1. Introduction
Staphylococcal food poisoning (SFP) is one of the most common food-borne intoxication diseases, caused by enterotoxins produced mainly by
Staphylococcus aureus (
S. aureus) strains. Staphylococcal enterotoxins have been the cause of 6.4% of food-borne outbreaks in the European Union (EU) in 2012, placing bacterial toxins as the third most common outbreak causative agent in the EU [
1]. In the USA,
S. aureus was ranked as one of the five most frequent causes of food-borne outbreaks with more than 240,000 illnesses annually [
2]. This reflects how staphylococcal outbreaks are not only a public health problem, but also an economical challenge for the social health system in developed countries.
Until now, 22 staphylococcal enterotoxins (SEs) and enterotoxin-like proteins (SEls) are known [
3]. They differ on the encoding genetic element (
i.e., plasmids, prophages, staphylococcal pathogenicity islands (SaPIs), vSa genomic islands or the staphylococcal cassette chromosome SCC), molecular weight (22–29 kDa), amino acid sequence or mode of action [
3,
4]. All SEs though, have been characterized with superantigenic activity, proteinase resistance and high stability in a wide range of pH and temperature [
3,
5,
6]. The latter features make them extremely challenging for food safety, since they can persist even after food has been processed or cooked.
The enterotoxin involved in around 80% of SFP outbreaks is enterotoxin A (SEA) [
3,
5,
7]. Unlike most other enterotoxins, SEA is not regulated by the accessory gene regulator (
agr) or the staphylococcal accessory regulator (
sar) and thus still poses a challenge regarding its regulatory mechanism. What is known is that the gene encoding for SEA, the
sea gene, is located on the genome of
Siphoviridae bacteriophages. These phages are temperate bacteriophages closely related to lambda (λ) and their life cycle is characterized by two phases, the lysogenic and the lytic phase [
8]. During lysogeny the phage DNA is integrated into the bacterial chromosome and is steadily transferred through generations. In the lytic phase, the phage genome is excised from the chromosome, circularizes and replicates using the cell’s machinery. These dsDNA genome copies are often referred to as the replicative form (RF). Ultimately new phages will be formed that will lyse the bacterial cell [
9,
10].
Borst and Betley in 1994 [
11] were the first to observe differences on the SEA levels produced by different
S. aureus strains (high and low SEA-producing strains) and suggested an association with the
sea-carrying prophage. Later, Wallin-Carlquist
et al. [
12] showed the existence of two
sea variants,
sea1 and
sea2, where high SEA-producing strains bore the
sea1 variant while
sea2 was found in low SEA producing strains. In 2012, Cao
et al. [
