
Cancer Therapy by Catechins Involves Redox Cycling of Copper Ions and Generation of Reactive Oxygen Species

 ,
,  , ,
, ,
Abstract
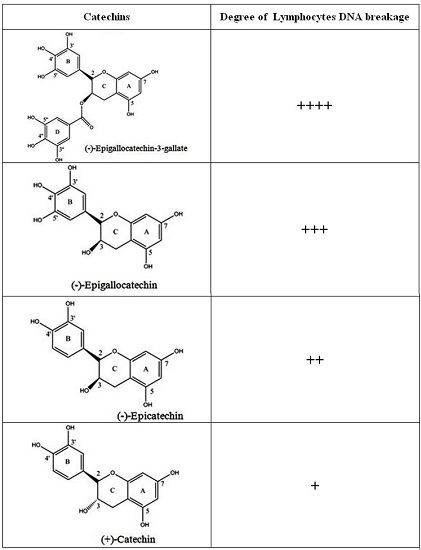
:
1. Introduction
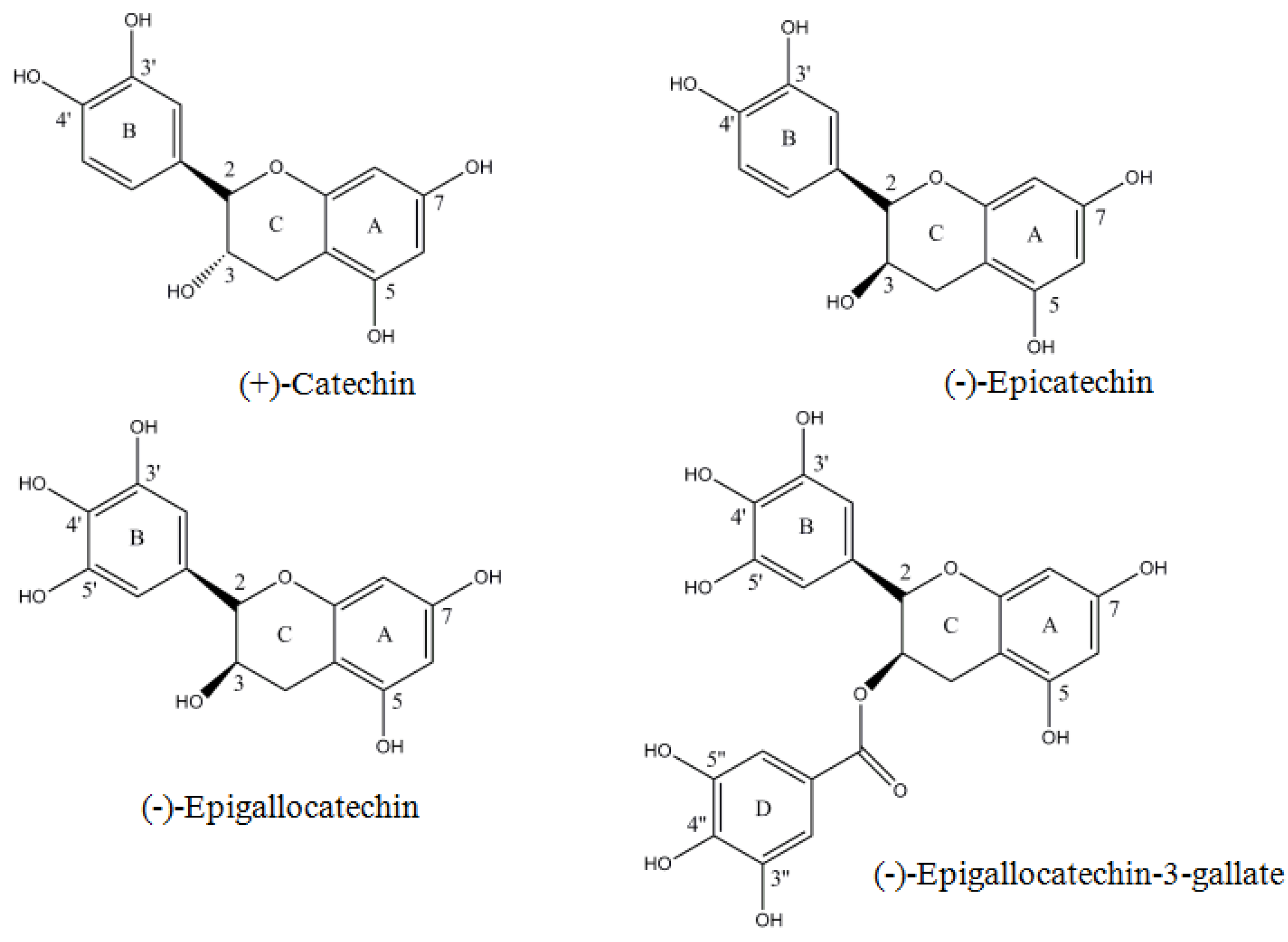
2. Results
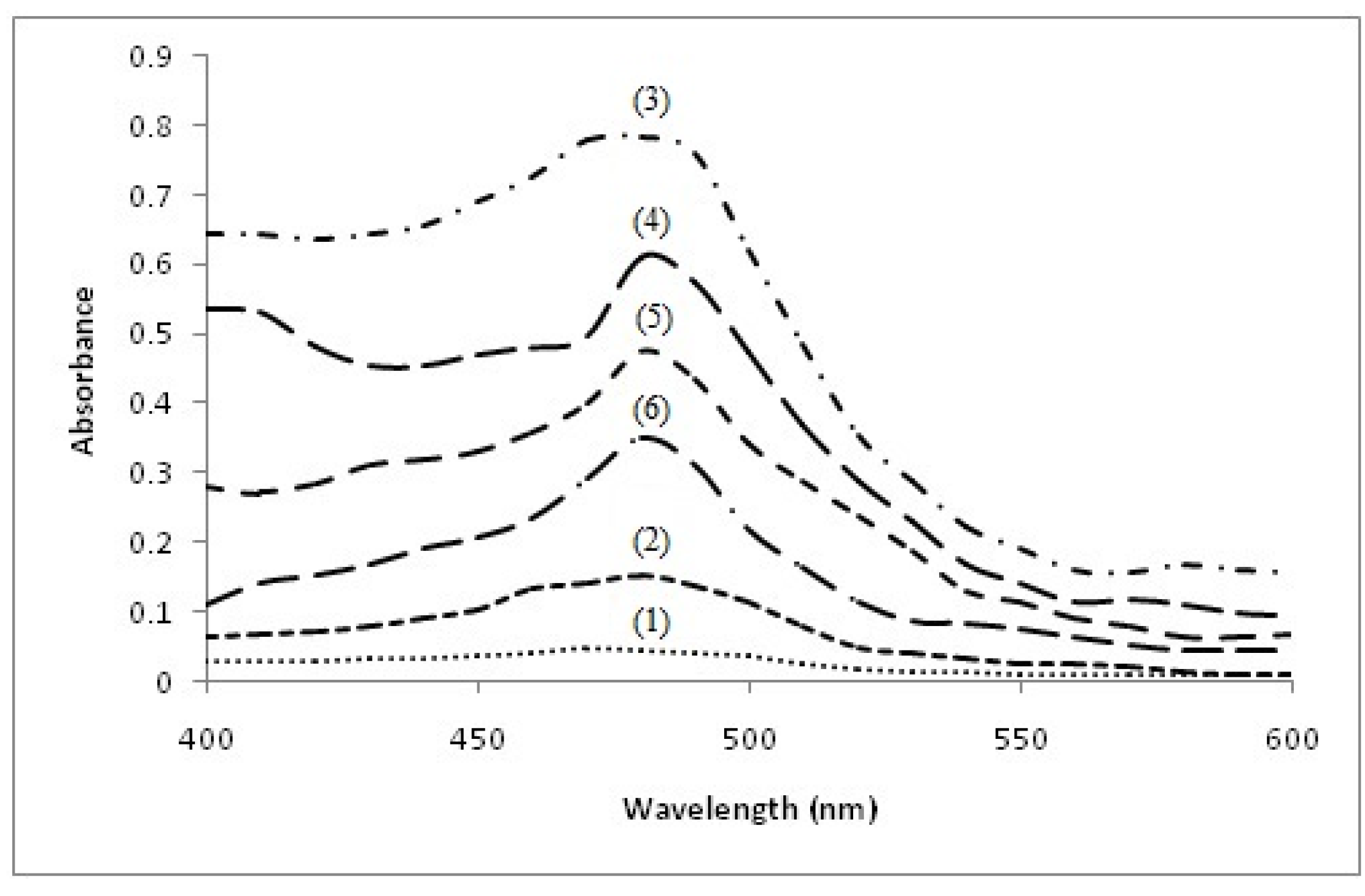
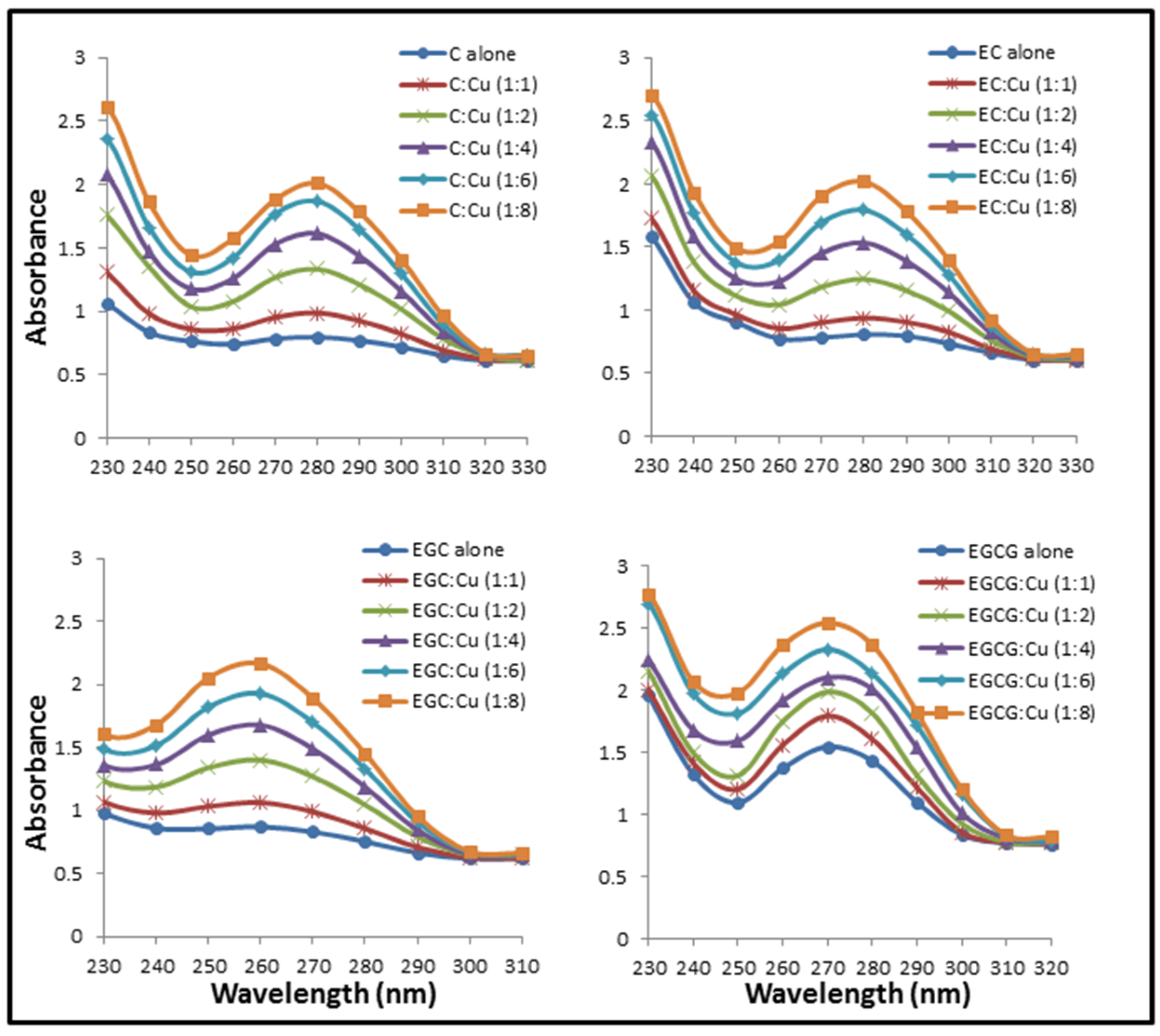
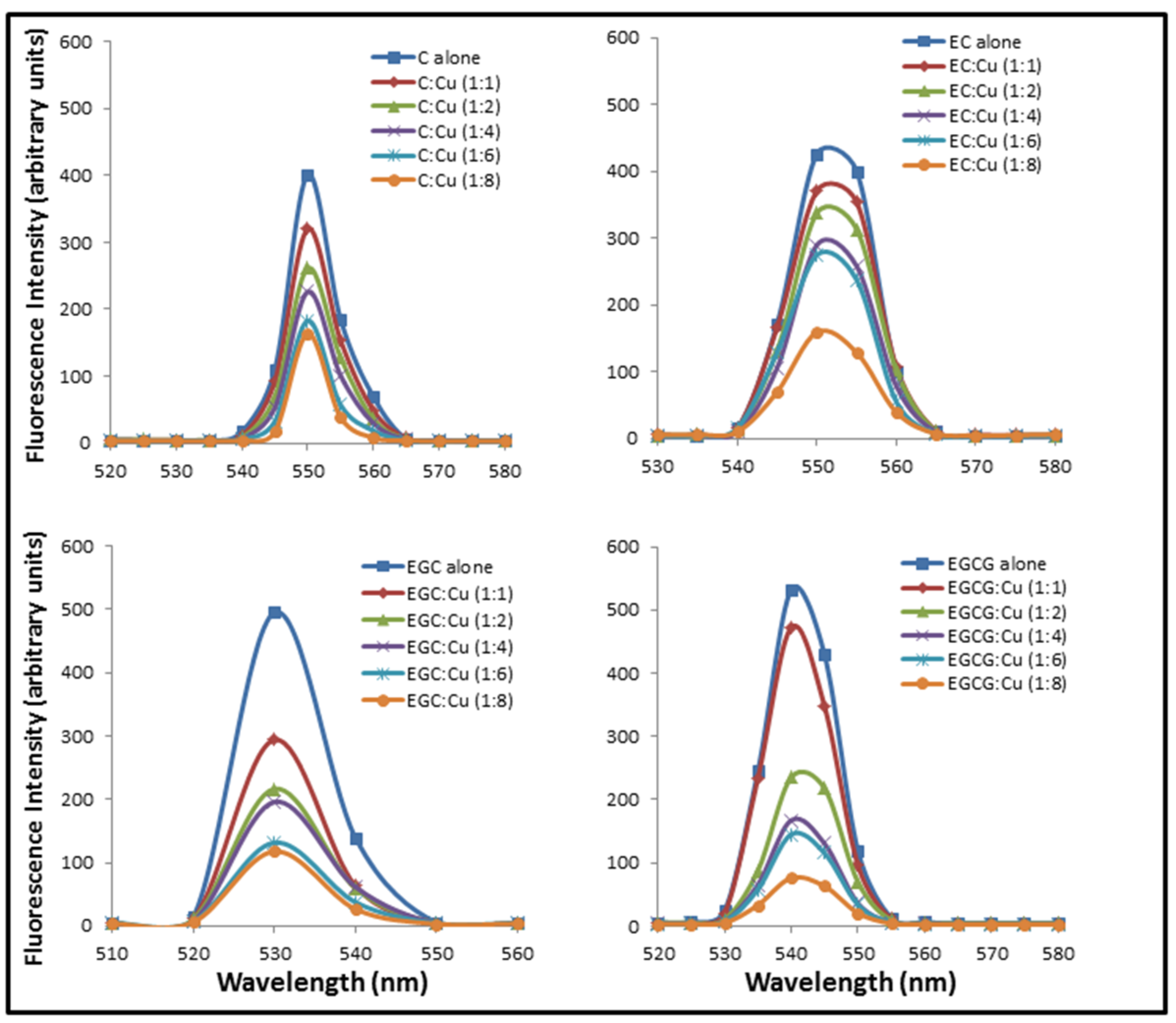
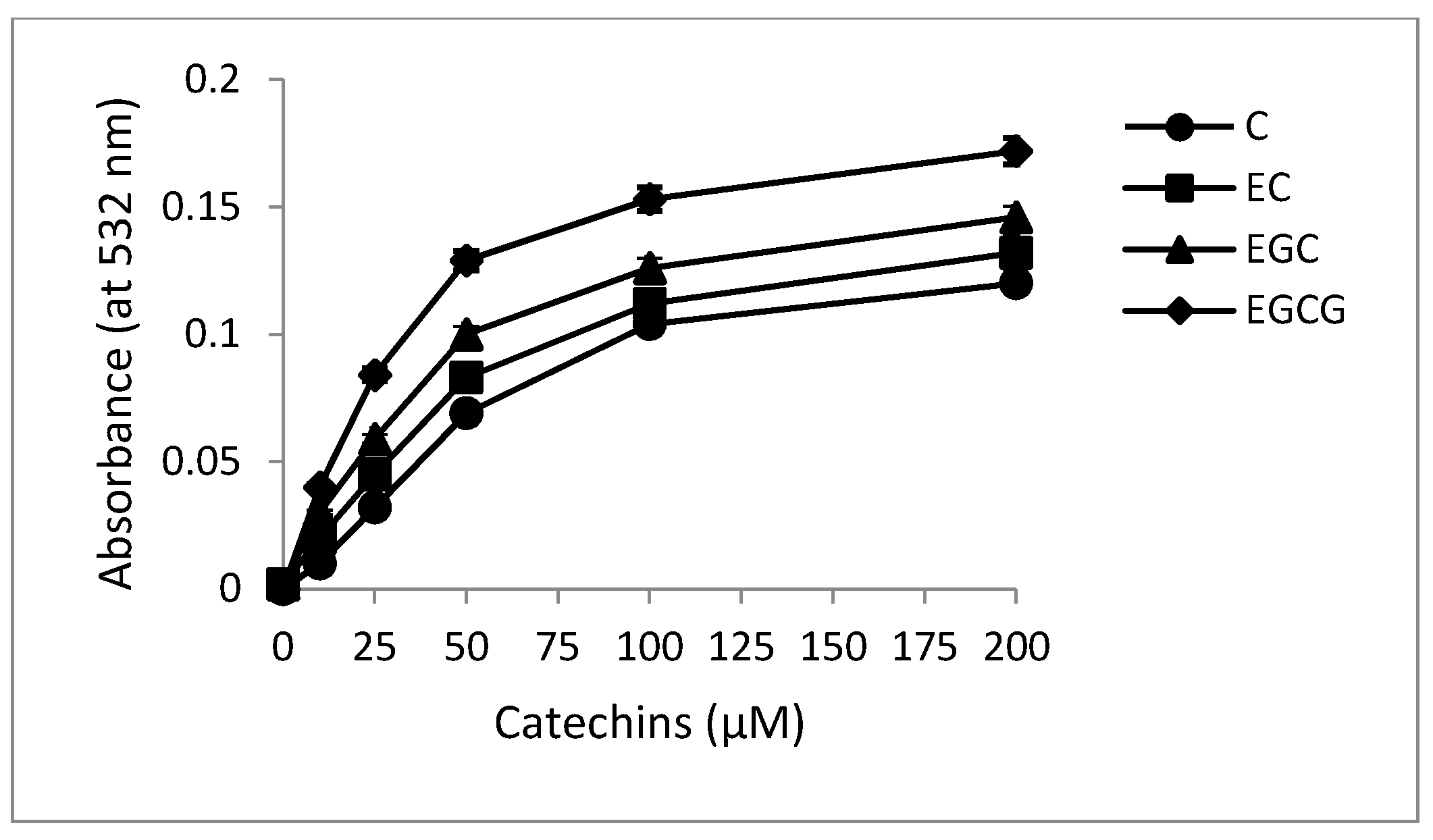
2.1. Formation of Catechins-Cu(II) Complex

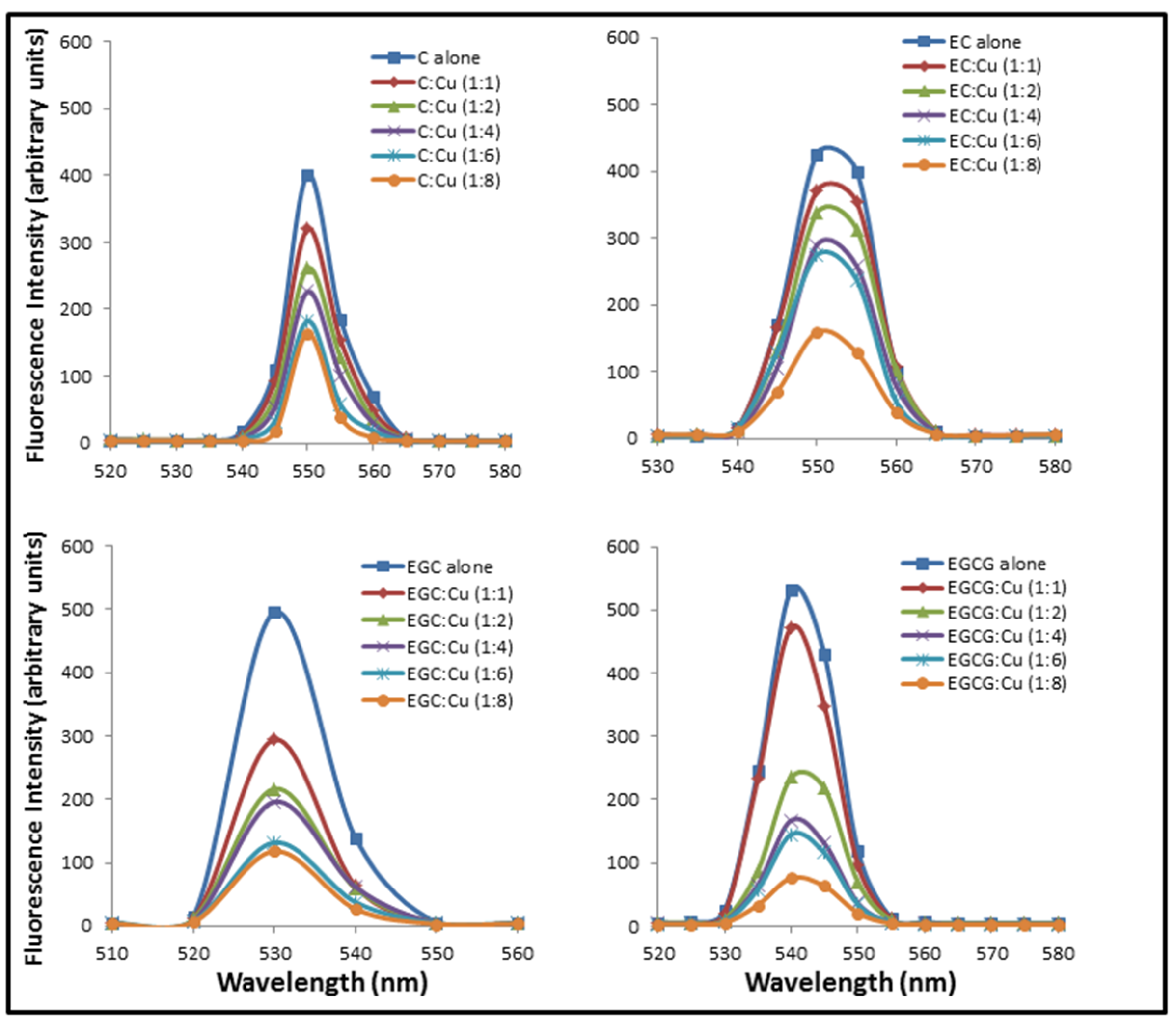
2.2. Formation of Complexes Involving Cu(II) with Catechins
2.3. Detection of Catechins Induced Cu(I) Production by Bathocuproine
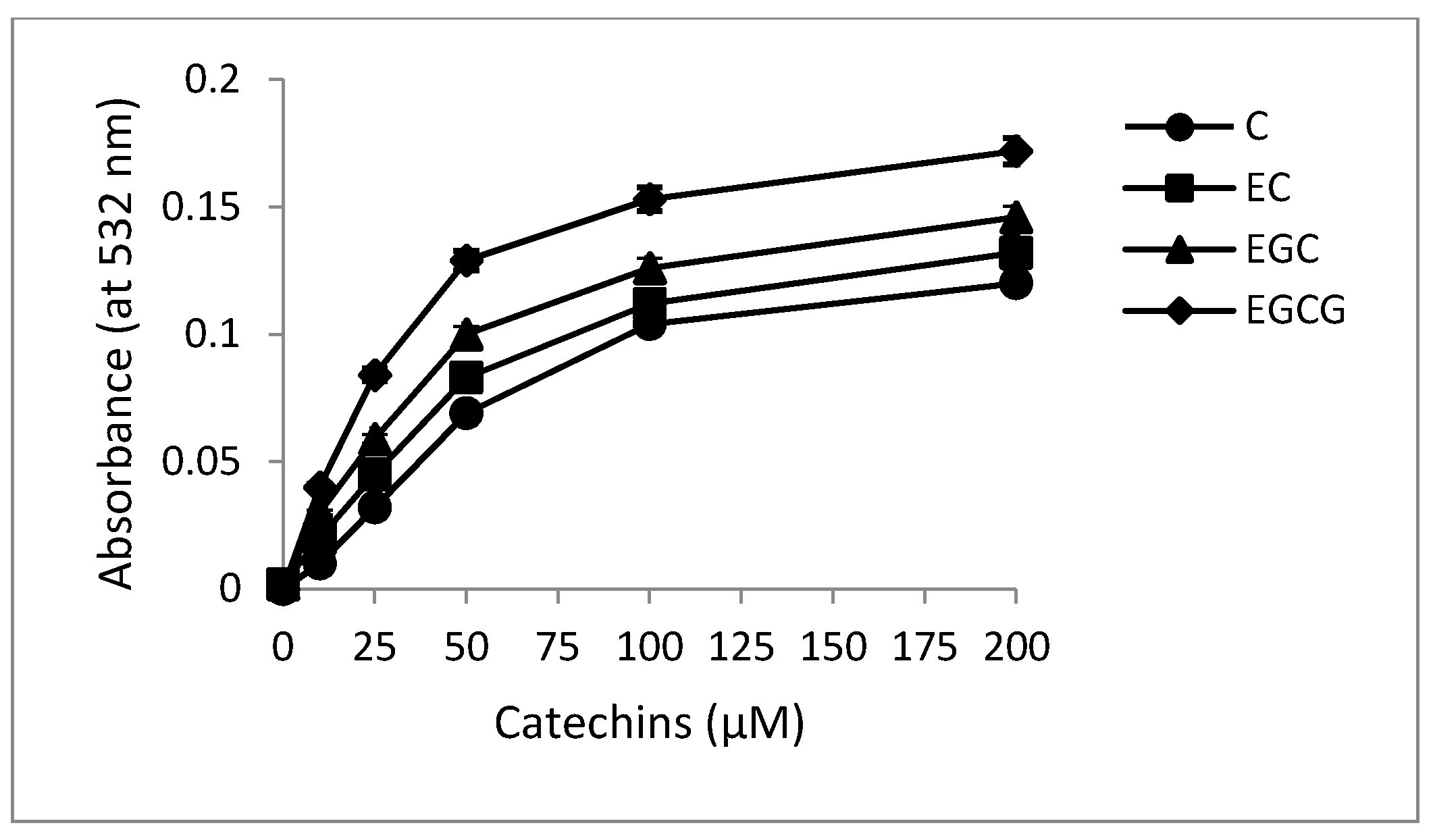
2.4. Superoxide Production by Catechins
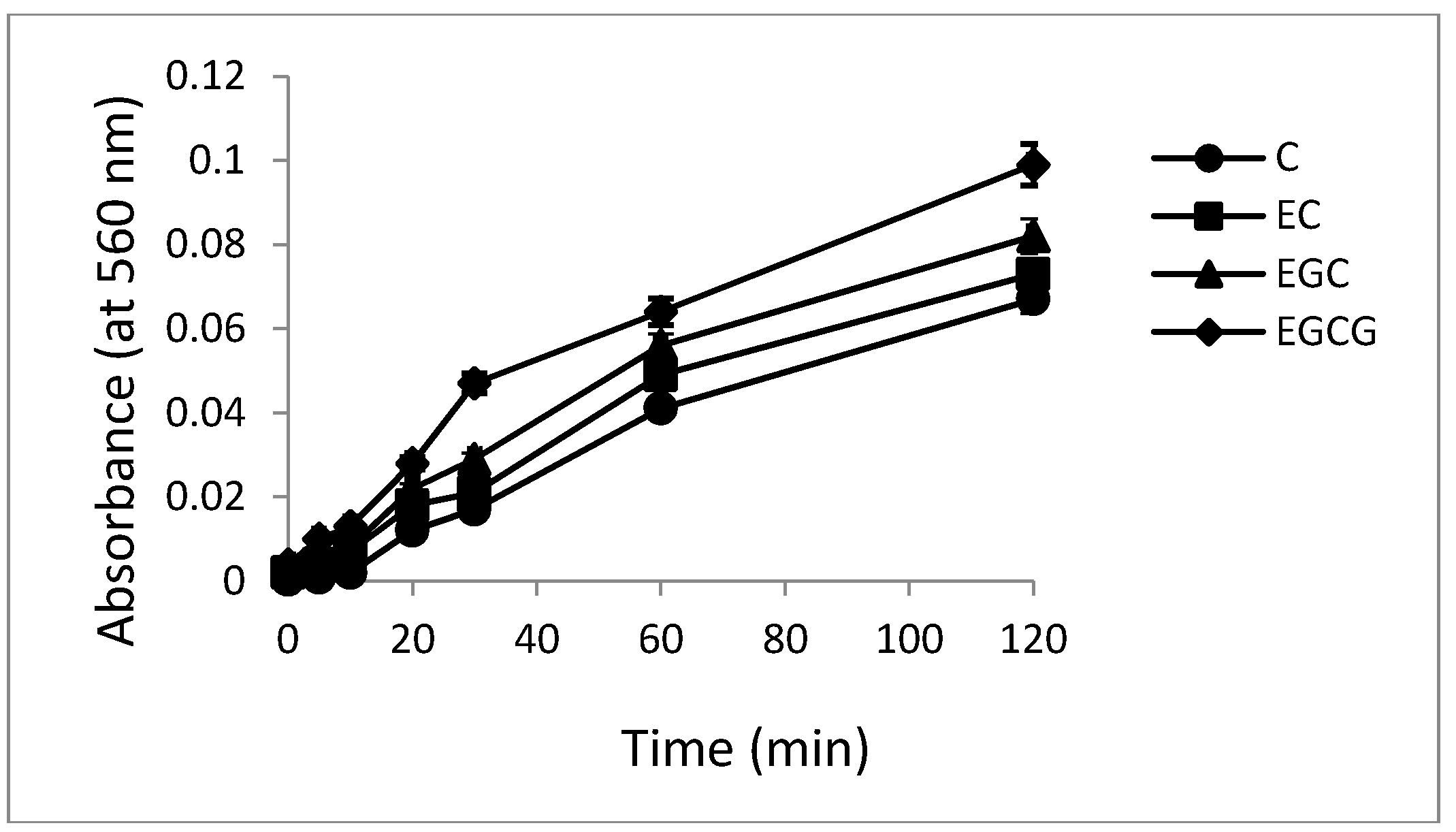
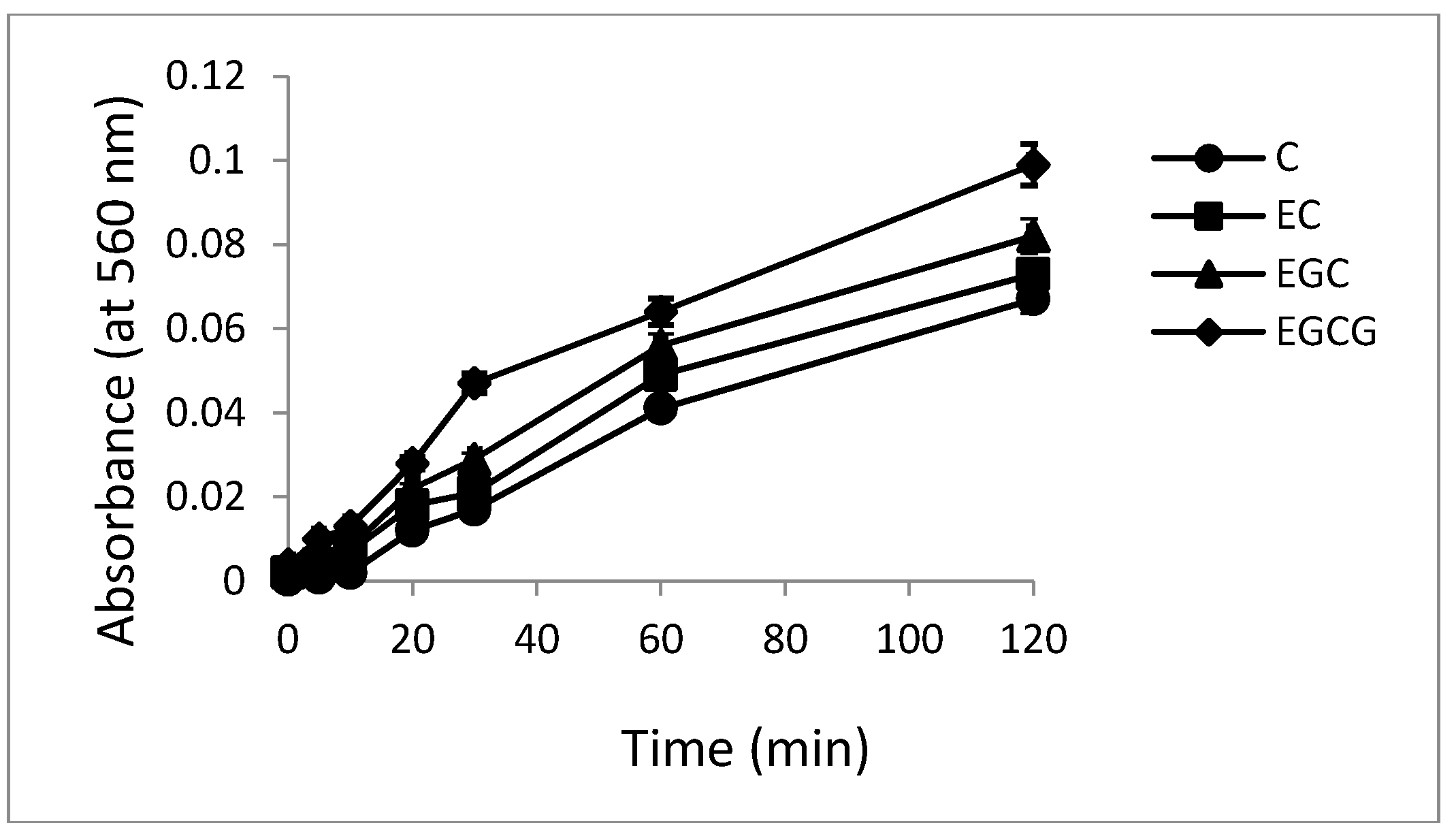
2.5. Hydroxyl Radical Generation by Catechins
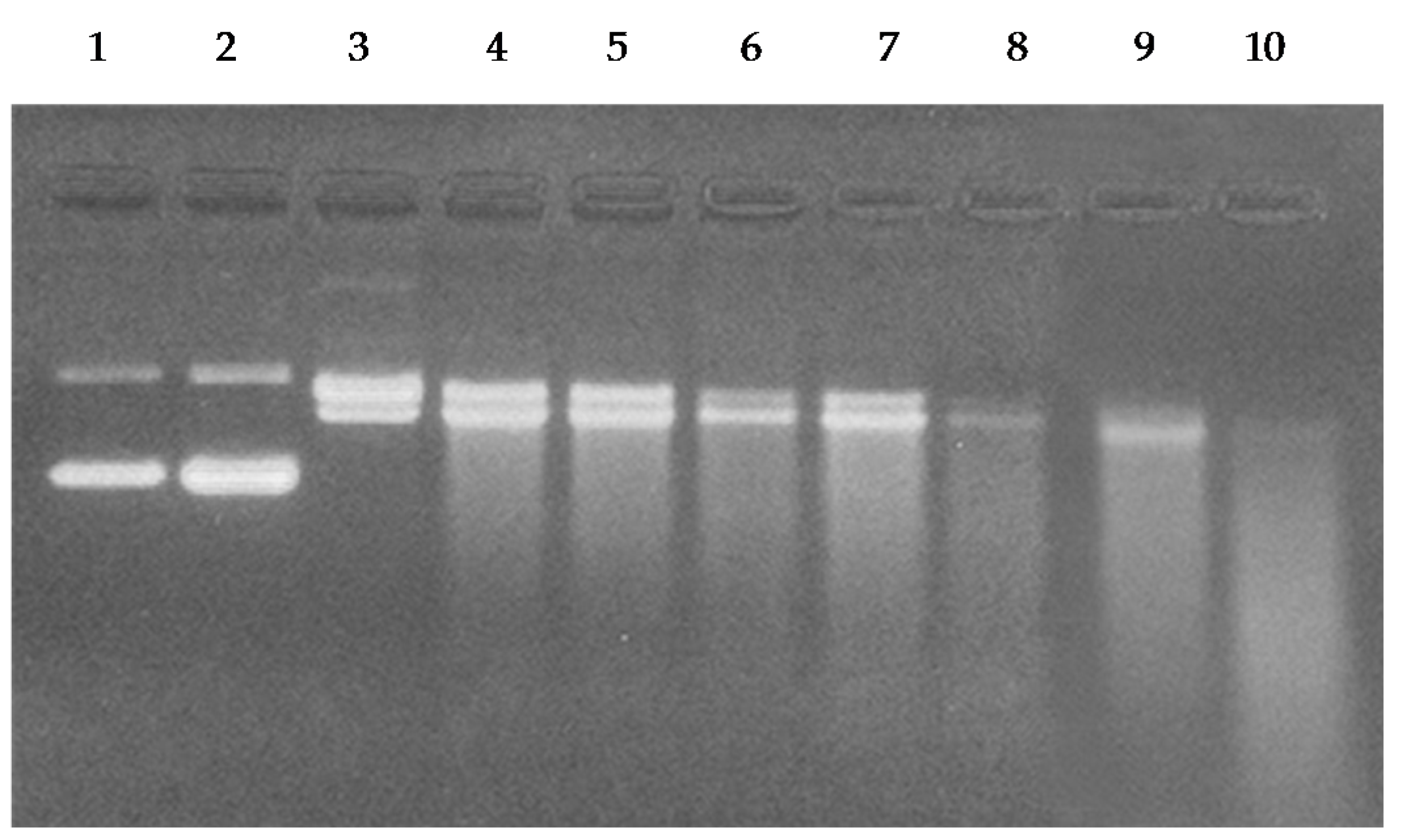
2.6. Breakage of Calf Thymus DNA by Catechins in the Presence of Cu(II)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Catechins | Concentration (µM) | % DNA Hydrolyzed | |
|---|---|---|---|
| without Cu(II) | with Cu(II) | ||
| C | 50 | 1.53 ± 0.21 | 8.87 ± 0.46 |
| 100 | 3.09 ± 0.37 | 11.93 ± 0.62 | |
| 200 | 5.98 ± 0.26 | 17.65 ± 0.73 | |
| 300 | 7.11 ± 0.67 | 26.34 ± 0.91 | |
| EC | 50 | 2.62 ± 0.29 | 10.43 ± 0.41 |
| 100 | 4.18 ± 0.35 | 15.07 ± 0.48 | |
| 200 | 7.26 ± 0.32 | 22.32 ± 0.81 | |
| 300 | 9.12 ± 0.56 | 28.56 ± 0.98 | |
| EGC | 50 | 3.03 ± 0.43 | 12.71 ± 0.51 |
| 100 | 5.21 ± 0.72 | 16.27 ± 0.81 | |
| 200 | 8.11 ± 0.54 | 22.45 ± 0.94 | |
| 300 | 10.87 ± 0.76 | 31.17 ± 1.13 | |
| EGCG | 50 | 4.26 ± 0.43 | 14.11 ± 0.52 |
| 100 | 8.67 ± 0.59 | 18.19 ± 0.77 | |
| 200 | 11.89 ± 0.74 | 29.58 ± 0.83 | |
| 300 | 14.35 ± 0.93 | 38.74 ± 1.71 | |
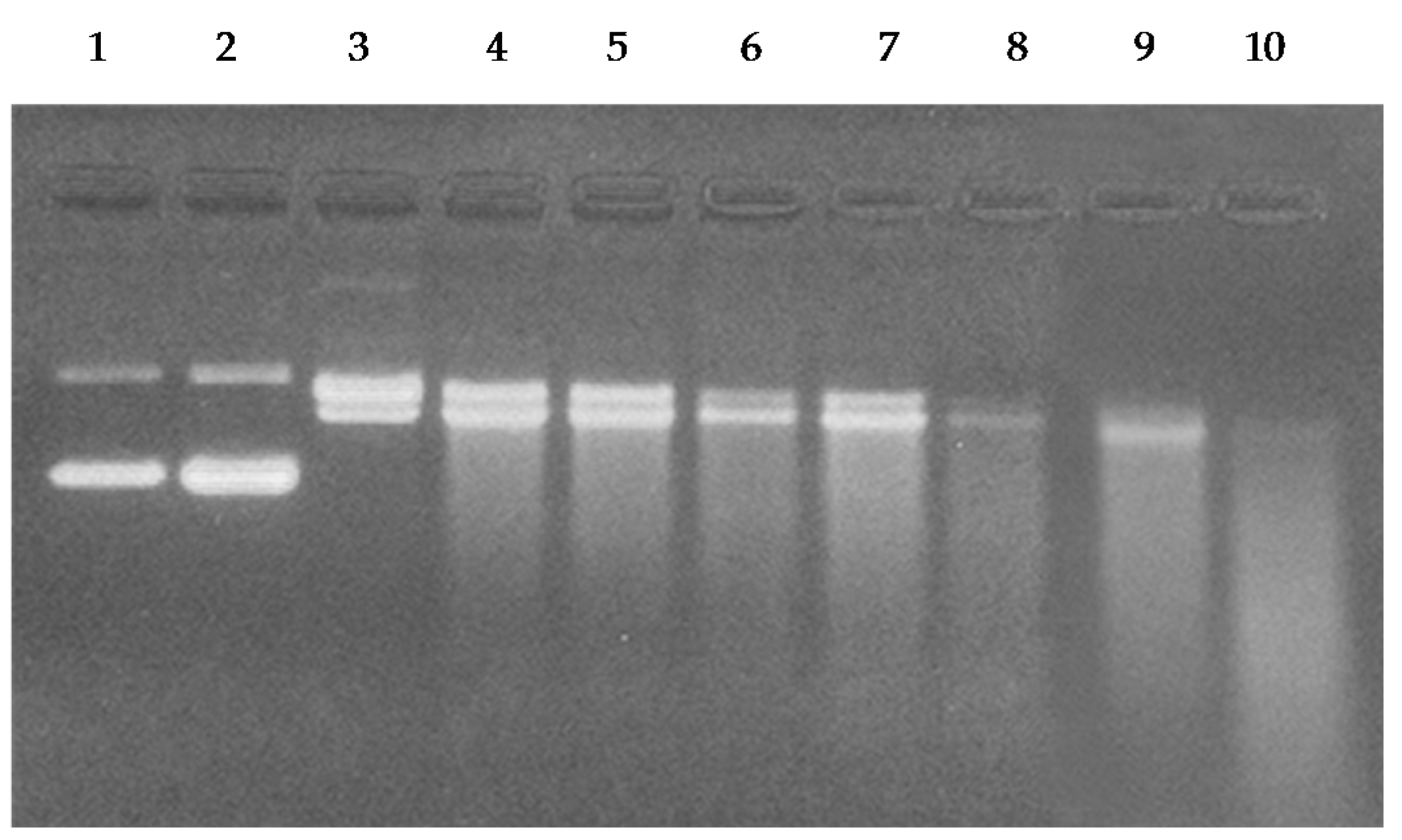
2.7. Cleavage of Plasmid pBR322 DNA by Catechins
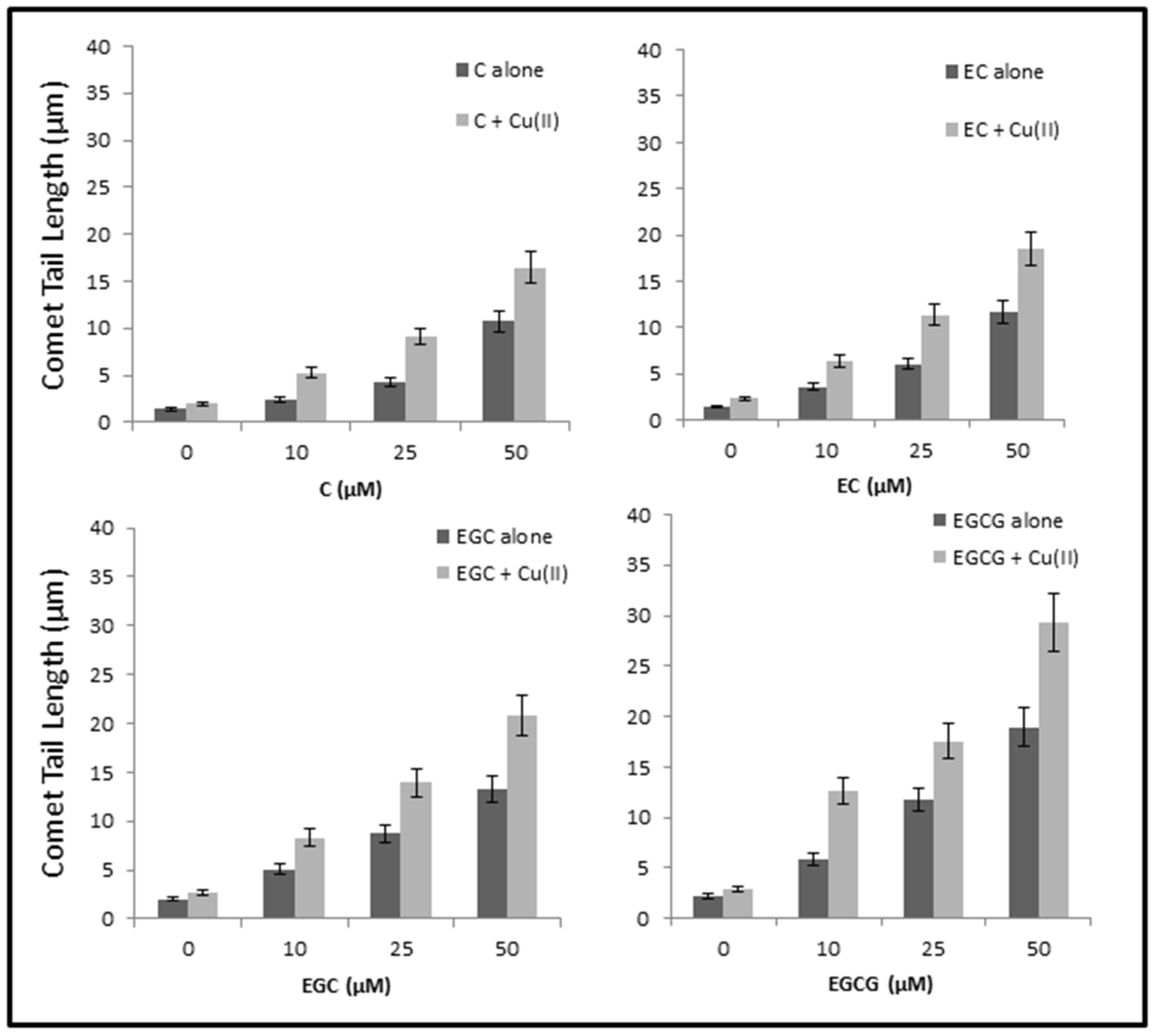
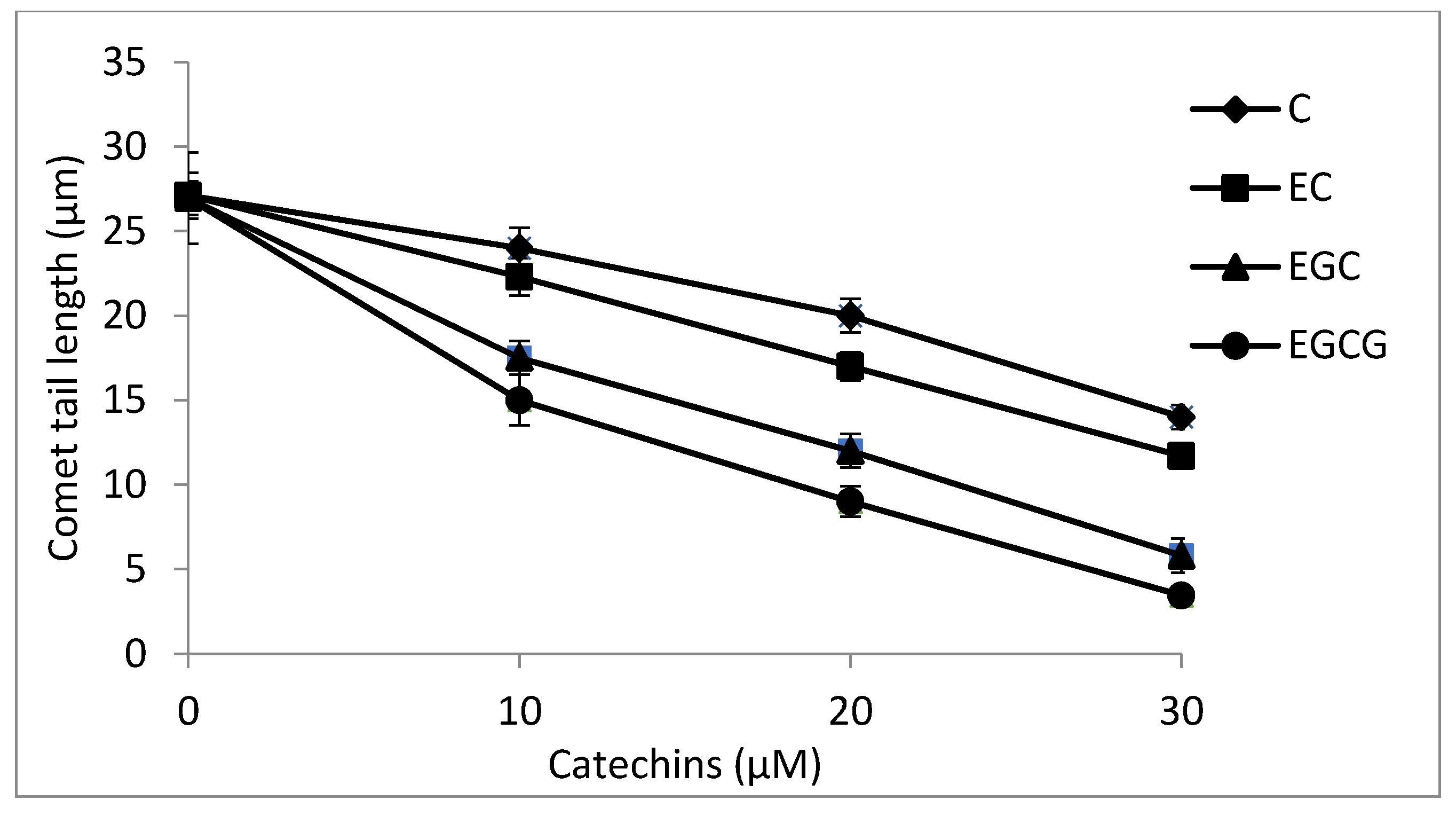
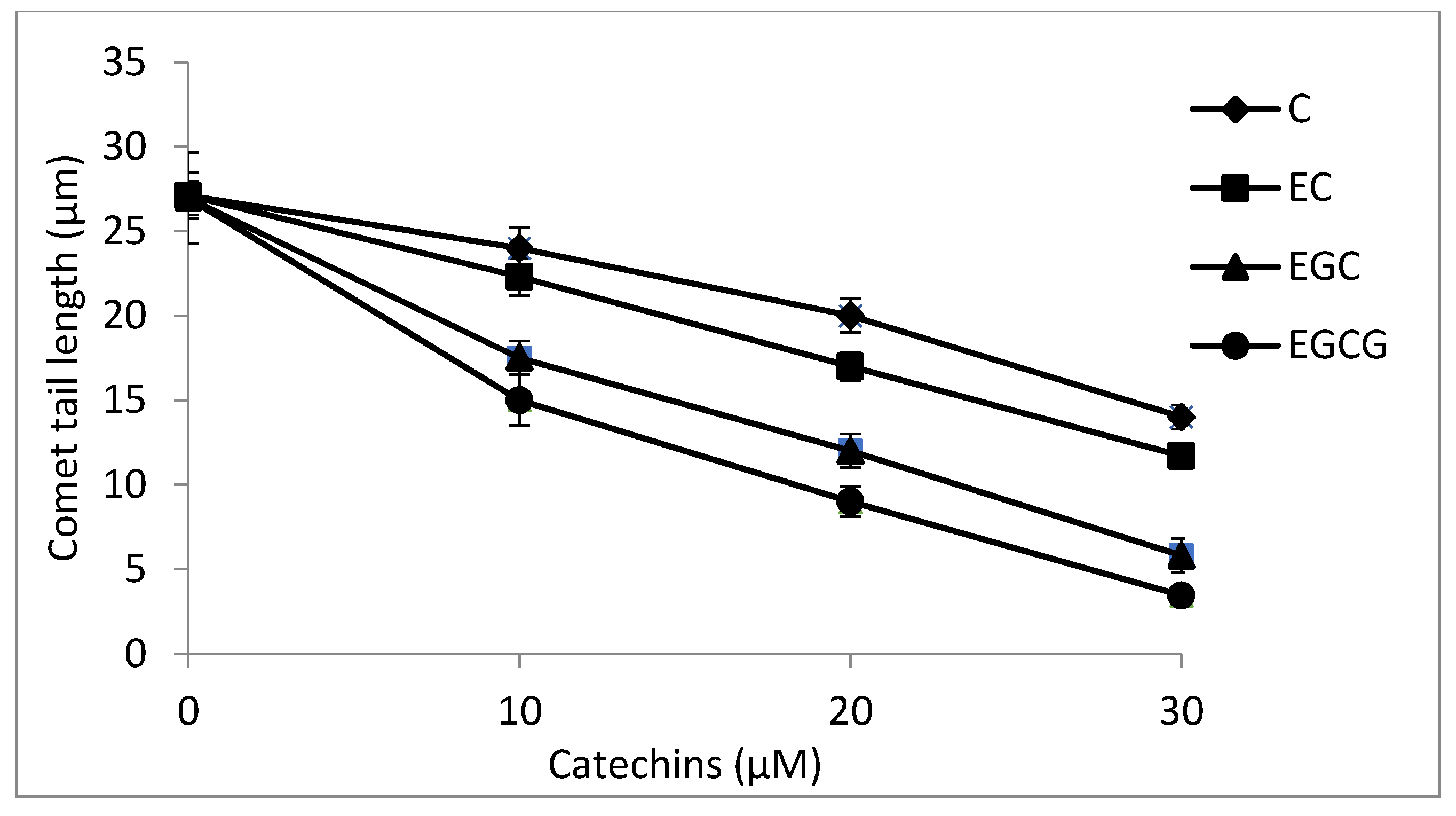
