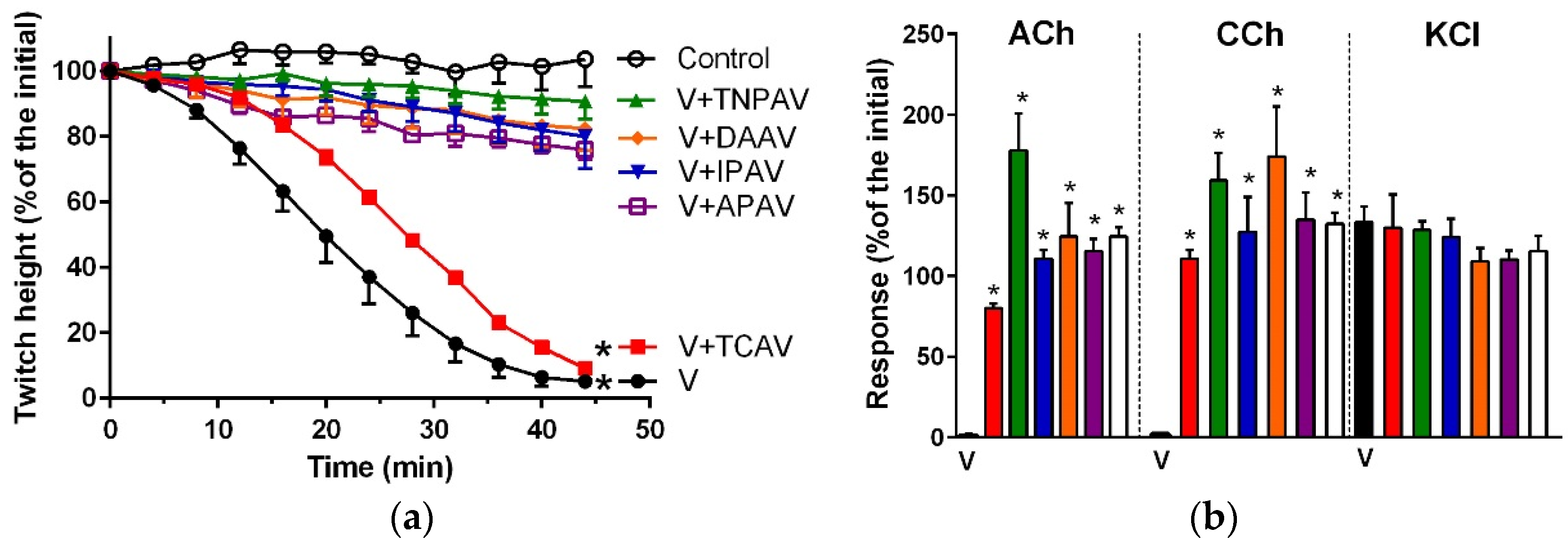
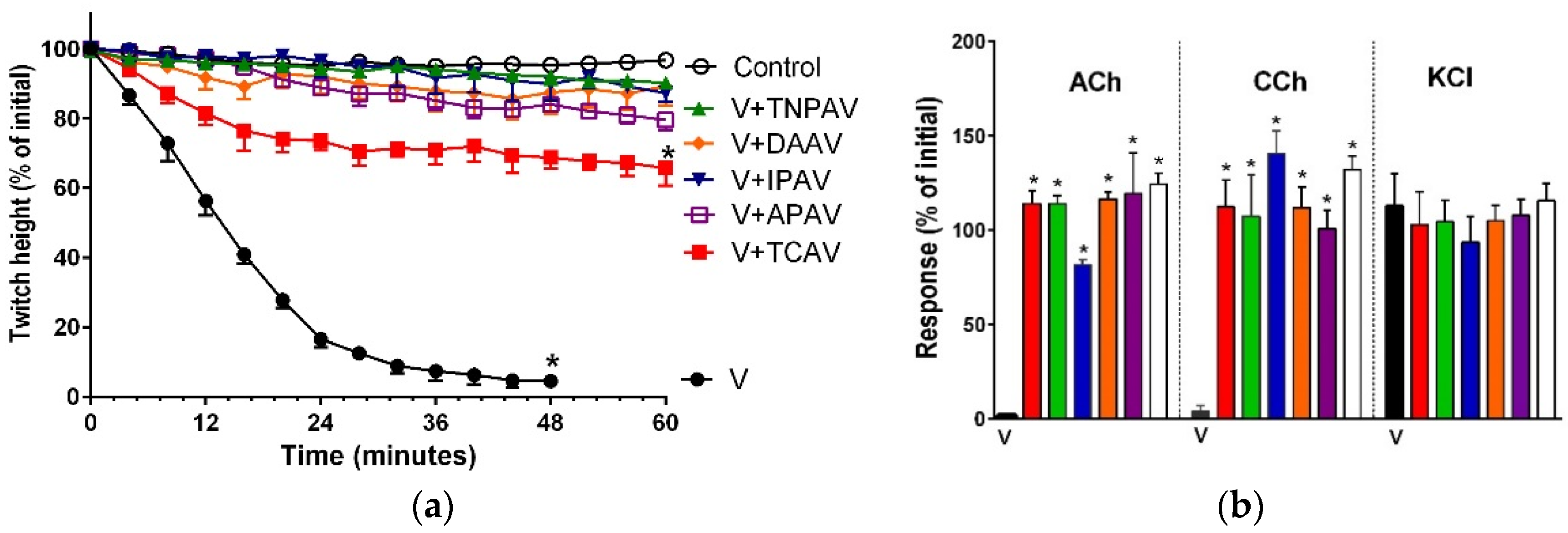
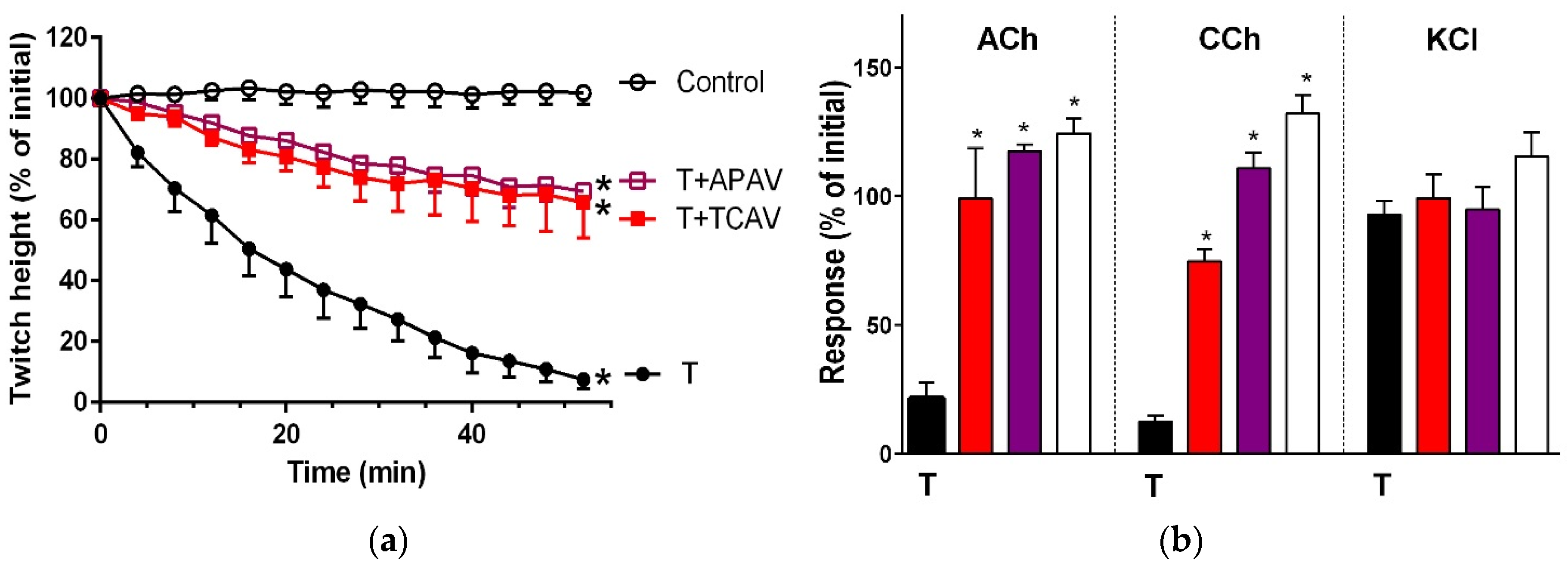
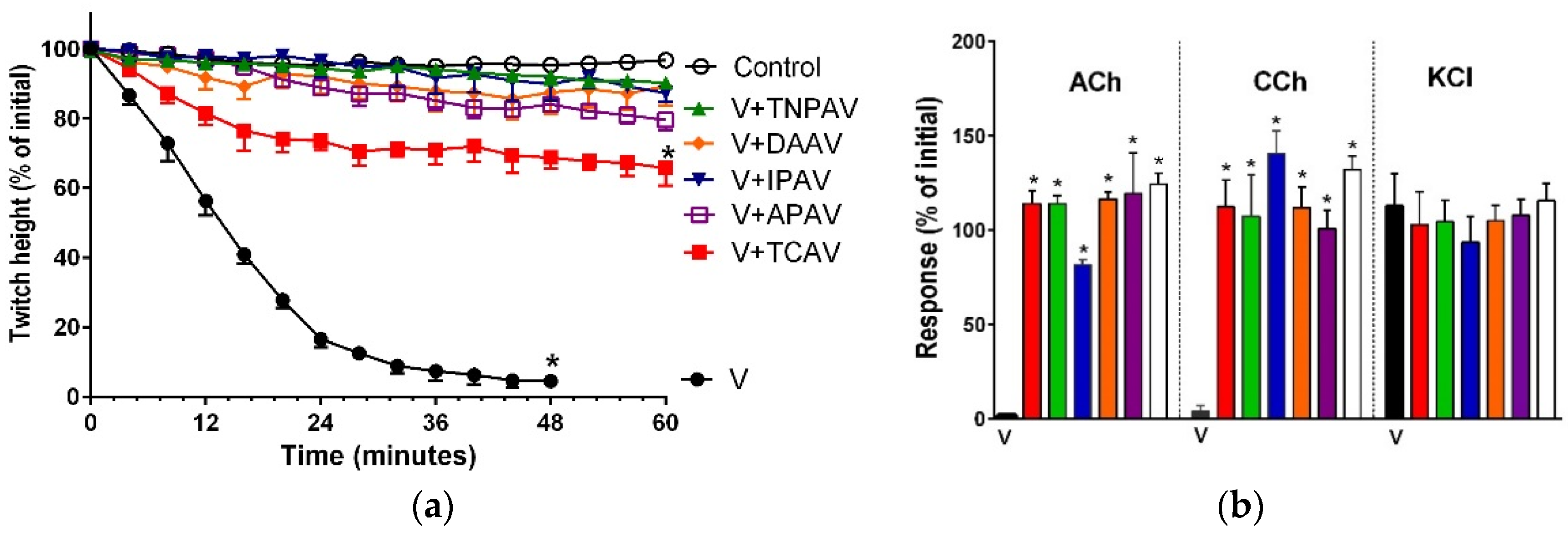
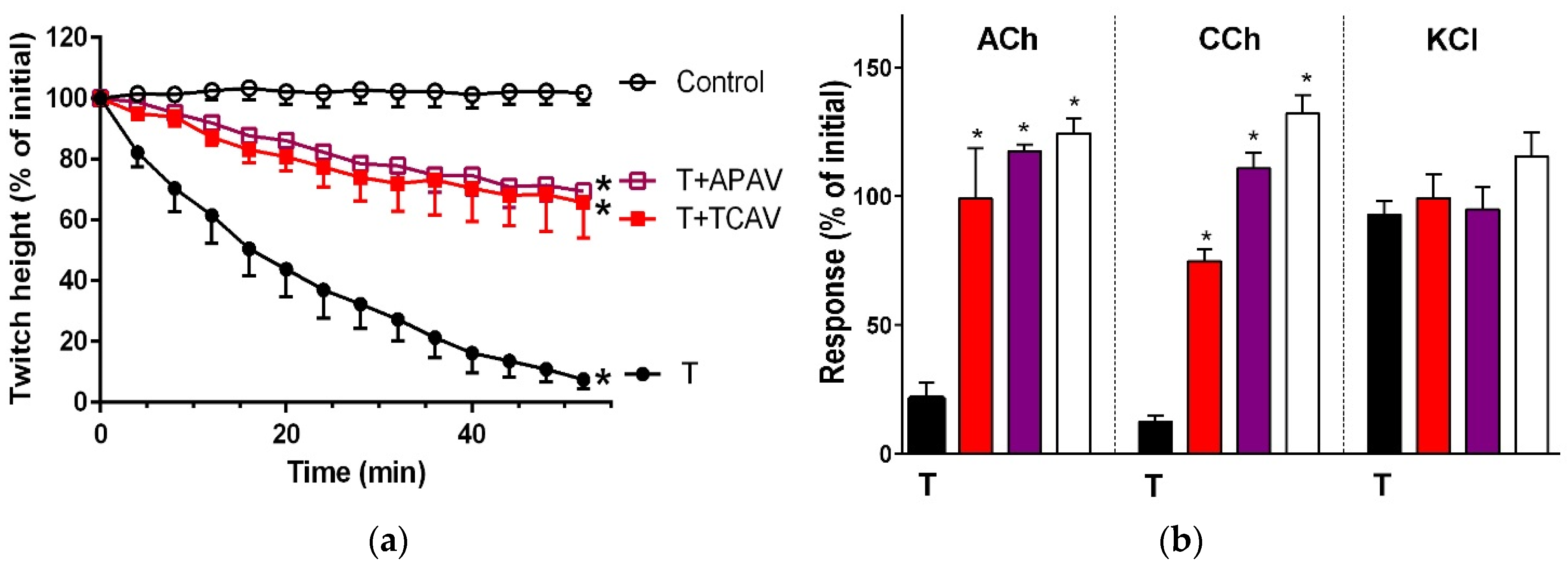
Figure 1.
Cross-neutralisation of the neurotoxicity of Bungarus caeruleus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms except TCAV prevented neurotoxicity (* significantly different from control at 44 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and the presence of antivenoms compared to control. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
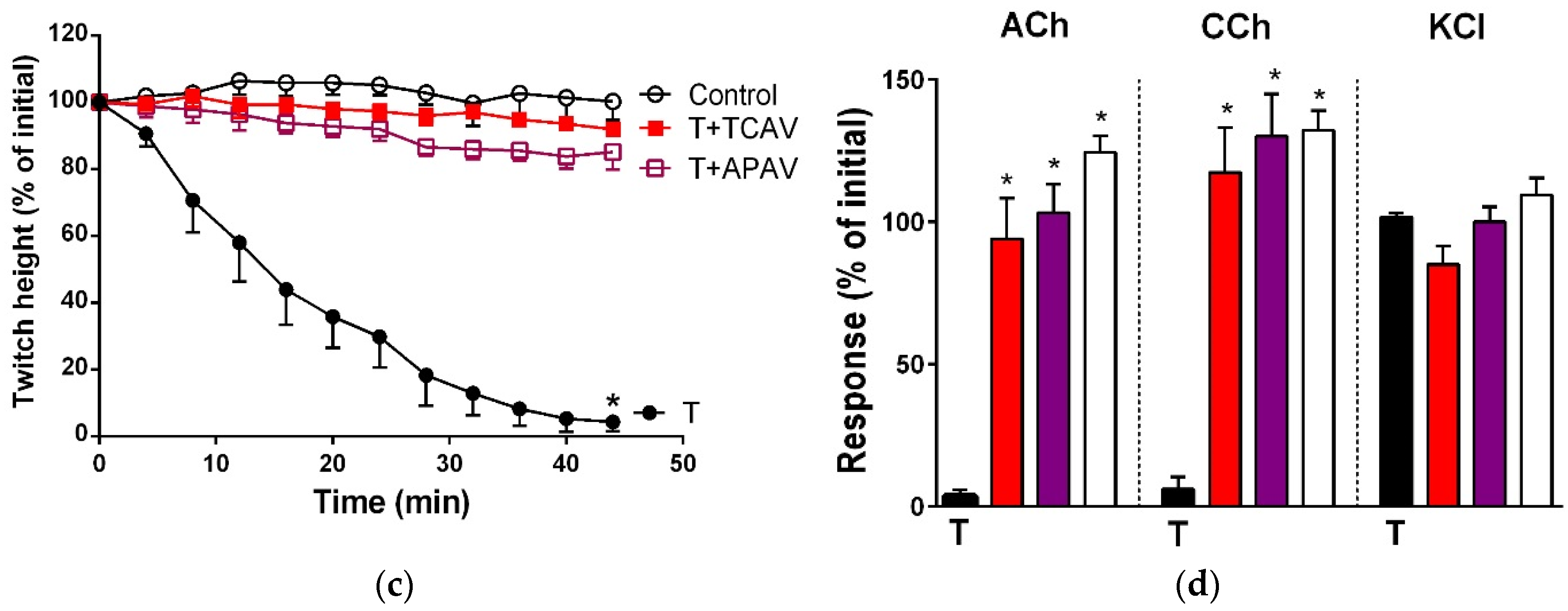
Figure 1.