13] demonstrated that the life cycle of the
sea-carrying phages influences the
sea gene expression and the levels of SEA produced by
S. aureus strains. It was also proved that some high SEA-producing strains had the ability to produce increased amounts of SEA when their cultures were subjected to prophage inducing conditions using mitomycin C (MMC). Accordingly, the high SEA-producing group was divided into two sub-groups, the inducible one, where higher SEA levels were observed and the non-inducible one, with no impact of induction on the SEA levels. A long
sea transcript, as it was designated, was also detected and quantified. It was presumed to originate from a latent promoter (P
2), located upstream the endogenous
sea promoter (P
1). This long transcript could only be detected in the inducible high SEA-producing strains.
It is known from studies on the λ-phage that the switch from lysogenic to lytic phase is controlled by the cell’s SOS response mechanism and specifically by the RecA activator protein [
14]. When the SOS response is activated the RecA protein directly stimulates auto-proteolysis of the lytic repressor,
cI, which in turn allows transcription of the anti-repressor,
cro, that induces the lytic cycle of the phage [
9,
10,
15], where transcription of relevant early and late phage lytic genes is initiated. The
sea gene is located in the late gene region on the
Siphoviridae phage genome; downstream the late lytic promoters, and therefore transition to the lytic phase could potentially activate and/or even enhance its transcription along with the phage’s late lytic genes.
The aim of the present study was to determine the role of the Siphoviridae phages on sea gene transcription and SEA production especially by identifying and characterizing the nature of the link between the phage life cycle and SEA produced. The impact of S. aureus’ SOS response mechanism on the phage life cycle and SEA production was investigated. Five S. aureus strains representing the three different genetic lines of the sea region were grown and thoroughly investigated under optimal and MMC induced laboratory growth conditions. A recA-disruption mutant strain was constructed and studied under the same growth conditions. To understand how the behavior of the sea-carrying phages affects the SEA produced by different strains, the increase in phage RF copies was explicitly followed, to our knowledge, for the first time in S. aureus and correlated with that of the sea gene copies. The presence of sea transcripts under both control and MMC induced conditions was recorded and correlated time wise with the presence of RF in the cells and the amounts of SEA produced. By specifically monitoring RF the impact of the phage life cycle on sea transcription and SEA production was directly demonstrated. A significant (p < 0.05) increase in RF copy levels was detected under induced conditions, and this was further related to the activation of the SOS response mechanism of S. aureus.