2.8. Cellular DNA Breakage by catechins-Cu(II) in Lymphocytes as Measured by Comet Assay

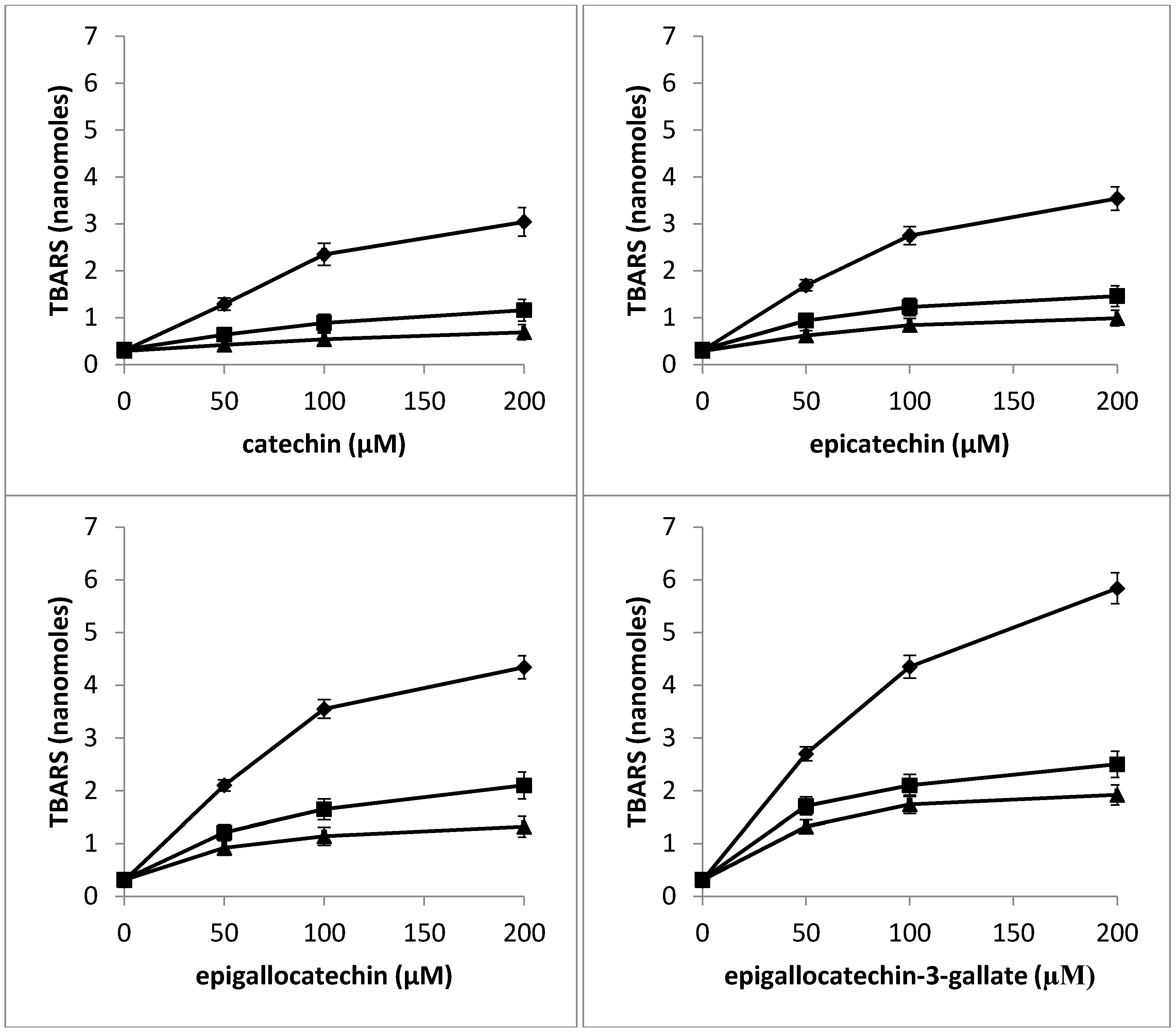
2.9. Determination of TBARS as a Measure of Oxidative Stress in Nuclei by Catechins in the Presence of Neocuproine and Thiourea

2.10. Antioxidant Activity of Catechins against TBHP-Induced Oxidative Stress in Lymphocytes
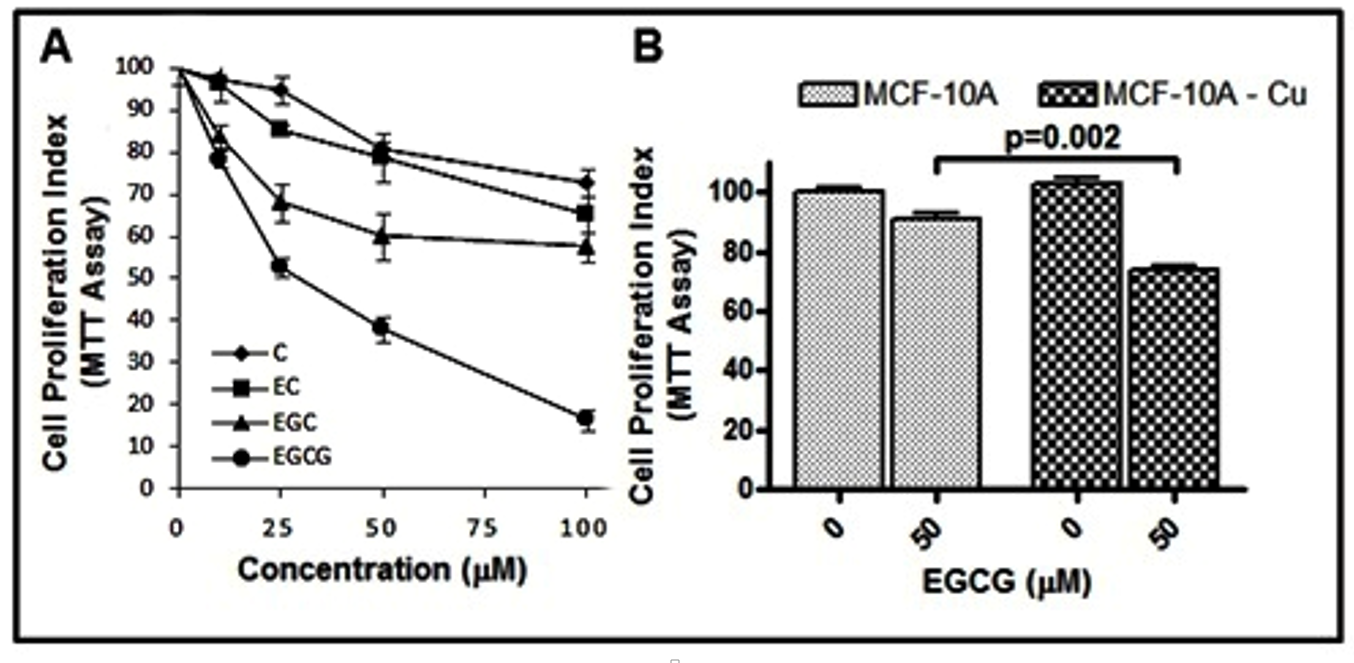
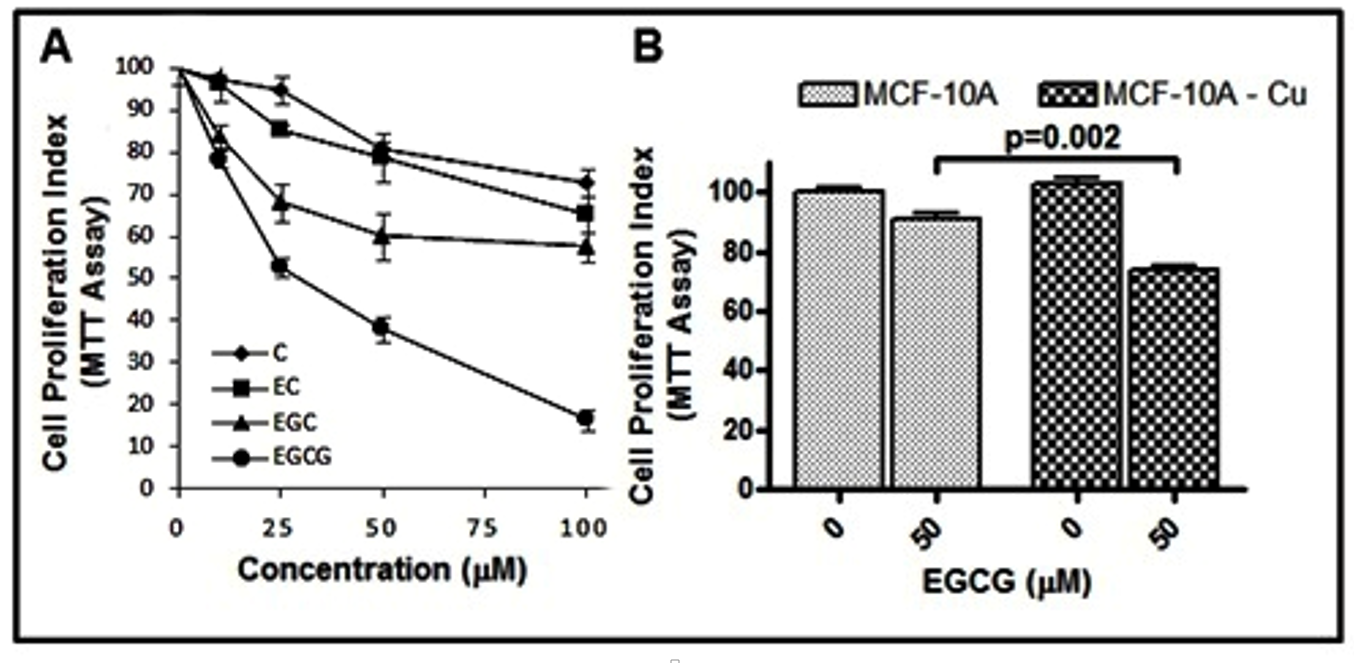
2.11. Catechins Cause Inhibition of Cell Growth in MDA-MB-231 Breast Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals, Reagents and Cell Lines
4.2. Absorbance Studies
4.3. Flourescence Studies
4.4. Detection of Cu(II) Reduction
4.5. Detection of Superoxide Anion Generation
4.6. Detection of Hydroxyl Radical Generation
4.7. Degradation of Calf Thymus DNA
4.8. Treatment of pBR322 DNA
4.9. Isolation of Lymphocytes
4.10. Viability Assessment of Lymphocytes
4.11. Comet Assay
4.12. Determination of TBARS
4.13. Cell Growth Inhibition Studies by MTT Assay
4.14. Statistics
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Adlercreutz, C.H.; Goldin, B.R.; Gorbach, S.L.; Hockerstedt, K.A.; Watanabe, S.; Hamalainen, E.K.; Markkanen, M.H.; Makela, T.H.; Wahala, K.T.; Adlercreutz, T. Soybean phytoestrogen intake and cancer risk. J. Nutr. 1995, 125, 757S–770S. [Google Scholar] [PubMed]
- Park, O.J.; Surh, Y.J. Chemopreventive potential of epigallocatechin gallate and genistein: Evidence from epidemiological and laboratory studies. Toxicol. Lett. 2004, 150, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Barnes, S.; Peterson, G.; Grubbs, C.; Setchell, K. Potential role of dietary isoflavones in the prevention of cancer. Adv. Exp. Med. Biol. 1994, 354, 135–147. [Google Scholar] [PubMed]
