Different Assay Conditions for Detecting the Production and Release of Heat-Labile and Heat-Stable Toxins in Enterotoxigenic Escherichia coli Isolates

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
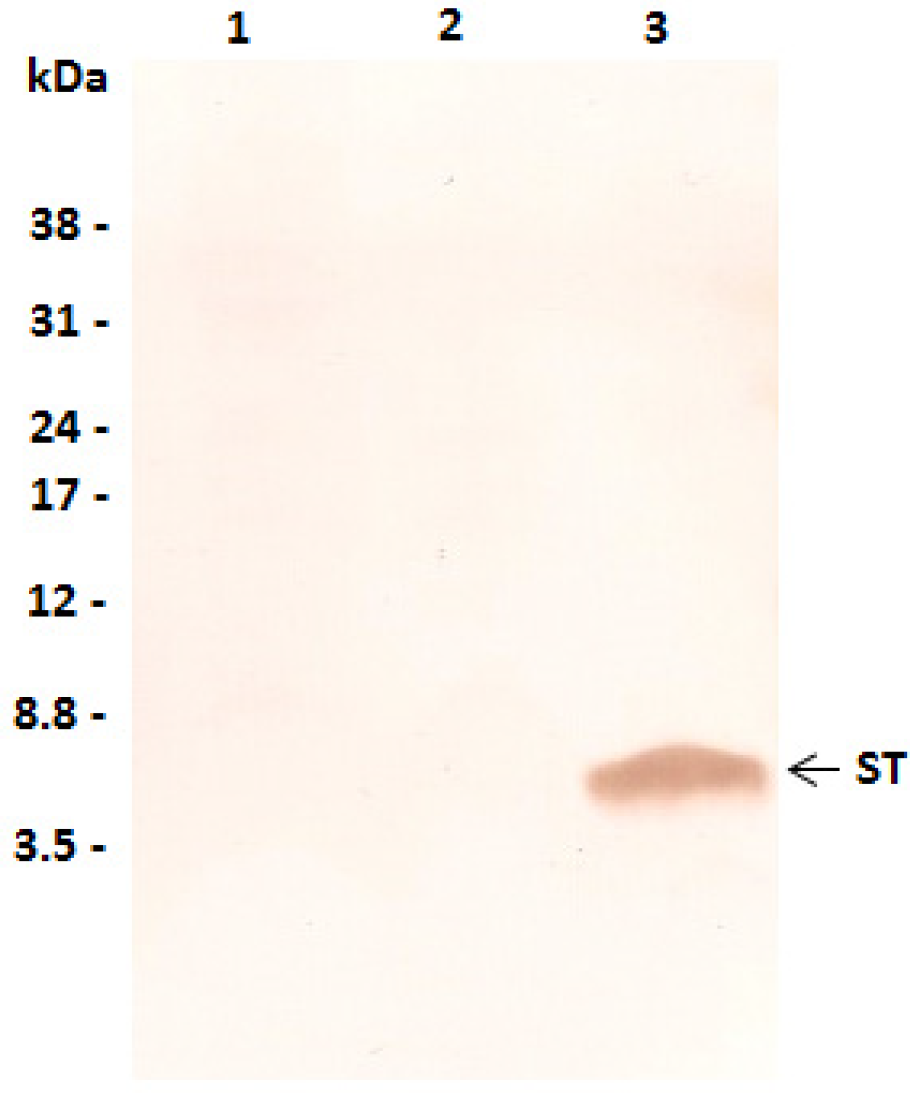
2.1. Characterization of ST MAb

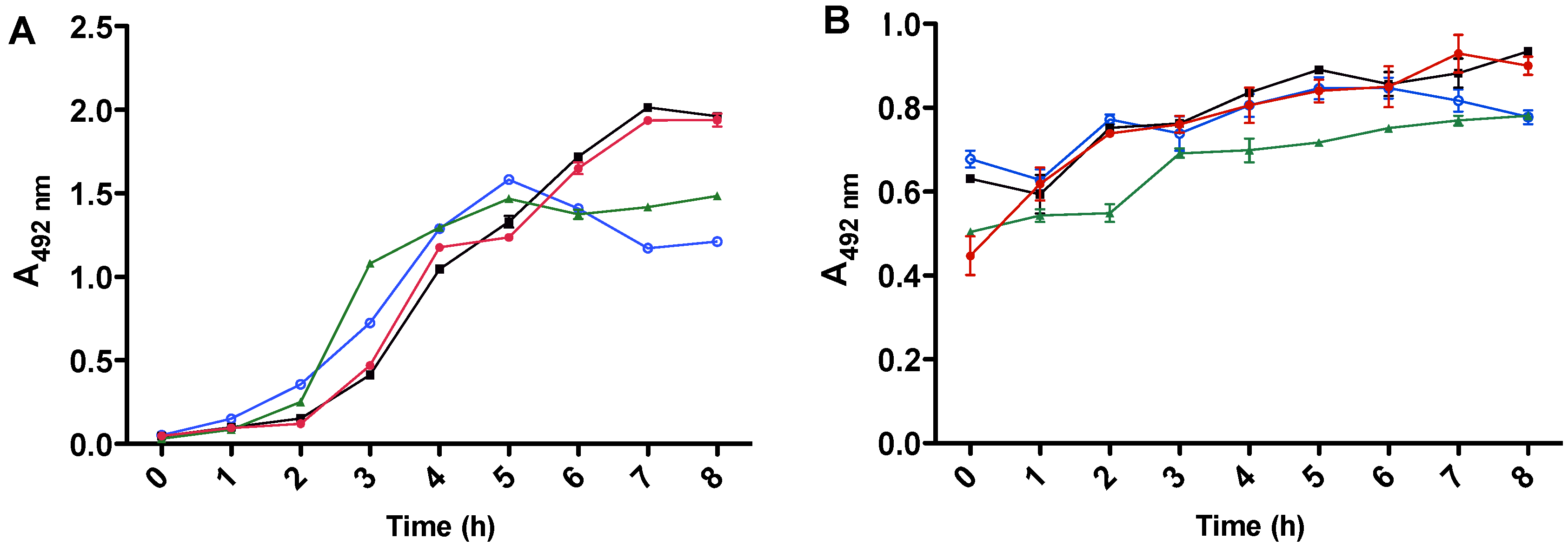
2.2. Comparison of Effects of Different Compounds on Production of LT and ST by E. coli H10407
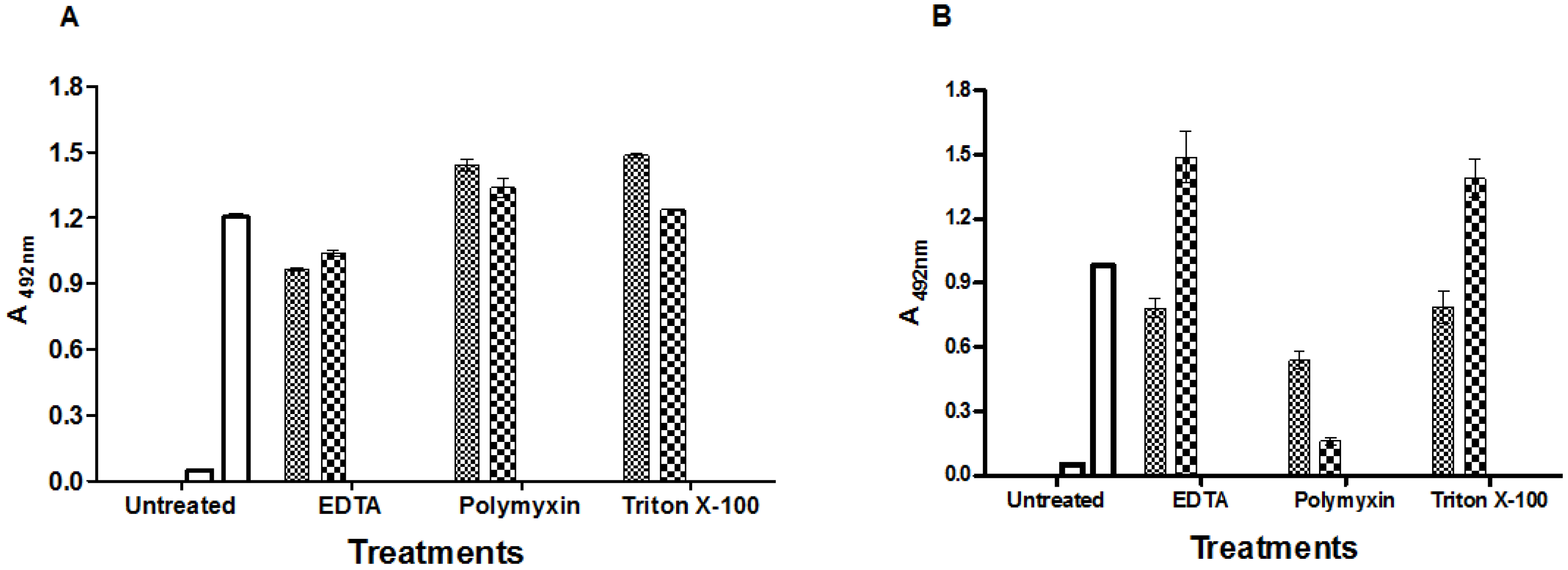
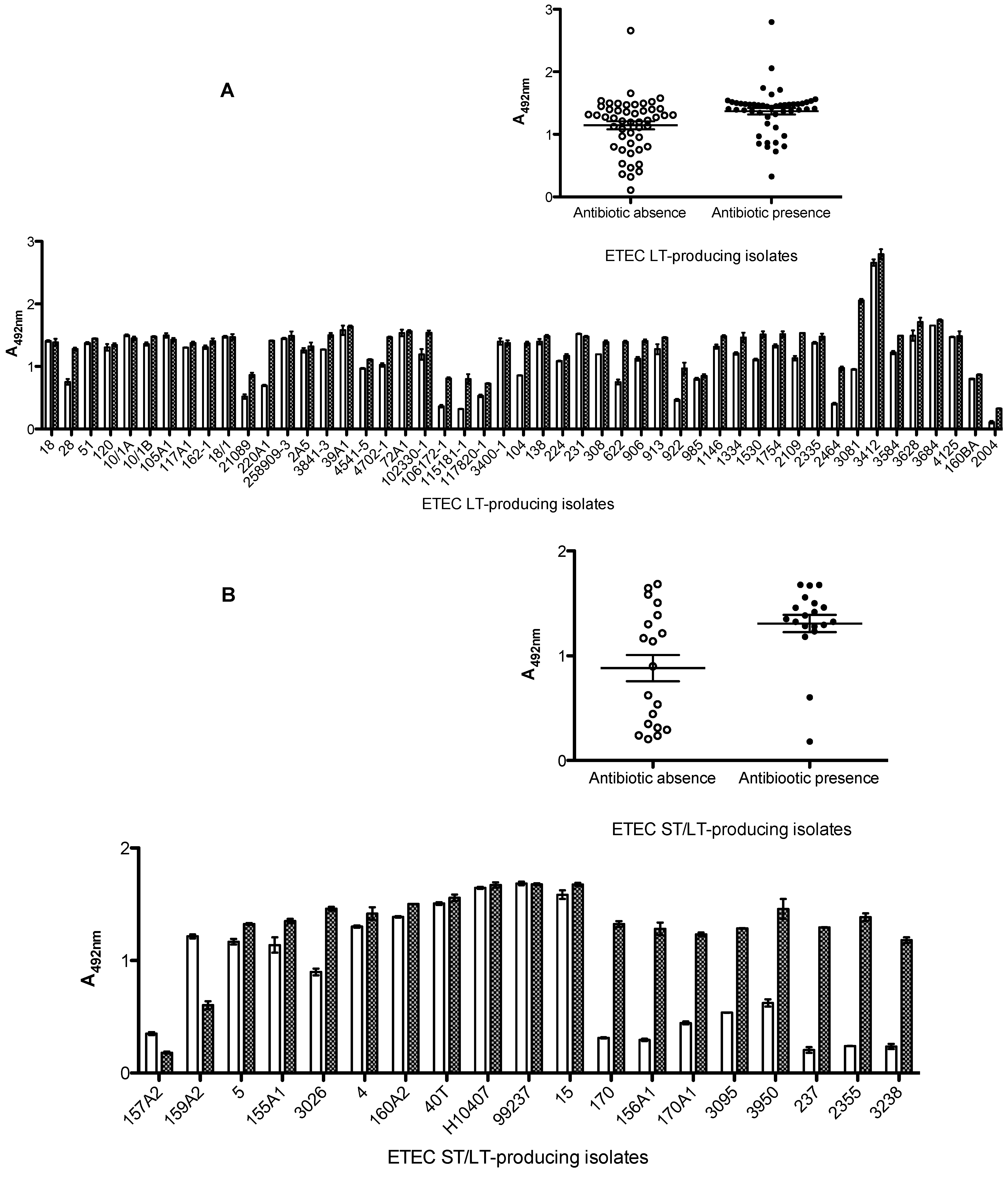
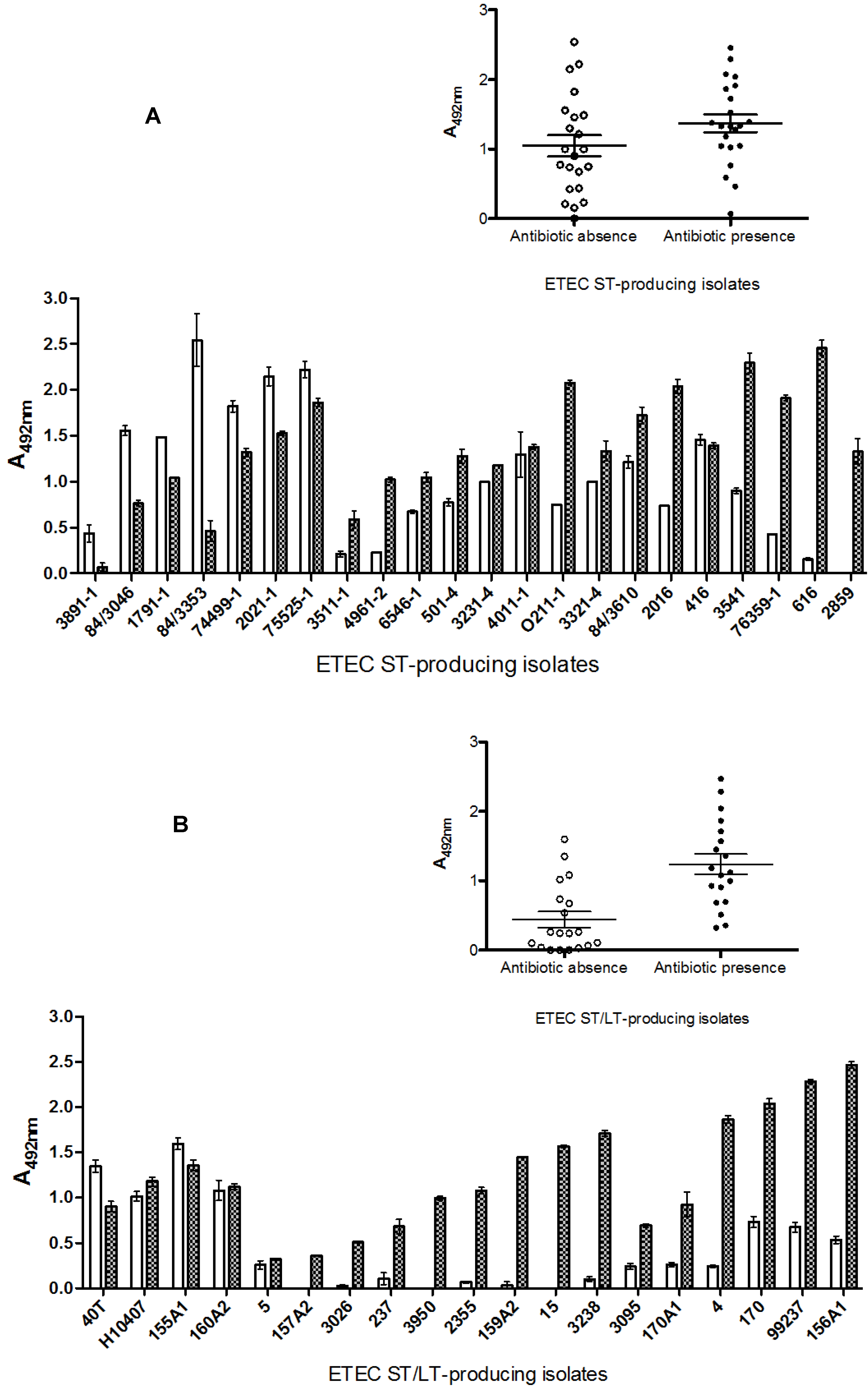
2.3. Effects of Antibiotics on Toxin Production
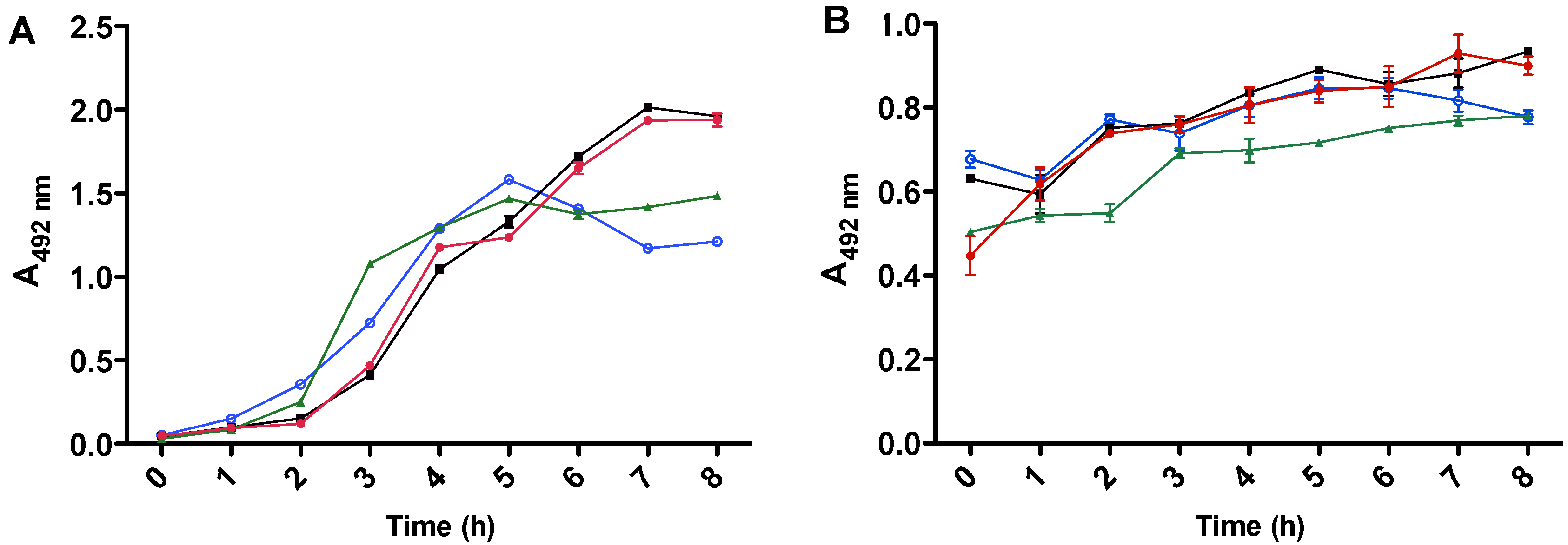
2.4. Characterization of LT and ST Production/Release in ETEC Strains
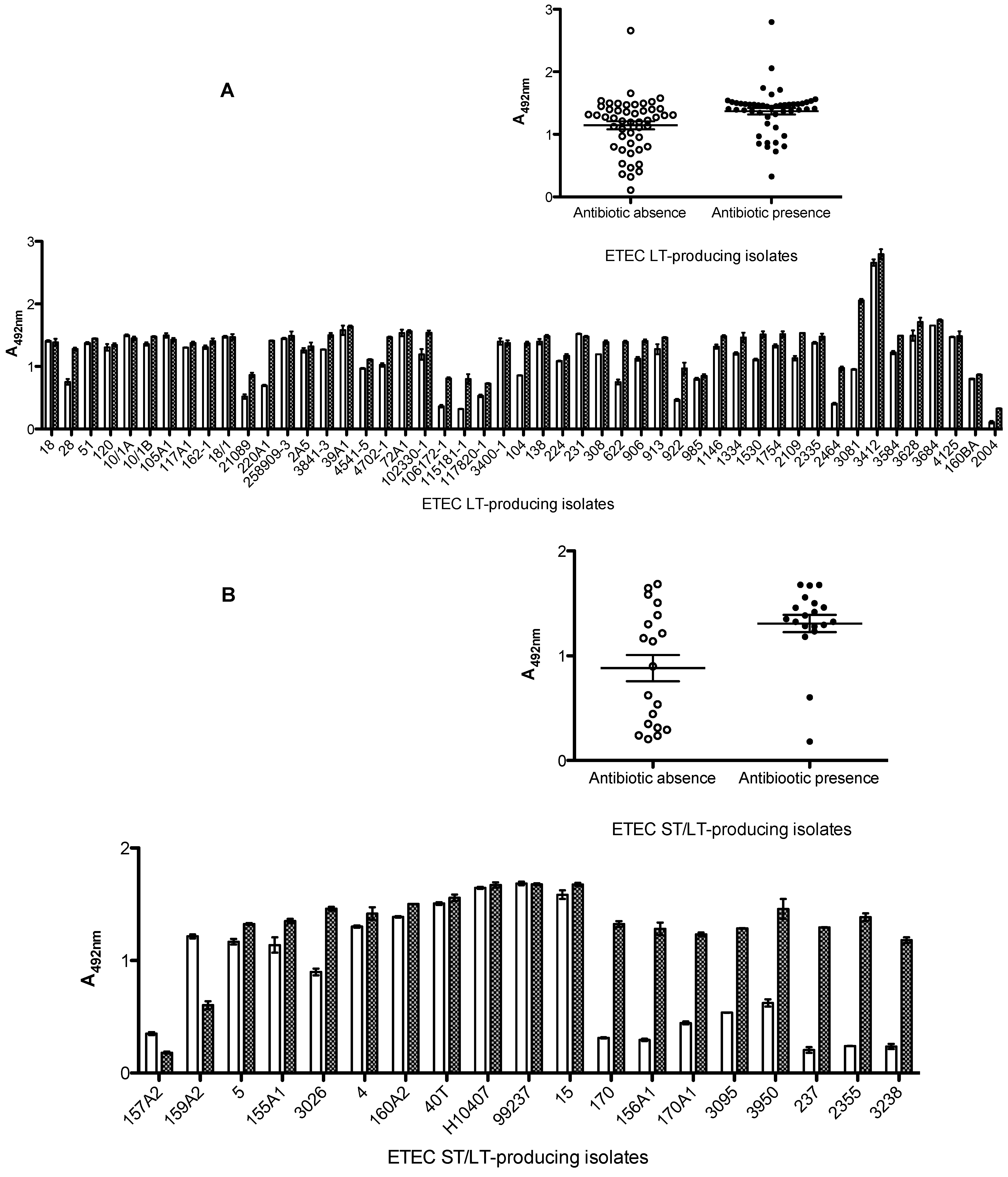
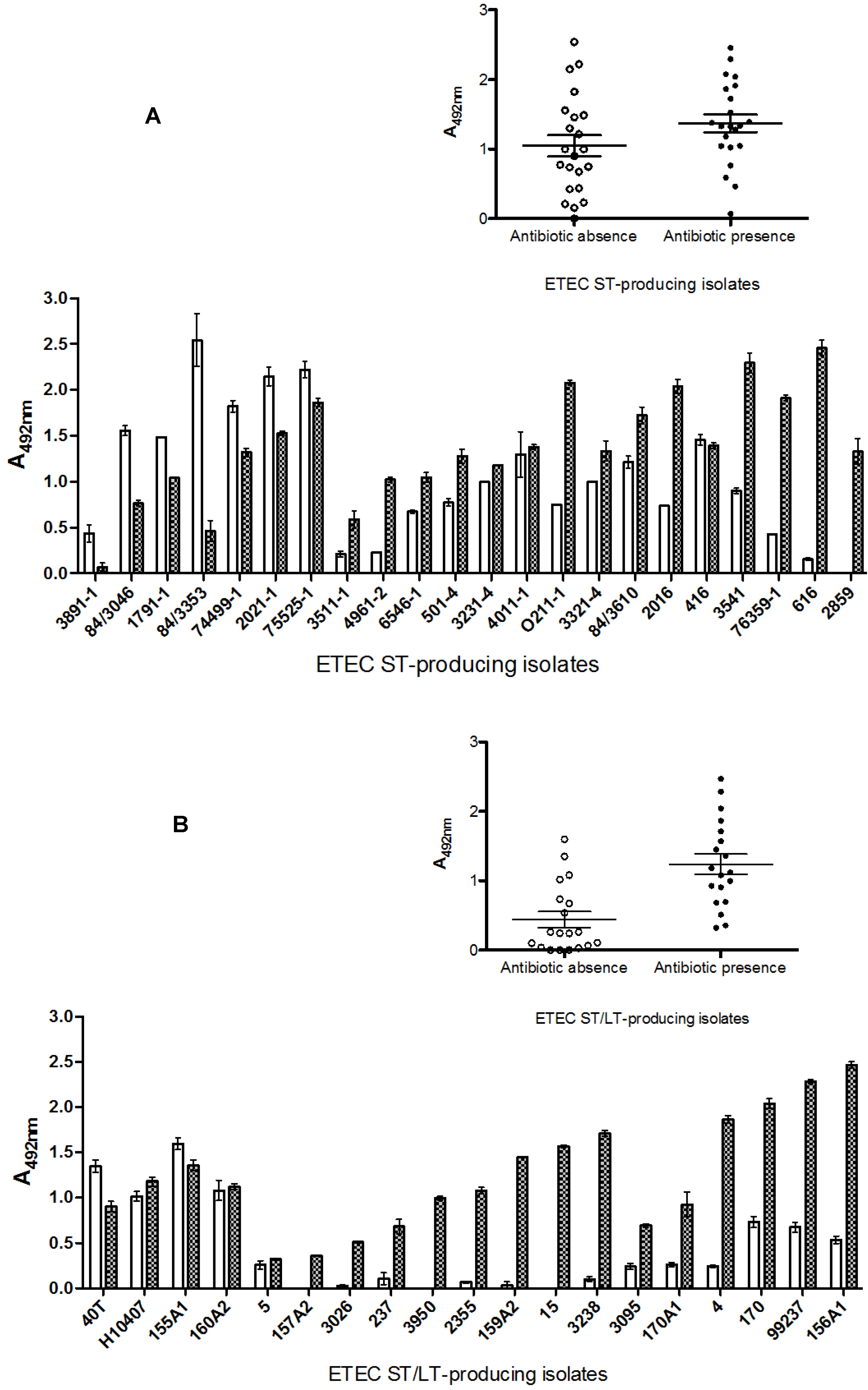
2.5. Discussion
3. Experimental Section
3.1. Bacterial Strains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Serotype | Gene | Toxin expression | Geographic origin/period of isolation |
|---|---|---|---|---|
| 18 | O8:H9 | elt | (LT+) | São Paulo, Brazil, 1994–1996 |
| 28 | O112:H10 | elt | (LT+) | São Paulo, Brazil, 1994–1996 |
| 51 | O62:H19 | elt | (LT+) | São Paulo, Brazil, 1994–1996 |
| 120 | O88:H25 | elt | (LT+) | São Paulo, Brazil, 1994–1996 |
| 10/1A | O114:H21 | elt | (LT+) | Rio de Janeiro, Brazil, 1998 |
| 10/1B | O114:H21 | elt | (LT+) | Rio de Janeiro, Brazil, 1998 |
| 105A1 | ONT:H4 | elt | (LT+) | Paraiba, Brazil, 2000–2001 |
| 117A1 | O6:H16 | elt | (LT+) | Paraiba, Brazil, 2000–2001 |
| 162-1 | O88:H25 | elt | (LT+) | São Paulo, Brazil, 1989–1990 |
| 18/1 | ONT:HNT | elt | (LT+) | Rio de Janeiro, Brazil, 1998 |
| 21089 | ONT:HNT | elt | (LT+) | Rio de Janeiro, Brazil, 1998 |
| 220A1 | O25:H16 | elt | (LT+) | Paraiba, Brazil, 2000–2001 |
| 258909-3 | O128:H1 | elt | (LT+) | Bangladesh, 1979–1984 |
| 2A5 | O88:H25 | elt | (LT+) | Paraiba, Brazil, 2000–2001 |
| 3841-3 | O88:H- | elt | (LT+) | São Paulo, Brazil, 1989–1990 |
| 39A1 | O109:H19 | elt | (LT+) | Paraiba, Brazil, 2000–2001 |
| 4541-5 | O64:H- | elt | (LT+) | São Paulo, Brazil, 1989–1990 |
| 4702-1 | O167:H5 | elt | (LT+) | São Paulo, Brazil, 1989–1990 |
| 72A1 | O15:H40 | elt | (LT+) | Paraiba, Brazil, 2000–2001 |
| 102330-1 | O45:H16 | elt | (LT+) | Bahia, Brazil, 2001–2002 |
| 106172-1 | ONT:H10 | elt | (LT+) | Bahia, Brazil, 2001–2002 |
| 115181-1 | O64:H- | elt | (LT+) | Bahia, Brazil, 2001–2002 |
| 117820-1 | O64:H- | elt | (LT+) | Bahia, Brazil, 2001–2002 |
| 3400-1 | ONT:H4 | elt | (LT+) | Bahia, Brazil, 2001–2002 |
