
Recommended Immunological Strategies to Screen for Botulinum Neurotoxin-Containing Samples

Abstract
:1. Introduction
2. Results and Discussion
2.1. Assay Development and Validation
2.1.1. ELISA 4 and LFA 2
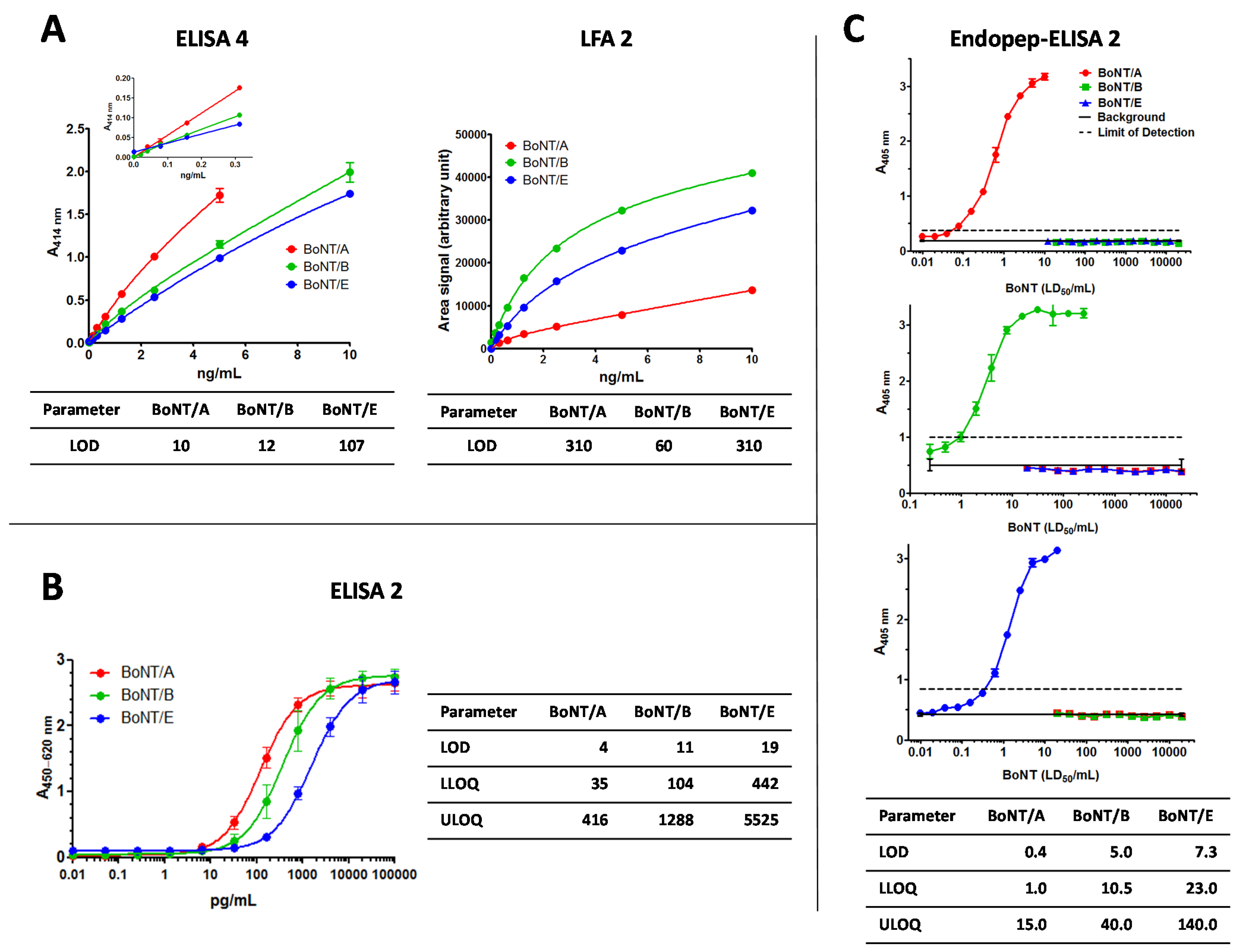
2.1.2. ELISA 2
2.1.3. Endopeptidase-ELISA 2
2.2. The Proficiency Test Scheme

{kind=link}
{kind=link}
{kind=link}
| Sample | Matrix | Serotype | xa Nominal Concentration (ng/mL) |
|---|---|---|---|
| S1 | meat extract | BoNT/A | 10.5 |
| S2 | 0.1% BSA/PBS | BoNT/A | 9.9 |
| S3 | 0.1% BSA/PBS | none | 0.0 |
| S4 | 0.1% BSA/PBS | BoNT/E | 10.9 |
| S5 | meat extract | BoNT/A | 108.0 |
| S6 | 0.1% BSA/PBS | BoNT/B | 9.0 |
| S7 | 0.1% BSA/PBS | BoNT/A | 100.0 |
| S8 | 0.1% BSA/PBS | BoNT/A | 4.7 |
| BoNT/B | 4.5 | ||
| S9 | 0.1% BSA/PBS | BoNT/A | 0.5 |
| S10 | cow’s milk | BoNT/A | 10.3 |
| S11 | human serum | BoNT/A | 9.8 |
| S12 | 0.1% BSA/PBS | BoNT/A | 1001.0 |
| S13 | cow’s milk | BoNT/A | 112.0 |
2.3. Overview of the Diagnostic Approaches Utilized by the Participating Laboratories
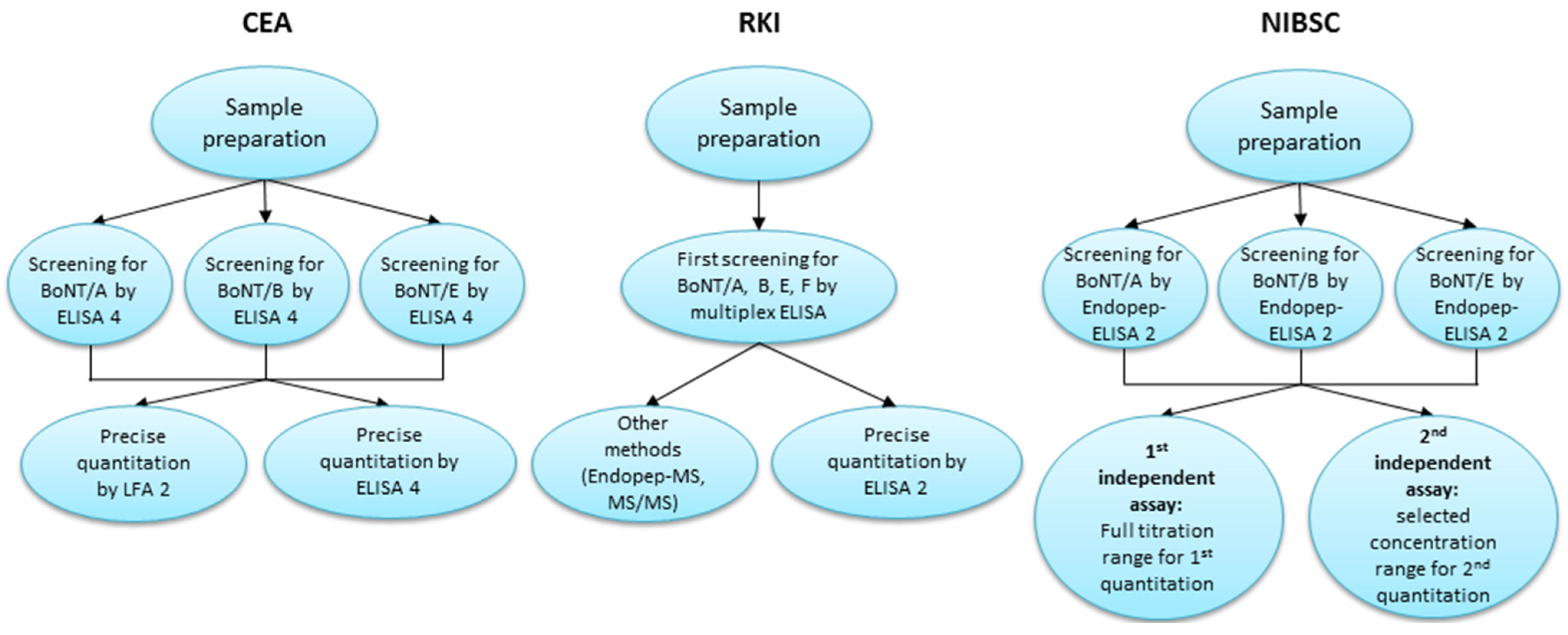
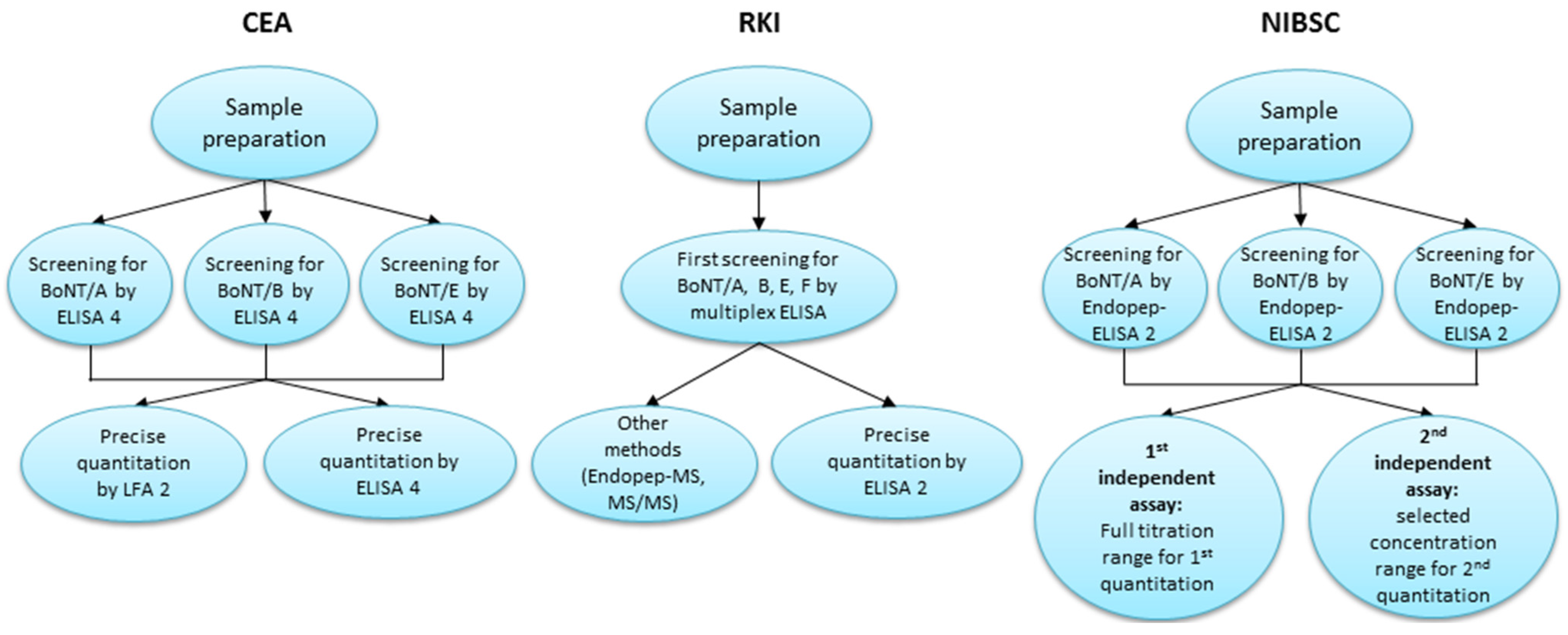
2.3.1. Diagnostic Approach of CEA
2.3.2. Diagnostic Approach of RKI
2.3.3. Diagnostic Approach of NIBSC
2.4. Individual Laboratory Assay Results
2.4.1. Laboratory Results from CEA
2.4.2. Laboratory Results from RKI
2.4.3. Laboratory Results from NIBSC
2.5. Overall Assessment of Immunological Results
2.5.1. Qualitative Results
| Method | PT Sample | BoNT | BoNT/A | BoNT/B | BoNT/E | BoNT/F |
|---|---|---|---|---|---|---|
| LFA 2 | S1 | 1 | 1 | −1 | −1 | - |
| S2 | 1 | 1 | −1 | −1 | - | |
| S3 | b | −1 | −1 | −1 | - | |
| S4 | 1 | −1 | −1 | 1 | - | |
| S5 | 1 | 1 | −1 | −1 | - | |
| S6 | 1 | −1 | 1 | −1 | - | |
| S7 | 1 | 1 | −1 | −1 | - | |
| S8 | 1 | 1 | 1 | −1 | - | |
| S9 | 1 | 1 | −1 | −1 | - | |
| S10 | 1 | 1 | −1 | −1 | - | |
| S11 | 1 | 1 | −1 | −1 | - | |
| S12 | 1 | 1 | −1 | −1 | - | |
| S13 | 1 | 1 | −1 | −1 | - | |
| ELISA 4 c | S1 | 1 | 1 | −1 | −1 | - |
| S2 | 1 | 1 | −1 | −1 | - | |
| S3 | b | −1 | −1 | −1 | - | |
| S4 | 1 | −1 | −1 | 1 | - | |
| S5 | 1 | 1 | −1 | −1 | - | |
| S6 | 1 | −1 | 1 | −1 | - | |
| S7 | 1 | 1 | −1 | −1 | - | |
| S8 | 1 | 1 | 1 | −1 | - | |
| S9 | 1 | 1 | −1 | −1 | - | |
| S10 | 1 | 1 | −1 | −1 | - | |
| S11 | 1 | 1 | −1 | −1 | - | |
| S12 | 1 | 1 | 10 | −1 | - | |
| S13 | 1 | 1 | −1 | −1 | - | |
| ELISA 2 | S1 | 1 | 1 | −1 | −1 | −1 |
| S2 | 1 | 1 | −1 | −1 | −1 | |
| S3 | −1 | −1 | −1 | −1 | −1 | |
| S4 | 1 | −1 | −1 | 1 | −1 | |