- Surh, Y.J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.Y.; Zubair, H.; Faisal, M.; Ullah, M.F.; Farhan, M.; Sarkar, F.H.; Ahmad, A.; Hadi, S.M. Plant polyphenol induced cell death in human cancer cells involves mobilization of intracellular copper ions and reactive oxygen species generation: A mechanism for cancer chemopreventive action. Mol. Nutr. Food Res. 2014, 58, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Liao, J.; Kim, K.; Yurkow, E.J.; Yang, C.S. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 1998, 19, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Shamim, U.; Hanif, S.; Ullah, M.F.; Azmi, A.S.; Bhat, S.H.; Hadi, S.M. Plant polyphenols mobilize nuclear copper in human peripheral lymphocytes leading to oxidatively generated DNA breakage: Implications for an anticancer mechanism. Free Radic. Res. 2008, 42, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.F.; Ahmad, A.; Khan, H.Y.; Zubair, H.; Sarkar, F.H.; Hadi, S.M. The prooxidant action of dietary antioxidants leading to cellular DNA breakage and anticancer effects: Implications for chemotherapeutic action against cancer. Cell Biochem. Biophys. 2013, 67, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.V.; Hirpara, J.L.; Chawdhury, S.H.; Pervaiz, S. Chemopreventive agent resveratrol, a natural product derived from grapes, triggers cd95 signaling-dependent apoptosis in human tumor cells. Blood 1998, 92, 996–1002. [Google Scholar] [PubMed]
- Kuo, M.L.; Huang, T.S.; Lin, J.K. Curcumin, an antioxidant and anti-tumor promoter, induces apoptosis in human leukemia cells. Biochim. Biophys. Acta 1996, 1317, 95–100. [Google Scholar] [CrossRef]
- Chang, K.L.; Cheng, H.L.; Huang, L.W.; Hsieh, B.S.; Hu, Y.C.; Chih, T.T.; Shyu, H.W.; Su, S.J. Combined effects of terazosin and genistein on a metastatic, hormone-independent human prostate cancer cell line. Cancer Lett. 2009, 276, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Hastak, K.; Ahmad, N.; Lewin, J.S.; Mukhtar, H. Inhibition of prostate carcinogenesis in tramp mice by oral infusion of green tea polyphenols. Proc. Natl. Acad. Sci. USA 2001, 98, 10350–10355. [Google Scholar] [CrossRef] [PubMed]
