
AaeAP1 and AaeAP2: Novel Antimicrobial Peptides from the Venom of the Scorpion, Androctonus aeneas: Structural Characterisation, Molecular Cloning of Biosynthetic Precursor-Encoding cDNAs and Engineering of Analogues with Enhanced Antimicrobial and Anticancer Activities

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification and Structural Analysis of Mature Novel AMPs from Venom
2.2. Cloning of AMP Precursor-Encoding cDNAs
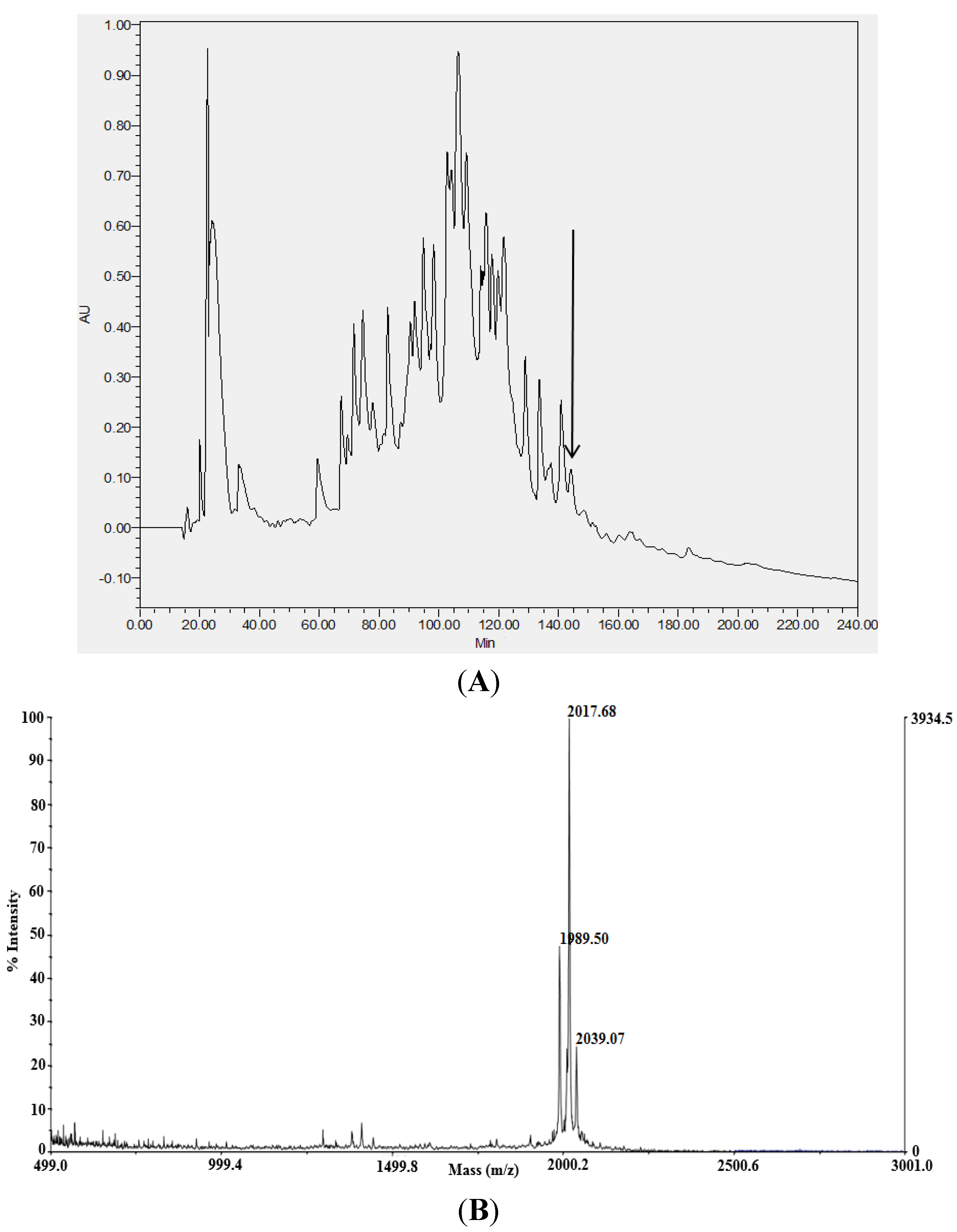

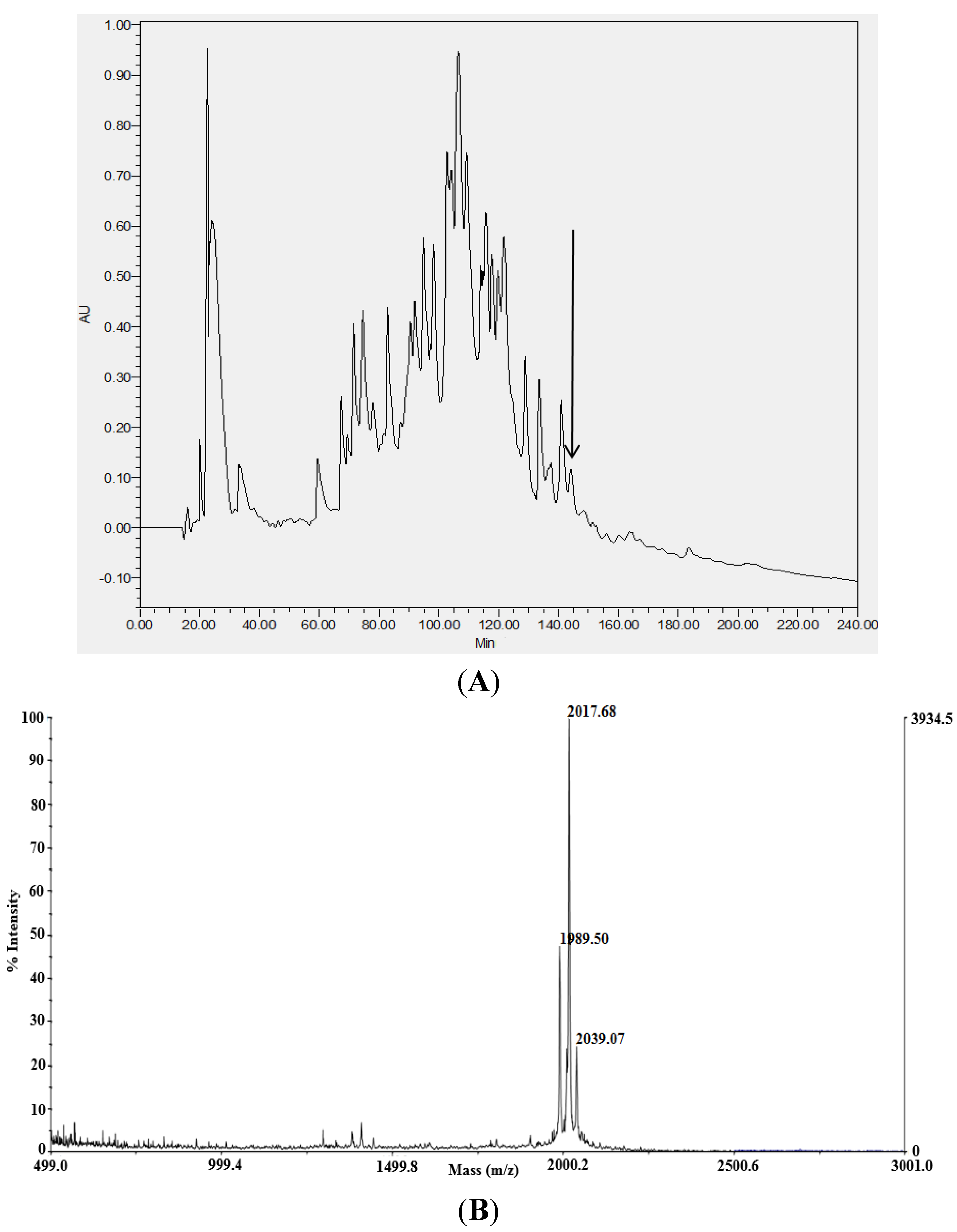{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| #1 | b(1+) | b(2+) | Sequence | y(1+) | y(2+) | #2 |
|---|---|---|---|---|---|---|
| 1 | 148.07570 | 74.54149 | F | - | - | 19 |
| 2 | 261.15977 | 131.08352 | L | 1869.13213 | 935.06970 | 18 |
| 3 | 408.22819 | 204.61773 | F | 1756.04806 | 878.52767 | 17 |
| 4 | 495.26022 | 248.13375 | S | 1608.97964 | 804.99346 | 16 |
| 5 | 608.34429 | 304.67578 | L | 1521.94761 | 761.47744 | 15 |
| 6 | 721.42836 | 361.21782 | I | 1408.86354 | 704.93541 | 14 |
| 7 | 818.48113 | 409.74420 | P | 1295.77947 | 648.39337 | 13 |
| 8 | 905.51316 | 453.26022 | S | 1198.72670 | 599.86699 | 12 |
| 9 | 1004.58158 | 502.79443 | V | 1111.69467 | 556.35097 | 11 |
| 10 | 1117.66565 | 559.33646 | I | 1012.62625 | 506.81676 | 10 |
| 11 | 1188.70277 | 594.85502 | A | 899.54218 | 450.27473 | 9 |
| 12 | 1245.72424 | 623.36576 | G | 828.50506 | 414.75617 | 8 |
| 13 | 1358.80831 | 679.90779 | L | 771.48359 | 386.24543 | 7 |
| 14 | 1457.87673 | 729.44200 | V | 658.39952 | 329.70340 | 6 |
| 15 | 1544.90876 | 772.95802 | S | 559.33110 | 280.16919 | 5 |
| 16 | 1615.94588 | 808.47658 | A | 472.29907 | 236.65317 | 4 |
| 17 | 1729.02995 | 865.01861 | I | 401.26195 | 201.13461 | 3 |
| 18 | 1885.13107 | 943.06917 | R | 288.17788 | 144.59258 | 2 |