| 104 | OR:H17 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 138 | OR:H25 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 224 | OR:H10 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 231 | OR:H25 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 308 | OR:H16 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 622 | OR:H9 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 906 | O15:H40 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 913 | O6:H16 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 922 | O64:HNT | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 985 | O166:H15 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 1146 | OR:H21 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 1334 | ONT:H16 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 1530 | OR:H19 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 1754 | OR:H21 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 2109 | ONT:H32 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 2335 | OR:H51 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 2464 | OR:H10 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 3081 | OR:H- | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 3412 | O109:H- | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 3584 | OR:H32 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 3628 | ONT:H40 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 3684 | O133:H25 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 4125 | O82:H40 | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 2004 | ONT:H- | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 160BA | OR:H- | elt | (LT+) | Bahia, Brazil, 2003–2004 |
| 3321-4 | O153:H45 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 616 | O25:H16 | estA | (ST+) | Bahia, Brazil, 2003–2004 |
| 2016 | O8:H2 | estA | (ST+) | Bahia, Brazil, 2003–2004 |
| 6546-1 | ONT:H32 | estA | (ST+) | Bahia, Brazil, 2001–2002 |
| 416 | O148:H27 | estA | (ST+) | Bahia, Brazil, 2003–2004 |
| 2859 | O78:H27 | estA | (ST+) | Bahia, Brazil, 2003–2004 |
| 3541 | ONT:HNT | estA | (ST+) | Bahia, Brazil, 2003–2004 |
| 74499-1 | O23:H15 | estA | (ST+) | Bahia, Brazil, 2003–2004 |
| 76359-1 | O166:H15 | estA | (ST+) | Bahia, Brazil, 2001–2002 |
| 1791-1 | O29:H21 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 4961-2 | O29:H21 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 501-4 | O78:H12 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| O211-1 | O6:H16 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 3231-4 | ONT:H[NT] | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 2021-1 | O128ac:H27 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 3511-1 | O128ac:H21 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 4011-1 | O153:H45 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 3891-1 | O153:H45 | estA | (ST+) | São Paulo, Brazil, 1989–1990 |
| 75525-1 | O27:H7 | estA | (ST+) | Bahia, Brazil, 2001–2002 |
