeld-Franklin, M.; Buck, E.; Foreman, K.; Landfair, D.; O’Connor, M.; Pirritt, C.; Sun, Y.; Yao, Y.; et al. Discovery of OSI-906: A selective and orally efficacious dual inhibitor of the IGF-1 receptor and insulin receptor. Future Med. Chem. 2009, 1, 1153–1171. [Google Scholar] [PubMed]
- McKinley, E.T.; Bugaj, J.E.; Zhao, P.; Guleryuz, S.; Mantis, C.; Gokhale, P.C.; Wild, R.; Manning, H.C. 18FDG-PET Predicts pharmacodynamic response to OSI-906, a dual IGF-1R/IR inhibitor, in preclinical mouse models of lung cancer. Clin. Cancer Res. 2011, 17, 3332–3340. [Google Scholar] [PubMed]
- Le Cesne, A.; Blay, J.Y.; Bui, B.N.; Bouche, O.; Adenis, A.; Domont, J.; Cioffi, A.; Ray-Coquard, I.; Lassau, N.; Bonvalot, S.; et al. Phase II study of oral masitinib mesilate in imatinib-naive patients with locally advanced or metastatic gastro-intestinal stromal tumour (GIST). Eur. J. Cancer 2010, 46, 1344–1351. [Google Scholar] [PubMed]
- Mitry, E.; Hammel, P.; Deplanque, G.; Mornex, F.; Levy, P.; Seitz, J.F.; Moussy, A.; Kinet, J.P.; Hermine, O.; Rougier, P.; et al. Safety and activity of masitinib in combination with gemcitabine in patients with advanced pancreatic cancer. Cancer Chemother. Pharmacol. 2010, 66, 395–403. [Google Scholar]
- Georgin-Lavialle, S.; Lhermitte, L.; Suarez, F.; Yang, Y.; Letard, S.; Hanssens, K.; Feger, F.; Renand, A.; Brouze, C.; Canioni, D.; et al. Mast cell leukemia: Identification of a new C-Kit mutation, dup(501–502), and response to masitinib, a C-Kit tyrosine kinase inhibitor. Eur. J. Haematol. 2012, 89, 47–52. [Google Scholar] [PubMed]
- Kathawala, R.J.; Chen, J.J.; Zhang, Y.K.; Wang, Y.J.; Patel, A.; Wang, D.S.; Talele, T.T.; Ashby, C.R., Jr.; Chen, Z.S. Masitinib antagonizes ATP-binding cassette subfamily G member 2-mediated multidrug resistance. Int. J. Oncol. 2014, 44, 1634–1642. [Google Scholar] [PubMed]
- Polverino, A.; Coxon, A.; Starnes, C.; Diaz, Z.; DeMelfi, T.; Wang, L.; Bready, J.; Estrada, J.; Cattley, R.; Kaufman, S.; et al. AMG 706, an oral, multikinase inhibitor that selectively targets vascular endothelial growth factor, platelet-derived growth factor, and kit receptors, potently inhibits angiogenesis and induces regression in tumor xenografts. Cancer Res. 2006, 66, 8715–8721. [Google Scholar] [PubMed]
- Gora-Tybor, J.; Robak, T. Targeted drugs in chronic myeloid leukemia. Curr. Med. Chem. 2008, 15, 3036–3051. [Google Scholar] [PubMed]
- Shah, N.P.; Nicoll, J.M.; Nagar, B.; Gorre, M.E.; Paquette, R.L.; Kuriyan, J.; Sawyers, C.L. Multiple BCR-ABL kinase domain mutations confer polyclonal resistance to the tyrosine kinase inhibitor imatinib (STI571) in chronic phase and blast crisis chronic myeloid leukemia. Cancer Cell 2002, 2, 117–125. [Google Scholar] [PubMed]
- Langenberg, M.H.; Witteveen, P.O.; Roodhart, J.M.; Verheul, H.M.; Mergui-Roelvink, M.; van der Sar, J.; Brendel, E.; Laferriere, N.; Schellens, J.H.; Voest, E.E. Phase I evaluation of telatinib, a vascular endothelial growth factor receptor tyrosine kinase inhibitor, in combination with irinotecan and capecitabine in patients with advanced solid tumors. Clin. Cancer Res. 2010, 16, 2187–2197. [Google Scholar] [PubMed]