| 19 | - | - | C-Amidated | 132.07676 | 66.54202 | 1 |
| #1 | b(1+) | b(2+) | Sequence | y(1+) | y(2+) | #2 |
|---|---|---|---|---|---|---|
| 1 | 148.07570 | 74.54149 | F | - | - | 19 |
| 2 | 261.15977 | 131.08352 | L | 1841.10083 | 921.05405 | 18 |
| 3 | 408.22819 | 204.61773 | F | 1728.01676 | 864.51202 | 17 |
| 4 | 495.26022 | 248.13375 | S | 1580.94834 | 790.97781 | 16 |
| 5 | 608.34429 | 304.67578 | L | 1493.91631 | 747.46179 | 15 |
| 6 | 721.42836 | 361.21782 | I | 1380.83224 | 690.91976 | 14 |
| 7 | 818.48113 | 409.74420 | P | 1267.74817 | 634.37772 | 13 |
| 8 | 905.51316 | 453.26022 | S | 1170.69540 | 585.85134 | 12 |
| 9 | 976.55028 | 488.77878 | A | 1083.66337 | 542.33532 | 11 |
| 10 | 1089.63435 | 545.32081 | I | 1012.62625 | 506.81676 | 10 |
| 11 | 1160.67147 | 580.83937 | A | 899.54218 | 450.27473 | 9 |
| 12 | 1217.69294 | 609.35011 | G | 828.50506 | 414.75617 | 8 |
| 13 | 1330.77701 | 665.89214 | L | 771.48359 | 386.24543 | 7 |
| 14 | 1429.84543 | 715.42635 | V | 658.39952 | 329.70340 | 6 |
| 15 | 1516.87746 | 758.94237 | S | 559.33110 | 280.16919 | 5 |
| 16 | 1587.91458 | 794.46093 | A | 472.29907 | 236.65317 | 4 |
| 17 | 1700.99865 | 851.00296 | I | 401.26195 | 201.13461 | 3 |
| 18 | 1857.09977 | 929.05352 | R | 288.17788 | 144.59258 | 2 |
| 19 | - | - | C-Amidated | 132.07676 | 66.54202 | 1 |

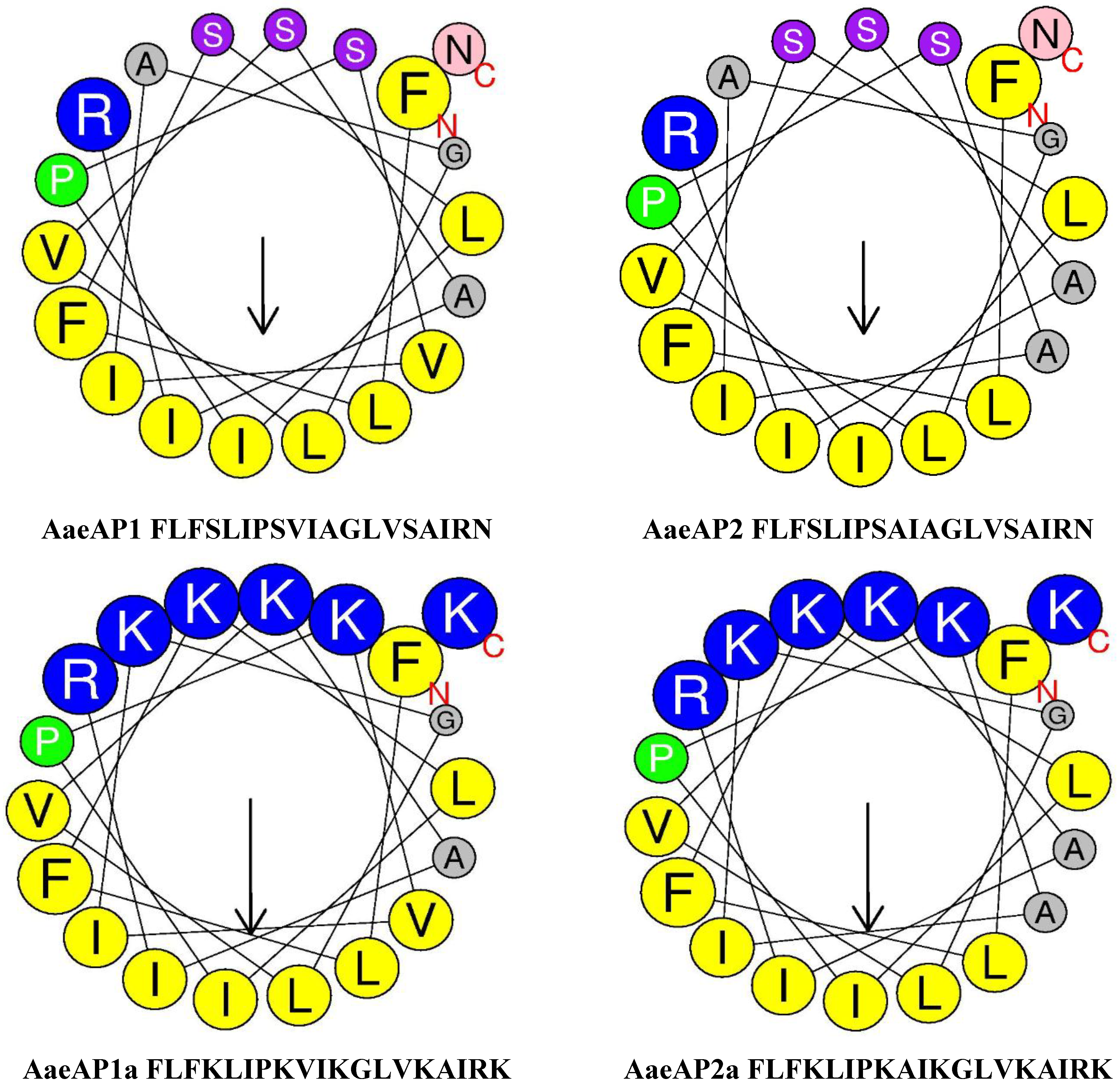
2.3. Secondary Structure Prediction and Physiochemical Properties of AaeAP1 and -2 and Their Cationicity-/Amphipathicity-Enhanced Analogues, AaeAP1a and -2a

| Species | Peptide | Primary structure | Nucleotide sequence database |
|---|---|---|---|
| Androctonus aeneas | AaeAP1 | FLFSLIPSVIAGLVSAIRNa | (Accession No. HG792997) |
| Androctonus aeneas | AaeAP2 | --------A----------a | (Accession No. HG792998) |