| S5 | 1 | 1 | −1 | −1 | −1 | |
| S6 | 1 | −1 | 1 | −1 | −1 | |
| S7 | 1 | 1 | −1 | −1 | −1 | |
| S8 | 1 | 1 | 1 | −1 | −1 | |
| S9 | 1 | 1 | −1 | −1 | −1 | |
| S10 | 1 | 1 | −1 | −1 | −1 | |
| S11 | 1 | 1 | −1 | −1 | −1 | |
| S12 | 1 | 1 | −1 | −1 | −1 | |
| S13 | 1 | 1 | −1 | −1 | −1 | |
| Endopep-ELISA 2 | S1 | 1 | 1 | −1 | −1 | - |
| S2 | 1 | 1 | −1 | −1 | - | |
| S3 | −1 | −1 | −1 | −1 | - | |
| S4 | 1 | −1 | −1 | 1 | - | |
| S5 | 1 | 1 | −1 | −1 | - | |
| S6 | 1 | −1 | 1 | −1 | - | |
| S7 | 1 | 1 | −1 | −1 | - | |
| S8 | 1 | 1 | 1 | −1 | - | |
| S9 | 1 | 1 | −1 | −1 | - | |
| S10 | 1 | 1 | −1 | −1 | - | |
| S11 | 1 | 1 | b | 10 | - | |
| S12 | 1 | 1 | −1 | −1 | - | |
| S13 | 1 | 1 | −1 | −1 | - |
2.5.2. Quantitative Results
| Sample | Matrix | Analyte | xa (ng/mL) | xELISA 4 (ng/mL) | zELISA 4 | xLFA 2 (ng/mL) | zLFA 2 | xELISA 2 (ng/mL) | zELISA 2 | xEndopep ELISA 2 (ng/mL) | zEndopep ELISA 2 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | meat extract | BoNT/A | 10.5 | 20.0 | 3.6 | 26.0 | 5.8 | 15.3 | 1.8 | 13.9 | 1.3 |
| S2 | 0.1% BSA/PBS | BoNT/A | 9.9 | 18.8 | 3.5 | 29.6 | 7.8 | 12.9 | 1.2 | 15.1 | 2.1 |
| S3 | 0.1% BSA/PBS | none | 0.0 | 0.0 | n/a | 0.0 | n/a | 0.0 | n/a | 0.0 | n/a |
| S4 | 0.1% BSA/PBS | BoNT/E | 10.9 | 12.5 | 0.6 | 20.7 | 3.6 | 6.56 | −1.5 | 21.3 | 3.8 |
| S5 | meat extract | BoNT/A | 108.0 | 205 | 3.5 | 173 | 2.4 | 148 | 1.4 | 161 | 1.9 |
| S6 | 0.1% BSA/PBS | BoNT/B | 9.0 | 8.00 | −0.4 | 9.30 | 0.1 | 7.91 | −0.5 | 27.1 | 7.9 |
| S7 | 0.1% BSA/PBS | BoNT/A | 100.0 | 216 | 4.5 | 195 | 3.7 | 163 | 2.5 | 191 | 3.6 |
| S8 | 0.1% BSA/PBS | BoNT/A | 4.7 | 9.65 | 4.1 | 10.5 | 4.8 | 7.47 | 2.3 | 7.69 | 2.4 |
| BoNT/B | 4.5 | 4.35 | −0.1 | 4.50 | 0.0 | 4.10 | −0.4 | 13.1 | 7.5 | ||
| S9 | 0.1% BSA/PBS | BoNT/A | 0.5 | 0.770 | 2.2 | 1.25 | 6.0 | 0.718 | 1.8 | 0.875 | 3.0 |
| S10 | milk | BoNT/A | 10.3 | 17.1 | 2.6 | 16.0 | 2.1 | 16.3 | 2.3 | 13.8 | 1.3 |
| S11 | serum | BoNT/A | 9.8 | 17.7 | 3.1 | 14.2 | 1.7 | 15.4 | 2.2 | 22.0 | 4.9 |
| S12 | 0.1% BSA/PBS | BoNT/A | 1001.0 | 1664 | 2.6 | 2173 | 4.6 | 1487 | 1.9 | 1542 | 2.1 |
| S13 | milk | BoNT/A | 112.0 | 211 | 3.5 | 218 | 3.7 | 162 | 1.7 | 130 | 0.6 |
3. Experimental Section
3.1. CEA
3.1.1. Ethics Statement
3.1.2. Reagents
3.1.3. Description of the Performed Tests
3.1.4. Sandwich ELISA
3.1.5. Lateral Flow Immunoassays
3.2. RKI
3.2.1. Ethics Statement
3.2.2. Antibodies
3.2.3. Multiplex Bead-Based ELISA
3.2.4. Singleplex Sandwich ELISA
3.3. NIBSC
3.3.1. Ethics Statement
3.3.2. Reference Toxins