| 84/3353 | O4:H1 | estA | (ST+) | Germany, 1984 |
| 84/3046 | O6:HNT | estA | (ST+) | Germany, 1984 |
| 84/3610 | ONT:H5 | estA | (ST+) | Germany, 1984 |
| H10407 | O78:H11 | elt/estA | (LT+ ST+) | Bangladesh, 1973 |
| 4 | O6:H16 | elt/estA | (LT+ ST+) | São Paulo, Brazil, 1994–1996 |
| 5 | O6:H16 | elt/estA | (LT+ ST+) | São Paulo, Brazil, 1994–1996 |
| 155A1 | O6:H16 | elt/estA | (LT+ ST+) | Paraiba, Brazil, 2000–2001 |
| 156A1 | O6:H16 | elt/estA | (LT+ ST+) | Paraiba, Brazil, 2000–2001 |
| 157A2 | O6:H16 | elt/estA | (LT+ ST+) | Paraiba, Brazil, 2000–2001 |
| 159A2 | O6:H16 | elt/estA | (LT+ ST+) | Paraiba, Brazil, 2000–2001 |
| 160A2 | O6:H16 | elt/estA | (LT+ ST+) | Paraiba, Brazil, 2000–2001 |
| 170A1 | O6:H16 | elt/estA | (LT+ ST+) | Paraiba, Brazil, 2000–2001 |
| 40T | OR:H- | elt/estA | (LT+ ST+) | São Paulo, Brazil, 1989–1990 |
| 99237 | O78:H12 | elt/estA | (LT+ ST+) | Rio de Janeiro, Brazil, 1998 |
| 237 | O6:H16 | elt/estA | (LT+ ST+) | Bahia, Brazil, 2003–2004 |
| 2355 | OR:H16 | elt/estA | (LT+ ST+) | Bahia, Brazil, 2003–2004 |
| 3026 | O6:H16 | elt/estA | (LT+ ST+) | Bahia, Brazil, 2003–2004 |
| 3095 | O6:H16 | elt/estA | (LT+ ST+) | Bahia, Brazil, 2003–2004 |
| 3238 | O6:H16 | elt/estA | (LT+ ST+) | Bahia, Brazil, 2003–2004 |
| 3950 | O6:H16 | elt/estA | (LT+ ST+) | Bahia, Brazil, 2003–2004 |
| 15 | ONT:H19 | elt/estA | (LT+ ST+) | São Paulo, Brazil, 1994–1996 |
| 170 | O6:H16 | elt/estA | (LT+ ST+) | São Paulo, Brazil, 1994–1996 |
3.2. O:H Identification
3.3. Media, Culture Conditions and Treatments for Toxin Release
3.4. Rabbit Polyclonal and Mouse Monoclonal (MAb) Anti-LT Antibodies
3.5. Rabbit Polyclonal and Mouse Monoclonal (MAb) Anti-ST Antibodies
3.6. Evaluation of Anti-ST MAb Reactivity
3.7. Indirect ELISA for ST
3.8. Capture ELISA for ST
3.9. cELISA for LT
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Diarrhea: Why Children are Still Dying and What can be Done. In United Nations Children’s Fund; WHO Press: New York, NY, USA, 2009; p. 68. [Google Scholar]
- Petri, W.A., Jr.; Miller, M.; Binder, H.J.; Levine, M.M.; Dillingham, R.; Guerrant, R.L. Enteric infections, diarrhea, and their impact on function and development. J. Clin. Investig. 2008, 118, 1277–1290. [Google Scholar] [CrossRef]
- Qadri, F.; Svennerholm, A.M.; Faruque, A.S.; Sack, R.B. Enterotoxigenic Escherichia coli in developing countries: Epidemiology, microbiology, clinical features, treatment, and prevention. Clin. Microbiol. Rev. 2005, 18, 465–483. [Google Scholar] [CrossRef]
- Gaastra, W.; Svennerholm, A.M. Colonization factors of human enterotoxigenic Escherichia coli (ETEC). Trends Microbiol. 1996, 4, 444–452. [Google Scholar] [CrossRef]