| Mesobuthus martensii | Kb1 | --------A-S--I--FKa | (Accession No. AF159979) |
| Mesobuthus eupeus | caerin-like AMP | --------A-S--I--FKa | (Accession No. KC108907) |
| Androctonus amoreuxi | AamAP1 | -------HA-G--I--FKa | (Accession No. FR821613) |
| Isometrus maculatus | imcroporin | -F---L--L-G------Ka | (Accession No. FJ750949) |
| Lychas mucronatus | mucroporin | --G----L-G-----FKa | (Accession No. EU669864) |
| Mesobuthus martensii | BmKb2 | --S-----A-S--I--FKa | (Accession No. AF543048) |
| Tityus costatus | AMP clone 5 | -F------L-G---F--Ka | (Accession No. AY740687) |
| Androctonus amoreuxi | AamAP2 | -P-----HA-GG-ISAIKa | (Accession No. FR821614) |
| Peptides | Secondary structure | α-Helix (%) | Hydrophobicity (H) | Hydrophobic moment (μH) | Net charge |
|---|---|---|---|---|---|
| AaeAP1 | FLFSLIPSVIAGLVSAIRN cccccccchhhheeeeeec | 21.05 | 0.849 | 0.455 | +2 |
| AaeAP2 | FLFSLIPSAIAGLVSAIRN cccccccchhhheeeeeec | 21.05 | 0.801 | 0.428 | +2 |
| AaeAP1a | FLFKLIPKAIKGLVKAIRK ccceecchhhhhheeeeec | 31.58 | 0.610 | 0.669 | +7 |
| AaeAP2a | FLFKLIPKVIKGLVKAIRK ccceecchhhhhheeeeec | 31.58 | 0.562 | 0.641 | +7 |
2.4. Antimicrobial/Haemolytic Activities of AaeAP1 and -2 and Their Cationicity-/Amphipathicity-Enhanced Analogues, AaeAP1a and -2a
| Peptides | MIC (mg/L) | MBC (mg/L) 100% hemolysis (mg/L) | |||||
|---|---|---|---|---|---|---|---|
| S. aureus | E. coli | C. albicans | S. aureus | E. coli | C. albicans | Horse red cells | |
| AaeAP1 | 16 | >512 | 32 | 32 | NT | 64 | 16 |
| AaeAP2 | 16 | >512 | 32 | 16 | NT | 64 | 64 |
| AaeAP1a | 4 | 16 | 4 | 32 | 32 | 16 | 32 |
| AaeAP2a | 4 | 16 | 4 | 32 | 32 | 16 | 64 |
| Peptide | Primary structure | Mass (Da) |
|---|---|---|
| AaeAP1 | FLFSLIPSVIAGLVSAIRNamide | 2016.18 |
| AaeAP2 | FLFSLIPSAIAGLVSAIRNamide | 1986.15 |