3. Discussion
The role of the
sea-carrying phages’ life cycle and impact of prophage induction on the regulation of
sea gene transcription, has been discussed in previous studies of Sumby and Waldor [
17] and Cao
et al. [
13] where a phage activated long
sea transcript was linked with increased SEA production. However, the transition from lysogeny to the lytic mode in the life cycle of temperate bacteriophages brings about changes both on the transcription pattern of phage-associated genes, and their copy numbers available for potential transcription, through the increase of RF levels.
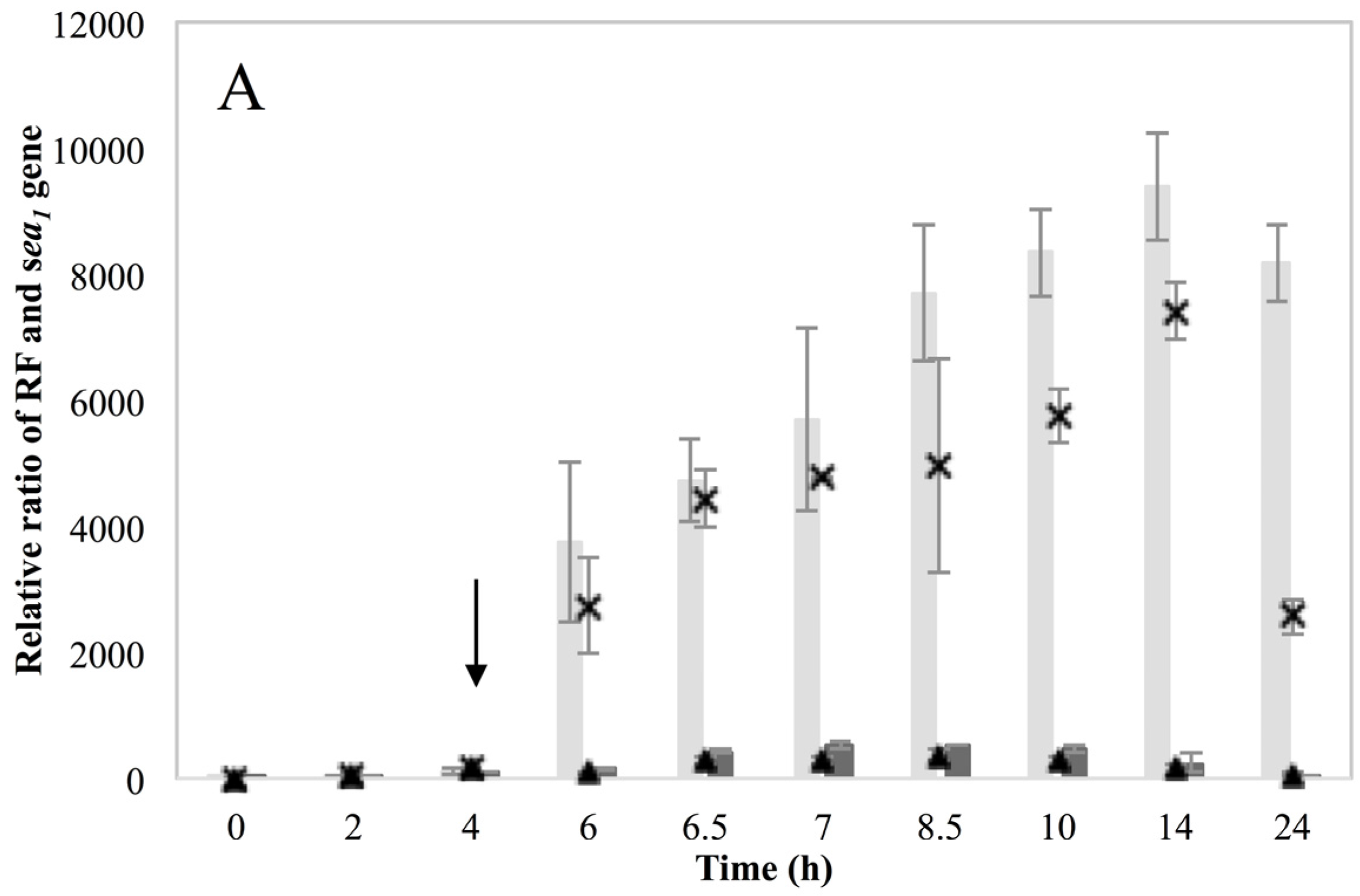
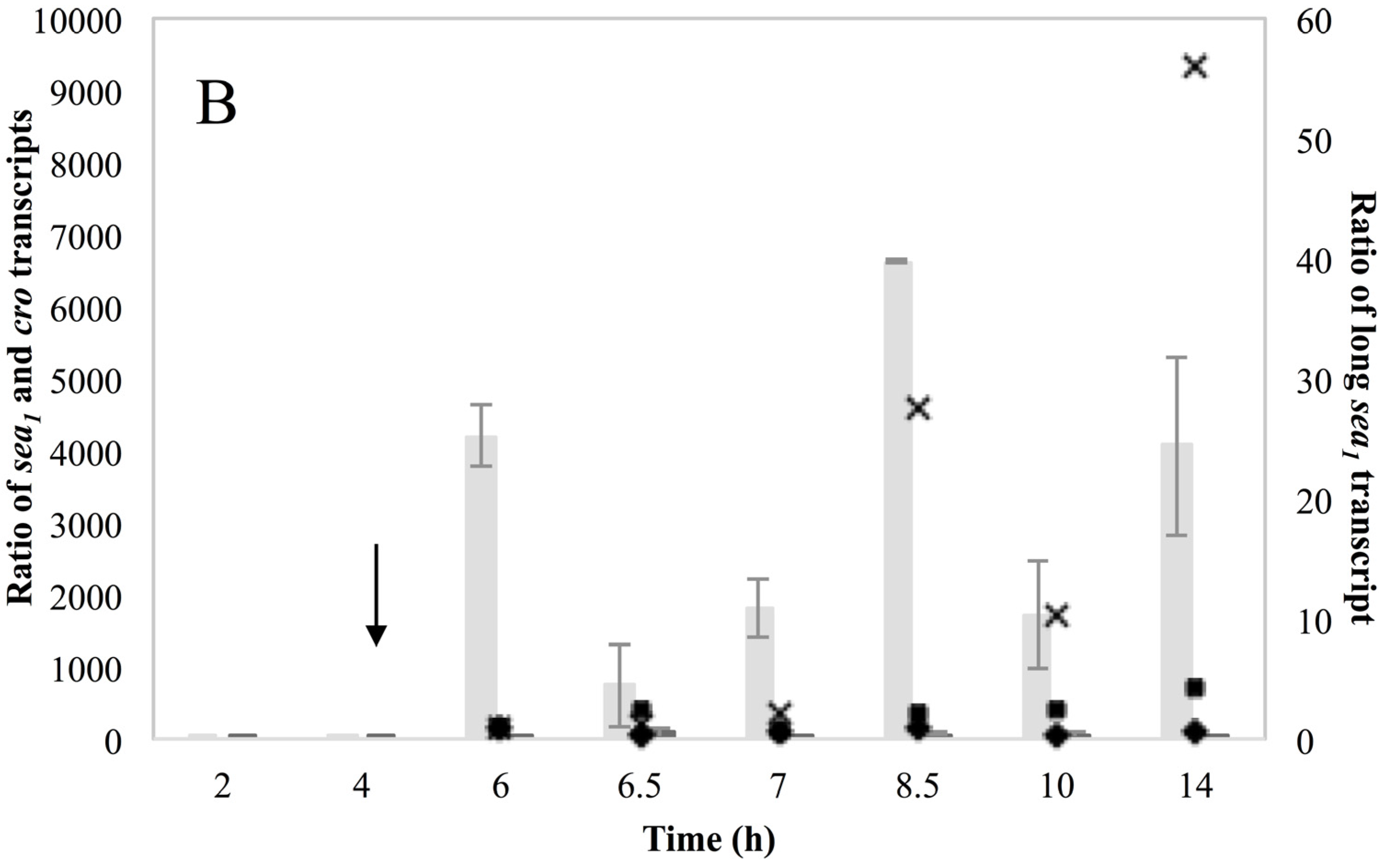
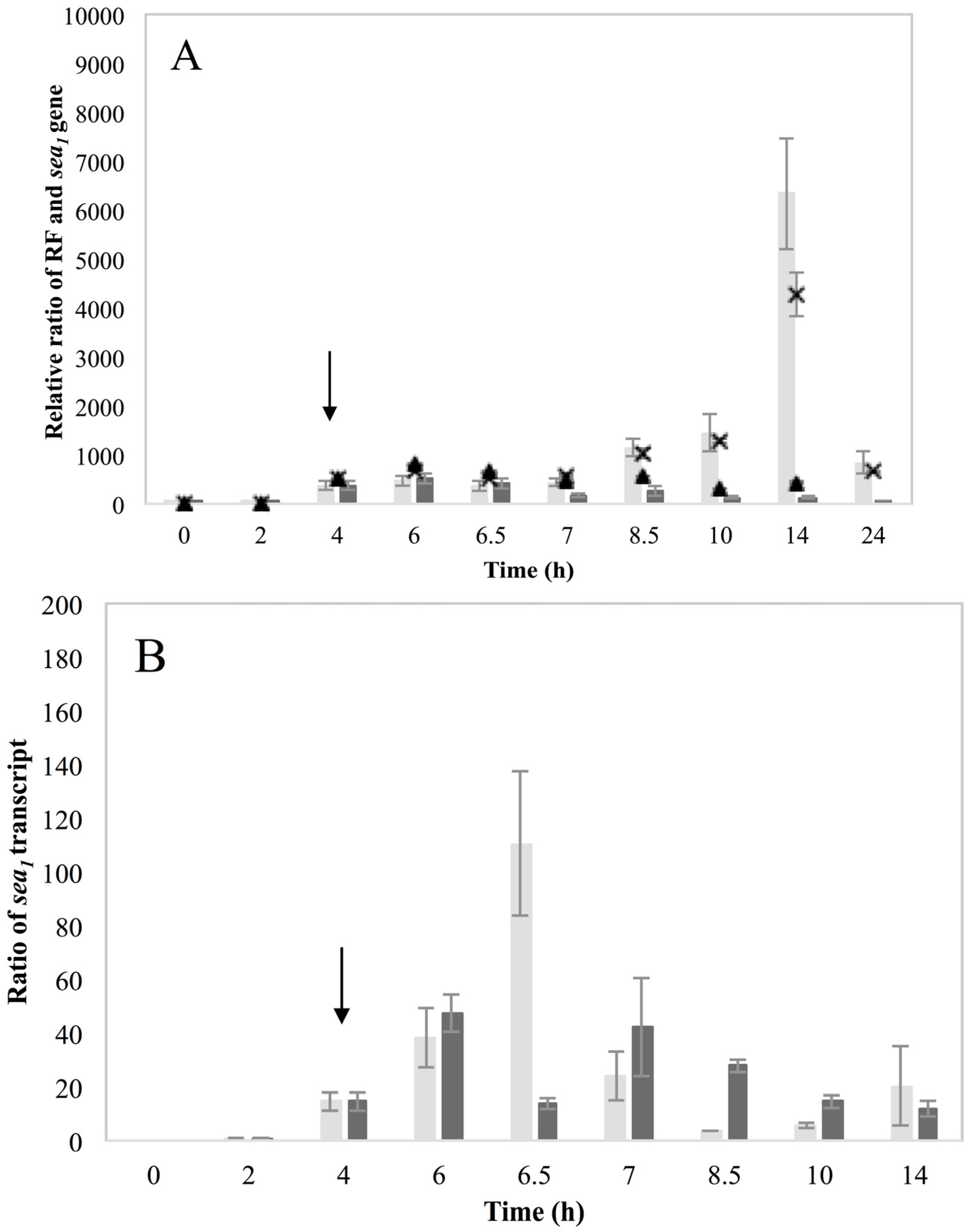
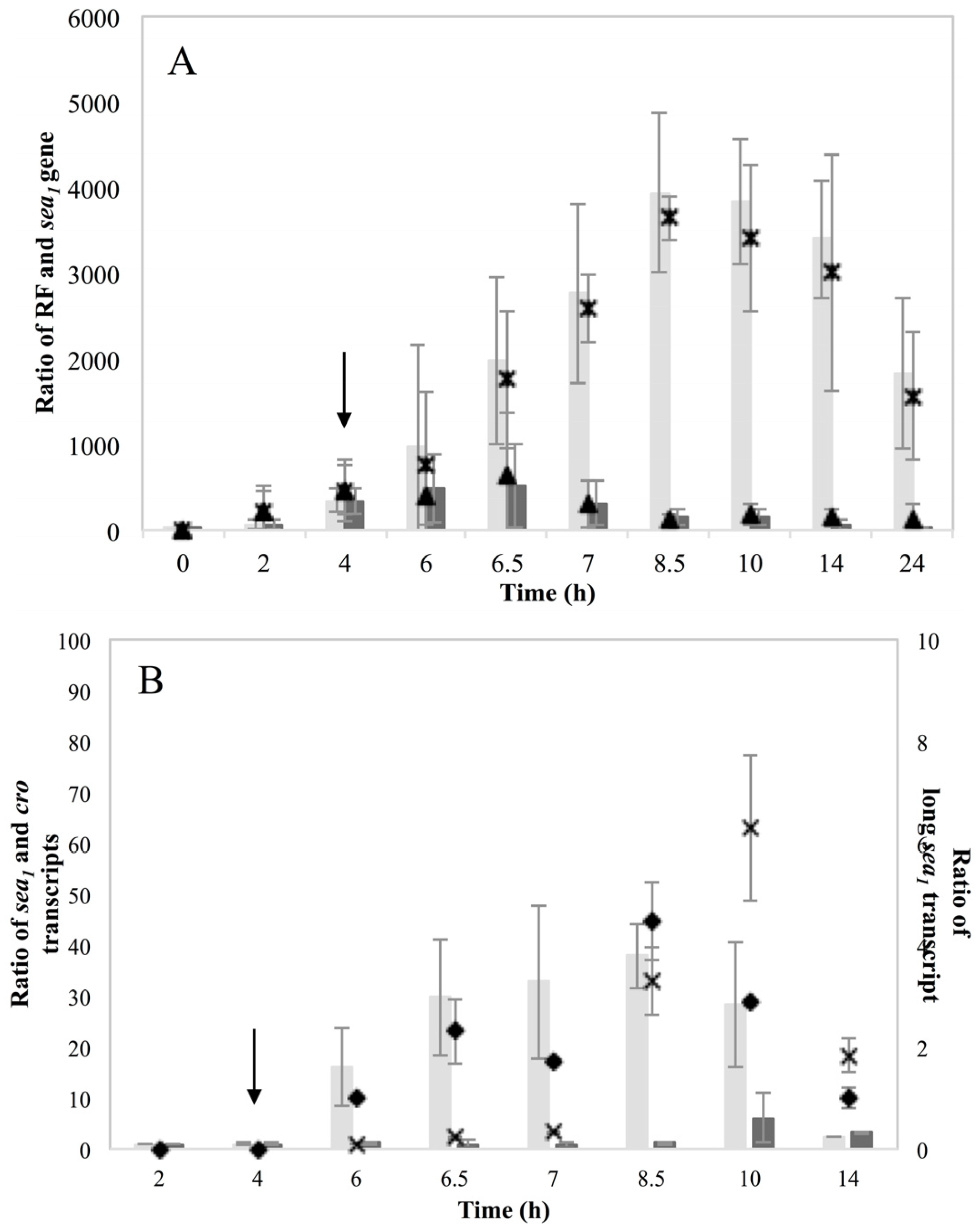
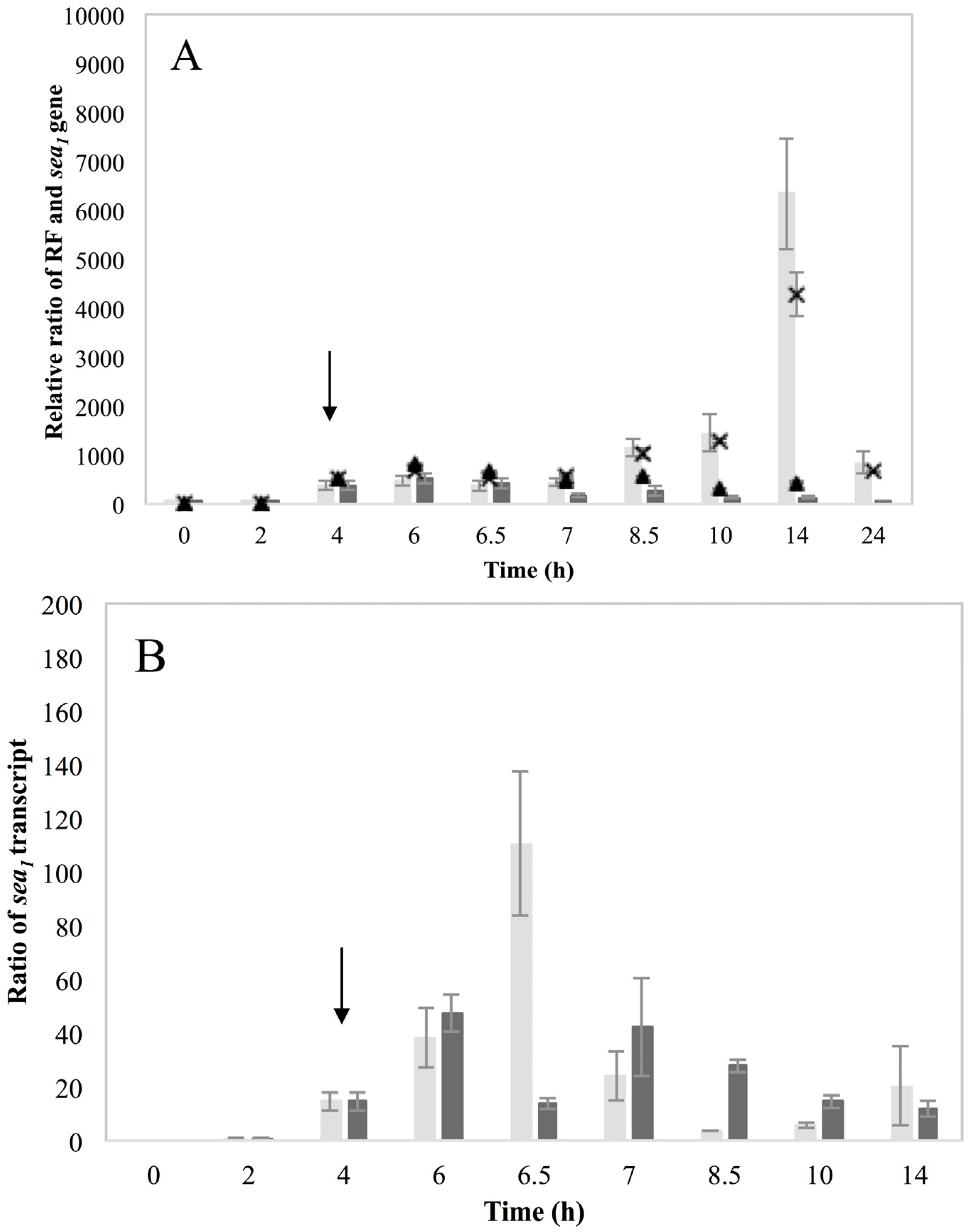
Investigation of RF levels in the present study concluded that the inducible high SEA-producing strains (Sa17 and Sa48), exhibited marked increase in the levels of RF copies after prophage induction, in a pattern that corresponded to the increase in the levels of
sea1 gene copies of the induced cells. This observation demonstrates the impact of prophage induction on
S. aureus virulence, as excision and replication of the phage genome increases the
sea gene copies in the cells. Detection of the long
sea1 transcript and the increase of total
sea1 transcripts noted, further shows the series of events leading to high SEA production by Sa17 and Sa48, after prophage induction. The observed similar transcriptional behaviour of
cro and the long
sea1 transcript indicates similar regulation of their expression and supports the conception of the phage activated P
2 promoter. The lysogenic repressor is known, from studies on phage λ, to increase in levels under prophage induction, while during lysogeny only basal transcription of
cro occurs [
10,
15]. The different levels of SEA observed between Sa17 and Sa48, were justified by the higher levels of RF,
sea1 gene copies and total
sea1 transcript in Sa48. Bearing in mind that the type of phages carrying the
sea gene are characterized by superinfection immunity [
10], this behavior could be attributed to enhanced phage replication and prolonged mRNA transcription or stability in Sa48.
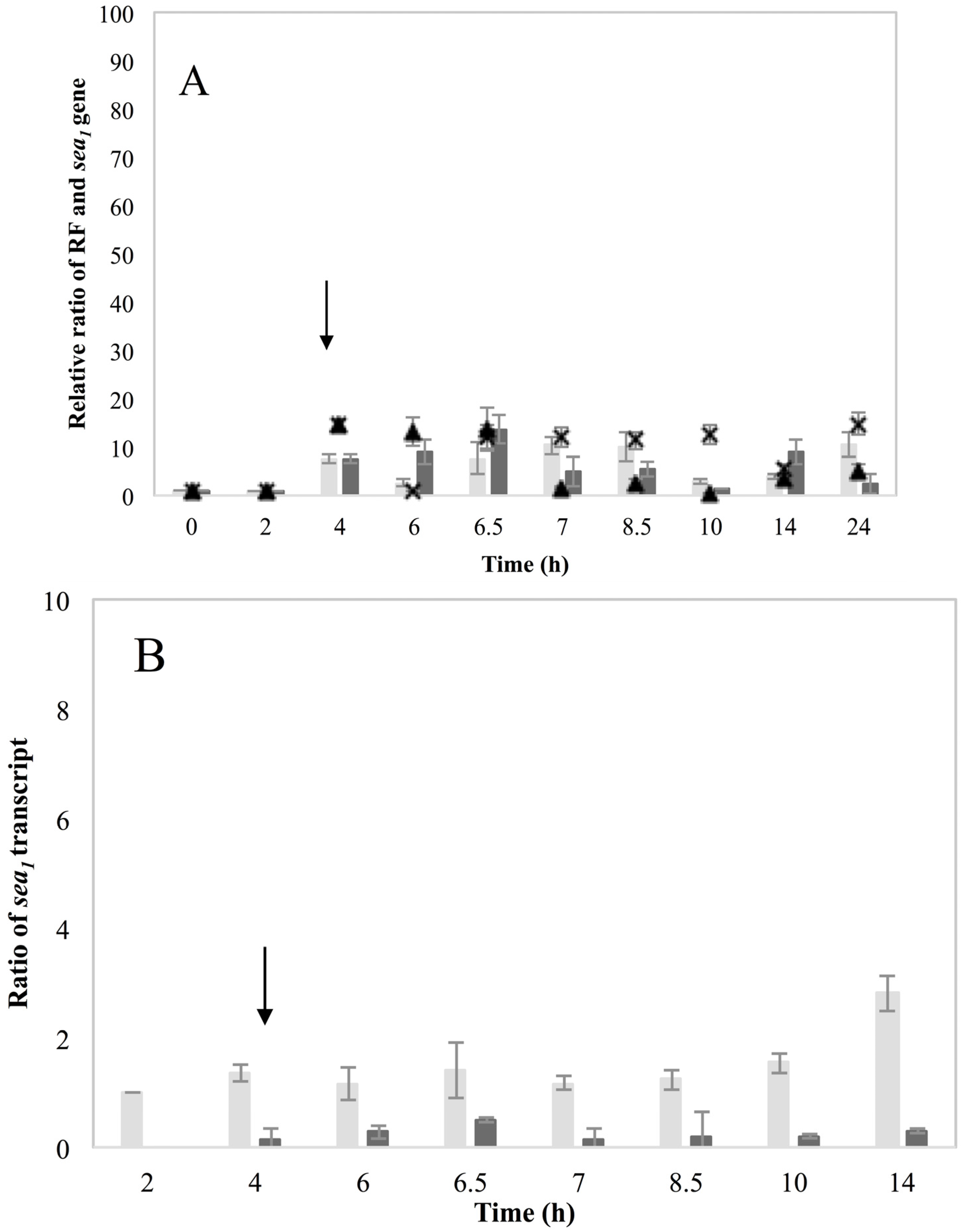
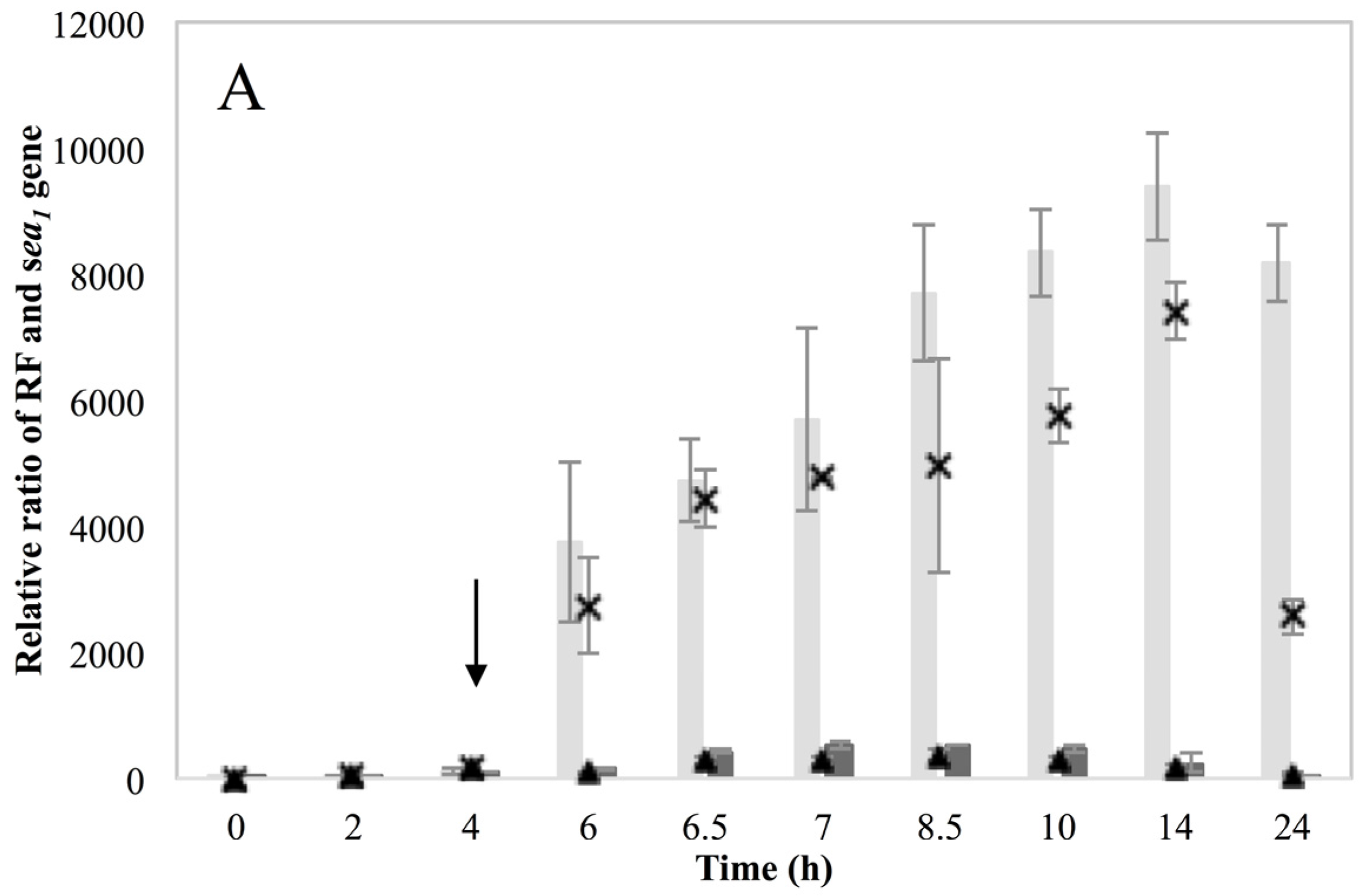
These new findings enlightening the link in which prophage induction affects SEA production, were also indirectly supported by the behavior of the non-inducible Sa21 strain. This strain behaved as if the changes in the environment by the addition of MMC were either not sensed by the cell or that the phage was unable to excise and replicate after the signal of environmental change was received. The dearth of transcripts from the latent