- So, M.; McCarthy, B.J. Nucleotide sequence of transposon Tn1681 encoding a heat-stable toxin (ST) and its identification in enterotoxigenic Escherichia coli strains. Proc. Natl. Acad. Sci. USA 1980, 77, 4011–4015. [Google Scholar] [CrossRef]
- Rasheed, J.K.; Guzmán-Verduzco, L.M.; Kupersztoch, Y.M. Two precursors of the heat-stable enterotoxin of Escherichia coli: Evidence of extracellular processing. Mol. Microbiol. 1990, 4, 265–273. [Google Scholar] [CrossRef]
- Thompson, M.R.; Giannella, R.A. Revised amino acid sequence for a heat-stable enterotoxin produced by an Escherichia coli strain (18D) that is pathogenic for humans. Infect. Immun. 1985, 47, 834–836. [Google Scholar]
- Schulz, S.; Green, C.K.; Yuen, P.S.; Garbers, D.L. Guanylyl cyclase is a heat-stable enterotoxin receptor. Cell 1990, 63, 941–948. [Google Scholar] [CrossRef]
- Hughes, J.M.; Murad, F.; Chang, B.; Guerrant, R.L. Role of cyclic GMP in the action of heat-stable enterotoxin of Escherichia coli. Nature 1978, 271, 755–756. [Google Scholar] [CrossRef]
- Spangler, B.D. Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol. Rev. 1992, 56, 622–647. [Google Scholar]
- Hirst, T.R.; Holmgren, J. Conformation of protein secreted across bacterial outer membranes: A study of enterotoxin translocation from Vibrio cholerae. Proc. Natl. Acad. Sci. USA 1987, 84, 7418–7422. [Google Scholar] [CrossRef]
- Wandersman, C. Secretion across the bacterial outer membrane. In Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed.; Neidhardt, F.C., Curtis, R., III, Ingraham, J.L., et al., Eds.; American Society for Microbiology: Washington, DC, USA, 1996; Volume 1, pp. 955–966. [Google Scholar]
- Tauschek, M.; Gorrell, R.J.; Strugnell, R.A.; Robins-Browne, R.M. Identification of a protein secretory pathway for the secretion of heat-labile enterotoxin by an enterotoxigenic strain of Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 14, 7066–7071. [Google Scholar]
- Brown, E.A.; Hardwige, P.R. Biochemical characterization of the enterotoxigenic Escherichia coli LeoA protein. Microbiology 2007, 153, 3776–3784. [Google Scholar] [CrossRef]
- Fleckenstein, J.M.; Hardwige, P.R.; Munson, G.P.; Rasko, D.A.; Sommerfelt, H.; Steinsland, H. Molecular mechanisms of enterotoxigenic Escherichia coli infection. Microbes Infect. 2010, 12, 89–98. [Google Scholar] [CrossRef]
- Lencer, W.I.; Hirst, T.R.; Holmes, R.K. Membrane traffic and the cellular uptake of cholera toxin. Biochim. Biophys. Acta 1999, 8, 177–190. [Google Scholar] [CrossRef]
- Zafriri, D.; Oron, Y.; Eisenstein, B.I.; Ofek, I. Growth advantage and enhanced toxicity of Escherichia coli adherent to tissue culture cells due to restricted diffusion of products secreted by the cells. J. Clin. Invest. 1987, 79, 1210–1216. [Google Scholar] [CrossRef]