Cross-neutralisation of the neurotoxicity of Bungarus caeruleus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms except TCAV prevented neurotoxicity (* significantly different from control at 44 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and the presence of antivenoms compared to control. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
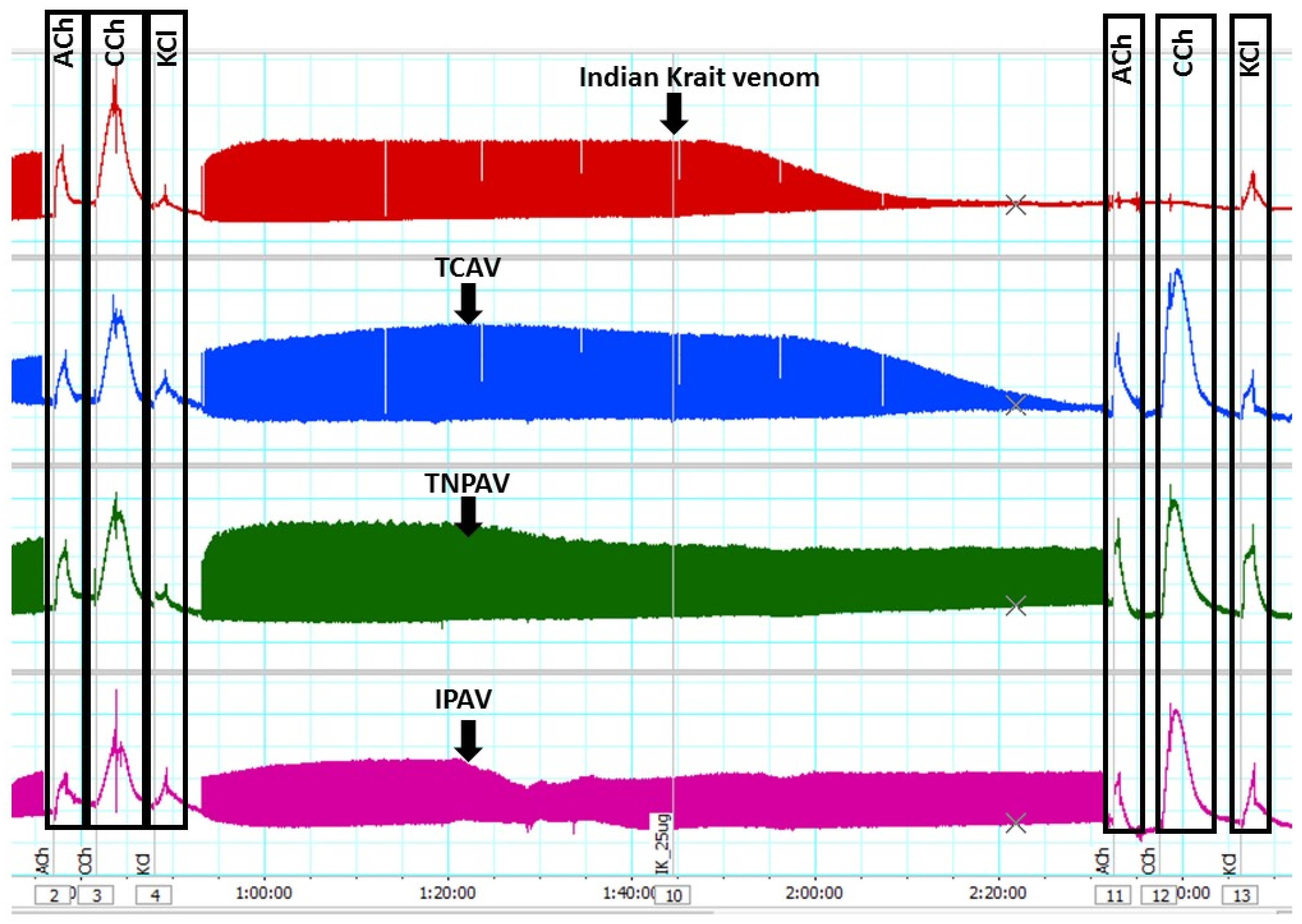
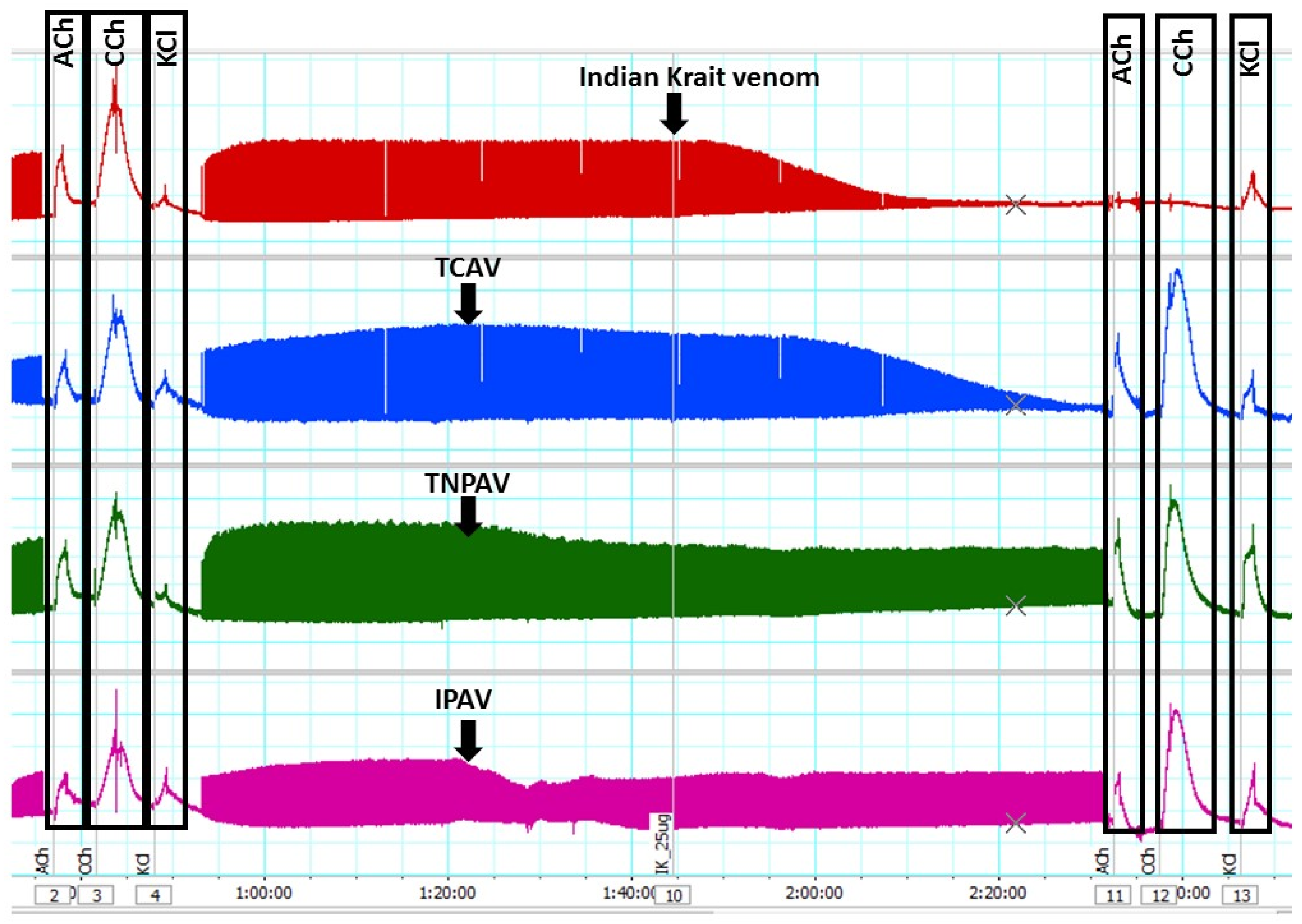
Figure 2.
Traces showing indirect twitch and agonist contractile responses of the chick-biventer nerve-muscle preparation for B. caeruleus venom (5 µg/mL) in the absence and the presence of Thai cobra antivenom (TCAV), Thai neuro polyvalent antivenom (TNPAV) and Indian polyvalent antivenom (IPAV). Note: the venom (red trace) inhibits indirect twitches and abolishes ACh and CCh responses indicating post-synaptic neurotoxicity. In the presence of TCAV (blue trace), the venom inhibits indirect twitches but does not abolish ACh and CCh responses indicating that the pre-synaptic neurotoxicity is not neutralised. In the presence of TNPAV (green trace) and IPAV (purple trace), venom fails to inhibit indirect twitches and fails to abolish ACh and CCh responses indicating neutralisation of both pre- and post-synaptic neurotoxicity of the venom.
Figure 2.