sea promoter, P
2, and the lysogenic repressor,
cro, favor the first hypothesis. Either way, the different behavior of Sa21 towards prophage induction could be due to key differences in the phage genome sequence, to differences in regulatory proteins of the SOS response system or to a combination of both. Equally intriguing was the equivocal behavior of the non-inducible strain Mu50. This strain has also been previously found to respond similarly towards prophage induction as Wallin-Carlquist
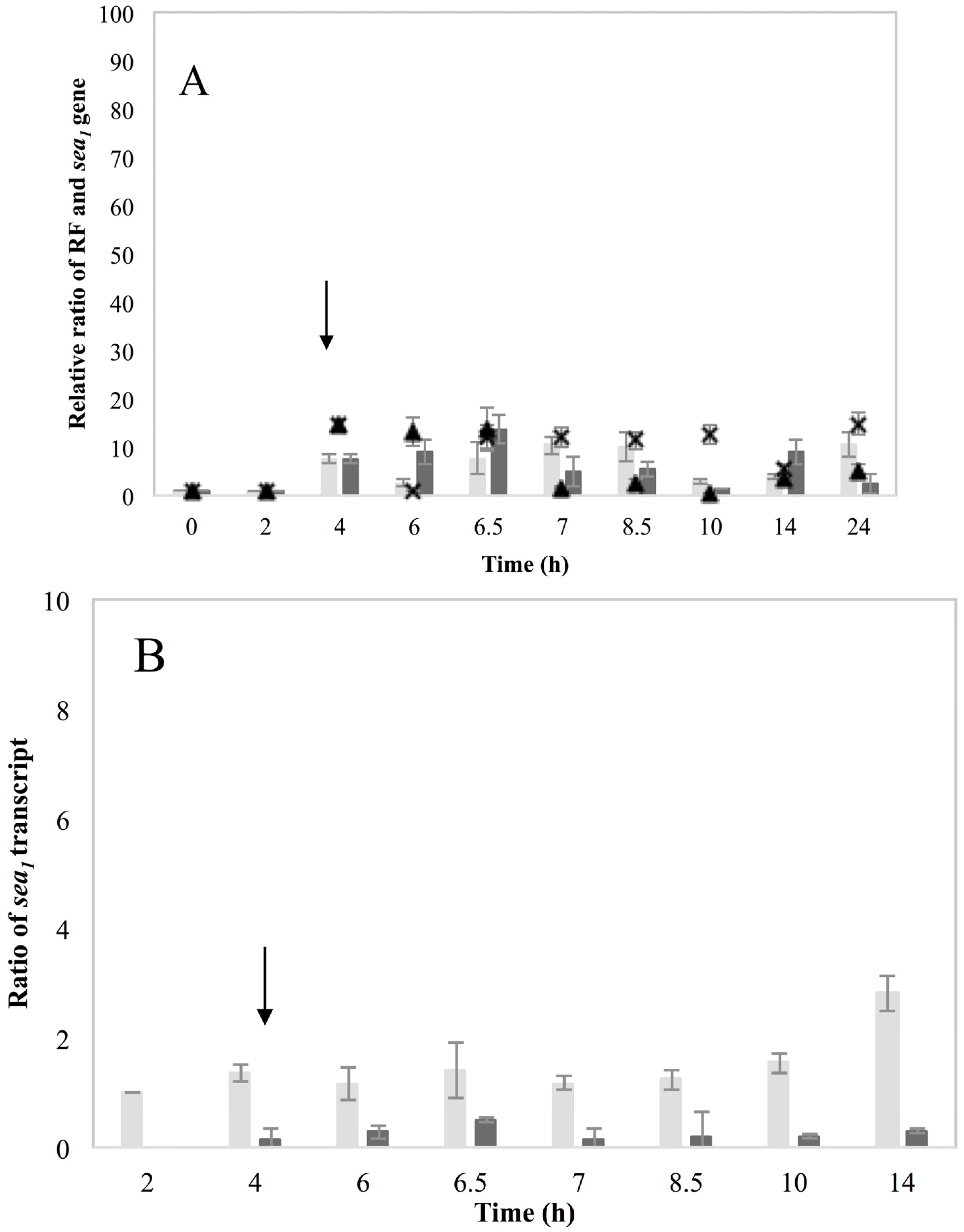
et al. [
12] showed that acetic acid increased Mu50
sea gene copy numbers at pH 5.5 but the expected boost on SEA levels was not observed. In regard to what has been observed so far for the different SEA-producing strains, it can be deduced that Mu50 could behave as an inducible high SEA-producer, when considering only the ability for prophage induction; however, possible impaired transcription efficiency could obstruct the transcription of the
sea gene. Likewise, the observation on the induction capacity of the low SEA-producer Sa51, which was only apparent on the RF and
sea2 gene copy levels, but not on
sea2 transcripts or the SEA produced, further relates to the issue of transcription efficiency in some strains and the role that sequence differences could have on the ability of the phage to excise and replicate.
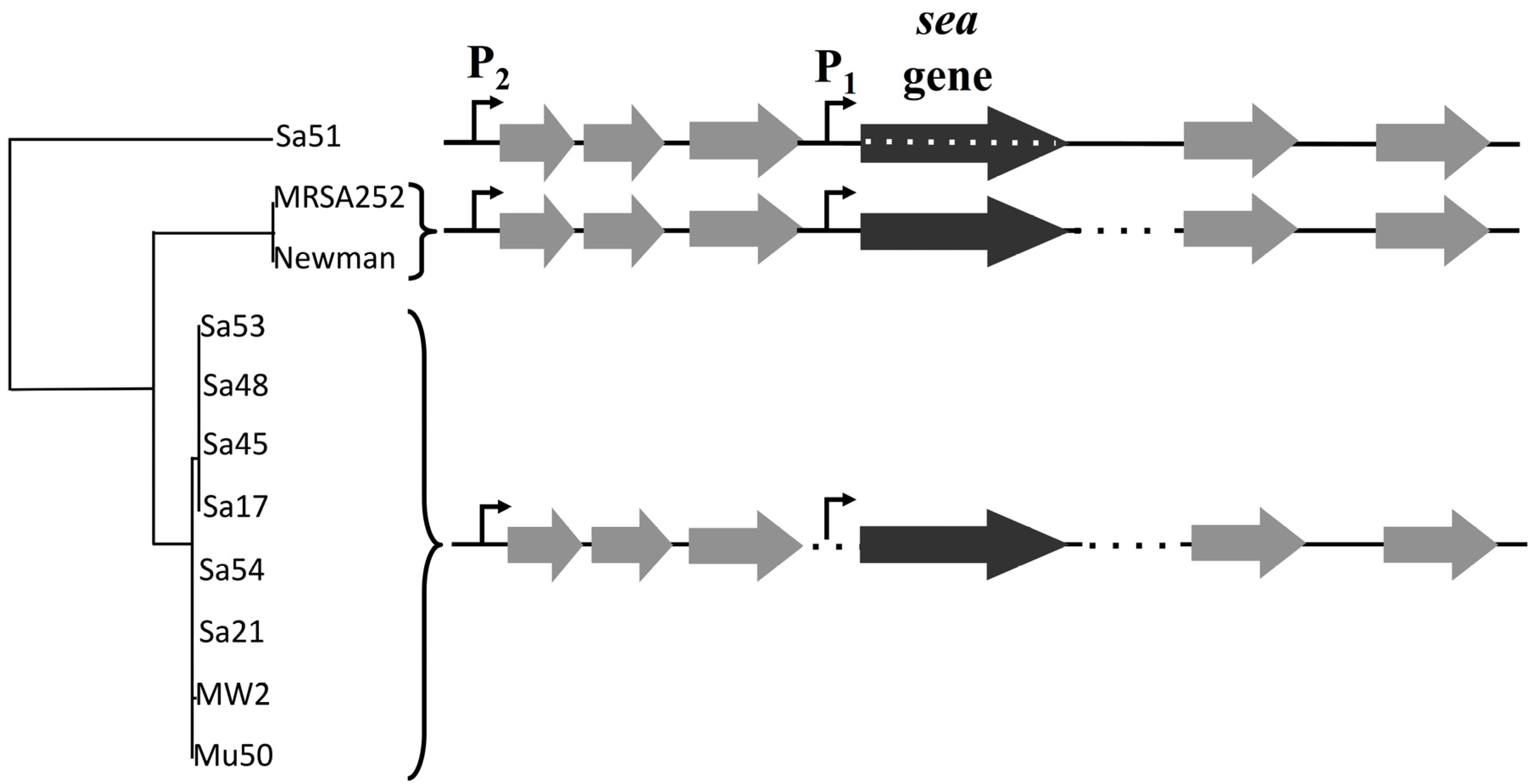
The analysis of the 3.6 kb gene region including the
sea gene complemented previous data by Wallin-Carlquist
et al. [
12], where a map for the
sea1 and
sea2 gene alleles had been created. The lack of key sequence differences in this region between the inducible and non-inducible high SEA-producers further supports the critical role of the different
sea-carrying phages on
S. aureus virulence. The possibility of sequence differences outside the region analyzed in this study, could, however, explain the capacity of the phages to excise and replicate, and thus needs further investigation. The differences observed for Sa51 could possibly explain its response to prophage induction, as the mutations in the
sea gene region alter not only the SEA amino acid sequence but could also affect the transcription and translation process in this strain. Interestingly, Soltis
et al. [
18] had also reported a gene named as
sezA that bore differences from the previously identified
sea gene. This
sea variant, though transcribed into stable mRNA could not be translated into a SEl protein, as it was deficient of an appropriate translation initiation codon. Specifically, the
sezA gene differed at the start codon by a single nucleotide deletion. Further investigation of the
sea2 gene will elucidate the present observations.