- Kesty, N.C.; Mason, K.M.; Reedy, M.; Miller, S.E.; Kuehn, M.J. Enterotoxigenic Escherichia coli vesicles target toxin delivery into mammalian cells. EMBO J. 2004, 23, 4538–4549. [Google Scholar] [CrossRef]
- Dorsey, F.C.; Fischer, J.F.; Fleckenstein, J.M. Directed delivery of heat-labile enterotoxin by enterotoxigenic Escherichia coli. Cell Microbiol. 2006, 8, 1516–1527. [Google Scholar] [CrossRef]
- Buddelmeijer, N.; Krehenbrink, M.; Pecorari, F.; Pugsley, A.P. Type II secretion system secretin PulD localizes in clusters in the Escherichia coli outer membrane. J. Bacteriol. 2009, 191, 161–168. [Google Scholar] [CrossRef]
- Chapman, P.A.; Swift, D.L. A simplified method for detecting the heat-labile enterotoxin of Escherichia coli. J. Med. Microbiol. 1984, 18, 399–403. [Google Scholar] [CrossRef]
- Levner, M.; Wiener, F.P.; Rubin, B.A. Induction of Escherichia coli and Vibrio cholerae enterotoxins by an inhibitor of protein synthesis. Infect. Immun. 1977, 15, 132–137. [Google Scholar]
- Ristaino, P.A.; Levine, M.M.; Young, C.R. Improved GM1-enzyme-linked immunosorbent assay for detection of Escherichia coli heat-labile enterotoxin. J. Clin. Microbiol. 1983, 18, 808–815. [Google Scholar]
- Menezes, C.A.; Gonçalves, D.S.; Amianti, J.; Fernandes, I.; Taddei, C.R.; Koga, P.C.M.; Trabulsi, L.R.; Martinez, M.B.; Piazza, R.M.F. Capture immunoassay for LT detection produced by enterotoxigenic Escherichia coli in bacterial isolates. Braz. J. Microbiol. 2003, 34, 11–13. [Google Scholar] [CrossRef]
- Menezes, C.A.; Imamura, S.Y.; Trabulsi, L.R.; Fernandes-Filho, A.; Martinez, M.B.; Guth, B.E.C.; Girão, D.M.; Piazza, R.M.F. Production, characterization and application of antibodies against heat-labile type-I toxin for detection of enterotoxigenic Escherichia coli. Mem. Inst. Oswaldo Cruz 2006, 101, 875–880. [Google Scholar]
- Rocha, L.B.; Piazza, R.M.F. Production of Shiga toxin by Shiga toxin-expressing Escherichia coli (STEC) in broth media: from divergence to definition. Lett. Appl. Microbiol. 2007, 45, 411–417. [Google Scholar]
- Sahl, J.W.; Rasko, D.A. Analysis of global transcriptional profiles of enterotoxigenic Escherichia coli isolate E24377A. Infect. Immun. 2012, 80, 1232–1242. [Google Scholar] [CrossRef]
- Hunt, P.D.; Hardy, S.J. Heat-labile enterotoxin can be released from Escherichia coli cells by host intestinal factors. Infect. Immun. 1991, 59, 168–171. [Google Scholar]
- Horstman, A.L.; Kuehn, M.J. Enterotoxigenic Escherichia coli secretes active heat-labile enterotoxin via outer membrane vesicles. J. Biol. Chem. 2000, 275, 12489–12496. [Google Scholar] [CrossRef]
- Levner, M.; Urbano, C.; Rubin, B.A. Polymyxin B release of unnicked cholera toxin subunit A. J. Bacteriol. 1980, 144, 1203–1204. [Google Scholar]