| AaeAP1a | FLFKLIPKVIKGLVKAIRKamide | 2209.48 |
| AaeAP2a | FLFKLIPKAIKGLVKAIRKamide | 2181.45 |
| * * * * * |
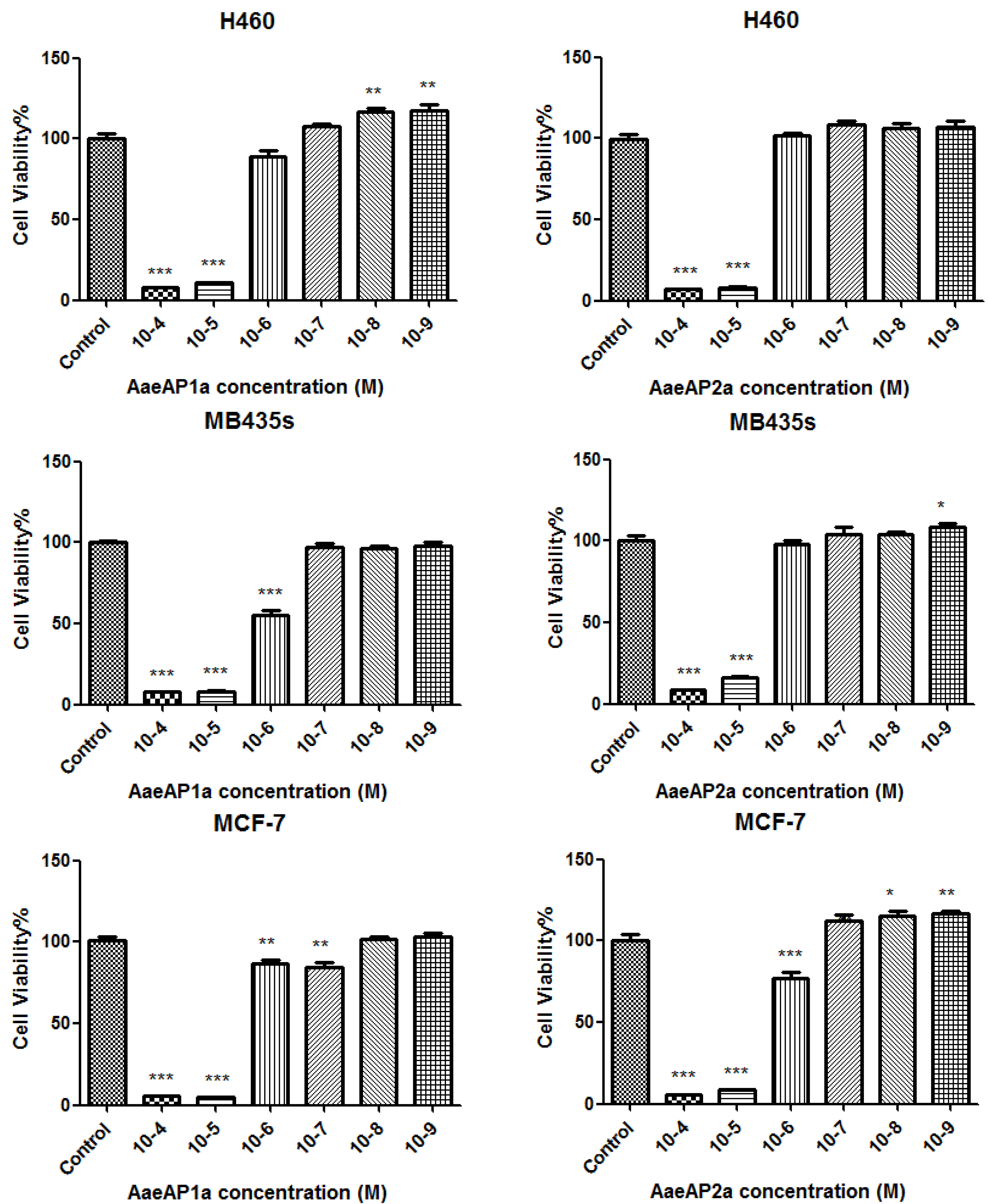
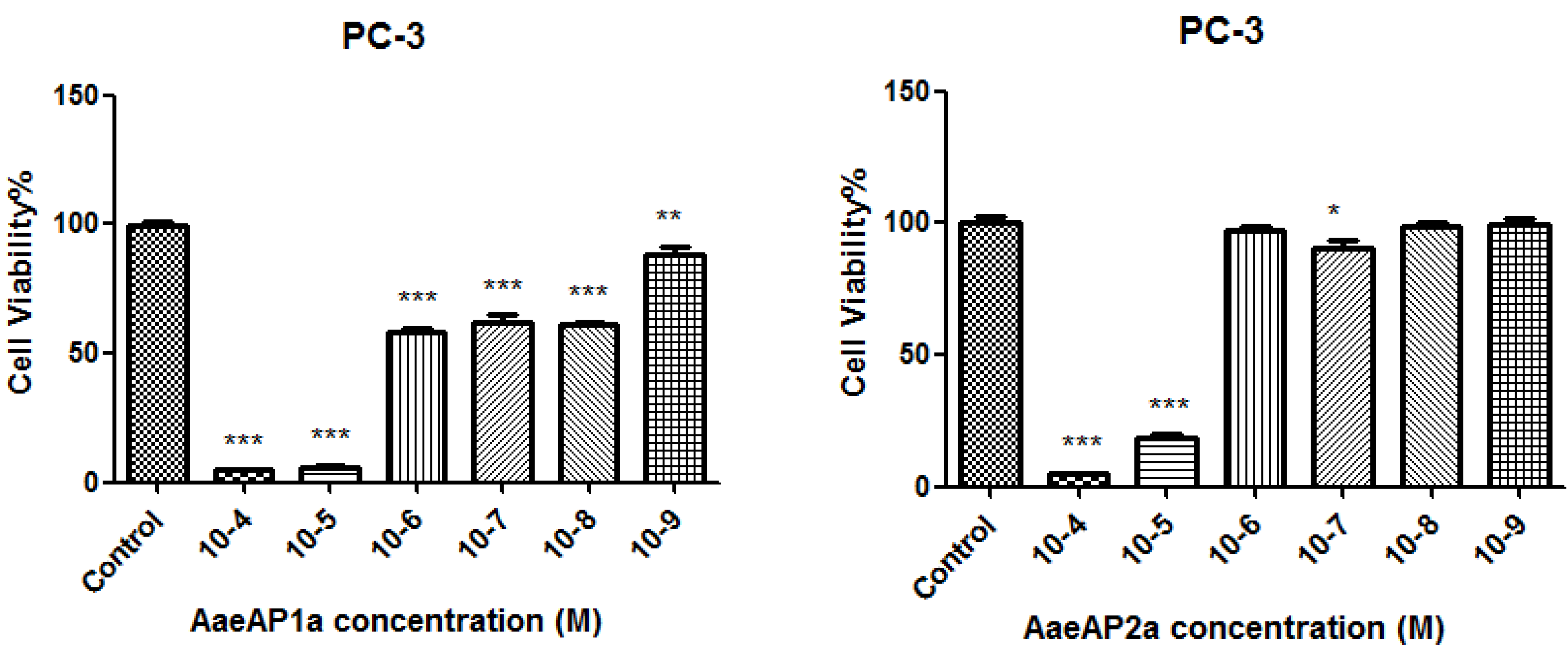
2.5. The Antiproliferative Effects of AaeAP1 and -2 and Their Cationicity-/Amphipathicity-Enhanced Analogues, AaeAP1a and -2a, against a Panel of Human Cancer Cell Lines
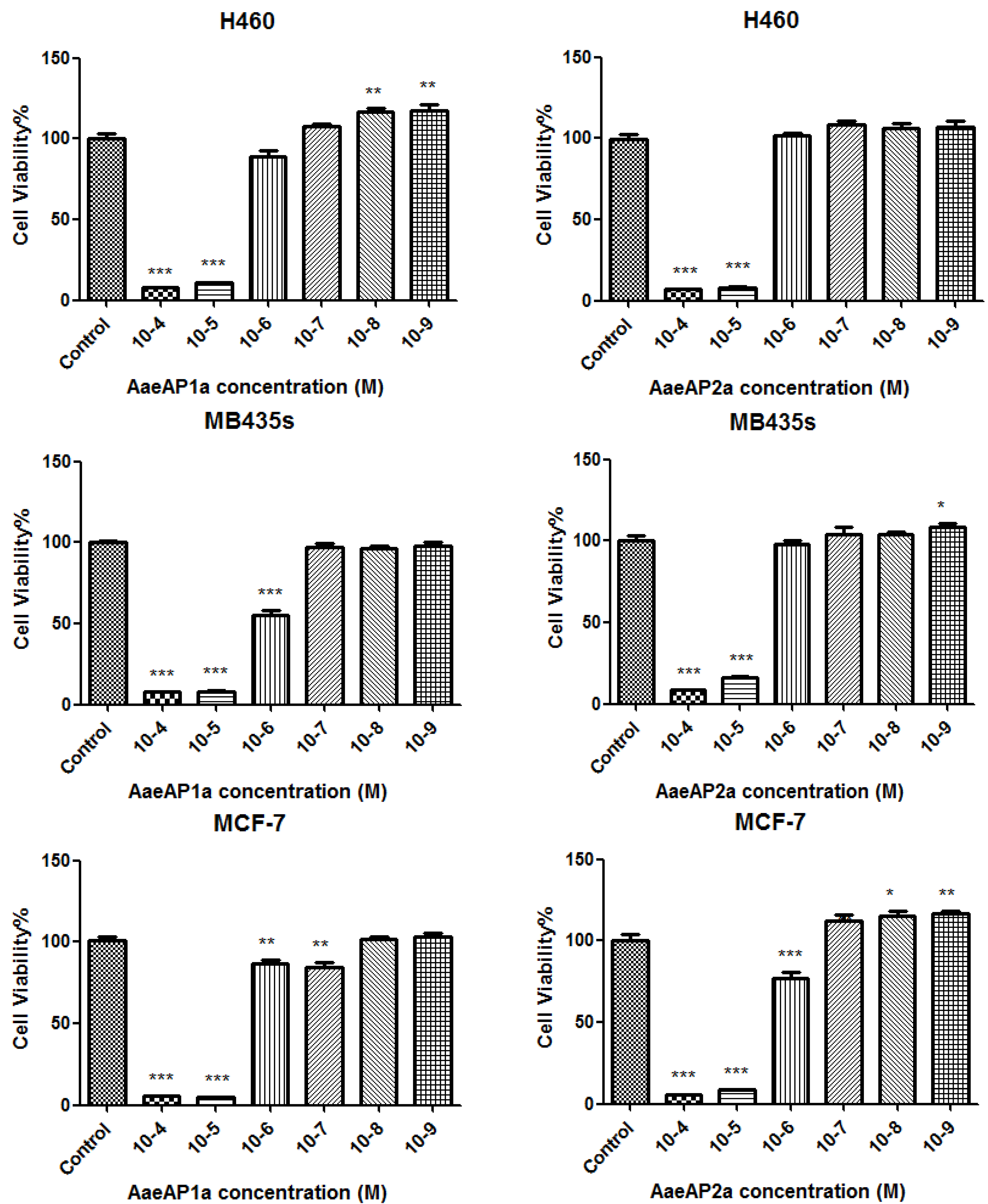
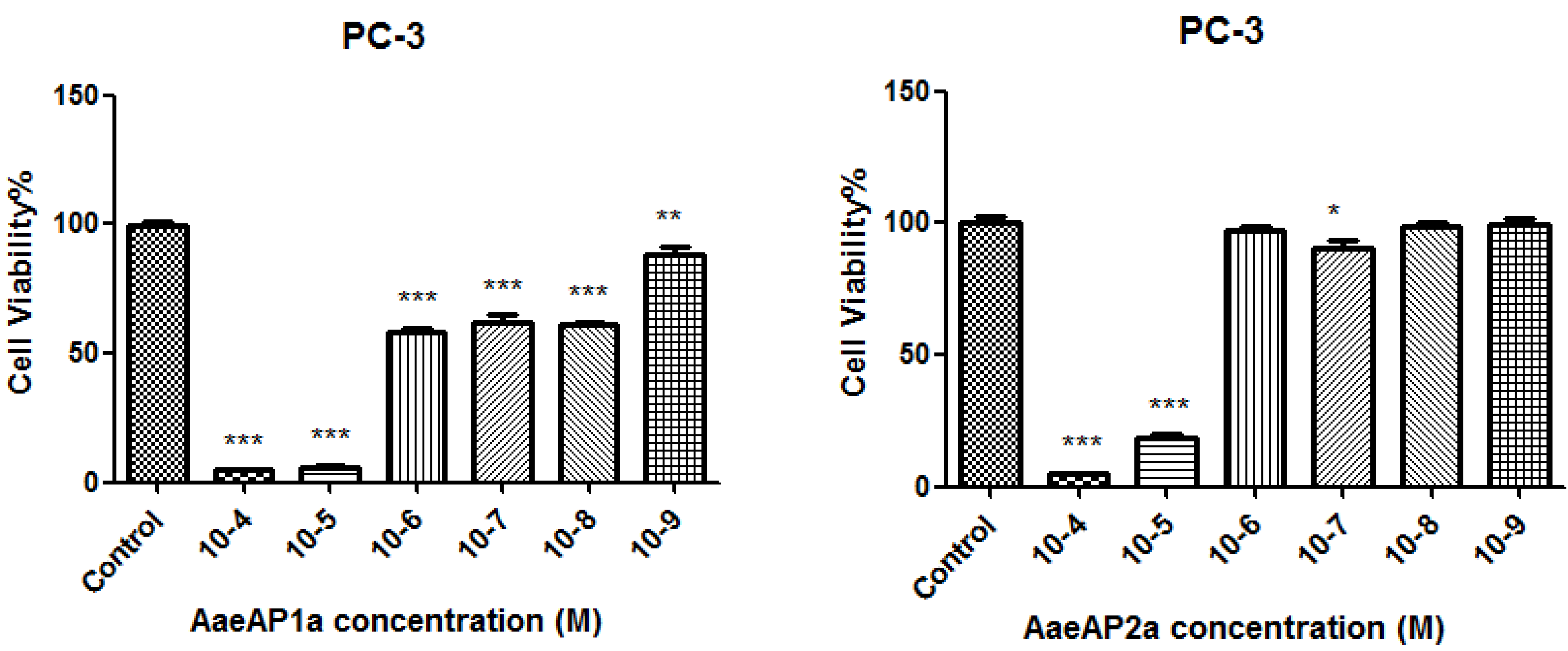
3. Discussion
4. Experimental Section
4.1. Acquisition of Scorpion Venom
4.2. Isolation and Structural Characterisation of Antimicrobial Activity from Lyophilised Venom
4.3. Antimicrobial Screening Assays
4.4. Molecular Cloning of Antimicrobial Peptide Biosynthetic Precursor-Encoding cDNAs from the Venom-Derived cDNA Library
4.5. Prediction of Putative AMP Secondary Structures and Physicochemical Properties
4.6. Peptide Synthesis and Purification
4.7. Antimicrobial Minimal Inhibitory Concentration and Minimum Bactericidal Assays
4.8. Haemolysis Assay
4.9. Culture and Maintenance of Human Cancer Cell Lines
4.10. Assessment of Anti-Proliferative Effects of AMPs on Human Cancer Cells Using the MTT Cell Viability Assay
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Quintero-Hernández, V.; Jiménez-Vargas, J.M.; Gurrola, G.B.; Valdivia, H.H.; Possani, L.D. Scorpion venom components that affect ion-channels function. Toxicon 2013, 76, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.N.; Gopalakrishnakone, P.; Tan, P.T.; Chew, K.C.; Cheng, B.; Kini, R.M.; Koh, J.L.; Seah, S.H.; Brusic, V. SCORPION, a molecular database of scorpion toxins. Toxicon 2002, 40, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.T.; Veeramani, A.; Srinivasan, K.N.; Ranganathan, S.; Brusic, V. SCORPION2: A database for structure-function