3.3.3. Endopeptidase ELISA
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Popoff, M.R. Ecology of neurotoxigenic strains of clostridia. Curr. Top. Microbiol. Immunol. 1995, 195, 1–29. [Google Scholar] [PubMed]
- Sobel, J. Botulism. Clin. Infect. Dis. 2005, 41, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Gu, S.; Jin, L.; Le, T.T.; Cheng, L.W.; Strotmeier, J.; Kruel, A.M.; Yao, G.; Perry, K.; Rummel, A.; et al. Structure of a bimodular botulinum neurotoxin complex provides insights into its oral toxicity. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Benefield, D.A.; Dessain, S.K.; Shine, N.; Ohi, M.D.; Lacy, D.B. Molecular assembly of botulinum neurotoxin progenitor complexes. Proc. Natl. Acad. Sci. USA 2013, 110, 5630–5635. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Zhong, X.; Gu, S.; Kruel, A.M.; Dorner, M.B.; Perry, K.; Rummel, A.; Dong, M.; Jin, R. Molecular basis for disruption of E-cadherin adhesion by botulinum neurotoxin A complex. Science 2014, 344, 1405–1410. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Rumpel, S.; Zhou, J.; Strotmeier, J.; Bigalke, H.; Perry, K.; Shoemaker, C.B.; Rummel, A.; Jin, R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science 2012, 335, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, T.; Sugawara, Y.; Yutani, M.; Amatsu, S.; Yagita, H.; Kohda, T.; Fukuoka, S.-I.; Nakamura, Y.; Fukuda, S.; Hase, K.; et al. Botulinum toxin A complex exploits intestinal M cells to enter the host and exert neurotoxicity. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pirazzini, M.; Tehran, D.A.; Leka, O.; Zanetti, G.; Rossetto, O.; Montecucco, C. On the translocation of botulinum and tetanus neurotoxins across the membrane of acidic intracellular compartments. Biochim. Biophys. Acta 2015. [Google Scholar] [CrossRef] [PubMed]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M.; et al. Botulinum toxin as a biological weapon: Medical and public health management. J. Am. Med. Assoc. 2001, 285, 1059–1070. [Google Scholar] [CrossRef]
- Dorner, M.B.; Schulz, K.M.; Kull, S.; Dorner, B.G. Complexity of botulinum neurotoxins: Challenges for detection technology. Curr. Top. Microbiol. Immunol. 2013, 364, 219–255. [Google Scholar] [PubMed]
- Adler, S.; Bicker, G.; Bigalke, H.; Bishop, C.; Blumel, J.; Dressler, D.; Fitzgerald, J.; Gessler, F.; Heuschen, H.; Kegel, B.; et al. The current scientific and legal status of alternative methods to the LD50 test for botulinum neurotoxin potency testing. The report and recommendations of a ZEBET Expert Meeting. Altern. Lab. Anim. 2010, 38, 315–330. [Google Scholar] [PubMed]