- Bowman, C.C.; Clements, J.D. Differential biological and adjuvant activities of cholera toxin and Escherichia coli heat labile enterotoxin hybrids. Infect. Immun. 2001, 69, 1528–1535. [Google Scholar] [CrossRef]
- Drevet, P.; Guinet, R. Comparison of sandwich-ELISA and GM1-ELISA for the detection of Escherichia coli thermolabile enterotoxin. J. Immunoass. 1991, 12, 293–304. [Google Scholar] [CrossRef]
- Lasaro, M.A.; Rodrigues, J.F.; Cabrera-Crespo, J.; Sbrogio-Almeida, M.E.; Lasaro, M.O.; Ferreira, L.C.S. Evaluation of experimental conditions for quantification of LT produced by human derived enterotoxigenic Escherichia coli strains. Braz. J. Microbiol. 2007, 38, 446–451. [Google Scholar] [CrossRef]
- Qu, Z.-H.; Boesman-Finkelstein, M.; Finkelstein, R.A. Urea-induced release of heat-labile enterotoxin from Escherichia coli. J. Clin. Microbiol. 1991, 29, 773–777. [Google Scholar]
- Clements, J.D.; Finkelstein, R.A. Isolation and characterization of homogeneous heat-labile enterotoxins with high specific activity from Escherichia coli cultures. Infect. Immun. 1979, 24, 760–769. [Google Scholar]
- Minami, J.; Okabe, A.; Nagata, A.; Hayashi, H. Quantitative analysis of the production of heat-labile enterotoxin by enterotoxigenic Escherichia coli. Acta Med. Okayama 1984, 38, 461–469. [Google Scholar]
- Yoh, M.; Yamamoto, K.; Honda, T.; Takeda, Y.; Miwatani, T. Effects of lincomycin and tetracycline on production and properties of enterotoxins of enterotoxigenic Escherichia coli. Infect. Immun. 1983, 42, 778–782. [Google Scholar]
- Sjöling, A.; Wiklund, G.; Savarino, S.J.; Cohen, D.I.; Svennerholm, A.M. Comparative analyses of phenotypic and genotypic methods for detection of enterotoxigenic Escherichia coli toxins and colonization factors. J. Clin. Microbiol. 2007, 45, 3295–3301. [Google Scholar] [CrossRef]
- Busque, P.; Letellier, A.; Harel, J.; Dubreuil, J.D. Production of Escherichia coli STb enterotoxin is subject to catabolite repression. Microbiology 1995, 141, 1621–1627. [Google Scholar] [CrossRef]
- Erume, J.; Berberov, E.M.; Moxley, R.A. Comparison of the effects of different nutrient media on production of heat-stable enterotoxin-b by Escherichia coli. Vet. Microbiol. 2010, 144, 160–165. [Google Scholar] [CrossRef]
- O’Brien, A.D.; Newland, J.W.; Miller, S.F.; Holmes, R.K. Shiga-like toxin-converting phages from Escherichia coli strains that cause hemorrhagic colitis or infantile diarrhea. Science 1984, 226, 694–696. [Google Scholar]