- Sudbeck, E.A.; Ghosh, S.; Liu, X.P.; Zheng, Y.; Myers, D.E.; Uckun, F.M. Tyrosine kinase inhibitors against EGF receptor-positive malignancies. Methods Mol. Biol. 2001, 166, 193–218. [Google Scholar] [PubMed]
- Marzec, M.; Kasprzycka, M.; Ptasznik, A.; Wlodarski, P.; Zhang, Q.; Odum, N.; Wasik, M.A. Inhibition of ALK enzymatic activity in T-cell lymphoma cells induces apoptosis and suppresses proliferation and STAT3 phosphorylation independently of Jak3. Lab. Invest. 2005, 85, 1544–1554. [Google Scholar]
- Stewart, C.F.; Leggas, M.; Schuetz, J.D.; Panetta, J.C.; Cheshire, P.J.; Peterson, J.; Daw, N.; Jenkins, J.J., 3rd; Gilbertson, R.; Germain, G.S.; et al. Gefitinib enhances the antitumor activity and oral bioavailability of irinotecan in mice. Cancer Res. 2004, 64, 7491–7499. [Google Scholar] [PubMed]
- Nakamura, Y.; Oka, M.; Soda, H.; Shiozawa, K.; Yoshikawa, M.; Itoh, A.; Ikegami, Y.; Tsurutani, J.; Nakatomi, K.; Kitazaki, T.; et al. Gefitinib (“Iressa”, ZD1839), an epidermal growth factor receptor tyrosine kinase inhibitor, reverses breast cancer resistance protein/ABCG2-mediated drug resistance. Cancer Res. 2005, 65, 1541–1546. [Google Scholar] [PubMed]
- Chen, K.G.; Sikic, B.I. Molecular pathways: Regulation and therapeutic implications of multidrug resistance. Clin. Cancer Res. 2012, 18, 1863–1869. [Google Scholar] [PubMed]
- Sikic, B.I.; Fisher, G.A.; Lum, B.L.; Halsey, J.; Beketic-Oreskovic, L.; Chen, G. Modulation and prevention of multidrug resistance by inhibitors of P-Glycoprotein. Cancer Chemother. Pharmacol. 1997, 40, S13–S19. [Google Scholar]
- Yahanda, A.M.; Alder, K.M.; Fisher, G.A.; Brophy, N.A.; Halsey, J.; Hardy, R.I.; Gosland, M.P.; Lum, B.L.; Sikic, B.I. Phase I trial of etoposide with cyclosporine as a modulator of multidrug resistance. J. Clin. Oncol. 1992, 10, 1624–1634. [Google Scholar] [PubMed]
- Lum, B.L.; Kaubisch, S.; Yahanda, A.M.; Adler, K.M.; Jew, L.; Ehsan, M.N.; Brophy, N.A.; Halsey, J.; Gosland, M.P.; Sikic, B.I. Alteration of etoposide pharmacokinetics and pharmacodynamics by cyclosporine in a phase I trial to modulate multidrug resistance. J. Clin. Oncol. 1992, 10, 1635–1642. [Google Scholar] [PubMed]
- Bartlett, N.L.; Lum, B.L.; Fisher, G.A.; Brophy, N.A.; Ehsan, M.N.; Halsey, J.; Sikic, B.I. Phase I trial of doxorubicin with cyclosporine as a modulator of multidrug resistance. J. Clin. Oncol. 1994, 12, 835–842. [Google Scholar] [PubMed]
- Advani, R.; Fisher, G.A.; Lum, B.L.; Hausdorff, J.; Halsey, J.; Litchman, M.; Sikic, B.I. A phase I trial of doxorubicin, paclitaxel, and valspodar (PSC 833), a modulator of multidrug resistance. Clin. Cancer Res. 2001, 7, 1221–1229. [Google Scholar] [PubMed]
- Baer, M.R.; George, S.L.; Dodge, R.K.; OʼLoughlin, K.L.; Minderman, H.; Caligiuri, M.A.; Anastasi, J.; Powell, B.L.; Kolitz, J.E.; Schiffer, C.A.; et al. Phase 3 study of the multidrug resistance modulator PSC-833 in previously untreated patients 60 years of age and older with acute myeloid leukemia: Cancer and leukemia group B study 9720. Blood 2002, 100, 1224–1232. [Google Scholar] [PubMed]
- List, A.F.; Kopecky, K.J.; Willman, C.L.; Head, D.R.; Persons, D.L.; Slovak, M.L.; Dorr, R.; Karanes, C.; Hynes, H.E.; Doroshow, J.H.; et al. Benefit of cyclosporine modulation of drug resistance in patients with poor-risk acute myeloid leukemia: A southwest oncology group study. Blood 2001, 98, 3212–3220. [Google Scholar] [PubMed]