- Orsolic, N.; Knezevic, A.; Sver, L.; Terzic, S.; Hackenberger, B.K.; Basic, I. Influence of honey bee products on transplantable murine tumours. Vet. Comp. Oncol. 2003, 1, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.P.; Schell, J.B.; Ho, C.T.; Chen, K.Y. Green tea epigallocatechin gallate shows a pronounced growth inhibitory effect on cancerous cells but not on their normal counterparts. Cancer Lett. 1998, 129, 173–179. [Google Scholar] [CrossRef]
- Said Ahmad, M.; Fazal, F.; Rahman, A.; Hadi, S.M.; Parish, J.H. Activities of flavonoids for the cleavage of DNA in the presence of cu(ii): Correlation with generation of active oxygen species. Carcinogenesis 1992, 13, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Gali, H.U.; Perchellet, E.M.; Klish, D.S.; Johnson, J.M.; Perchellet, J.P. Hydrolyzable tannins: Potent inhibitors of hydroperoxide production and tumor promotion in mouse skin treated with 12-o-tetradecanoylphorbol-13-acetate in vivo. Int. J. Cancer 1992, 51, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Hadi, S.M.; Asad, S.F.; Singh, S.; Ahmad, A. Putative mechanism for anticancer and apoptosis-inducing properties of plant-derived polyphenolic compounds. IUBMB Life 2000, 50, 167–171. [Google Scholar] [PubMed]
- Hadi, S.M.; Bhat, S.H.; Azmi, A.S.; Hanif, S.; Shamim, U.; Ullah, M.F. Oxidative breakage of cellular DNA by plant polyphenols: A putative mechanism for anticancer properties. Semin. Cancer Biol. 2007, 17, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.H.; Azmi, A.S.; Hanif, S.; Hadi, S.M. Ascorbic acid mobilizes endogenous copper in human peripheral lymphocytes leading to oxidative DNA breakage: A putative mechanism for anticancer properties. Int. J. Biochem. Cell Biol. 2006, 38, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Kagawa, T.F.; Geierstanger, B.H.; Wang, A.H.; Ho, P.S. Covalent modification of guanine bases in double-stranded DNA. The 1.2-A Z-DNA structure of d(CGCGCG) in the presence of CuCl2. J. Biol. Chem. 1991, 266, 20175–20184. [Google Scholar] [PubMed]