- Sesardic, T. Bioassays for evaluation of medical products derived from bacterial toxins. Curr. Opin. Microbiol. 2012, 15, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Jones, R.G.; Liu, Y.; Sesardic, D. Measurement of botulinum types A, B and E neurotoxicity using the phrenic nerve-hemidiaphragm: Improved precision with in-bred mice. Toxicon 2009, 53, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Bigalke, B.; Rummel, A. Botulinum neurotoxins: Qualitative and quantitative analysis using the mouse phrenic nerve hemidiaphragm assay (MPN). Toxins 2015, 7. [Google Scholar] [CrossRef]
- Kiris, E.; Kota, K.P.; Burnett, J.C.; Soloveva, V.; Kane, C.D.; Bavari, S. Recent developments in cell-based assays and stem cell technologies for botulinum neurotoxin research and drug discovery. Expert Rev. Mol. Diagn. 2014, 14, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Boyer, A.E.; Moura, H.; Woolfitt, A.R.; Kalb, S.R.; McWilliams, L.G.; Pavlopoulos, A.; Schmidt, J.G.; Ashley, D.L.; Barr, J.R. From the mouse to the mass spectrometer: Detection and differentiation of the endoproteinase activities of botulinum neurotoxins A-G by mass spectrometry. Anal. Chem. 2005, 77, 3916–3924. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Moura, H.; Boyer, A.E.; McWilliams, L.G.; Pirkle, J.L.; Barr, J.R. The use of Endopep-MS for the detection of botulinum toxins A, B, E, and F in serum and stool samples. Anal. Biochem. 2006, 351, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Pirkle, J.L.; Barr, J.R. Mass spectrometric detection of botulinum neurotoxin by measuring its activity in serum and milk. In Detection of Biological Agents for the Prevention of Bioterrorism; Banoub, J., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2011; pp. 115–129. [Google Scholar]
- Parks, B.A.; Shearer, J.D.; Baudys, J.; Kalb, S.R.; Sanford, D.C.; Pirkle, J.L.; Barr, J.R. Quantification of botulinum neurotoxin serotypes A and B from serum using mass spectrometry. Anal. Chem. 2011, 83, 9047–9053. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Baudys, J.; Kalb, S.R.; Barr, J.R. Improved detection of botulinum neurotoxin type A in stool by mass spectrometry. Anal. Biochem. 2011, 412, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Björnstad, K.; Tevell Åberg, A.; Kalb, S.R.; Wang, D.; Barr, J.R.; Bondesson, U.; Hedeland, M. Validation of the Endopep-MS method for qualitative detection of active botulinum neurotoxins in human and chicken serum. Anal. Bioanal. Chem. 2014, 406, 7149–7161. [Google Scholar] [CrossRef] [PubMed]