analysis of scorpion toxins. Toxicon 2006, 47, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez de la Vega, R.C.; Schwartz, E.F.; Possani, L.D. Mining on scorpion venom biodiversity. Toxicon 2010, 56, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2001. [Google Scholar]
- Ashcroft, F. Ion Channels and Disease; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Almaaytah, A.; Albalas, Q. Scorpion venom peptides with no disulfide bridges: A review. Peptides 2014, 51, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Chhatwal, G.S.; Habermann, E. Neurotoxins, protease inhibitors and histamine releasers in the venom of the Indian red scorpion (Buthus tamulus): Isolation and partial characterization. Toxicon 1981, 19, 807–823. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Yasuda, A.; Naoki, H.; Corzo, G.; Andriantsiferana, M.; Nakajima, T. IsCT, a novel cytotoxic linear peptide from scorpion Opisthacanthus madagascariensis. Biochem. Biophys. Res. Commun. 2001, 286, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Arpornsuwan, T.; Buasakul, B.; Jaresitthikunchai, J.; Roytrakul, S. Potent and rapid antigonococcal activity of the venom peptide BmKn2 and its derivatives against different Maldi biotype of multidrug-resistant Neisseria gonorrhoeae. Peptides 2014, 53, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Polis, G.A. The Biology of Scorpions; Stanford University Press: Stanford, CA, USA, 1990. [Google Scholar]
- Possani, L.D. Structure of scorpion toxins. In Handbook of Natural Toxins; Tu, A.T., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1984; Volume 2, pp. 513–550. [Google Scholar]