Traces showing indirect twitch and agonist contractile responses of the chick-biventer nerve-muscle preparation for B. caeruleus venom (5 µg/mL) in the absence and the presence of Thai cobra antivenom (TCAV), Thai neuro polyvalent antivenom (TNPAV) and Indian polyvalent antivenom (IPAV). Note: the venom (red trace) inhibits indirect twitches and abolishes ACh and CCh responses indicating post-synaptic neurotoxicity. In the presence of TCAV (blue trace), the venom inhibits indirect twitches but does not abolish ACh and CCh responses indicating that the pre-synaptic neurotoxicity is not neutralised. In the presence of TNPAV (green trace) and IPAV (purple trace), venom fails to inhibit indirect twitches and fails to abolish ACh and CCh responses indicating neutralisation of both pre- and post-synaptic neurotoxicity of the venom.
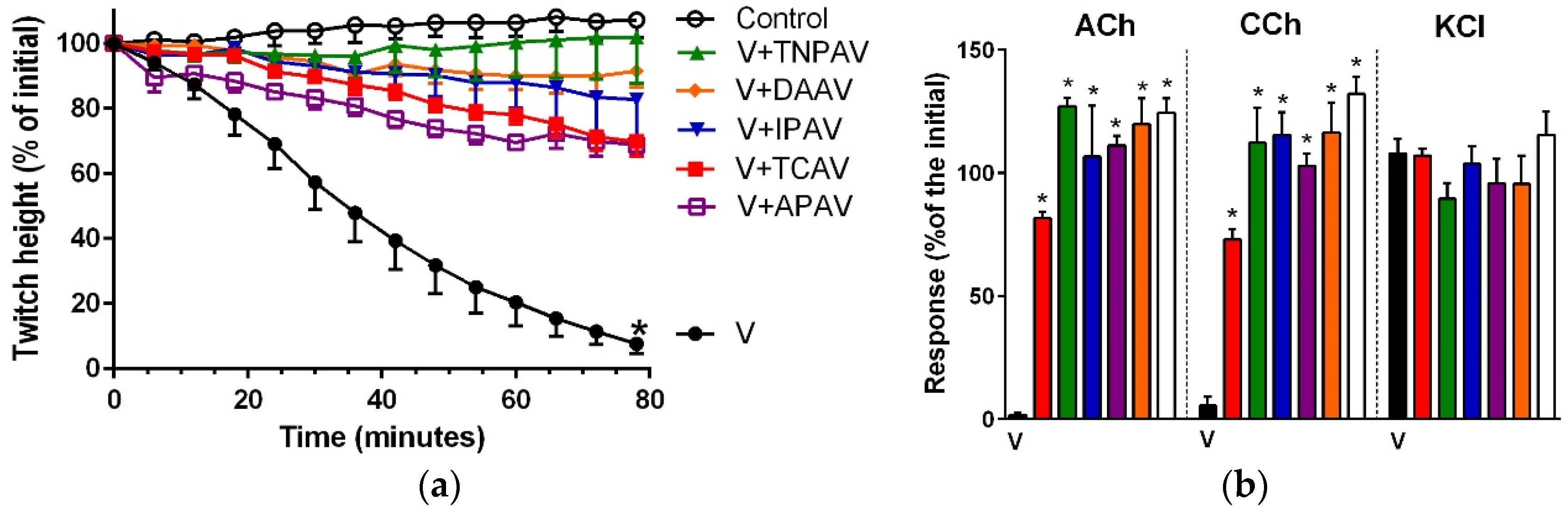
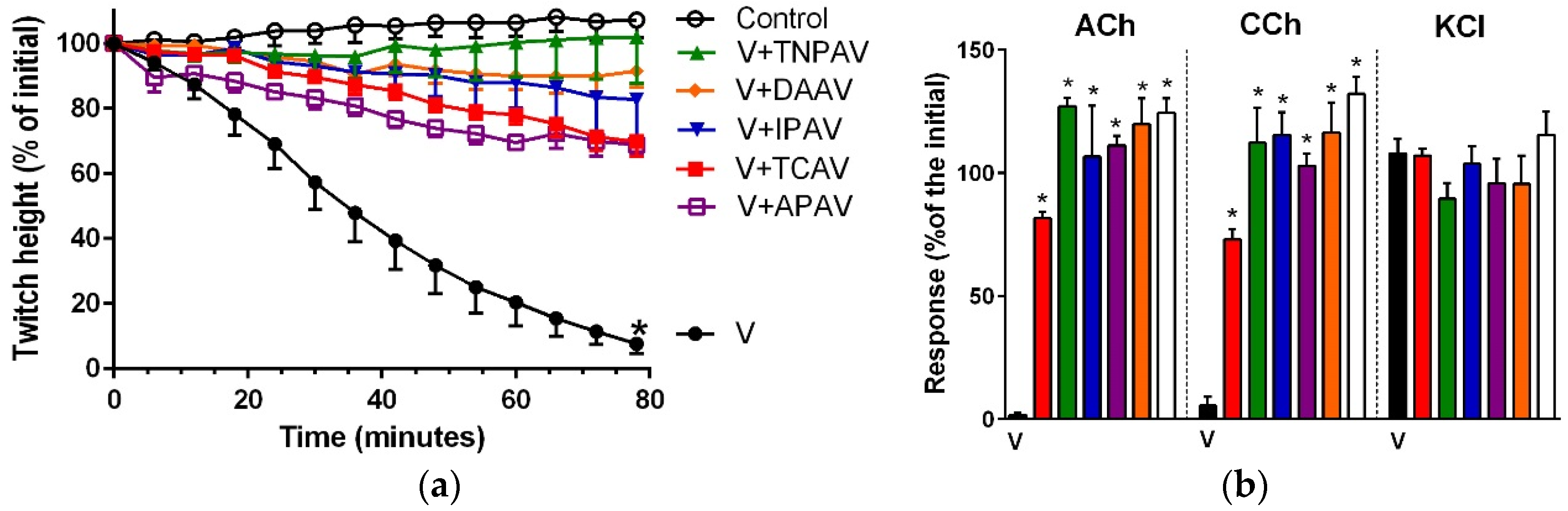
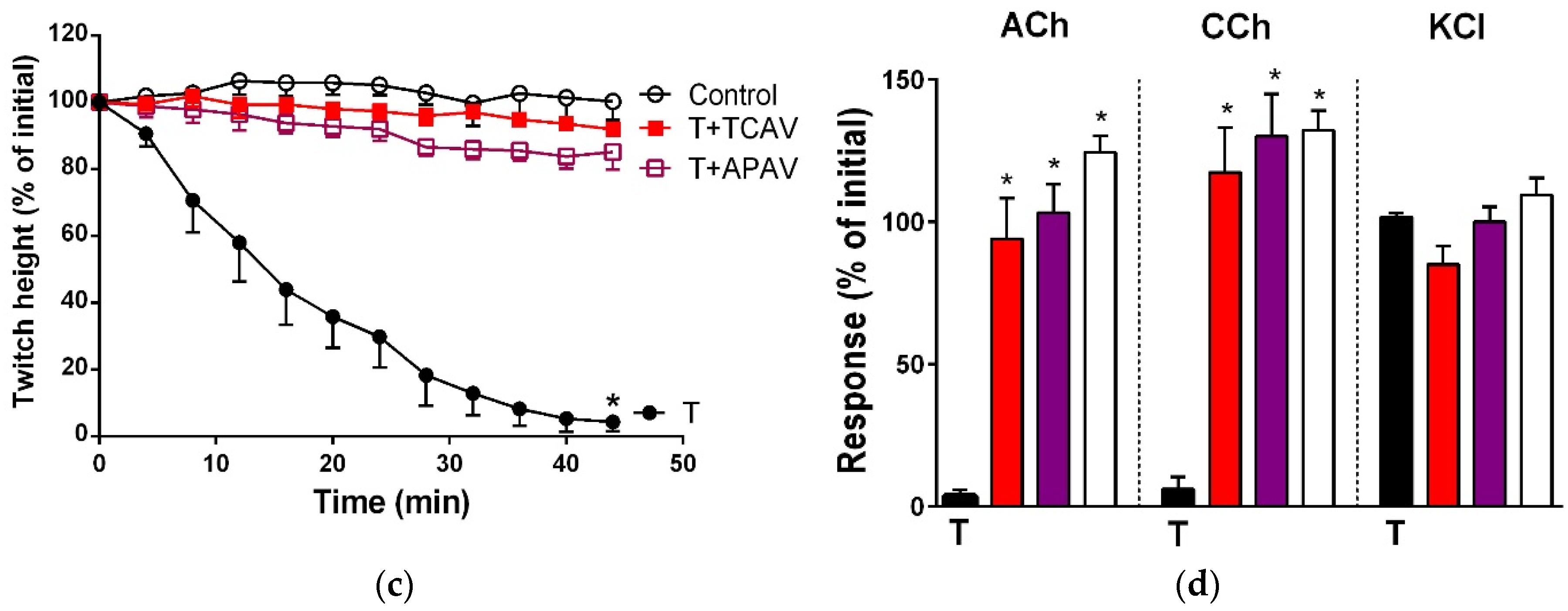
Figure 3.