- Mazuet, C.; Ezan, E.; Volland, H.; Popoff, M.R.; Becher, F. Toxin detection in patients’ sera by mass spectrometry during two outbreaks of type A botulism in France. J. Clin. Microbiol. 2012, 50, 4091–4094. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Baudys, J.; Wang, D.; Barr, J.R. Recommended mass spectrometry-based strategies to identify botulinum neurotoxin-containing samples. Toxins 2015, 7, 1765–1778. [Google Scholar] [CrossRef] [PubMed]
- Morineaux, V.; Mazuet, C.; Hilaire, D.; Enche, J.; Popoff, M.R. Characterization of botulinum neurotoxin type A subtypes by immunocapture enrichment and liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 5559–5570. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Liu, Y.; Sesardic, D. New highly specific botulinum type C1 endopeptidase immunoassays utilising SNAP25 or Syntaxin substrates. J. Immunol. Methods 2009, 343, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.A.; Marks, J.D. Use of a new functional dual coating (FDC) assay to measure low toxin levels in serum and food samples following an outbreak of human botulism. J. Med. Microbiol. 2013, 62, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Rigsby, P.; Sesardic, D.; Marks, J.D.; Jones, R.G. A functional dual-coated (FDC) microtiter plate method to replace the botulinum toxin LD50 test. Anal. Biochem. 2012, 425, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Ekong, T.A.N.; Feavers, I.M.; Sesardic, D. Recombinant SNAP-25 is an effective substrate for Clostridium botulinum type A toxin endopeptidase activity in vitro. Microbiology 1997, 143, 3337–3347. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.R.; Skipper, P.J.; Shone, C.C. An assay for botulinum toxin types A, B and F that requires both functional binding and catalytic activities within the neurotoxin. J. Appl. Microbiol. 2009, 107, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Ekong, T.A.N.; McLellan, K.; Sesardic, D. Immunological detection of Clostridium botulinum toxin type A in therapeutic preparations. J. Immunol. Methods 1995, 180, 181–191. [Google Scholar] [CrossRef]
- Bagramyan, K.; Barash, J.R.; Arnon, S.S.; Kalkum, M. Attomolar detection of botulinum toxin type A in complex biological matrices. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- EQuATox. Available online: http://www.equatox.eu/ (accessed on 30 October 2015).