- Ebadi, M.; Swanson, S. The status of zinc, copper, and metallothionein in cancer patients. Prog. Clin. Biol. Res. 1988, 259, 161–175. [Google Scholar] [PubMed]
- Margalioth, E.J.; Udassin, R.; Cohen, C.; Maor, J.; Anteby, S.O.; Schenker, J.G. Serum copper level in gynecologic malignancies. Am. J. Obstet. Gynecol. 1987, 157, 93–96. [Google Scholar] [CrossRef]
- Yoshida, D.; Ikeda, Y.; Nakazawa, S. Quantitative analysis of copper, zinc and copper/zinc ratio in selected human brain tumors. J. Neuro-Oncol. 1993, 16, 109–115. [Google Scholar] [CrossRef]
- Ebara, M.; Fukuda, H.; Hatano, R.; Saisho, H.; Nagato, Y.; Suzuki, K.; Nakajima, K.; Yukawa, M.; Kondo, F.; Nakayama, A.; et al. Relationship between copper, zinc and metallothionein in hepatocellular carcinoma and its surrounding liver parenchyma. J. Hepatol. 2000, 33, 415–422. [Google Scholar] [CrossRef]
- Zheng, L.F.; Wei, Q.Y.; Cai, Y.J.; Fang, J.G.; Zhou, B.; Yang, L.; Liu, Z.L. DNA damage induced by resveratrol and its synthetic analogues in the presence of Cu (II) ions: Mechanism and structure-activity relationship. Free Radic. Biol. Med. 2006, 41, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Farhan, M.; Zafar, A.; Chibber, S.; Khan, H.Y.; Arif, H.; Hadi, S.M. Mobilization of copper ions in human peripheral lymphocytes by catechins leading to oxidative DNA breakage: A structure activity study. Arch. Biochem. Biophys. 2015, 580, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.A.; Narita, S.; Gieseg, S.; Gebicki, S.; Gebicki, J.M.; Dean, R.T. Long-lived reactive species on free-radical-damaged proteins. Biochem. J. 1992, 282, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Kimura, T.; Kodama, M.; Nagata, C. Generation of hydrogen peroxide and superoxide anion from active metabolites of