Cross-neutralisation of the neurotoxicity of Bungarus fasciatus venom (7.5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom. Note: all antivenoms prevented neurotoxicity (twitch height as a percentage of the initial at 78 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and the presence of antivenoms compared to control. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 3.
Cross-neutralisation of the neurotoxicity of Bungarus fasciatus venom (7.5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom. Note: all antivenoms prevented neurotoxicity (twitch height as a percentage of the initial at 78 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and the presence of antivenoms compared to control. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
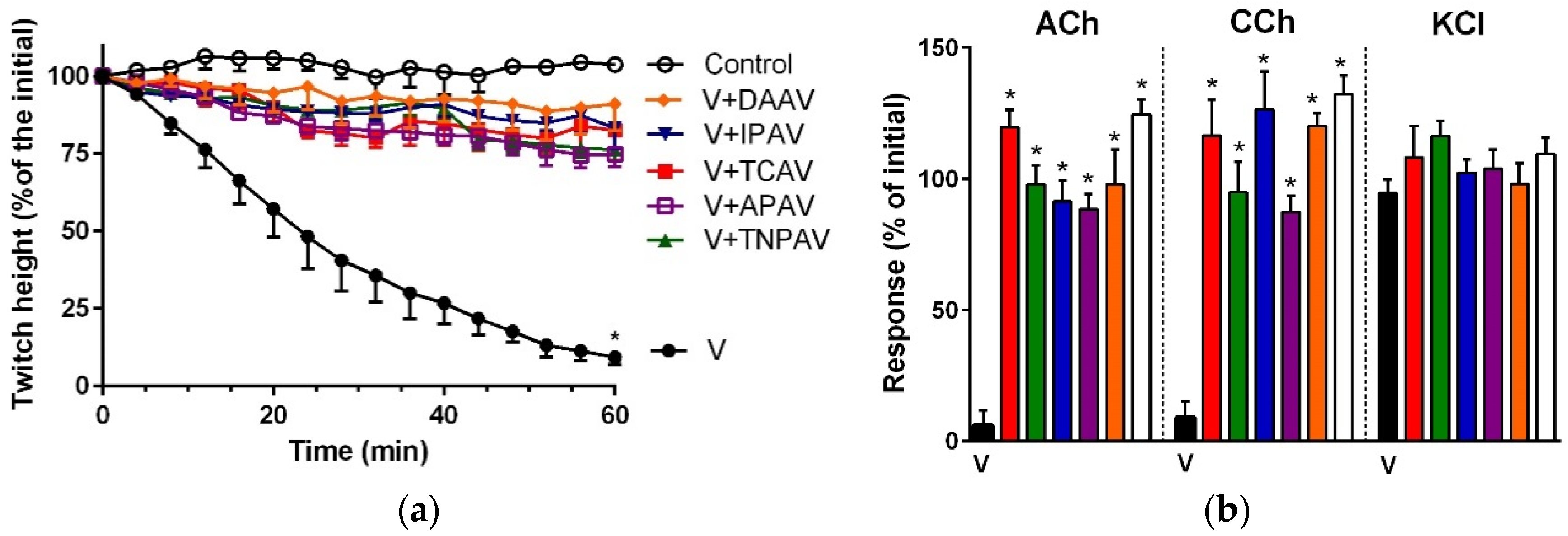
Figure 4.
Cross-neutralisation of the neurotoxicity of Ophiophagus hannah venom (5 µg/mL, V, black) by, Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPVA), and Australian polyvalent antivenom (APAV) in chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms prevented neurotoxicity (twitch height as a percentage of the initial at 60 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and the presence of antivenoms compared to control. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 4.
Cross-neutralisation of the neurotoxicity of Ophiophagus hannah venom (5 µg/mL, V, black) by, Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPVA), and Australian polyvalent antivenom (APAV) in chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms prevented neurotoxicity (twitch height as a percentage of the initial at 60 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and the presence of antivenoms compared to control. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 5.
Cross-neutralisation of the neurotoxicity of Naja kaouthia venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPVA), and Australian polyvalent antivenom (APAV) in chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms except IPAV prevented neurotoxicity (* significantly lower from the control at 44 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and presence of antivenoms compared to control. Note: all antivenoms except IPAV prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 5.
Cross-neutralisation of the neurotoxicity of Naja kaouthia venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), death adder antivenom (DAAV; orange), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPVA), and Australian polyvalent antivenom (APAV) in chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms except IPAV prevented neurotoxicity (* significantly lower from the control at 44 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence of antivenom and presence of antivenoms compared to control. Note: all antivenoms except IPAV prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 6.
Cross-neutralisation of the neurotoxicity of Notechis scutatus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antiveom (IPAV; blue), death adder antivenom (DAAV; orange), and Australian polyvalent antivenom (APAV; purple) in chick biventer nerve-muscle preparation compared to control: (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms except TCAV prevented neurotoxicity (* significantly lower from the control at 48 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of N. scutatus venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to controls. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 6.
Cross-neutralisation of the neurotoxicity of Notechis scutatus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antiveom (IPAV; blue), death adder antivenom (DAAV; orange), and Australian polyvalent antivenom (APAV; purple) in chick biventer nerve-muscle preparation compared to control: (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: all antivenoms except TCAV prevented neurotoxicity (* significantly lower from the control at 48 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of N. scutatus venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to controls. Note: all antivenoms prevented the abolition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 7.
Cross-neutralisation of the neurotoxicity of Oxyuranus scutellatus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), death adder antivenom (DAAV; orange), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control: (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: TCAV and IPAV failed to prevent neurotoxicity (twitch height as a percentage of the initial at 140 min is different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to controls. Note: all antivenoms prevented the inhibition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 7.
Cross-neutralisation of the neurotoxicity of Oxyuranus scutellatus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), death adder antivenom (DAAV; orange), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control: (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: TCAV and IPAV failed to prevent neurotoxicity (twitch height as a percentage of the initial at 140 min is different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to controls. Note: all antivenoms prevented the inhibition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 8.
Cross-neutralisation of the neurotoxicity of Acanthophis antarcticus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), death adder antivenom (DAAV; orange), Australian polyvalent antivenom (APAV; purple) in chick biventer nerve-muscle preparation compared to control: (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: TCAV and IPAV failed to prevent neurotoxicity (twitch height as a percentage of the initial at 48 min is different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to controls. Note: all antivenoms prevented the inhibition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 8.
Cross-neutralisation of the neurotoxicity of Acanthophis antarcticus venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), death adder antivenom (DAAV; orange), Australian polyvalent antivenom (APAV; purple) in chick biventer nerve-muscle preparation compared to control: (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: TCAV and IPAV failed to prevent neurotoxicity (twitch height as a percentage of the initial at 48 min is different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of venom on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to controls. Note: all antivenoms prevented the inhibition of agonist responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
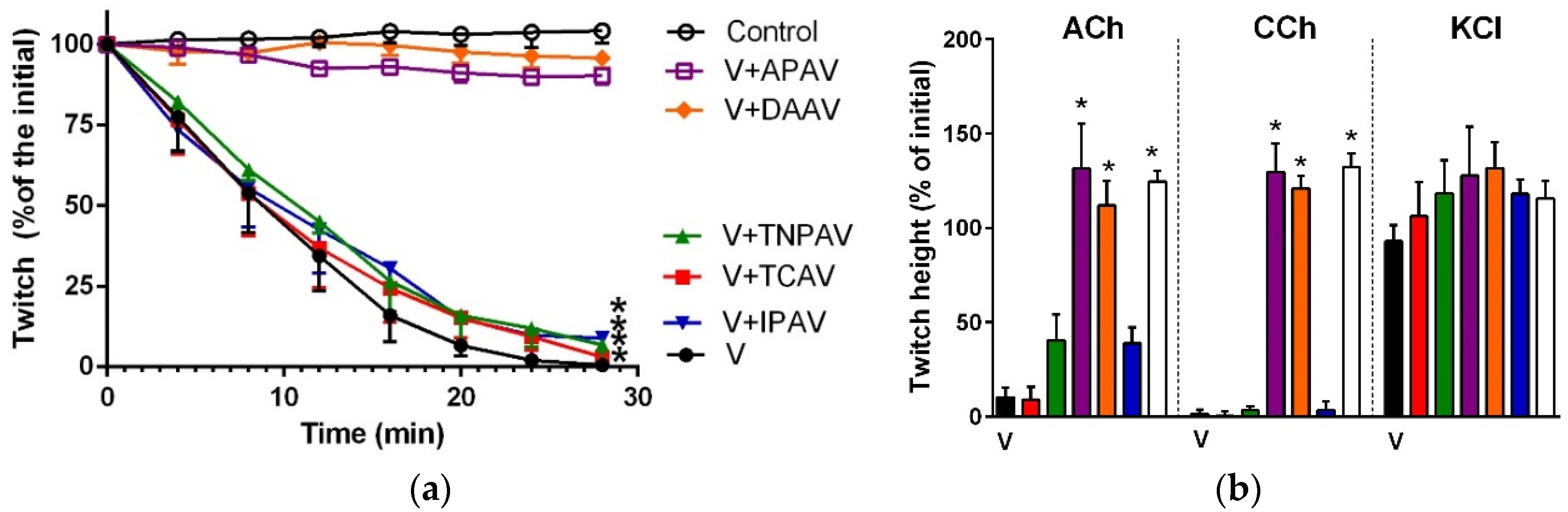
Figure 9.