- Weisemann, J.; Krez, N.; Fiebig, U.; Worbs, S.; Skiba, M.; Endermann, T.; Dorner, M.B.; Bergström, T.; Muñoz, A.; Zegers, I.; et al. Generation and characterisation of six recombinant botulinum neurotoxins as reference material to serve in an international proficiency test. Toxins 2015, 7. [Google Scholar] [CrossRef]
- Worbs, S.; Fiebig, U.; Zeleny, R.; Schimmel, H.; Rummel, A.; Luginbühl, W.; Dorner, B.G. Qualitative and quantitative detection of botulinum neurotoxins from complex matrices: Results of the first international proficiency test. Toxins 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Dorner, M.; Zhang, X.; Hlinak, A.; Dorner, B.; Schade, R. Monitoring of laying capacity, immunoglobulin Y concentration, and antibody titer development in chickens immunized with ricin and botulinum toxins over a two-year period. Poult. Sci. 2009, 88, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Kirchner, S.; Stoermann, B.; Schreiber, T.; Kaulfuss, S.; Schade, R.; Zbinden, R.; Avondet, M.A.; Dorner, M.B.; Dorner, B.G. Simultaneous quantification of five bacterial and plant toxins from complex matrices using a multiplexed fluorescent magnetic suspension assay. Analyst 2009, 134, 2028–2039. [Google Scholar] [CrossRef] [PubMed]
- Kull, S.; Pauly, D.; Störmann, B.; Kirchner, S.; Stämmler, M.; Dorner, M.B.; Lasch, P.; Naumann, D.; Dorner, B.G. Multiplex detection of microbial and plant toxins by immunoaffinity enrichment and matrix-assisted laser desorption/ionization mass spectrometry. Anal. Chem. 2010, 82, 2916–2924. [Google Scholar] [CrossRef] [PubMed]
- Weingart, O.G.; Schreiber, T.; Mascher, C.; Pauly, D.; Dorner, M.B.; Berger, T.F.H.; Egger, C.; Gessler, F.; Loessner, M.J.; Avondet, M.-A.; et al. The case of botulinum toxin in milk: Experimental data. Appl. Environ. Microbiol. 2010, 76, 3293–3300. [Google Scholar] [CrossRef] [PubMed]
- Stevens, G.B.; Silver, D.A.; Zgaga-Griesz, A.; Bessler, W.G.; Vashist, S.K.; Patel, P.; Achazi, K.; Strotmeier, J.; Worbs, S.; Dorner, M.B.; et al. Bioluminescence assay for the highly sensitive detection of botulinum neurotoxin A activity. Analyst 2013, 138, 6154–6162. [Google Scholar] [CrossRef] [PubMed]
- Kull, S.; Schulz, K.M.; Strotmeier, J.W.; Kirchner, S.; Schreiber, T.; Bollenbach, A.; Dabrowski, P.W.; Nitsche, A.; Kalb, S.R.; Dorner, M.B.; et al. Isolation and functional characterization of the novel Clostridium botulinum neurotoxin A8 subtype. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D.; Jones, R.G.; Leung, T.; Alsop, T.; Tierney, R. Detection of antibodies against botulinum toxins. Mov. Disord. 2004, 19, S85–S91. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Ochiai, M.; Liu, Y.; Ekong, T.; Sesardic, D. Development of improved SNAP25 endopeptidase immuno-assays for botulinum type A and E toxins. J. Immunol. Methods 2008, 329, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.G.; Liu, Y.; Halls, C.; Thorpe, S.J.; Longstaff, C.; Matejtschuk, P.; Sesardic, D. Release of proteolytic activity following reduction in therapeutic human serum albumin containing products: Detection with a new neoepitope endopeptidase immunoassay. J. Pharm. Biomed. Anal. 2011, 54, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D.; Leung, T.; Gaines Das, R. Role for standards in assays of botulinum toxins: International collaborative study of three preparations of botulinum type A toxin. Biologicals 2003, 31, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Garcia-Rodriguez, C.; Lou, J.; Baudys, J.; Smith, T.J.; Marks, J.D.; Smith, L.A.; Pirkle, J.L.; Barr, J.R. Extraction of BoNT/A, /B, /E, and /F with a single, high affinity monoclonal antibody for detection of botulinum neurotoxin by Endopep-MS. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Hauschild, A.H.W. Epidemiology of human foodborne botulism. In Clostridium botulinum: Ecology and Control in Foods; Hauschild, A.H.W., Dodds, K.L., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1992; pp. 69–104. [Google Scholar]
- Hill, K.K.; Smith, T.J. Genetic diversity within Clostridium botulinum serotypes, botulinum neurotoxin gene clusters and toxin subtypes. Curr. Top. Microbiol. Immunol. 2013, 364, 1–20. [Google Scholar] [PubMed]
- Thompson, M.; Ellison, S.L.R.; Wood, R. The International Harmonized Protocol for the proficiency testing of analytical chemistry laboratories (IUPAC Technical Report). Pure Appl. Chem. 2006, 78, 145–196. [Google Scholar] [CrossRef]
- Moura, H.; Terilli, R.R.; Woolfitt, A.R.; Gallegos-Candela, M.; McWilliams, L.G.; Solano, M.I.; Pirkle, J.L.; Barr, J.R. Studies on botulinum neurotoxins type/C1 and mosaic/DC using Endopep-MS and proteomics. FEMS Immunol. Med. Microbiol. 2011, 61, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Frevert, J. Pharmaceutical, biological, and clinical properties of botulinum neurotoxin type A products. Drugs R&D 2015, 15, 1–9. [Google Scholar]