- Liu Yin, J.A.; Wheatley, K.; Rees, J.K.; Burnett, A.K.; UK MRC Adult Leukemia Working Party. Comparison of “Sequential” versus “Standard” chemotherapy as re-induction treatment, with or without cyclosporine, in refractory/relapsed acute myeloid leukaemia (AML): Results of the UK medical research council AML-R trial. Br. J. Haematol. 2001, 113, 713–726. [Google Scholar] [PubMed]
- Cripe, L.D.; Uno, H.; Paietta, E.M.; Litzow, M.R.; Ketterling, R.P.; Bennett, J.M.; Rowe, J.M.; Lazarus, H.M.; Luger, S.; Tallman, M.S. Zosuquidar, a novel modulator of P-Glycoprotein, does not improve the outcome of older patients with newly diagnosed acute myeloid leukemia: A randomized, Placebo-controlled trial of the eastern Cooperative Oncology Group 3999. Blood 2010, 116, 4077–4085. [Google Scholar] [PubMed]
- Qadir, M.; O’Loughlin, K.L.; Fricke, S.M.; Williamson, N.A.; Greco, W.R.; Minderman, H.; Baer, M.R. Cyclosporin A is a broad-spectrum multidrug resistance modulator. Clin. Cancer Res. 2005, 11, 2320–2326. [Google Scholar] [PubMed]
- Matsubara, H.; Watanabe, M.; Imai, T.; Yui, Y.; Mizushima, Y.; Hiraumi, Y.; Kamitsuji, Y.; Watanabe, K.; Nishijo, K.; Toguchida, J.; et al. Involvement of extracellular signal-regulated kinase activation in human osteosarcoma cell resistance to the histone deacetylase inhibitor FK228 [(1S,4S,7Z,10S,16E,21R)-7-Ethylidene-4,21-Bis(Propan-2-Yl)-2-Oxa-12,13-Dithia-5,8,20,23-Tetraazabicyclo[8.7.6]Tricos-16-Ene-3,6,9,19,22-Pentone. J. Pharmacol. Exp. Ther. 2009, 328, 839–848. [Google Scholar] [PubMed]
- Tabe, Y.; Konopleva, M.; Contractor, R.; Munsell, M.; Schober, W.D.; Jin, L.; Tsutsumi-Ishii, Y.; Nagaoka, I.; Igari, J.; Andreeff, M. Up-regulation of MDR1 and induction of doxorubicin resistance by histone deacetylase inhibitor depsipeptide (FK228) and ATRA in acute promyelocytic leukemia cells. Blood 2006, 107, 1546–1554. [Google Scholar] [PubMed]
- Van den Heuvel-Eibrink, M.M.; Wiemer, E.A.; Prins, A.; Meijerink, J.P.; Vossebeld, P.J.; van der Holt, B.; Pieters, R.; Sonneveld, P. Increased expression of the breast cancer resistance protein (BCRP) in relapsed or refractory acute myeloid leukemia (AML). Leukemia 2002, 16, 833–839. [Google Scholar] [PubMed]
- Tang, S.C.; Lagas, J.S.; Lankheet, N.A.; Poller, B.; Hillebrand, M.J.; Rosing, H.; Beijnen, J.H.; Schinkel, A.H. Brain accumulation of sunitinib is restricted by P-Glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) and can be enhanced by oral elacridar and sunitinib coadministration. Int. J. Cancer 2012, 130, 223–233. [Google Scholar] [PubMed]
- Plasschaert, S.L.; van der Kolk, D.M.; de Bont, E.S.; Kamps, W.A.; Morisaki, K.; Bates, S.E.; Scheffer, G.L.; Scheper, R.J.; Vellenga, E.; de Vries, E.G. The role of breast cancer resistance protein in acute lymphoblastic leukemia. Clin. Cancer Res. 2003, 9, 5171–5177. [Google Scholar]
- Sauerbrey, A.; Sell, W.; Steinbach, D.; Voigt, A.; Zintl, F. Expression of the BCRP gene (ABCG2/MXR/ABCP) in childhood acute lymphoblastic leukaemia. Br. J. Haematol. 2002, 118, 147–150. [Google Scholar] [PubMed]