Cross-neutralisation of the neurotoxicity of Pseudonaja textillis venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), death adder antivenom (DAAV; orange), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: only APAV prevented neurotoxicity (* twitch height as a percentage of the initial at 28 min significantly different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of P. textillis venom on the response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms. Note: only APAV prevented the abolition of ACh and CCh responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 9.
Cross-neutralisation of the neurotoxicity of Pseudonaja textillis venom (5 µg/mL, V, black) by Thai cobra antivenom (TCAV; red), Thai neuro polyvalent antivenom (TNPAV; green), Indian polyvalent antivenom (IPAV; blue), death adder antivenom (DAAV; orange), and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by venom (V). Note: only APAV prevented neurotoxicity (* twitch height as a percentage of the initial at 28 min significantly different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of P. textillis venom on the response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms. Note: only APAV prevented the abolition of ACh and CCh responses by the venom (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
![Toxins 08 00302 g009]()
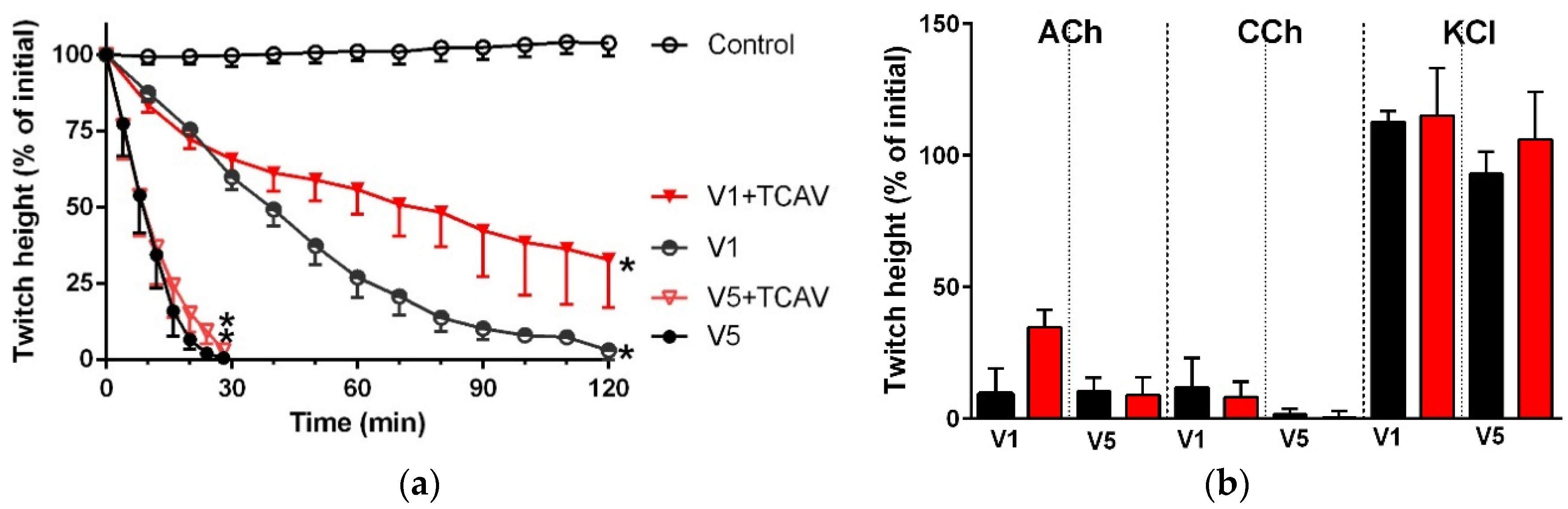
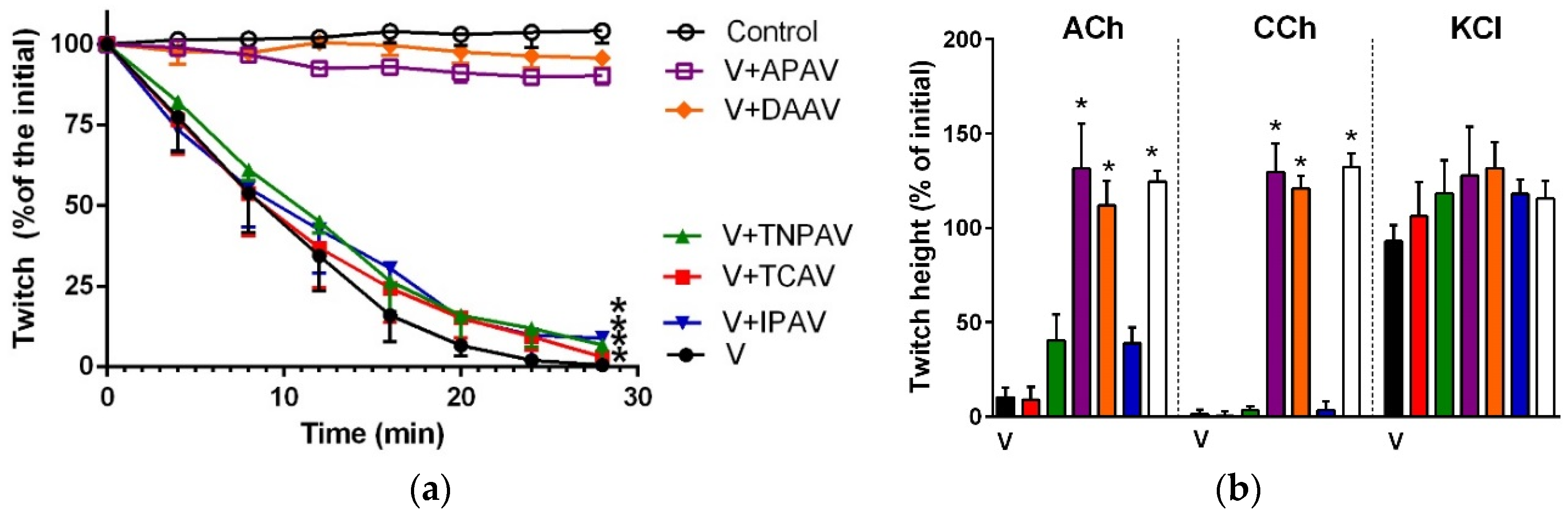
Figure 10.
Cross-neutralisation of the neurotoxicity of 1 µg/mL vs. 5 µg/mL P. textillis venom by Thai cobra antivenom (TCAV; red): (a) Inhibition of indirect twitches caused by 1 µg/mL (V1) vs. 5 µg/mL (V5) P. textillis venom in the presence of TCAV. Note: TCAV does not prevent the twitch abolition caused by 1 µg/mL P. textillis venom (* significantly lower from the control at 120 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of 1 µg/mL (V1) vs. 5 µg/mL (V5) P. textillis venom on the response to exogenous nicotinic agonists (ACh, CCh and KCl) in the absence and presence of TCAV. Note: TCAV failed to prevent the abolition of agonist responses caused by 1 µg/mL venom (no significant difference from the venom, one-way ANOVA followed by Bonferroni’s post-hoc test).