- Brin, M.F.; James, C.; Maltman, J. Botulinum toxin type A products are not interchangeable: A review of the evidence. Biologics 2014, 8, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Lévêque, C.; Ferracci, G.; Maulet, Y.; Mazuet, C.; Popoff, M.; Seagar, M.; el Far, O. Direct biosensor detection of botulinum neurotoxin endopeptidase activity in sera from patients with type A botulism. Biosens. Bioelectron. 2014, 57, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Lindström, M.; Korkeala, H. Laboratory diagnostics of botulism. Clin. Microbiol. Rev. 2006, 19, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.W.; Stanker, L.H. Detection of botulinum neurotoxin serotypes A and B using a chemiluminescent versus electrochemiluminescent immunoassay in food and serum. J. Agric. Food Chem. 2013, 61, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Guglielmo-Viret, V.; Attree, O.; Blanco-Gros, V.; Thullier, P. Comparison of electrochemiluminescence assay and ELISA for the detection of Clostridium botulinum type B neurotoxin. J. Immunol. Methods 2005, 301, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.-Y.; Wang, Y.-C.; Tang, S.-S.; Liu, H.-W. A highly sensitive immuno-polymerase chain reaction assay for Clostridium botulinum neurotoxin type A. Toxicon 2004, 43, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Gessler, F.; Pagel-Wieder, S.; Avondet, M.A.; Böhnel, H. Evaluation of lateral flow assays for the detection of botulinum neurotoxin type A and their application in laboratory diagnosis of botulism. Diagn. Microbiol. Infect. Dis. 2007, 57, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Ferreira, J.L.; Eblen, B.S.; Whiting, R.C. Detection of type A, B, E, and F Clostridium botulinum neurotoxins in foods by using an amplified enzyme-linked immunosorbent assay with digoxigenin-labeled antibodies. Appl. Environ. Microbiol. 2006, 72, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Rasooly, R.; Stanker, L.H.; Carter, J.M.; Do, P.M.; Cheng, L.W.; He, X.; Brandon, D.L. Detection of botulinum neurotoxin-A activity in food by peptide cleavage assay. Int. J. Food Microbiol. 2008, 126, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.; Liu, C.Q. Label-free detection of botulinum neurotoxins using a surface plasmon resonance biosensor. In Advances in Immunoassay Technology; Chiu, N.H.L., Ed.; In Tech: Rijeka, Croatia, 2012. [Google Scholar]
- Bruno, J.G.; Richarte, A.M.; Carrillo, M.P.; Edge, A. An aptamer beacon responsive to botulinum toxins. Biosens. Bioelectron. 2012, 31, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Lou, J.; Garcia-Rodriguez, C.; Geren, I.N.; Smith, T.J.; Moura, H.; Marks, J.D.; Smith, L.A.; Pirkle, J.L.; Barr, J.R. Extraction and inhibition of enzymatic activity of botulinum neurotoxins/A1, /A2, and /A3 by a panel of monoclonal anti-BoNT/A antibodies. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Santana, W.I.; Geren, I.N.; Garcia-Rodriguez, C.; Lou, J.; Smith, T.J.; Marks, J.D.; Smith, L.A.; Pirkle, J.L.; Barr, J.R. Extraction and inhibition of enzymatic activity of botulinum neurotoxins /B1, /B2, /B3, /B4, and /B5 by a panel of monoclonal anti-BoNT/B antibodies. BMC Biochem. 2011, 12, 58. [Google Scholar] [CrossRef] [PubMed]
- Kalb, S.R.; Smith, T.J.; Moura, H.; Hill, K.; Lou, J.; Geren, I.N.; Garcia-Rodriguez, C.; Marks, J.D.; Smith, L.A.; Pirkle, J.L.; et al. The use of Endopep-MS to detect multiple subtypes of botulinum neurotoxins A, B, E, and F. Int. J. Mass Spectrom. 2008, 278, 101–108. [Google Scholar] [CrossRef]
- Kalb, S.R.; Baudys, J.; Rees, J.C.; Smith, T.J.; Smith, L.A.; Helma, C.H.; Hill, K.; Kull, S.; Kirchner, S.; Dorner, M.B.; et al. De novo subtype and strain identification of botulinum neurotoxin type B through toxin proteomics. Anal. Bioanal. Chem. 2012, 403, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Gu, S.; Lam, K.H.; Carter, L.G.; Rummel, A.; Mathews, I.I.; Jin, R. Structural basis of the pH-dependent assembly of a botulinum neurotoxin complex. J. Mol. Biol. 2014, 426, 3773–3782. [Google Scholar] [CrossRef] [PubMed]
- Eisele, K.H.; Fink, K.; Vey, M.; Taylor, H.V. Studies on the dissociation of botulinum neurotoxin type A complexes. Toxicon 2011, 57, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Volland, H.; Lamourette, P.; Nevers, M.-C.; Mazuet, C.; Ezan, E.; Neuburger, L.-M.; Popoff, M.; Créminon, C. A sensitive sandwich enzyme immunoassay for free or complexed Clostridium botulinum neurotoxin type A. J. Immunol. Methods 2008, 330, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.L.; Maslanka, S.; Johnson, E.; Goodnough, M. Detection of botulinal neurotoxins A, B, E, and F by amplified enzyme-linked immunosorbent assay: Collaborative study. J. AOAC Int. 2003, 86, 314–331. [Google Scholar] [PubMed]
- Maslanka, S.E.; Lúquez, C.; Raphael, B.H.; Dykes, J.K.; Joseph, L.A. Utility of botulinum toxin ELISA A, B, E, F kits for clinical laboratory investigations of human botulism. Botulinum J. 2011, 2, 72–92. [Google Scholar] [CrossRef]
- Jenko, K.L.; Zhang, Y.; Kostenko, Y.; Fan, Y.; Garcia-Rodriguez, C.; Lou, J.; Marks, J.D.; Varnum, S.M. Development of an ELISA microarray assay for the sensitive and simultaneous detection of ten biodefense toxins. Analyst 2014, 139, 5093–5102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lou, J.; Jenko, K.L.; Marks, J.D.; Varnum, S.M. Simultaneous and sensitive detection of six serotypes of botulinum neurotoxin using enzyme-linked immunosorbent assay-based protein antibody microarrays. Anal. Biochem. 2012, 430, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Garber, E.A.; Venkateswaran, K.V.; O’Brien, T.W. Simultaneous multiplex detection and confirmation of the proteinaceous toxins abrin, ricin, botulinum toxins, and Staphylococcus enterotoxins A, B, and C in food. J. Agric. Food Chem. 2010, 58, 6600–6607. [Google Scholar] [CrossRef] [PubMed]