- Jordanides, N.E.; Jorgensen, H.G.; Holyoake, T.L.; Mountford, J.C. Functional ABCG2 is overexpressed on primary CML CD34+ cells and is Inhibited by imatinib mesylate. Blood 2006, 108, 1370–1373. [Google Scholar] [PubMed]
- Kim, J.E.; Singh, R.R.; Cho-Vega, J.H.; Drakos, E.; Davuluri, Y.; Khokhar, F.A.; Fayad, L.; Medeiros, L.J.; Vega, F. Sonic hedgehog signaling proteins and ATP-binding cassette G2 are aberrantly expressed in diffuse large B-Cell lymphoma. Mod. Pathol. 2009, 22, 1312–1320. [Google Scholar] [PubMed]
- Szczuraszek, K.; Materna, V.; Halon, A.; Mazur, G.; Wrobel, T.; Kuliczkowski, K.; Maciejczyk, A.; Zabel, M.; Drag, M.; Dietel, M.; et al. Positive correlation between cyclooxygenase-2 and ABC-transporter expression in non-hodgkin’s lymphomas. Oncol. Rep. 2009, 22, 1315–1323. [Google Scholar] [PubMed]
- Kim, Y.H.; Ishii, G.; Goto, K.; Ota, S.; Kubota, K.; Murata, Y.; Mishima, M.; Saijo, N.; Nishiwaki, Y.; Ochiai, A. Expression of breast cancer resistance protein is associated with a poor clinical outcome in patients with small-cell lung cancer. Lung Cancer 2009, 65, 105–111. [Google Scholar] [PubMed]
- Rijavec, M.; Silar, M.; Triller, N.; Kern, I.; Cegovnik, U.; Kosnik, M.; Korosec, P. Expressions of topoisomerase IIalpha and BCRP in metastatic cells are associated with overall survival in small cell lung cancer patients. Pathol. Oncol. Res. 2011, 17, 691–696. [Google Scholar] [PubMed]
- Yamada, A.; Ishikawa, T.; Ota, I.; Kimura, M.; Shimizu, D.; Tanabe, M.; Chishima, T.; Sasaki, T.; Ichikawa, Y.; Morita, S.; et al. High expression of ATP-binding cassette transporter ABCC11 in breast tumors is associated with aggressive subtypes and low disease-free survival. Breast Cancer Res. Treat. 2013, 137, 773–782. [Google Scholar] [PubMed]
- Hlavata, I.; Mohelnikova-Duchonova, B.; Vaclavikova, R.; Liska, V.; Pitule, P.; Novak, P.; Bruha, J.; Vycital, O.; Holubec, L.; Treska, V.; et al. The role of ABC transporters in progression and clinical outcome of colorectal cancer. Mutagenesis 2012, 27, 187–196. [Google Scholar] [PubMed]
- Kunicka, T.; Soucek, P. Importance of ABCC1 for cancer therapy and prognosis. Drug Metab. Rev. 2014, 46, 325–342. [Google Scholar] [PubMed]
- Young, L.C.; Campling, B.G.; Cole, S.P.; Deeley, R.G.; Gerlach, J.H. Multidrug resistance proteins MRP3, MRP1, and MRP2 in lung cancer: Correlation of protein levels with drug response and messenger RNA levels. Clin. Cancer Res. 2001, 7, 1798–1804. [Google Scholar] [PubMed]
- Zalcberg, J.; Hu, X.F.; Slater, A.; Parisot, J.; El-Osta, S.; Kantharidis, P.; Chou, S.T.; Parkin, J.D. MRP1 Not MDR1 Gene expression is the predominant mechanism of acquired multidrug resistance in two prostate carcinoma cell lines. Prostate Cancer Prostatic Dis. 2000, 3, 66–75. [Google Scholar] [PubMed]
- Chauhan, P.S.; Bhushan, B.; Singh, L.C.; Mishra, A.K.; Saluja, S.; Mittal, V.; Gupta, D.K.; Kapur, S. Expression of genes related to multiple drug resistance and apoptosis in acute leukemia: Response to induction chemotherapy. Exp. Mol. Pathol. 2012, 92, 44–49. [Google Scholar]
- Cui, Y.; Konig, J.; Buchholz, J.K.; Spring, H.; Leier, I.; Keppler, D. Drug resistance and ATP-dependent conjugate transport mediated by the apical multidrug resistance protein, MRP2, permanently expressed in human and canine cells. Mol. Pharmacol. 1999, 55, 929–937. [Google Scholar] [PubMed]