naphthylamines and aminoazo dyes: Its possible role in carcinogenesis. Carcinogenesis 1983, 4, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Shahabuddin; Hadi, S.M.; Parish, J.H.; Ainley, K. Strand scission in DNA induced by quercetin and Cu(II): Role of Cu(I) and oxygen free radicals. Carcinogenesis 1989, 10, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, G.J.; Gutteridge, J.M. Oxygen radical damage to DNA by rifamycin sv and copper ions. Biochem. Pharmacol. 1987, 36, 3629–3633. [Google Scholar] [CrossRef]
- Smith, C.; Halliwell, B.; Aruoma, O.I. Protection by albumin against the pro-oxidant actions of phenolic dietary components. Food Chem. Toxicol. 1992, 30, 483–489. [Google Scholar] [CrossRef]
- Dubuisson, M.L.; de Wergifosse, B.; Trouet, A.; Baguet, F.; Marchand-Brynaert, J.; Rees, J.F. Antioxidative properties of natural coelenterazine and synthetic methyl coelenterazine in rat hepatocytes subjected to tert-butyl hydroperoxide-induced oxidative stress. Biochem. Pharmacol. 2000, 60, 471–478. [Google Scholar] [CrossRef]
- Suzuki, Y.; Apostolova, M.D.; Cherian, M.G. Astrocyte cultures from transgenic mice to study the role of metallothionein in cytotoxicity of tert-butyl hydroperoxide. Toxicology 2000, 145, 51–62. [Google Scholar] [CrossRef]
- Ahmad, N.; Feyes, D.K.; Nieminen, A.L.; Agarwal, R.; Mukhtar, H. Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J. Natl. Cancer Inst. 1997, 89, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Chevion, M. A site-specific mechanism for free radical induced biological damage: The essential role of redox-active transition metals. Free Radic. Biol. Med. 1988, 5, 27–37. [Google Scholar] [CrossRef]
- Zimmer, C.; Luck, G.; Fritzsche, H.; Triebel, H. DNA-copper (II) complex and the DNA conformation. Biopolymers 1971, 10, 441–463. [Google Scholar] [CrossRef] [PubMed]