Altogether, the findings presented in this study, lead to the realization that the phage life cycle affects SEA production in two different ways. Initially, activation of the phage with consequent replication of its genome increases the pool of the sea gene copies in the cell and thus the possibility to produce more sea mRNA. This gene dose effect is one of the reasons behind the observed increase in SEA production, which is noted for the inducible high SEA-producing strains but not for the non-inducible one. Furthermore, as stated above, the sea gene is located in the late lytic region of the phage genome. Activation of the lytic promoters could accordingly have an impact on the level of sea transcription. Detection of the long sea1 transcript only for the inducible strains and during induced conditions is a strong indication that the putative latent promoter P2 is regulated by the phage and specifically activated when it is entering the lytic phase generating transcripts including sea. The demonstration that prophage induction is linked with the activation of the RecA protein highlights the critical role of environmental changes on phage activation and thus the risk for SFP.
4. Experimental Section
4.1. Bacterial Strains and Growth Conditions
Four food isolates of S. aureus, Sa17, Sa21, Sa48, and Sa51 (donated from SP Food and Bioscience, the former Swedish Institute for Food and Biotechnology, SIK, Göteborg, Sweden), and a fully sequenced strain Mu50 (LGC Promochem, London, UK), were studied. Brain Heart Infusion (BHI) broth (Difco Laboratories, BD Diagnostic Systems, Le Point de Claix, France), was used as growth medium throughout the study. Prior to inoculation, each strain was streaked on BHI agar plates (Difco Laboratories, BD Diagnostic Systems, Le Point de Claix, France), from −80 °C glycerol stocks, and incubated for 16 to 18 h (overnight) at 37 °C. A single colony of each strain was selected and sterilely transferred to a 50 mL falcon tube containing 25 mL of BHI broth. The pre-cultures were grown overnight in a rotating incubator (New Brunswick Scientific, Innova 40/40R incubator Shakers, Eppendorf AG, Hamburg, Germany) at 37 °C and 200 rpm shaking. To remove secreted metabolites and enterotoxins produced by S. aureus during overnight growth, pre-cultures were centrifuged (3220× g, 10 min, 4 °C, 5810R table centrifuge, Eppendorf AG, Hamburg, Germany) and cell pellets were washed twice in 25 mL of 0.9% sterile NaCl solution (Merck Millipore, Darmstadt, Germany) and re-suspended by vortexing in the same volume of 0.9% sterile NaCl solution. Optical density (OD) of the washed pre-cultures was measured at 620 nm (UV/Visible Spectrophotometer Ultrospec 2100 pro, GE Healthcare, Little Chalfont, UK) and the adequate inoculum volume for a starting OD of 0.1 in 400 mL BHI (1 L flask) was calculated. Thereafter, cultures were grown under optimal conditions (37 °C, 200 rpm) for about 4 h until OD ≈ 5 (mid-exponential growth phase). At this point, they were sterilely divided into half volumes and transferred to new flasks of 500 mL size. One half was then induced using mitomycin C (MMC) (Duchefa Biochemie, Haarlem, The Netherlands) in a concentration of 1 mg·mL−1 of culture, while the other was kept as a control (optimal conditions). Both cultures were then further incubated at 37 °C, 200 rpm. Samples were collected at 0 h, 2 h, 4 h, 6 h, 6.5 h, 7 h, 8.5 h, 10 h, 14 h and 24 h for OD, CFU (colony forming units), DNA and RNA isolation and ELISA analysis. For strains Sa17 and Sa21 three independent experiments were performed, while strains Sa48, Sa51 and Mu50 were investigated once. CFU determination was performed by serially diluting 1 mL of sample into 9 mL of 0.9% sterile NaCl solution. A volume of 100 μL of the appropriate dilutions was then plated, in duplicates, on BHI agar plates. The plates were incubated approximately 24 h at 37 °C before enumeration of colonies. Results were calculated and expressed as log cfu·mL−1.
4.2. DNA Extraction