- Huisman, M.T.; Chhatta, A.A.; van Tellingen, O.; Beijnen, J.H.; Schinkel, A.H. MRP2 (ABCC2) transports taxanes and confers paclitaxel resistance and both processes are stimulated by probenecid. Int. J. Cancer 2005, 116, 824–829. [Google Scholar] [PubMed]
- Guminski, A.D.; Balleine, R.L.; Chiew, Y.E.; Webster, L.R.; Tapner, M.; Farrell, G.C.; Harnett, P.R.; Defazio, A. MRP2 (ABCC2) and cisplatin sensitivity in hepatocytes and human ovarian carcinoma. Gynecol. Oncol. 2006, 100, 239–246. [Google Scholar] [PubMed]
- Zelcer, N.; Saeki, T.; Reid, G.; Beijnen, J.H.; Borst, P. Characterization of drug transport by the human multidrug resistance protein 3 (ABCC3). J. Biol. Chem. 2001, 276, 46400–46407. [Google Scholar] [PubMed]
- Zeng, H.; Chen, Z.S.; Belinsky, M.G.; Rea, P.A.; Kruh, G.D. Transport of methotrexate (MTX) and folates by multidrug resistance protein (MRP) 3 and MRP1: Effect of polyglutamylation on MTX transport. Cancer Res. 2001, 61, 7225–7232. [Google Scholar] [PubMed]
- Chen, Z.S.; Lee, K.; Kruh, G.D. Transport of cyclic nucleotides and estradiol 17-Beta-D-Glucuronide by multidrug resistance protein 4. Resistance to 6-mercaptopurine and 6-thioguanine. J. Biol. Chem. 2001, 276, 33747–33754. [Google Scholar]
- Tian, Q.; Zhang, J.; Tan, T.M.; Chan, E.; Duan, W.; Chan, S.Y.; Boelsterli, U.A.; Ho, P.C.; Yang, H.; Bian, J.S.; et al. Human multidrug resistance associated protein 4 confers resistance to camptothecins. Pharm. Res. 2005, 22, 1837–1853. [Google Scholar]
- Pratt, S.; Shepard, R.L.; Kandasamy, R.A.; Johnston, P.A.; Perry, W., 3rd; Dantzig, A.H. The multidrug resistance protein 5 (ABCC5) confers resistance to 5-fluorouracil and transports its monophosphorylated metabolites. Mol. Cancer. Ther. 2005, 4, 855–863. [Google Scholar] [PubMed]
- Belinsky, M.G.; Chen, Z.S.; Shchaveleva, I.; Zeng, H.; Kruh, G.D. Characterization of the drug resistance and transport properties of multidrug resistance protein 6 (MRP6, ABCC6). Cancer Res. 2002, 62, 6172–6177. [Google Scholar]
- Wang, P.; Zhang, Z.; Gao, K.; Deng, Y.; Zhao, J.; Liu, B.; Li, X. Expression and clinical significance of ABCC10 in the patients with non-small cell lung cancer (in Chinese). Zhongguo Fei Ai Za Zhi 2009, 12, 875–878. [Google Scholar] [PubMed]
- Hu, S.; Niu, H.; Inaba, H.; Orwick, S.; Rose, C.; Panetta, J.C.; Yang, S.; Pounds, S.; Fan, Y.; Calabrese, C.; et al. Activity of the multikinase inhibitor sorafenib in combination with cytarabine in acute myeloid leukemia. J. Natl. Cancer Inst. 2011, 103, 893–905. [Google Scholar] [PubMed]
- Walle, T.; Walle, U.K.; Kumar, G.N.; Bhalla, K.N. Taxol metabolism and disposition in cancer patients. Drug Metab. Dispos. 1995, 23, 506–512. [Google Scholar] [PubMed]
- Sparreboom, A.; van Asperen, J.; Mayer, U.; Schinkel, A.H.; Smit, J.W.; Meijer, D.K.; Borst, P.; Nooijen, W.J.; Beijnen, J.H.; van Tellingen, O. Limited oral bioavailability and active epithelial excretion of paclitaxel (Taxol) caused by P-Glycoprotein in the intestine. Proc. Natl. Acad. Sci. USA 1997, 94, 2031–2035. [Google Scholar] [PubMed]