- Lasaro, M.A.; Rodrigues, J.F.; Mathias-Santos, C.; Guth, B.E.; Régua-Mangia, A.; Piantino-Ferreira, A.J.; Takagi, M.; Cabrera-Crespo, J.; Sbrogio-Almeida, M.E.; Ferreira, L.C.S. Production and release of heat-labile toxin by wild-type human-derived enterotoxigenic Escherichia coli. FEMS Immunol. Med. Microbiol. 2006, 48, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.J.; Evans, D.G. Three characteristics associated with enterotoxigenic Escherichia coli isolated from man. Infect. Immun. 1973, 8, 322–328. [Google Scholar]
- Guth, B.E.C.; Aguiar, E.G.; Griffin, P.M.; Ramos, S.R.; Gomes, T.A.T. Prevalence of colonization factor antigens (CFAs) and adherence to HeLa cells in enterotoxigenic Escherichia coli isolated from feces of children in São Paulo. Microbiol. Immunol. 1994, 38, 695–701. [Google Scholar] [CrossRef]
- Bueris, V.; Sircili, M.P.; Taddei, C.R.; Santos, M.F.; Franzolin, M.R.; Martinez, M.B.; Ferrer, S.R.; Barreto, M.L.; Trabulsi, L.R. Detection of diarrheagenic Escherichia coli from children with and without diarrhea in Salvador, Bahia, Brazil. Mem. Inst. Oswaldo Cruz 2007, 102, 839–844. [Google Scholar] [CrossRef]
- Gomes, T.A.T.; Rassi, V.; MacDonald, K.L.; Ramos, S.R.T.S.; Trabulsi, L.R.; Vieira, M.A.M.; Guth, B.E.C.; Candeias, J.A.N.; Ivey, C.; Toledo, M.R.F.; et al. Enteropathogens associated with acute diarrheal disease in urban infants in São Paulo, Brazil. J. Infect. Dis. 1991, 164, 331–337. [Google Scholar] [CrossRef]
- Moreno, A.C.R.; Fernandes-Filho, A.; Gomes, T.A.T.; Ramos, S.T.S.; Montemor, L.P.G.; Tavares, V.C.; Santos-Filho, L.; Irino, K.; Martinez, M.B. Etiology of childhood diarrhea in the northeast of Brazil: significant emergent diarrheal pathogens. Diagn. Microbiol. Infect. Dis. 2010, 66, 50–57. [Google Scholar] [CrossRef]
- Nishimura, L.S.; Ferreira, L.C.S.; Pacheco, A.B.F.; Guth, B.E.C. Relationship between outer membrane protein and lipopolysaccharide profiles and serotypes of enterotoxigenic Escherichia coli isolated in Brazil. FEMS Microbiol. Lett. 1996, 143, 253–258. [Google Scholar] [CrossRef]
- Prada, J.; Baljer, G.; de Rycke, J.; Steinrück, H.; Zimmermann, S.; Stephan, R.; Beutin, L. Characteristics of alpha-hemolytic strains of Escherichia coli isolated from dogs with gastroenteritis. Vet. Microbiol. 1991, 29, 59–73. [Google Scholar] [CrossRef]
- Gothefors, L.; Ahren, C.; Stoll, B.; Barua, D.K.; Orskov, F.; Salek, M.A.; Svennerholm, A.M. Presence of colonization factor antigens on fresh isolates of fecal Escherichia coli: A prospective study. J. Infect. Dis. 1985, 152, 1128–1133. [Google Scholar]
- Franzolin, M.R.; Alves, R.C.; Keller, R.; Gomes, T.A.T.; Beutin, L.; Barreto, M.L.; Milroy, C.; Strina, A.; Ribeiro, H.; Trabulsi, L.R. Prevalence of diarrheagenic Escherichia coli in children with diarrhea in Salvador, Bahia, Brazil. Mem. Inst. Oswaldo Cruz. 2005, 100, 359–363. [Google Scholar] [CrossRef]