Two methods, genomic and plasmid DNA extraction were used in this study to evaluate the quantity of extracted RF and the quality of the subsequent qPCR analysis. Prior to extraction, cells were washed once with sterile 0.9% NaCl solution. Purification of plasmid DNA was performed using the GeneJET Plasmid Miniprep Kit (Thermo Fisher Scientific, Waltham, MA, USA), as described in the producer’s protocol. Genomic DNA was extracted with the GeneJET Genomic DNA Purification Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the producer’s protocol for Gram-positive bacteria, including one modification of the lysis buffer. Specifically, lysis buffer was prepared using 5% lysozyme and 1% lysostaphine for more efficient extraction. Concentration of extracted DNA (ng·µL−1) and purity (A260/280) were measured using BioDrop spectrophotometer (BioDrop TOUCH UV/Visible Spectrophotometer, Integrated Scientific Solutions Inc., Walnut Creek, CA, USA). All DNA samples were stored at −20 °C until further analysis. Samples from both extraction methods were analyzed in qPCR and compared regarding the levels of the extracted RF. Both methods were equally successful in the isolation of RF and thus, plasmid extraction was chosen to continue further as it was the fastest isolation method.
4.3. RF Assay
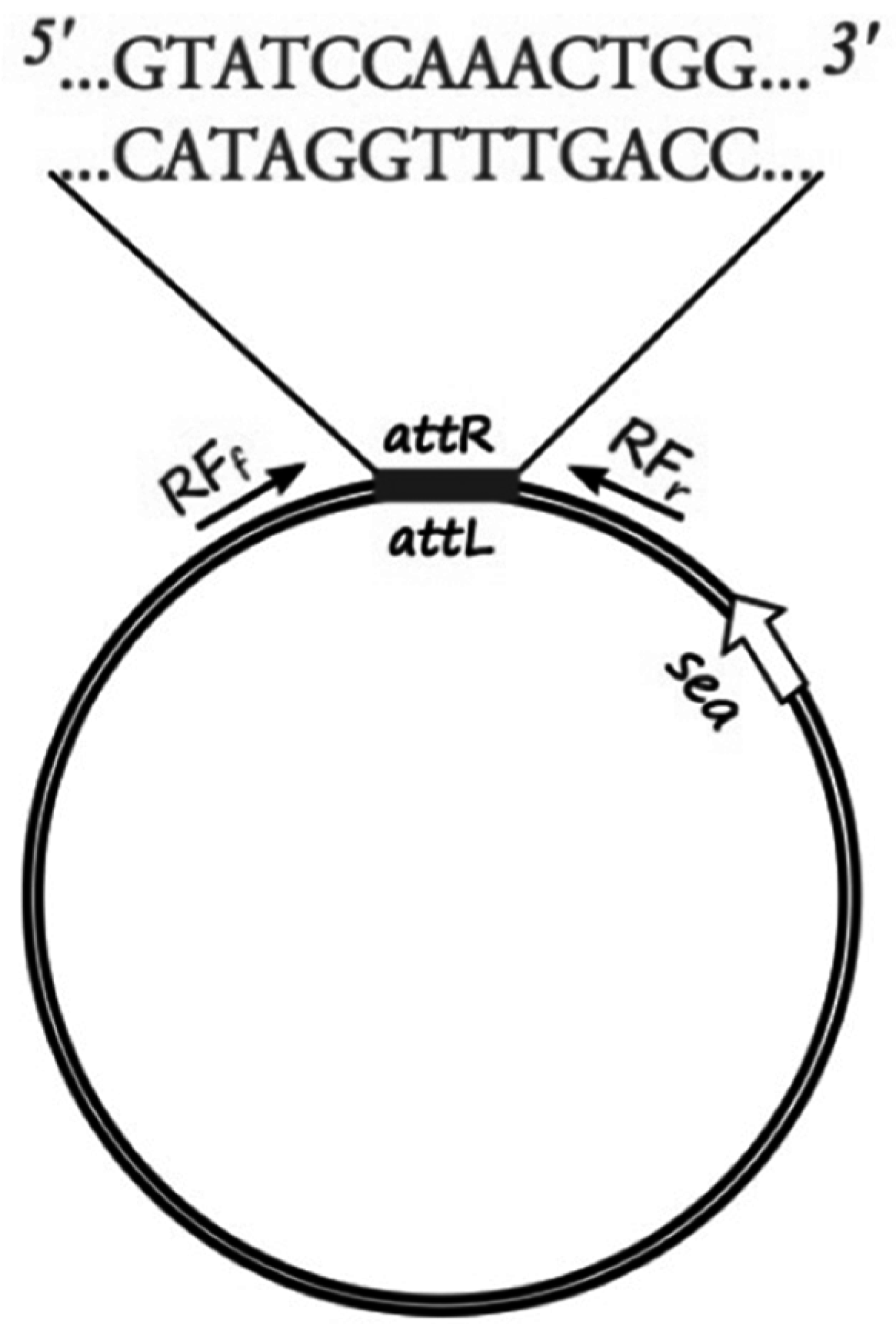
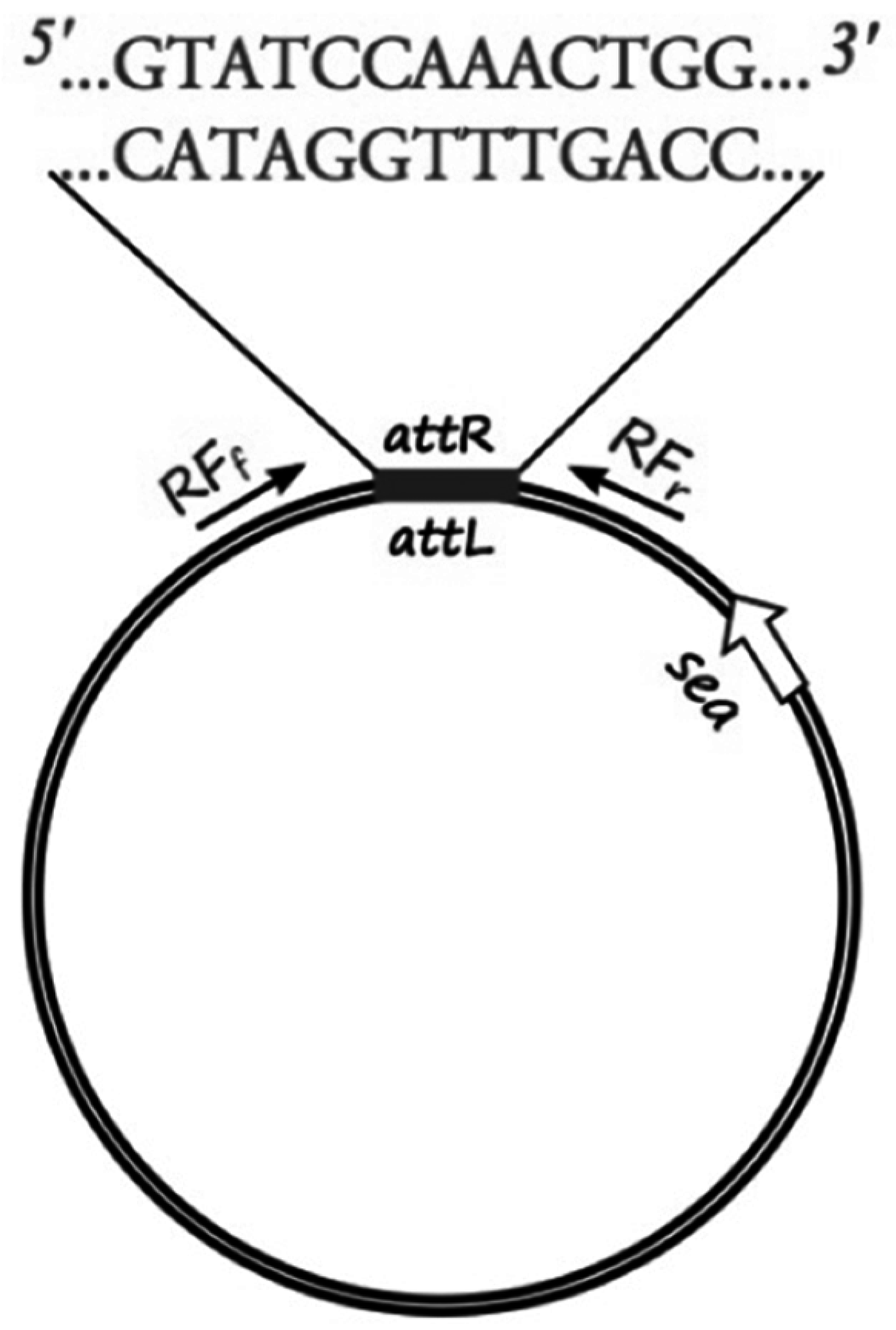
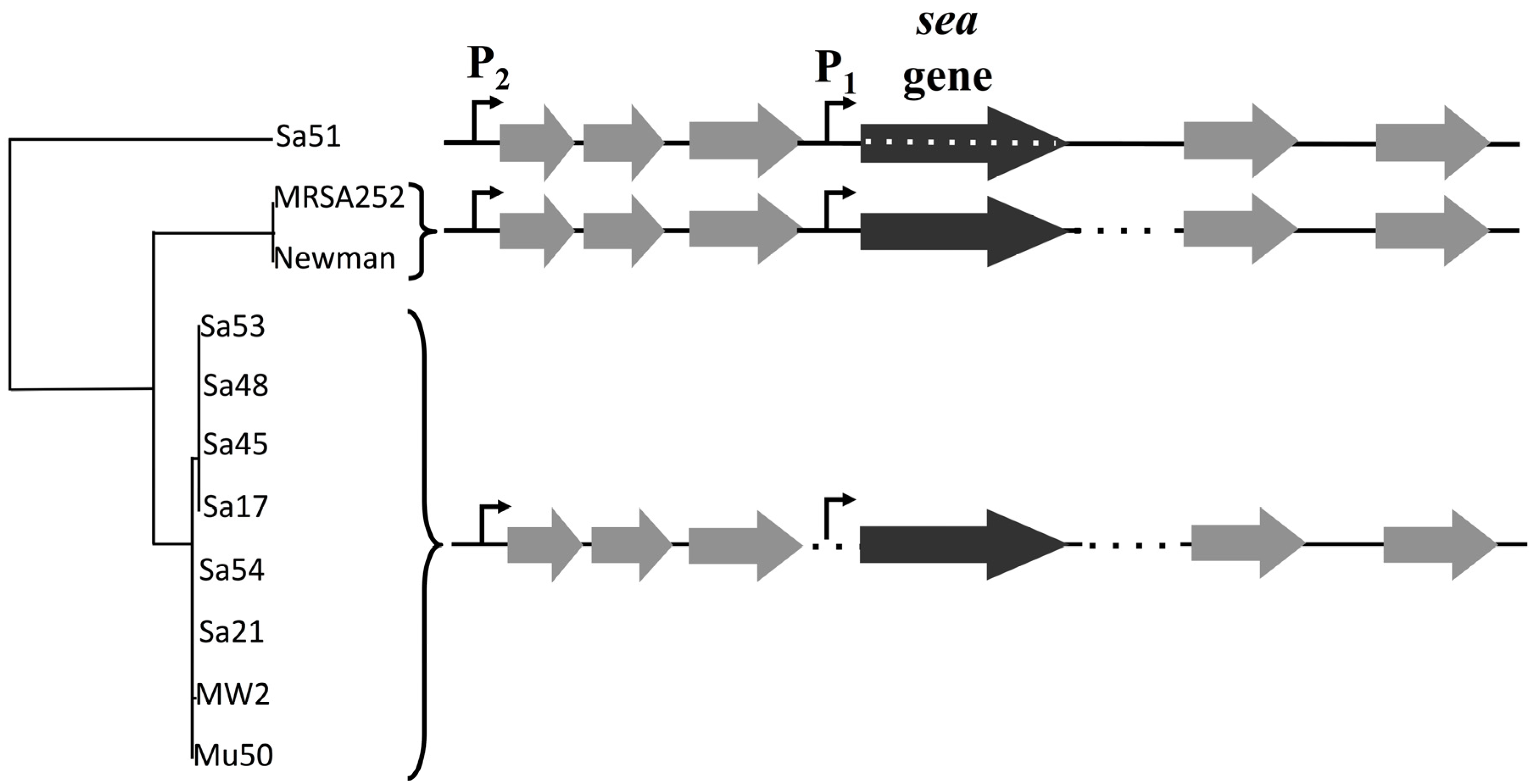
The sequenced phage ΦSa3mw of
S. aureus MW2 (Ac. No. BA000033.2) was used to pinpoint the region of the attachment sites of the prophage and bacterial chromosome ligation. Specific primers for sequencing (RF sequencing primers,
Table 2) were designed, as shown in
Figure 1, to amplify the region around the cohesive ends of the phage, only when in the circular form, in order to determine the exact sequence of the cohesive ends of six
S. aureus strains (Sa17, Sa21, Sa48, Sa51, Sa54 and Mu50). PCR was conducted with Gene Amp 9700 thermal cycler (Perkin-Elmer, Waltham, MA, USA) using the proof-reading enzyme
Pfu DNA polymerase (Thermo Fisher Scientific, Waltham, MA, USA). The following PCR protocol was used: initial denaturation at 95 °C for 1 min, followed by 30 cycles of denaturation at 95 °C for 30 s, annealing at 54 °C for 30 s and elongation at 72 °C for 3 min. A final extension step was added at 72 °C for 5 min and hold at 4 °C. Obtained PCR products were purified using GeneJET PCR Purification Kit from Thermo Fisher Scientific, Waltham, MA, USA. DNA concentration was measured using BioDrop spectrophotometer and diluted to a concentration of 10 ng·μL
−1. 15 μL of each PCR product and 10 μL of 10 μM of each primer (forward, reverse) were sent for sequencing at Eurofins MWG GmbH, Germany. The individual sequence reads were assembled and confirmed by the overlapping regions. Manual quality check and assembly of the sequencing results were performed using Serial Cloner 2.6.1 (Franck Perez, Serial Basics 2004-2013,
http://serialbasics.free.fr/Serial_Cloner.html). Analyses of the obtained sequences demonstrated a common 13-nucleotide region among four out of the five investigated strains (no sequence data obtained for Sa21) that functioned as cohesive ends for the respective phages (
Figure 1,
Supplementary Table S1). The 13-nucleotide sequence region was aligned and verified through BLAST with the existing fully sequenced strains.
The sequence region surrounding and including the 13-nucleotide cohesive ends was used to create a specific qPCR assay that would allow the quantification of explicitly circular RF copies. Specific primers were designed, as shown in
Figure 1, to amplify a 200-nucleotide long target with specific probes designed on the cohesive ends part of the target sequence.
Table 2.
Sequences of primers and probes with fluorescent dyes used for qPCR analyses of sea gene, the replicative form of the bacteriophage genome (RF) and the sequencing of RF cohesive ends.
Table 2.
Sequences of primers and probes with fluorescent dyes used for qPCR analyses of sea gene, the replicative form of the bacteriophage genome (RF) and the sequencing of RF cohesive ends.