- Green, H.; Soderkvist, P.; Rosenberg, P.; Horvath, G.; Peterson, C. ABCB1 G1199A polymorphism and ovarian cancer response to paclitaxel. J. Pharm. Sci. 2008, 97, 2045–2048. [Google Scholar] [PubMed]
- Green, H.; Soderkvist, P.; Rosenberg, P.; Horvath, G.; Peterson, C. Mdr-1 single nucleotide polymorphisms in ovarian cancer tissue: G2677T/A correlates with response to paclitaxel chemotherapy. Clin. Cancer Res. 2006, 12, 854–859. [Google Scholar] [PubMed]
- Sissung, T.M.; Mross, K.; Steinberg, S.M.; Behringer, D.; Figg, W.D.; Sparreboom, A.; Mielke, S. Association of ABCB1 genotypes with paclitaxel-mediated peripheral neuropathy and neutropenia. Eur. J. Cancer 2006, 42, 2893–2896. [Google Scholar]
- Nakajima, M.; Fujiki, Y.; Kyo, S.; Kanaya, T.; Nakamura, M.; Maida, Y.; Tanaka, M.; Inoue, M.; Yokoi, T. Pharmacokinetics of paclitaxel in ovarian cancer patients and genetic polymorphisms of CYP2C8, CYP3A4, and MDR1. J. Clin. Pharmacol. 2005, 45, 674–682. [Google Scholar] [PubMed]
- Henningsson, A.; Marsh, S.; Loos, W.J.; Karlsson, M.O.; Garsa, A.; Mross, K.; Mielke, S.; Vigano, L.; Locatelli, A.; Verweij, J.; et al. Association of CYP2C8, CYP3A4, CYP3A5, and ABCB1 polymorphisms with the pharmacokinetics of paclitaxel. Clin. Cancer Res. 2005, 11, 8097–8104. [Google Scholar] [PubMed]
- Grimm, C.; Polterauer, S.; Zeillinger, R.; Tong, D.; Heinze, G.; Wolf, A.; Natter, C.; Reinthaller, A.; Hefler, L.A. Two multidrug-resistance (ABCB1) gene polymorphisms as prognostic parameters in women with ovarian cancer. Anticancer Res. 2010, 30, 3487–3491. [Google Scholar] [PubMed]
- Marsh, S.; Paul, J.; King, C.R.; Gifford, G.; McLeod, H.L.; Brown, R. Pharmacogenetic assessment of toxicity and outcome after platinum plus taxane chemotherapy in ovarian cancer: The Scottish randomised trial in ovarian cancer. J. Clin. Oncol. 2007, 25, 4528–4535. [Google Scholar] [PubMed]
- Wojnowski, L.; Kulle, B.; Schirmer, M.; Schluter, G.; Schmidt, A.; Rosenberger, A.; Vonhof, S.; Bickeboller, H.; Toliat, M.R.; Suk, E.K.; et al. NAD(P)H oxidase and multidrug resistance protein genetic polymorphisms are associated with doxorubicin-induced cardiotoxicity. Circulation 2005, 112, 3754–3762. [Google Scholar] [PubMed]
- Colombo, S.; Soranzo, N.; Rotger, M.; Sprenger, R.; Bleiber, G.; Furrer, H.; Buclin, T.; Goldstein, D.; Decosterd, L.; Telenti, A.; et al. Influence of ABCB1, ABCC1, ABCC2, and ABCG2 haplotypes on the cellular exposure of nelfinavir in vivo. Pharmacogenet. Genomics 2005, 15, 599–608. [Google Scholar] [PubMed]
- Kartenbeck, J.; Leuschner, U.; Mayer, R.; Keppler, D. Absence of the canalicular isoform of the MRP gene-encoded conjugate export pump from the hepatocytes in dubin-johnson syndrome. Hepatology 1996, 23, 1061–1066. [Google Scholar] [PubMed]
- Gradhand, U.; Kim, R.B. Pharmacogenomics of MRP transporters (ABCC1–5) and BCRP (ABCG2). Drug Metab. Rev. 2008, 40, 317–354. [Google Scholar] [PubMed]
- Kato, T.; Hamada, A.; Mori, S.; Saito, H. Genetic polymorphisms in metabolic and cellular transport pathway of methotrexate impact clinical outcome of methotrexate monotherapy in Japanese patients with rheumatoid arthritis. Drug Metab. Pharmacokinet. 2012, 27, 192–199. [Google Scholar] [PubMed]