- Gupte, A.; Mumper, R.J. Elevated copper and oxidative stress in cancer cells as a target for cancer treatment. Cancer Treat. Rev. 2009, 35, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Devi, G.S.; Prasad, M.H.; Saraswathi, I.; Raghu, D.; Rao, D.N.; Reddy, P.P. Free radicals antioxidant enzymes and lipid peroxidation in different types of leukemias. Clin. Chim. Acta 2000, 293, 53–62. [Google Scholar] [CrossRef]
- Oberley, T.D.; Oberley, L.W. Antioxidant enzyme levels in cancer. Histol. Histopathol. 1997, 12, 525–535. [Google Scholar] [PubMed]
- Kong, Q.; Beel, J.A.; Lillehei, K.O. A threshold concept for cancer therapy. Med. Hypotheses 2000, 55, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Van Amelsvoort, J.M.; Van Hof, K.H.; Mathot, J.N.; Mulder, T.P.; Wiersma, A.; Tijburg, L.B. Plasma concentrations of individual tea catechins after a single oral dose in humans. Xenobiotica: Fate Foreign Compd. Biol. Syst. 2001, 31, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.Y.; Zubair, H.; Ullah, M.F.; Ahmad, A.; Hadi, S.M. Oral administration of copper to rats leads to increased lymphocyte cellular DNA degradation by dietary polyphenols: Implications for a cancer preventive mechanism. Biometals 2011, 24, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.R.; Wang, D.; Zhang, H.; Peng, S.; Shin, H.J.; Brandes, J.C.; Tighiouart, M.; Khuri, F.R.; Chen, Z.G.; Shin, D.M. Enhanced anti-tumor activity by the combination of the natural compounds (−)-epigallocatechin-3-gallate and luteolin: Potential role of p53. J. Biol. Chem. 2010, 285, 34557–34565. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.A.; Hadi, S.M. Partial purification and properties of an endonuclease from germinating pea seeds specific for single-stranded DNA. Arch. Biochem. Biophys. 1979, 196, 138–146. [Google Scholar] [CrossRef]
- Schneider, W.C. Determination of nucleic acids in tissues by pentose analysis. Methods Enzymol 1957, 3, 880–884. [Google Scholar]