- Froy, O.; Gurevitz, M. New insight on scorpion divergence inferred from comparative analysis of toxin structure, pharmacology and distribution. Toxicon 2003, 42, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Martin-Eauclaire, M.F.; Couraud, F. Scorpion neurotoxins: Effects and mechanisms. In Handbook of Neurotoxicology; Chang, L.W., Dyer, R.S., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 683–716. [Google Scholar]
- Goyffon, M.; Landon, C. Scorpion toxins and defensins. C. R. Seances Soc. Biol. Fil. 1998, 192, 445–462. [Google Scholar] [PubMed]
- Dehesa-Davila, M.; Alagon, A.C.; Possini, L.D. Clinical toxicology of scorpion stings. In Handbook of Clinical Toxicology of Animal Venoms and Poisons; Meier, J., White, J., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 221–238. [Google Scholar]
- Quan, D. North American poisonous bites and stings. Crit. Care Clin. 2012, 28, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Skolnik, A.B.; Ewald, M.B. Pediatric scorpion envenomation in the United States: Morbidity, mortality, and therapeutic innovations. Pediatr. Emerg. Care 2013, 29, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Harrison, P.L.; Abdel-Rahman, M.A.; Miller, K.; Strong, P.N. Antimicrobial peptides from scorpion venoms. Toxicon 2014, 88, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Torres-Larios, A.; Gurrola, G.B.; Zamudio, F.Z.; Possani, L.D. Hadrurin, a new antimicrobial peptide from the venom of the scorpion Hadrurus aztecus. Eur. J. Biochem. 2000, 267, 5023–5031. [Google Scholar] [CrossRef] [PubMed]
- Conde, R.; Zamudio, F.Z.; Rodríguez, M.H.; Possani, L.D. Scorpine, an anti-malaria and anti-bacterial agent purified from scorpion venom. FEBS Lett. 2000, 471, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Hou, X.; Ge, L.; Li, R.; Zhou, M.; Wang, H.; Wang, L.; Wei, M.; Chen, T.; Shaw, C. Cationicity-Enhanced Analogues of the Antimicrobial Peptides, AcrAP1 