- Pushpakom, S.P.; Liptrott, N.J.; Rodriguez-Novoa, S.; Labarga, P.; Soriano, V.; Albalater, M.; Hopper-Borge, E.; Bonora, S.; di Perri, G.; Back, D.J.; et al. Genetic variants of ABCC10, a novel tenofovir transporter, are associated with kidney tubular dysfunction. J. Infect. Dis. 2011, 204, 145–153. [Google Scholar] [PubMed]
- Liptrott, N.J.; Pushpakom, S.; Wyen, C.; Fatkenheuer, G.; Hoffmann, C.; Mauss, S.; Knechten, H.; Brockmeyer, N.H.; Hopper-Borge, E.; Siccardi, M.; et al. Association of ABCC10 polymorphisms with nevirapine plasma concentrations in the german competence network for HIV/AIDS. Pharmacogenet. Genomics 2012, 22, 10–19. [Google Scholar] [PubMed]
- Ciccacci, C.; di Fusco, D.; Marazzi, M.C.; Liotta, G.; Palombi, L.; Novelli, G.; Borgiani, P. ABCC10 rs2125739 polymorphism and nevirapine-induced hepatotoxicity: Lack of association in a population from mozambique. Pharmacogenet. Genomics 2013, 23, 38–39. [Google Scholar]
- Cusatis, G.; Sparreboom, A. Pharmacogenomic importance of ABCG2. Pharmacogenomics 2008, 9, 1005–1009. [Google Scholar] [PubMed]
- Cusatis, G.; Gregorc, V.; Li, J.; Spreafico, A.; Ingersoll, R.G.; Verweij, J.; Ludovini, V.; Villa, E.; Hidalgo, M.; Sparreboom, A.; et al. Pharmacogenetics of ABCG2 and adverse reactions to gefitinib. J. Natl. Cancer Inst. 2006, 98, 1739–1742. [Google Scholar] [PubMed]
- Zamboni, W.C.; Ramanathan, R.K.; McLeod, H.L.; Mani, S.; Potter, D.M.; Strychor, S.; Maruca, L.J.; King, C.R.; Jung, L.L.; Parise, R.A.; et al. Disposition of 9-nitrocamptothecin and its 9-aminocamptothecin metabolite in relation to ABC transporter genotypes. Invest. New Drugs 2006, 24, 393–401. [Google Scholar]
- Sparreboom, A.; Gelderblom, H.; Marsh, S.; Ahluwalia, R.; Obach, R.; Principe, P.; Twelves, C.; Verweij, J.; McLeod, H.L. Diflomotecan pharmacokinetics in relation to ABCG2 421C > A genotype. Clin. Pharmacol. Ther. 2004, 76, 38–44. [Google Scholar] [PubMed]
- Zhou, Q.; Sparreboom, A.; Tan, E.H.; Cheung, Y.B.; Lee, A.; Poon, D.; Lee, E.J.; Chowbay, B. Pharmacogenetic profiling across the irinotecan pathway in asian patients with cancer. Br. J. Clin. Pharmacol. 2005, 59, 415–424. [Google Scholar] [PubMed]
- Zhang, W.; Yu, B.N.; He, Y.J.; Fan, L.; Li, Q.; Liu, Z.Q.; Wang, A.; Liu, Y.L.; Tan, Z.R.; Jiang, F.; et al. Role of BCRP 421C > A polymorphism on rosuvastatin pharmacokinetics in healthy Chinese males. Clin. Chim. Acta 2006, 373, 99–103. [Google Scholar] [PubMed]
- Urquhart, B.L.; Ware, J.A.; Tirona, R.G.; Ho, R.H.; Leake, B.F.; Schwarz, U.I.; Zaher, H.; Palandra, J.; Gregor, J.C.; Dresser, G.K.; et al. Breast cancer resistance protein (ABCG2) and drug disposition: Intestinal expression, polymorphisms and sulfasalazine as an in vivo probe. Pharmacogenet. Genomics 2008, 18, 439–448. [Google Scholar] [PubMed]
- Yamasaki, Y.; Ieiri, I.; Kusuhara, H.; Sasaki, T.; Kimura, M.; Tabuchi, H.; Ando, Y.; Irie, S.; Ware, J.; Nakai, Y.; et al. Pharmacogenetic characterization of sulfasalazine disposition based on NAT2 and ABCG2 (BCRP) gene polymorphisms in humans. Clin. Pharmacol. Ther. 2008, 84, 95–103. [Google Scholar] [PubMed]
- Sparreboom, A.; Loos, W.J.; Burger, H.; Sissung, T.M.; Verweij, J.; Figg, W.D.; Nooter, K.; Gelderblom, H. Effect of ABCG2 genotype on the oral bioavailability of topotecan. Cancer Biol. Ther. 2005, 4, 650–658. [Google Scholar]