- Pool-Zobel, B.L.; Guigas, C.; Klein, R.; Neudecker, C.; Renner, H.W.; Schmezer, P. Assessment of genotoxic effects by lindane. Food Chem. Toxicol. 1993, 31, 271–283. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Azmi, A.S.; Bhat, S.H.; Hadi, S.M. Resveratrol-Cu(II) induced DNA breakage in human peripheral lymphocytes: Implications for anticancer properties. FEBS Lett. 2005, 579, 3131–3135. [Google Scholar] [CrossRef] [PubMed]
- Kanupriya; Dipti, P.; Sharma, S.K.; Sairam, M.; Ilavazhagan, G.; Sawhney, R.C.; Banerjee, P.K. Flavonoids protect u-937 macrophages against tert-butylhydroperoxide induced oxidative injury. Food Chem. Toxicol. 2006, 44, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, R.; Das, N.P.; Tan, C.H. Effects of gamma-linolenic acid, flavonoids, and vitamins on cytotoxicity and lipid peroxidation. Free Radic. Biol. Med. 1994, 16, 43–48. [Google Scholar] [CrossRef]
- Tice, R.R.; Strauss, G.H. The single cell gel electrophoresis/comet assay: A potential tool for detecting radiation-induced DNA damage in humans. Stem Cells 1995, 13 (Suppl. 1), 207–214. [Google Scholar] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farhan, M.; Khan, H.Y.; Oves, M.; Al-Harrasi, A.; Rehmani, N.; Arif, H.; Hadi, S.M.; Ahmad, A. Cancer Therapy by Catechins Involves Redox Cycling of Copper Ions and Generation of Reactive Oxygen Species. Toxins 2016, 8, 37. https://doi.org/10.3390/toxins8020037
Farhan M, Khan HY, Oves M, Al-Harrasi A, Rehmani N, Arif H, Hadi SM, Ahmad A. Cancer Therapy by Catechins Involves Redox Cycling of Copper Ions and Generation of Reactive Oxygen Species. Toxins. 2016; 8(2):37. https://doi.org/10.3390/toxins8020037
Chicago/Turabian StyleFarhan, Mohd, Husain Yar Khan, Mohammad Oves, Ahmed Al-Harrasi, Nida Rehmani, Hussain Arif, Sheikh Mumtaz Hadi, and Aamir Ahmad. 2016. "Cancer Therapy by Catechins Involves Redox Cycling of Copper Ions and Generation of Reactive Oxygen Species" Toxins 8, no. 2: 37. https://doi.org/10.3390/toxins8020037