- Souza, E.C.; Martinez, M.B.; Taddei, C.R.; Mukai, L.; Gilio, A.E.; Racz, M.L.; Silva, L.; Ejzenberg, B.; Okay, Y. Etiologic profile of acute diarrhea in children in São Paulo. J. Pediatr. 2002, 78, 31–38. [Google Scholar] [CrossRef]
- Ewing, W.H. Edwards and Ewing’s Identification of Enterobacteriaceae, 4th ed.; Elsevier: New York, NY, USA, 1986. [Google Scholar]
- Staples, S.J.; Acher, S.E.; Gianella, R.A. Purification and characterization of heat-stable enterotoxin produced by a strain of E. coli pathogenic for man. J. Biol. Chem. 1980, 255, 4716–4721. [Google Scholar]
- Lockwood, D.E.; Robertson, D.C. Development of a competitive enzyme-linked immunosorbent assay (ELISA) for Escherichia coli heat-stable enterotoxin (STa). J. Immunol. Methods 1984, 75, 295–307. [Google Scholar] [CrossRef]
- McKinney, M.M.; Parkinson, A. A simple, nonchromatographic procedure to purify immunoglobulins from serum and ascites fluid. J. Immunol. Methods 1987, 96, 271–278. [Google Scholar] [CrossRef]
- Menezes, M.A.; Rocha, L.B.; Koga, P.C.; Fernandes, I.; Nara, J.M.; Magalhães, C.A.; Abe, C.M.; Ayala, C.O.; Burgos, Y.K.; Elias, W.P.; et al. Identification of enteropathogenic and enterohaemorrhagic Escherichia coli strains by immunoserological detection of intimin. J. Appl. Microbiol. 2010, 108, 878–887. [Google Scholar] [CrossRef]
- Rocha, L.B.; Luz, D.E.; Moraes, C.T.; Caravelli, A.; Fernandes, I.; Guth, B.E.; Horton, D.S.P.Q.; Piazza, R.M.F. Interaction between Shiga toxin and monoclonal antibodies: Binding characteristics and in vitro neutralizing abilities. Toxins 2012, 4, 729–747. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rocha, L.B.; Ozaki, C.Y.; Horton, D.S.P.Q.; Menezes, C.A.; Silva, A.; Fernandes, I.; Magnoli, F.C.; Vaz, T.M.I.; Guth, B.E.C.; Piazza, R.M.F. Different Assay Conditions for Detecting the Production and Release of Heat-Labile and Heat-Stable Toxins in Enterotoxigenic Escherichia coli Isolates. Toxins 2013, 5, 2384-2402. https://doi.org/10.3390/toxins5122384
Rocha LB, Ozaki CY, Horton DSPQ, Menezes CA, Silva A, Fernandes I, Magnoli FC, Vaz TMI, Guth BEC, Piazza RMF. Different Assay Conditions for Detecting the Production and Release of Heat-Labile and Heat-Stable Toxins in Enterotoxigenic Escherichia coli Isolates. Toxins. 2013; 5(12):2384-2402. https://doi.org/10.3390/toxins5122384
Chicago/Turabian StyleRocha, Letícia B., Christiane Y. Ozaki, Denise S. P. Q. Horton, Caroline A. Menezes, Anderson Silva, Irene Fernandes, Fabio C. Magnoli, Tania M. I. Vaz, Beatriz E. C. Guth, and Roxane M. F. Piazza. 2013. "Different Assay Conditions for Detecting the Production and Release of Heat-Labile and Heat-Stable Toxins in Enterotoxigenic Escherichia coli Isolates" Toxins 5, no. 12: 2384-2402. https://doi.org/10.3390/toxins5122384