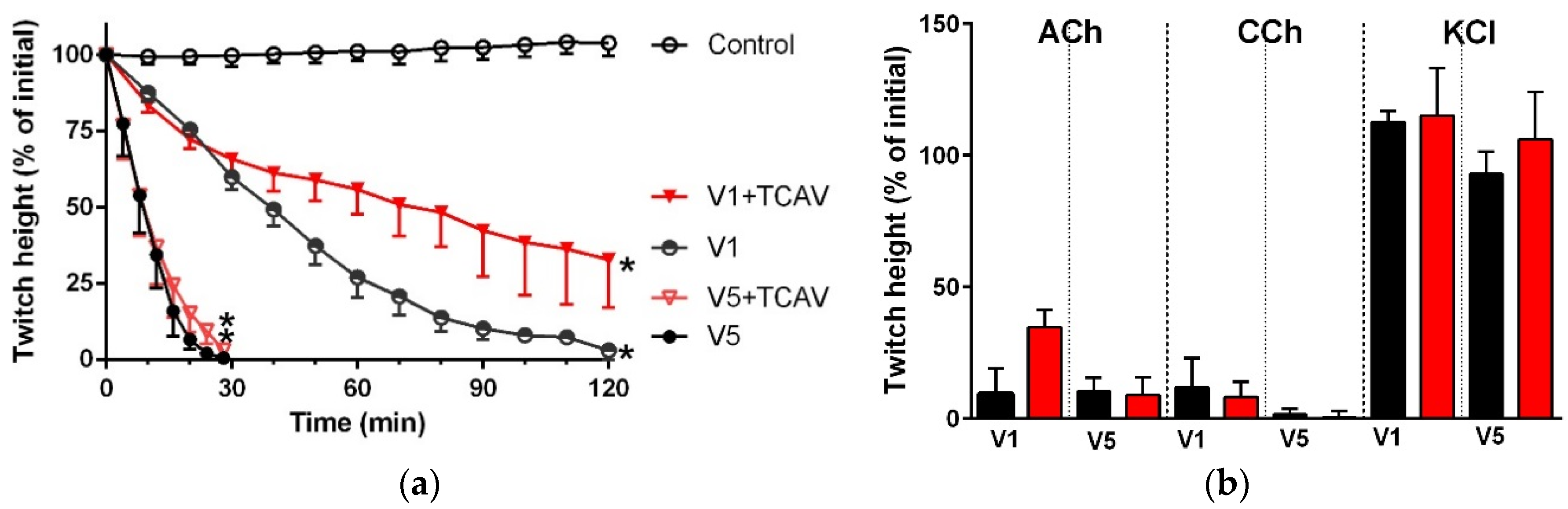
Figure 10.
Cross-neutralisation of the neurotoxicity of 1 µg/mL vs. 5 µg/mL P. textillis venom by Thai cobra antivenom (TCAV; red): (a) Inhibition of indirect twitches caused by 1 µg/mL (V1) vs. 5 µg/mL (V5) P. textillis venom in the presence of TCAV. Note: TCAV does not prevent the twitch abolition caused by 1 µg/mL P. textillis venom (* significantly lower from the control at 120 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of 1 µg/mL (V1) vs. 5 µg/mL (V5) P. textillis venom on the response to exogenous nicotinic agonists (ACh, CCh and KCl) in the absence and presence of TCAV. Note: TCAV failed to prevent the abolition of agonist responses caused by 1 µg/mL venom (no significant difference from the venom, one-way ANOVA followed by Bonferroni’s post-hoc test).
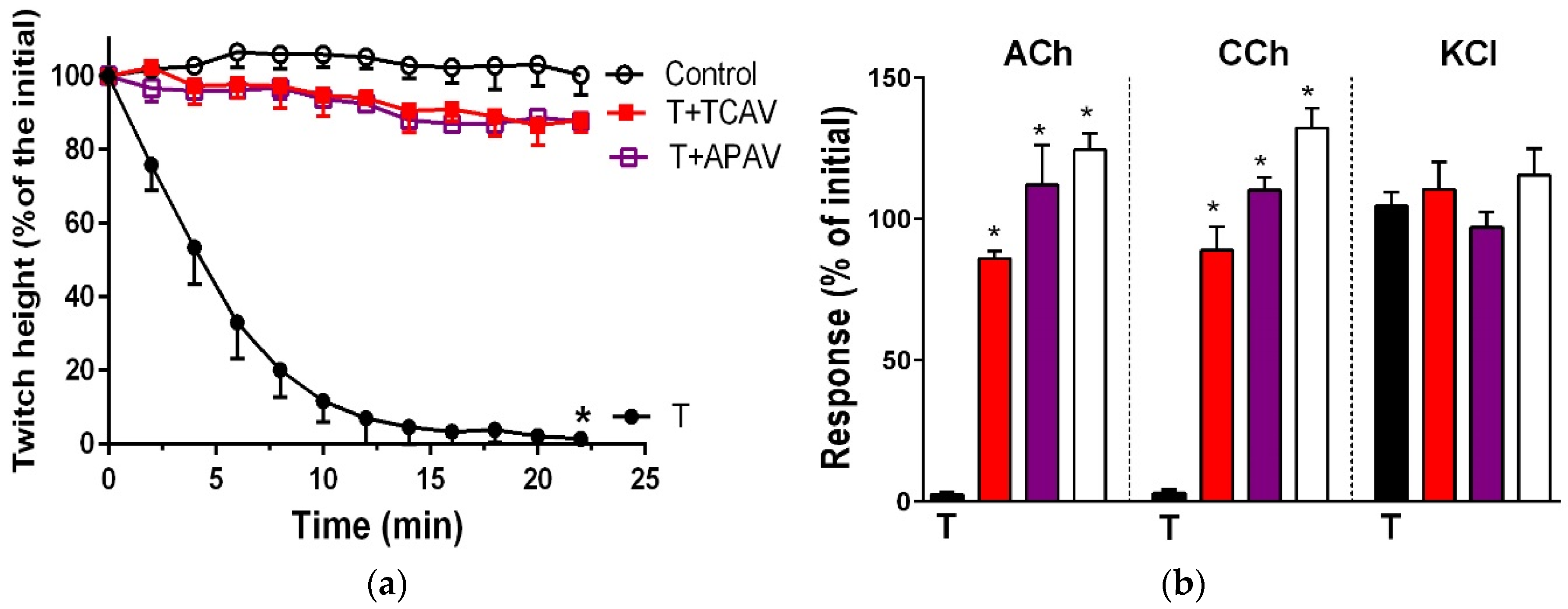
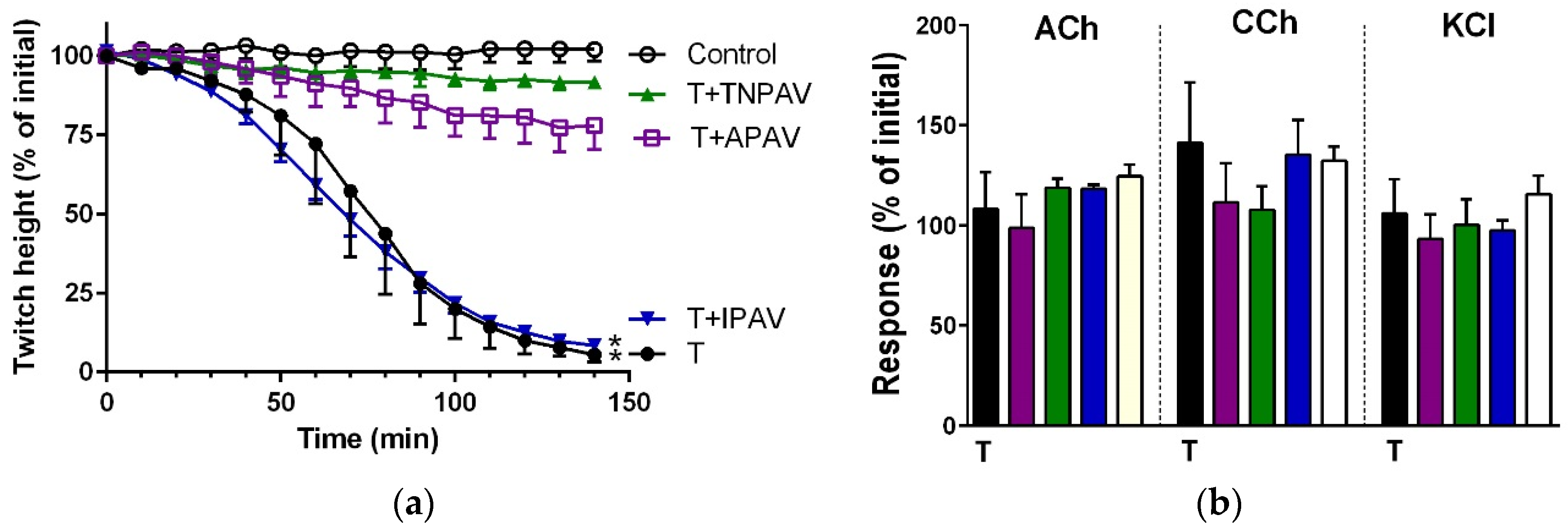
Figure 11.
Cross-neutralisation of the neurotoxicity of the long-chain α-neurotoxins (α-bungarotoxin and α-elapitoxin-Nk2a; black) by Thai cobra antivenom (TCAV; red) and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by 100 nM α-bungarotoxin (T). Note: APAV and TCAV effectively prevented neurotoxicity (no difference from the control at 22 min, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of α-bungarotoxin on response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms. Note: APAV and TCAV prevented the abolition of agonist responses caused by the toxin (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (c) Prevention of the inhibition of indirect twitches caused by 100 nM α-elapitoxin-Nk2a (T). Note: APAV and TCAV prevented neurotoxicity (twitch height as a percentage of the initial at 44 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (d) The effect of 100 nM α-elapitoxin-Nk2a on response to exogenous agonists (ACh, CCh and KCl) in the presence of antivenoms. Note: APAV and TCAV prevented the abolition of agonist responses by the venom (* significantly different from the toxin, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 11.