- Lal, S.; Wong, Z.W.; Sandanaraj, E.; Xiang, X.; Ang, P.C.; Lee, E.J.; Chowbay, B. Influence of ABCB1 and ABCG2 polymorphisms on doxorubicin disposition in Asian breast cancer patients. Cancer Sci. 2008, 99, 816–823. [Google Scholar] [PubMed]
- Gardner, E.R.; Burger, H.; van Schaik, R.H.; van Oosterom, A.T.; de Bruijn, E.A.; Guetens, G.; Prenen, H.; de Jong, F.A.; Baker, S.D.; Bates, S.E.; et al. Association of enzyme and transporter genotypes with the pharmacokinetics of imatinib. Clin. Pharmacol. Ther. 2006, 80, 192–201. [Google Scholar] [PubMed]
- Ieiri, I.; Suwannakul, S.; Maeda, K.; Uchimaru, H.; Hashimoto, K.; Kimura, M.; Fujino, H.; Hirano, M.; Kusuhara, H.; Irie, S.; et al. SLCO1B1 (OATP1B1, an uptake transporter) and ABCG2 (BCRP, an efflux transporter) variant alleles and pharmacokinetics of pitavastatin in healthy volunteers. Clin. Pharmacol. Ther. 2007, 82, 541–547. [Google Scholar] [PubMed]
- Yoshioka, S.; Katayama, K.; Okawa, C.; Takahashi, S.; Tsukahara, S.; Mitsuhashi, J.; Sugimoto, Y. The identification of two germ-line mutations in the human breast cancer resistance protein gene that result in the expression of a low/non-functional protein. Pharm. Res. 2007, 24, 1108–1117. [Google Scholar] [PubMed]
- Rudin, C.M.; Liu, W.; Desai, A.; Karrison, T.; Jiang, X.; Janisch, L.; Das, S.; Ramirez, J.; Poonkuzhali, B.; Schuetz, E.; et al. Pharmacogenomic and pharmacokinetic determinants of erlotinib toxicity. J. Clin. Oncol. 2008, 26, 1119–1127. [Google Scholar] [PubMed]
- Poonkuzhali, B.; Lamba, J.; Strom, S.; Sparreboom, A.; Thummel, K.; Watkins, P.; Schuetz, E. Association of breast cancer resistance protein/ABCG2 phenotypes and novel promoter and intron 1 single nucleotide polymorphisms. Drug Metab. Dispos. 2008, 36, 780–795. [Google Scholar] [PubMed]
- Innocenti, F.; Kroetz, D.L.; Schuetz, E.; Dolan, M.E.; Ramirez, J.; Relling, M.; Chen, P.; Das, S.; Rosner, G.L.; Ratain, M.J. Comprehensive pharmacogenetic analysis of irinotecan neutropenia and pharmacokinetics. J. Clin. Oncol. 2009, 27, 2604–2614. [Google Scholar]
- Cecchin, E.; D’Andrea, M.; Lonardi, S.; Zanusso, C.; Pella, N.; Errante, D.; de Mattia, E.; Polesel, J.; Innocenti, F.; Toffoli, G. A prospective validation pharmacogenomic study in the adjuvant setting of colorectal cancer patients treated with the 5-Fluorouracil/Leucovorin/Oxaliplatin (FOLFOX4) regimen. Pharmacogenomics J. 2013, 13, 403–409. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-J.; Zhang, Y.-K.; Kathawala, R.J.; Chen, Z.-S. Repositioning of Tyrosine Kinase Inhibitors as Antagonists of ATP-Binding Cassette Transporters in Anticancer Drug Resistance. Cancers 2014, 6, 1925-1952. https://doi.org/10.3390/cancers6041925
Wang Y-J, Zhang Y-K, Kathawala RJ, Chen Z-S. Repositioning of Tyrosine Kinase Inhibitors as Antagonists of ATP-Binding Cassette Transporters in Anticancer Drug Resistance. Cancers. 2014; 6(4):1925-1952. https://doi.org/10.3390/cancers6041925
Chicago/Turabian StyleWang, Yi-Jun, Yun-Kai Zhang, Rishil J. Kathawala, and Zhe-Sheng Chen. 2014. "Repositioning of Tyrosine Kinase Inhibitors as Antagonists of ATP-Binding Cassette Transporters in Anticancer Drug Resistance" Cancers 6, no. 4: 1925-1952. https://doi.org/10.3390/cancers6041925