and AcrAP2, from the Venom of the Scorpion, Androctonus crassicauda, Display Potent Growth Modulation Effects on Human Cancer Cell Lines. Int. J. Biol. Sci. 2014, 10, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, C.; Du, Q.; Wei, R.; Wang, L.; Zhou, M.; Chen, T.; Shaw, C. Two peptides, TsAP-1 and TsAP-2, from the venom of the Brazilian yellow scorpion, Tityus serrulatus: Evaluation of their antimicrobial and anticancer activities. Biochimie 2013, 95, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Tian, C.; Zhu, S. Inducible antibacterial response of scorpion venom gland. Peptides 2007, 28, 2299–2305. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez de la Vega, R.C.; García, B.I.; D’Ambrosio, C.; Diego-García, E.; Scaloni, A.; Possani, L.D. Antimicrobial peptide induction in the haemolymph of the Mexican scorpion Centruroides limpidus limpidus in response to septic injury. Cell Mol. Life Sci. 2004, 61, 1507–1519. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Xia, L.; Ng, T.B. A review of defensins of diverse origins. Curr. Protein Pept. Sci. 2007, 8, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Dennison, S.R.; Harris, F.; Bhatt, T.; Singh, J.; Phoenix, D.A. The effect of C-terminal amidation on the efficacy and selectivity of antimicrobial and anticancer peptides. Mol. Cell Biochem. 2009, 332, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Dawson, R.M.; Liu, C.Q. Properties and applications of antimicrobial peptides in biodefense against biological warfare threat agents. Crit. Rev. Microbiol. 2008, 34, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Engström, A.; Andersson, K.; Steiner, H.; Bennich, H.; Boman, H.G. Insect immunity. Attacins a family of antibacterial proteins from Hyalophora cecropia. EMBO J. 1983, 2, 571–576. [Google Scholar]