Cross-neutralisation of the neurotoxicity of the long-chain α-neurotoxins (α-bungarotoxin and α-elapitoxin-Nk2a; black) by Thai cobra antivenom (TCAV; red) and Australian polyvalent antivenom (APAV; purple) in the chick biventer nerve-muscle preparation compared to control (white): (a) Prevention of the inhibition of indirect twitches caused by 100 nM α-bungarotoxin (T). Note: APAV and TCAV effectively prevented neurotoxicity (no difference from the control at 22 min, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of α-bungarotoxin on response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms. Note: APAV and TCAV prevented the abolition of agonist responses caused by the toxin (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (c) Prevention of the inhibition of indirect twitches caused by 100 nM α-elapitoxin-Nk2a (T). Note: APAV and TCAV prevented neurotoxicity (twitch height as a percentage of the initial at 44 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (d) The effect of 100 nM α-elapitoxin-Nk2a on response to exogenous agonists (ACh, CCh and KCl) in the presence of antivenoms. Note: APAV and TCAV prevented the abolition of agonist responses by the venom (* significantly different from the toxin, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
![Toxins 08 00302 g011a]()
![Toxins 08 00302 g011b]()
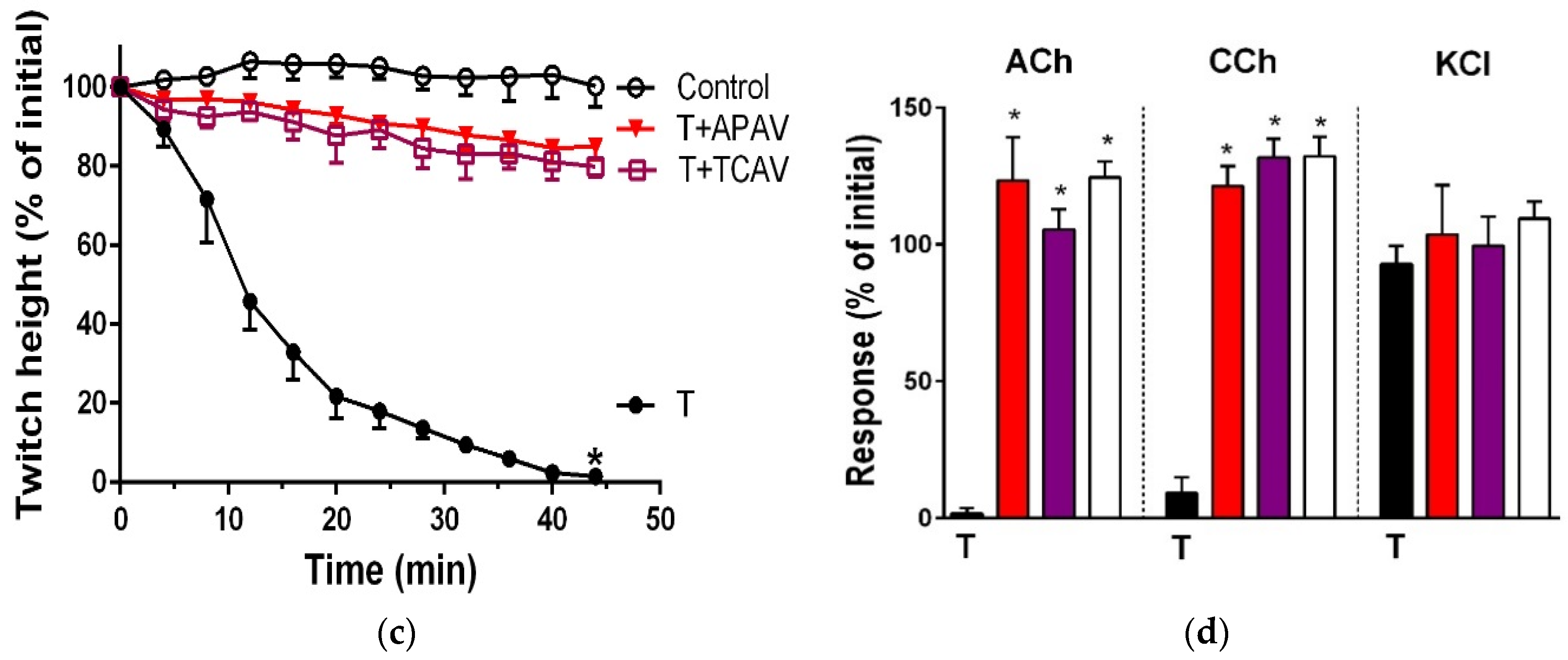
Figure 12.
Cross-neutralisation of the neurotoxicity of the short-chain α-neurotoxins (α-scutoxin and α-elapitoxin-Ppr-1; T, black) by Thai cobra antivenom (TCAV; red) and Australian polyvalent antivenom (APAV; purple) in chick biventer nerve-muscle preparation compared to control (white); (a) Prevention of the inhibition of indirect twitches caused by α-scutoxin (T). Note: APAV and TCAV partially prevented neurotoxicity (* significantly lower from the control and the toxin at 54 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of 100 nM α-scutoxin on the response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to control. Note: APAV and TCAV prevented the abolition of agonist responses caused by the toxin (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (c) Prevention of the inhibition of indirect twitches caused by 100 nM α-elapitoxin-Ppr-1 (T). Note: APAV and TCAV prevented neurotoxicity (twitch height as a percentage of the initial at 44 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (d) The effect of 100 nM α-elapitoxin-Ppr1 on the response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to control. Note: APAV and TCAV prevented the abolition of agonist responses by the venom (* significantly different from the toxin, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 12.
Cross-neutralisation of the neurotoxicity of the short-chain α-neurotoxins (α-scutoxin and α-elapitoxin-Ppr-1; T, black) by Thai cobra antivenom (TCAV; red) and Australian polyvalent antivenom (APAV; purple) in chick biventer nerve-muscle preparation compared to control (white); (a) Prevention of the inhibition of indirect twitches caused by α-scutoxin (T). Note: APAV and TCAV partially prevented neurotoxicity (* significantly lower from the control and the toxin at 54 min, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of 100 nM α-scutoxin on the response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to control. Note: APAV and TCAV prevented the abolition of agonist responses caused by the toxin (* significantly different from the venom, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test); (c) Prevention of the inhibition of indirect twitches caused by 100 nM α-elapitoxin-Ppr-1 (T). Note: APAV and TCAV prevented neurotoxicity (twitch height as a percentage of the initial at 44 min not different from the control, one-way ANOVA followed by Bonferroni’s post-hoc test); (d) The effect of 100 nM α-elapitoxin-Ppr1 on the response to exogenous nicotinic receptor agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to control. Note: APAV and TCAV prevented the abolition of agonist responses by the venom (* significantly different from the toxin, p < 0.05, one-way ANOVA followed by Bonferroni’s post-hoc test).
![Toxins 08 00302 g012a]()
![Toxins 08 00302 g012b]()
Figure 13.
Cross-neutralisation of the neurotoxicity of the pre-synaptic neurotoxin from Oxyuranus scutellatus, taipoxin (100 nM, T, black), by Thai neuro polyvalent antivenom (TNPAV, green), Indian polyvalent antivenom (IPAV, blue) and Australian polyvalent antivenom (APAV, orange) in the chick biventer nerve-muscle preparation: (a) Prevention of the inhibition of indirect twitches caused by 100 nM taipoxin (T). Note: All antivenoms except IPAV effectively prevented the neurotoxicity (* significant difference from the control at 140 min, One-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of taipoxin on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to control. Note: neither the toxin alone nor the toxin in the presence of antivenoms was able to abolish the agonist responses (no significant difference among groups, one-way ANOVA followed by Bonferroni’s post-hoc test).
Figure 13.
Cross-neutralisation of the neurotoxicity of the pre-synaptic neurotoxin from Oxyuranus scutellatus, taipoxin (100 nM, T, black), by Thai neuro polyvalent antivenom (TNPAV, green), Indian polyvalent antivenom (IPAV, blue) and Australian polyvalent antivenom (APAV, orange) in the chick biventer nerve-muscle preparation: (a) Prevention of the inhibition of indirect twitches caused by 100 nM taipoxin (T). Note: All antivenoms except IPAV effectively prevented the neurotoxicity (* significant difference from the control at 140 min, One-way ANOVA followed by Bonferroni’s post-hoc test); (b) The effect of taipoxin on the response to exogenous agonists (ACh, CCh and KCl) in the absence and the presence of antivenoms compared to control. Note: neither the toxin alone nor the toxin in the presence of antivenoms was able to abolish the agonist responses (no significant difference among groups, one-way ANOVA followed by Bonferroni’s post-hoc test).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}