| Target | Primer/Probe | Sequence (5′ T 3′) |
|---|
| sea1 gene | ESA-1 | 5-ATGAGTTGGGCAAGATGGTT-3 |
| Tox A reverse | 5-GGACTTGTTGTCCACGTTAGG-3 |
| Tox A-Fluo 1 | CCTTTGGAAACGGTTAAAACGAATAAGAA-FL |
| Tox A-Red 1 | LC-R640-TGTAACTGTTCAGGAGTTGGATCTTCA p |
| sea2 gene | sea2 forward | CAATATATACAAAGGGAAAAAAGTG |
| sea2 reverse | CAGAAGAAGGGTGAAACTCA |
| sea2 FL | ATCCATAAGTTAATCGGTACTTTCTTTTC-FL |
| sea2 LC | 640-TCCTCCAATTGATTATTATCATGTAACGT p |
| RF assay | forward | AAAATATAGCAATAACTACATCCG |
| reverse | AAGTCCCTAAAAAGTCCCTA |
| FL | ATGTTAAAAGTCTCCAGTTTGGATACA-FL |
| LC | 640-AGAAACCTTGTAACAACAGTATTTATTGGG p |
| Long transcript | forward | ATGAGTTGGGCAAGATGGTT |
| reverse | GGACTTGTTGTCCACGTTAGG |
| TaqMan BBQ | FAM-CATACTGCAAGTGAAGTTGGGAAGTGT-BBQ |
| RF sequencing primers | forward | CGCACGCATTAAGACACACT |
| reverse | ATGCCCCATAAACAACCCTT |
| cro gene | cro forward | TTAGCAATTTTTAAAGCACG |
| cro reverse | GCAATAAGTGCAAGAGAGTTATAT |
| cro FL | GCTTTCTCTACTTGGATGAAATACTCTCT-FL |
| cro LC | LC640-AAATCAAAACCTTTTTCTGTACCTGACA-PH |
4.4. RNA Extraction
A modified version of the protocol described by Lövenklev
et al. 2004 [
19] was used for RNA extraction. In brief, cells from 5 mL culture were harvested at 4 °C, 3220×
g. The resulting pellet was immediately frozen in liquid nitrogen and stored at −80 °C until extraction of RNA. For RNA extraction, each sample pellet was resuspended in 500 μL ice cold TES buffer, pH 7.5 (50 mM Tris (BDH Prolabo, VWR International, Stockholm, Sweden), 5 mM EDTA and 50 mM NaCl (Merck Millipore, Germany)) and the suspension was transferred to Precelly lysing kit tubes (VK 0.1) (Bertin technologies, Montigny-le-Bretonneux, France). Cells were disrupted in a Precellys 24 unit, in a three cycle run of 60 s at 6500 rpm. RNA was isolated using two steps of phenol-chloroform (600 μL:100 μL) extraction followed by one chloroform (600 μL) purification step. Precipitation of RNA was achieved in 0.1 vol of 3 M NaAc (sodium acetate) (pH 4.8) and 2.5 vol of 95% EtOH for at least for 1 h at −80 °C and further washed with 600 μL of 70% EtOH (ethanol). The remaining traces of EtOH after expiration were removed by evaporation in room temperature, before the RNA was dissolved in 100 μL RNA storage solution (Ambion, Thermo Fisher Scientific, Waltham, MA, USA). Concentration of extracted RNA (ng·μL
−1) and purity (A
260/280) were measured using BioDrop spectrophotometer. Prior to transformation into cDNA by reverse transcription, all RNA samples were treated with DNase to degrade contaminating traces of DNA. For this reaction, 15 μL of each RNA sample was mixed with 15 μL autoclaved diethyl pyrocarbonate (Fluka Analytical, Sigma-Aldrich, Buchs SG, Switzerland)-treated water, 15U RQ1 RNase-free DNase (Promega Co., Madison, WI, USA) and 1× reaction buffer. The suspension was incubated at 37 °C for 45 min. Stop solution, in combination with incubation at 65 °C for 10 min, was further added to inactivate the DNase. All samples were stored at −80 °C until further analysis.
4.5. cDNA Synthesis
First-strand cDNA was synthesized in two separate reverse-transcription reactions using reverse primers specific to the total
sea, long
sea and
cro transcripts (
Table 2). The reactions were performed with a Gene Amp 9700 thermal cycler (Perkin-Elmer Cetus, Norwalk, CT, USA). The total volume of the mixture was 20 μL and contained 0.5 μg of total RNA, the reverse primer for each target at a concentration of 0.5 mM, each deoxynucleoside triphosphate (dATP, dTTP, dCTP, and dGTP; Roche Diagnostics GmbH, Penzberg, Germany) at a concentration of 5 mM, 20 U of RNasin RNase inhibitor (Promega Co., Madison, WI, USA), 5 mM dithiothreitol, 1× first-strand buffer, and 200 U of Superscript II RNase reverse transcriptase (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA). Autoclaved ddH
2O was used both in the reaction mixture and to dilute mRNA samples. Before RT enzyme was added, the reaction mixture was heated to 65 °C for 5 min and then chilled on ice. After a brief centrifugation and addition of the RT enzymes, the reaction mixture was incubated at 42 °C for 50 min, and the reaction was terminated by incubation at 70 °C for 15 min. The cDNA was diluted 10-fold in autoclaved ddH
2O before qPCR analysis. Transcript analysis was performed on samples until 14 h of growth, as it was shown before that transcription decreases notably at late growth phases [
12,
13].
4.6. Quantification of RF, sea Gene Copies, sea and cro Transcripts
To follow the levels of the bacteriophage RF and the
sea gene, qPCR analysis of the purified plasmid samples was performed. To investigate the presence of the
sea and
cro transcripts, the cDNA synthesized from the extracted RNA was used for qPCR analysis. Specifically, a 20 μL qPCR reaction, including 10 ng of plasmid DNA template, was performed in a LightCycler 2.0 Carousel-Based System
® (Roche Diagnostics GmbH, Penzberg, Germany), using hybridization probes. The PCR mixture consisted of 1 PCR
Tth buffer, 3.25 mM MgCl
2, 0.2 mM of dNTPs mixture, 0.5 μM of both forward and reverse primers, 0.03 μM of each hybridization probe and 0.05 U of
Tth polymerase. All reagents mentioned above were purchased from Roche Diagnostics GmbH, Germany, apart from probes and primers, which were supplied by TIB Molbiol GmbH, Berlin, Germany (listed in
Table 2). Autoclaved ddH
2O was used for all preparations and as negative control. The PCR protocol consisted of initial denaturation at 95 °C for 1 min, followed by 45 cycles of denaturation at 95 °C for 0 s (
i.e., no hold at 95 °C), primer annealing at 46 °C for
sea1, 49 °C for
sea2, 50 °C for
cro and 49 °C for RF, for 5 s, and extension at 72 °C for 25 s. A single fluorescence measurement was made at the end of the extension step. Cq values obtained were accepted when efficiency of amplification was between 1.8 and 2.0. Results are presented as calculated ratios between the reference sample (0 h sample; denoted “control” in the equation) and the target sample according to the equation of Pfaffl (ratio = (E
target)
ΔCq target(control-sample) [
20]).
4.7. ELISA
ELISA for SEA and SEE was performed according to the revised laboratory protocol for staphylococcal enterotoxin A, described before by Wallin-Carlquist
et al. [
12]. Absorbance was measured at 405 nm with Multiskan Ascent
® spectrophotometer (Electron Corporation, Thermo Fisher Scientific, Waltham, MA, USA). Obtained absorbance values were plotted against toxin concentrations. Absorbance values for the standard samples were plotted against the known concentrations of SEA and SEE and standard curves were created. The concentrations of the unknown samples were calculated using the linear regression and expressed in ng·mL
−1 of toxin.
4.8. Sequencing and Analysis of sea Gene Region
A 3.6 kb region including the
sea gene and its 2 putative promoters was sequenced in 11 different isolates of
S. aureus strains (
Table 1). Sequencing primers for the food isolates (Sa17, Sa21, Sa45, Sa48, Sa51, Sa53, Sa54) were designed by multiple alignment of known sequences of
S. aureus strains Mu50, MW2, Newman and MRSA252 from NCBI database with MUSCLE [
21] and the primers were designed to have a 0.5 kb distance between them to support assembly and confirmation processes by overlapping regions. PCR was conducted with Gene Amp 9700 thermal cycler (Perkin-Elmer Cetus, Norwalk, CT, USA) using the proofreading enzyme Phusion DNA polymerase (Thermo Fisher Scientific, Waltham, MA, USA) and 1X Phusion GC Buffer. For strains MRSA252 and Newman the primer set YBSEA_9 and YBSEA_19 was used for amplification while the rest of the strains were amplified with the primer set of YBSEA_9 and YBSEA_18 (
Table 3). The following PCR protocol was used: initial denaturation at 98 °C for 1 min, followed by 30 cycles of denaturation at 98 °C for 10 s, annealing at 47 °C and 55 °C, respectively, for 30 s and elongation at 72 °C for 3 min. A final extension step was added at 72 °C for 7 min and held at 4 °C. Obtained PCR products were purified using GeneJET PCR Purification Kit. DNA concentration was measured and diluted to a concentration of 10 ng·μL
−1. Fifteen microliters of each PCR product was premixed with 2 μL of 10 μM of appropriate primer set and sequenced by Eurofins Genomics, Germany. Each individual sequence reads were assembled and confirmed by the overlapping regions. Manual quality check and assembly of the sequencing results were performed using Serial Cloner 2.6.1 and FinchTV 1.5.0 (Geospiza Inc., Seattle, WA, USA) followed by multiple sequence alignment of all obtained sequences using MUSCLE [
21] and phylogenetic tree construction using TreeDyn [
22].
Table 3.
Primer sequences used for sequencing the 3.6 kb gene region including the sea gene and for the construction of the recA-disruption mutant.
Table 3.
Primer sequences used for sequencing the 3.6 kb gene region including the sea gene and for the construction of the recA-disruption mutant.