- Rivera, V.R.; Gamez, F.J.; Keener, W.K.; White, J.A.; Poli, M.A. Rapid detection of Clostridium botulinum toxins A, B, E, and F in clinical samples, selected food matrices, and buffer using paramagnetic bead-based electrochemiluminescence detection. Anal. Biochem. 2006, 353, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Eblen, B.S.; Bull, R.L.; Burr, D.H.; Whiting, R.C. Evaluation of lateral-flow Clostridium botulinum neurotoxin detection kits for food analysis. Appl. Environ. Microbiol. 2005, 71, 3935–3941. [Google Scholar] [CrossRef] [PubMed]
- Chiao, D.J.; Wey, J.J.; Shyu, R.H.; Tang, S.S. Monoclonal antibody-based lateral flow assay for detection of botulinum neurotoxin type A. Hybridoma (Larchmt) 2008, 27, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Chiao, D.J.; Shyu, R.H.; Hu, C.S.; Chiang, H.Y.; Tang, S.S. Colloidal gold-based immunochromatographic assay for detection of botulinum neurotoxin type B. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2004, 809, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Klewitz, T.; Gessler, F.; Beer, H.; Pflanz, K.; Scheper, T. Immunochromatographic assay for determination of botulinum neurotoxin type D. Sens. Actuators B Chem. 2006, 113, 582–589. [Google Scholar] [CrossRef]
- Ching, K.H.; Lin, A.; McGarvey, J.A.; Stanker, L.H.; Hnasko, R. Rapid and selective detection of botulinum neurotoxin serotype-A and -B with a single immunochromatographic test strip. J. Immunol. Methods 2012, 380, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Kohda, T.; Seto, Y.; Mukamoto, M.; Kozaki, S. Improved detection methods by genetic and immunological techniques for botulinum C/D and D/C mosaic neurotoxins. Vet. Microbiol. 2013, 162, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Brunt, J.; Webb, M.D.; Peck, M.W. Rapid affinity immunochromatography column-based tests for sensitive detection of Clostridium botulinum neurotoxins and Escherichia coli O157. Appl. Environ. Microbiol. 2010, 76, 4143–4150. [Google Scholar] [CrossRef] [PubMed]
- Ahn-Yoon, S.; DeCory, T.R.; Durst, R.A. Ganglioside-liposome immunoassay for the detection of botulinum toxin. Anal. Bioanal. Chem. 2004, 378, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Attrée, O.; Guglielmo-Viret, V.; Gros, V.; Thullier, P. Development and comparison of two immunoassay formats for rapid detection of botulinum neurotoxin type A. J. Immunol. Methods 2007, 325, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Gessler, F.; Hampe, K.; Böhnel, H. Sensitive detection of botulinum neurotoxin types C and D with an immunoaffinity chromatographic column test. Appl. Environ. Microbiol. 2005, 71, 7897–7903. [Google Scholar] [CrossRef] [PubMed]
- Tevell Åberg, A.; Björnstad, K.; Hedeland, M. Mass spectrometric detection of protein-based toxins. Biosecur. Bioterror. 2013, 11 (Suppl. S1), S215–S226. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.L. Comparison of amplified ELISA and mouse bioassay procedures for determination of botulinal toxins A, B, E, and F. J. AOAC Int. 2001, 84, 85–88. [Google Scholar] [PubMed]
- Grassi, J.; Frobert, Y.; Pradelles, P.; Chercuitte, F.; Gruaz, D.; Dayer, J.M.; Poubelle, P.E. Production of monoclonal antibodies against interleukin-1α and -1β: Development of two enzyme immunometric assays (EIA) using acetylcholinesterase and their application to biological media. J. Immunol. Methods 1989, 123, 193–210. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Laporte, J.; Savin, C.; Lamourette, P.; Devilliers, K.; Volland, H.; Carniel, E.; Créminon, C.; Simon, S. Fast and sensitive detection of enteropathogenic Yersinia by immunoassays. J. Clin. Microbiol. 2015, 53, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Pauly, D.; Worbs, S.; Kirchner, S.; Shatohina, O.; Dorner, M.B.; Dorner, B.G. Real-time cytotoxicity assay for rapid and sensitive detection of ricin from complex matrices. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simon, S.; Fiebig, U.; Liu, Y.; Tierney, R.; Dano, J.; Worbs, S.; Endermann, T.; Nevers, M.-C.; Volland, H.; Sesardic, D.; et al. Recommended Immunological Strategies to Screen for Botulinum Neurotoxin-Containing Samples. Toxins 2015, 7, 5011-5034. https://doi.org/10.3390/toxins7124860
Simon S, Fiebig U, Liu Y, Tierney R, Dano J, Worbs S, Endermann T, Nevers M-C, Volland H, Sesardic D, et al. Recommended Immunological Strategies to Screen for Botulinum Neurotoxin-Containing Samples. Toxins. 2015; 7(12):5011-5034. https://doi.org/10.3390/toxins7124860
Chicago/Turabian StyleSimon, Stéphanie, Uwe Fiebig, Yvonne Liu, Rob Tierney, Julie Dano, Sylvia Worbs, Tanja Endermann, Marie-Claire Nevers, Hervé Volland, Dorothea Sesardic, and et al. 2015. "Recommended Immunological Strategies to Screen for Botulinum Neurotoxin-Containing Samples" Toxins 7, no. 12: 5011-5034. https://doi.org/10.3390/toxins7124860