- Gautier, R.; Douquet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific. α-helical properties. Bioinformatics 2008, 24, 2010–2012. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, T.; Zhou, M.; Wang, L.; Shaw, C. PsT-1: A new tryptophyllin peptide from the skin secretion of Waxy Monkey Leaf Frog, Phyllomedusa sauvagei. Regul. Pept. 2013, 184, 14–21. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Q.; Hou, X.; Wang, L.; Zhang, Y.; Xi, X.; Wang, H.; Zhou, M.; Duan, J.; Wei, M.; Chen, T.; et al. AaeAP1 and AaeAP2: Novel Antimicrobial Peptides from the Venom of the Scorpion, Androctonus aeneas: Structural Characterisation, Molecular Cloning of Biosynthetic Precursor-Encoding cDNAs and Engineering of Analogues with Enhanced Antimicrobial and Anticancer Activities. Toxins 2015, 7, 219-237. https://doi.org/10.3390/toxins7020219
Du Q, Hou X, Wang L, Zhang Y, Xi X, Wang H, Zhou M, Duan J, Wei M, Chen T, et al. AaeAP1 and AaeAP2: Novel Antimicrobial Peptides from the Venom of the Scorpion, Androctonus aeneas: Structural Characterisation, Molecular Cloning of Biosynthetic Precursor-Encoding cDNAs and Engineering of Analogues with Enhanced Antimicrobial and Anticancer Activities. Toxins. 2015; 7(2):219-237. https://doi.org/10.3390/toxins7020219
Chicago/Turabian StyleDu, Qiang, Xiaojuan Hou, Lei Wang, Yingqi Zhang, Xinping Xi, Hui Wang, Mei Zhou, Jinao Duan, Minjie Wei, Tianbao Chen, and et al. 2015. "AaeAP1 and AaeAP2: Novel Antimicrobial Peptides from the Venom of the Scorpion, Androctonus aeneas: Structural Characterisation, Molecular Cloning of Biosynthetic Precursor-Encoding cDNAs and Engineering of Analogues with Enhanced Antimicrobial and Anticancer Activities" Toxins 7, no. 2: 219-237. https://doi.org/10.3390/toxins7020219