| Primer | Direction | Sequence | Target DNA | Target Strains |
|---|
| YBSEA_1 | Forward | AAGTGGTAAAAAGTTATCTG | 1.5–1 kb upstream sea | MW2, Sa17, Sa21, Sa45, Sa54 |
| YBSEA_2 | Forward | GTAGGCAGTTTTACAACGTC | 1–0.5 kb upstream sea | MW2, Sa17, Sa21, Sa45, Sa54, Sa48, Sa53 |
| YBSEA_3 | Forward | TAAATGGCTCGTAGGTGCCA | 0.5 kb upstream—start of sea | MW2, Sa17, Sa21, Sa45, Sa54, Sa48, Sa53 |
| YBSEA_4 | Forward | ATGAAAAAAACAGCATTTA | Start of SEA—0.5 kb inside sea | MW2, Sa17, Sa21, Sa45, Sa54, MRSA252, Sa48, Sa53 |
| YBSEA_5 | Forward | GTTAAAACGAATAAGAAAAA | 0.5 kb inside SEA—0.2 kb downstream sea | All strains |
| YBSEA_6 | Forward | ATTCCTTATTGCATTGATAG | 0.2–0.7 kb downstream sea | All strains |
| YBSEA_7 | Forward | TATGAAGAACCAAGAAGAAA | 0.7–1.2 kb downstream sea | All strains |
| YBSEA_8 | Forward | CAAGTAAAATAACAGTTGGA | 1.2–1.5 kb downstream sea | All strains |
| YBSEA_9 | Reverse | AATCAATCTCTCATGCCATA | 1.5–1 kb downstream sea | All strains |
| YBSEA_18 | Forward | TCAAACGCTGATAGTGCATA | 2–1.5 kb upstream sea | MW2, Sa17, Sa21, Sa45, Sa54 |
| YBSEA_19 | Forward | CTGATGAGAACTATGATTAC | 2–1.5 kb upstream sea | Newman, Sa51, MRSA252 |
| YBSEA_20 | Forward | GGTTTGATTGCTGAAGAGGT | 1.5–1 kb upstream sea | Newman, Sa51, MRSA252 |
| YBSEA_21 | Forward | AAATCTGAAGAAAACGCTAA | 1–0.5 kb upstream sea | Newman, Sa51, MRSA252 |
| YBSEA_22 | Forward | TGCAGTCATTACTGCATCAA | 0.5 kb upstream—start of sea | Newman, Sa51, MRSA252 |
| YBSEA_101 | Forward | GGAGAAATNGAAGGTATNG | 2–1.5 kb upstream sea | Sa48 |
| YBSEA_105 | Reverse | TATCTAGTGGCTGAAGTAGC | 1.5–1 kb downstream sea | Sa53 |
| YB RecA-108 | Forward | AGTCAGTCGAGCTCTATGGAGAAATCTTTCGGTA |
| YB RecA-109 | Reverse | GTCAGTCGCGGCCGCAGTCTCTGGATTACCGAACA |
| YB RecA-110 | Forward | CCTTGTATAGTATTGGTAAGATA |
| YB RecA-111 | Reverse | GTATCGATAAGCTTGATATC |
4.9. Construction of recA Disruption Mutant
To create the
S. aureus Sa17
recA disruption mutant, recombination technique with the plasmid pIMAY containing a chloramphenicol resistance gene was used [
23]. A single
recA fragment was amplified from
S. aureus Sa17 strain with primers YBRecA-108 and YBRecA-109 (
Table 3) and inserted in pIMAY between
SacI and
NotI restriction sites. The amplified fragment was designed to recombine with the Sa17’s
recA gene in the chromosome, resulting in integration of the plasmid. For the amplification of the Sa17
recA fragment Phusion polymerase was used and a PCR reaction with initial denaturation at 9 °C for 30 s, a first denaturation, annealing and elongation step for 10 cycles at 98 °C for 10 s, 57 °C for 10 s, 72 °C for 30 s, respectively, a second denaturation, annealing and elongation step for 25 cycles at 98 °C for 10 s, 72 °C for 30 s, respectively, followed by a final elongation at 72 °C for 5 min. The product size was confirmed by gel electrophoresis and purified using GeneJet PCR purification kit (Thermo Fisher Scientific, Waltham, MA, USA). Both the PCR product and pIMAY were digested with
SacI and
NotI and ligated with a molar ratio of 1:3 creating a pIMAY+
recA-disruption mutant plasmid. Preparation of electro-competent
S. aureus Sa17 cells and transformation were performed according to the original protocol of Löfblom
et al. [
24] and as adjusted by Monk
et al. [
23].
Mutant selection was performed as described in Monk
et al. [
23]. Disruption of the
recA gene was confirmed by colony PCR using primer YBRecA-110 and YBRecA-111 (
Table 3). The PCR reaction was performed using Phire Hot Start II DNA Polymerase (Thermo Fisher Scientific, Waltham, MA, USA) and the following protocol: a part of colony was transferred in a PCR tube using a 1 μL loop, heated in the microwave at full power (800 W) for 5 min and immediately cooled down on ice. An amount of 50 μL of master mix containing 1 U·μL
−1 of DNA polymerase, as recommended by the manufacturer was then added to the PCR tube. Colony PCR reaction was performed at initial denaturation of 98 °C for 30 s, denaturation annealing and elongation (35 cycles) at 98 °C for 10 s, 51 °C for 10 s and 72 °C for 20 s, respectively, followed by final elongation at 72 °C for 5 min.
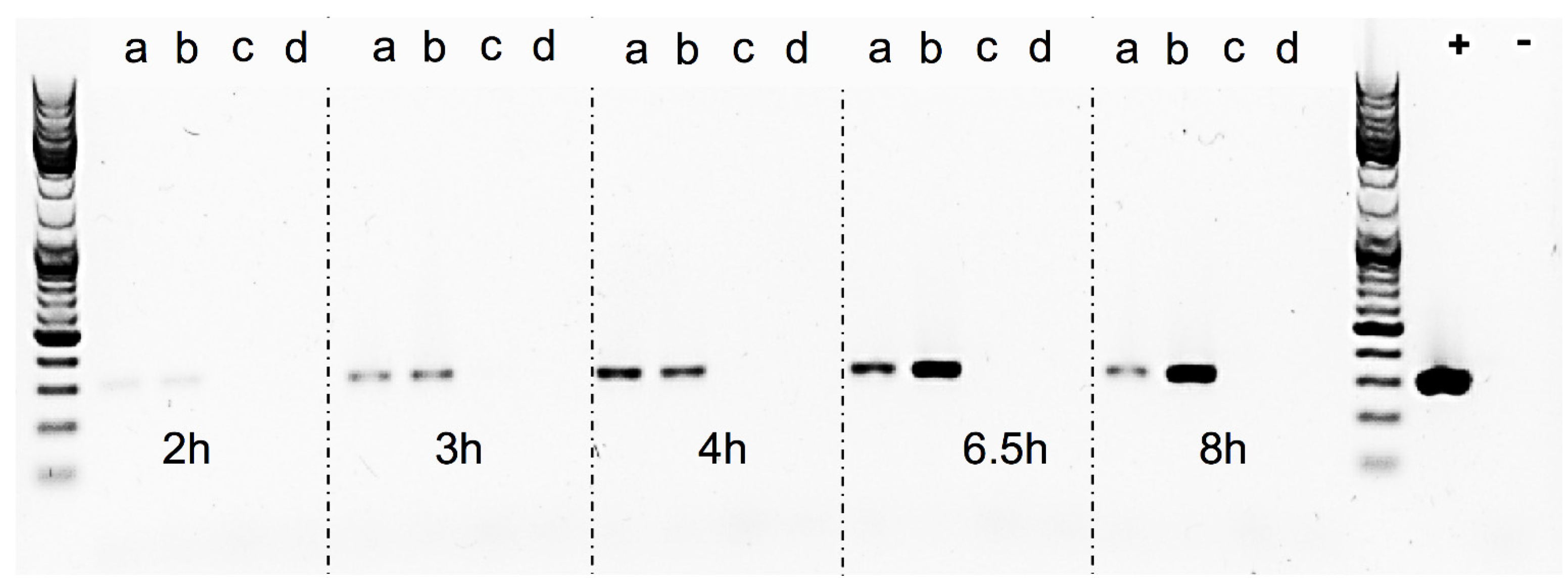
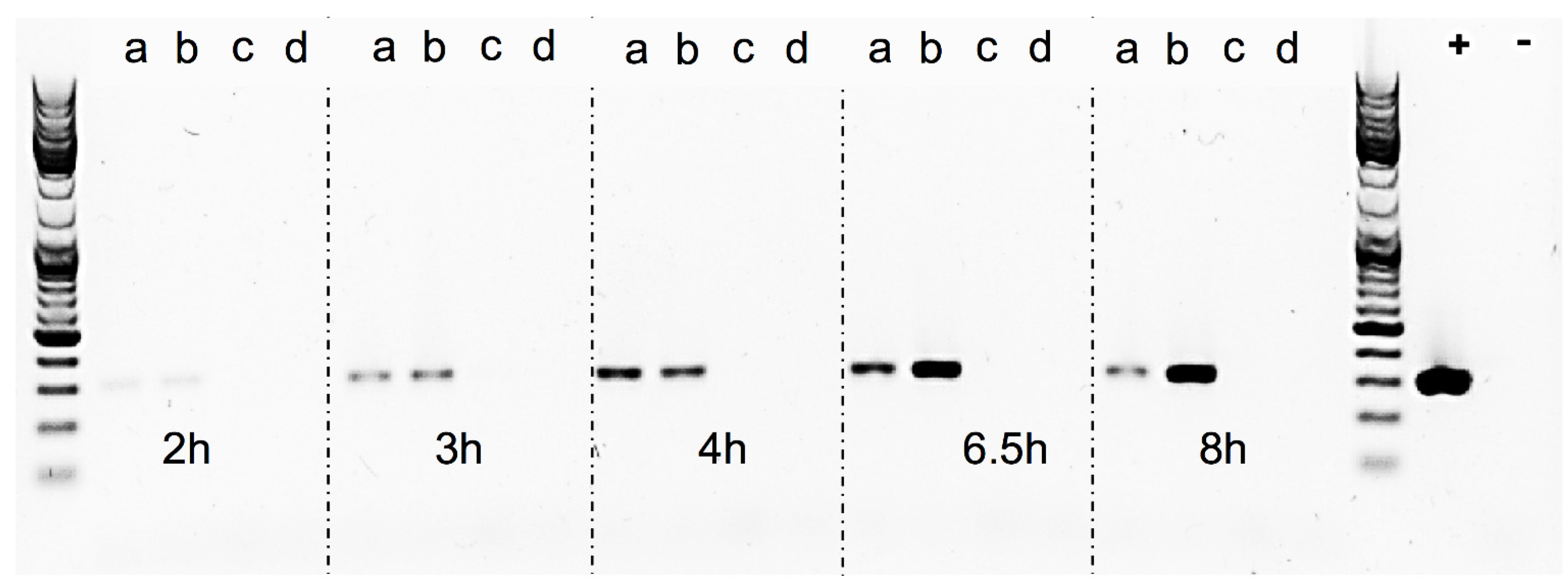
The Sa17
recA-disruption mutant and Sa17 wild type strains were investigated for their growth pattern and SEA production under controlled and induced growth conditions, as described in “Bacterial strains and growth conditions”. Both strains were then studied for the amount of RF copies produced, by amplification of the RF target in a conventional PCR reaction. The primers used were the same as described in “RF assay” (
Table 2), the DNA Polymerase was Phire Hot Start II (Thermo Fisher Scientific, Waltham, MA, USA). To normalize the reaction, the same amount of plasmid DNA (10 ng) isolated from each time point was used as template. The reaction set up was as follows: initial denaturation at 98 °C for 30 s, denaturation, annealing and elongation steps for 25 cycles at 98 °C for 10 s, 56.5 °C for 10 s and 72 °C for 10 s, respectively, followed by final elongation at 72 °C for 5 min. Assuming the efficiency of PCR in each reaction is the same, an equal volume of 5 μL from each sample along with GeneRuler DNA Ladder Mix (Thermo Fisher Scientific, Waltham, MA, USA) to verify the product size was visualized in gel electrophoresis using Biorad Universal Hood II (Biorad Laboratories Inc., Hercules, CA, USA). Density analysis of the bands was performed using Quantity one 1-D analysis software (Biorad Laboratories Inc., Hercules, CA, USA).
4.10. Statistical Analyses
The mean values for RF and sea gene levels between the MMC induced and control conditions were compared for statistical significance using unpaired t-test function of Microsoft Office Excel (2013, Microsoft, Stockholm, Sweden) at a significance levels of 0.05 (p value).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}