Effects of Vitamin D3, Calcipotriol and FTY720 on the Expression of Surface Molecules and Cytolytic Activities of Human Natural Killer Cells and Dendritic Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
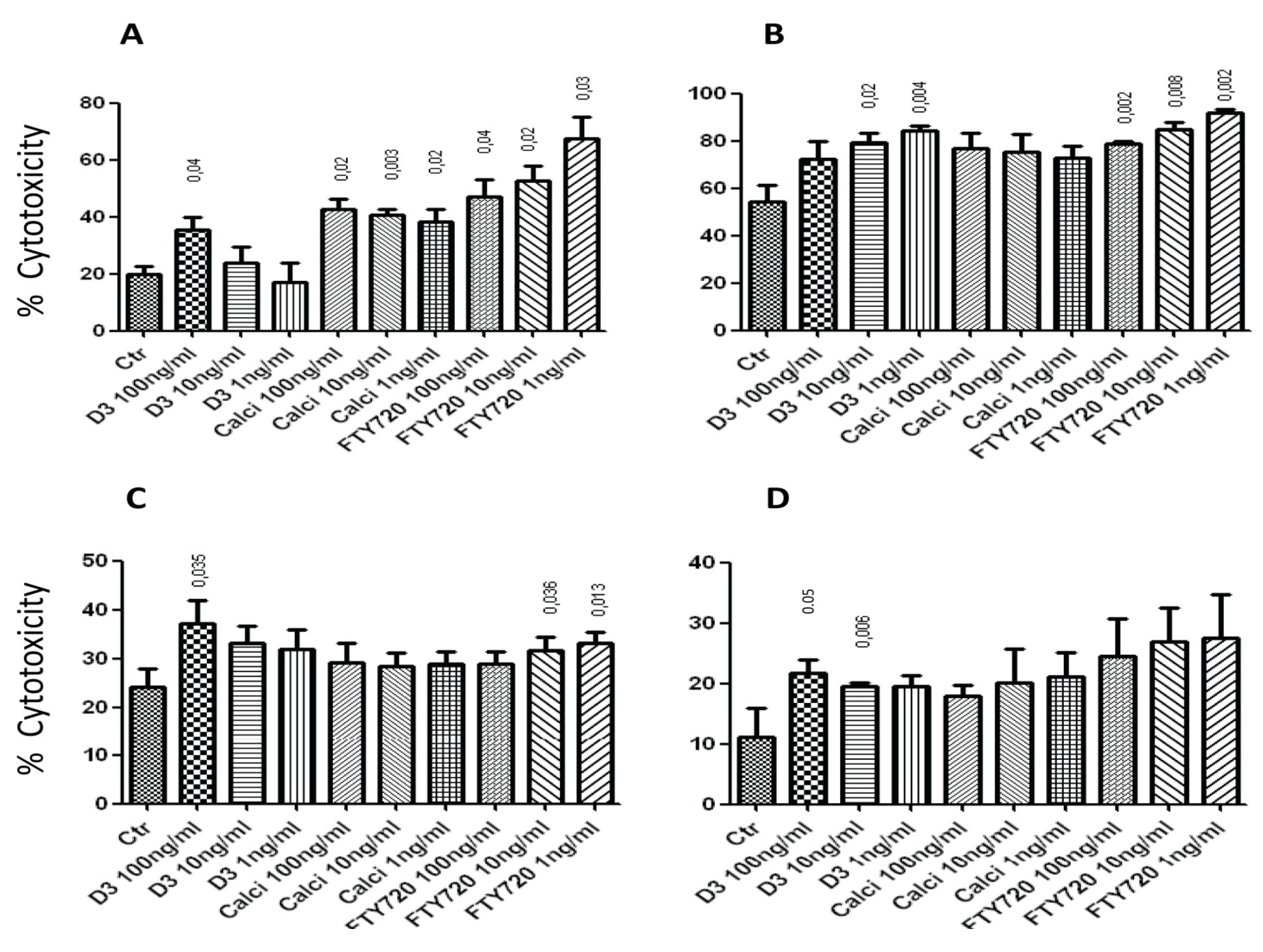
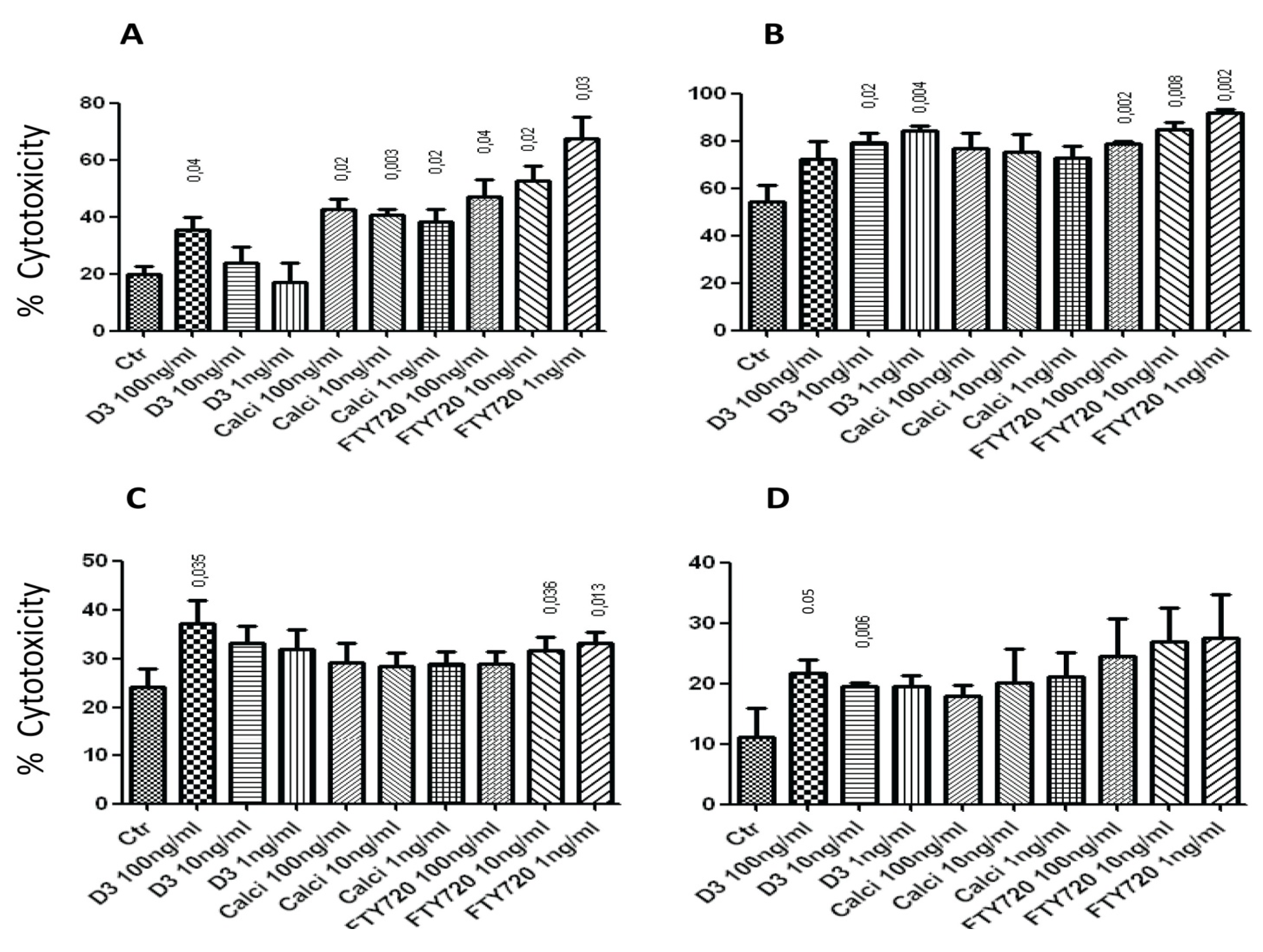
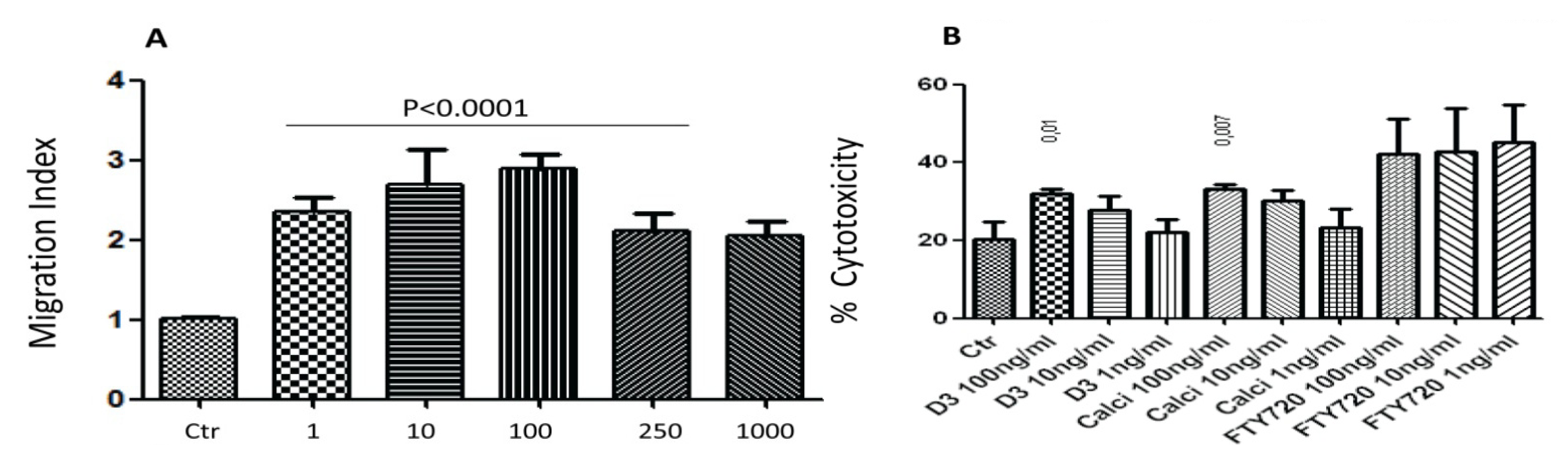
2.1. Effects of the Drugs on NK Cell Lysis of Target Cells
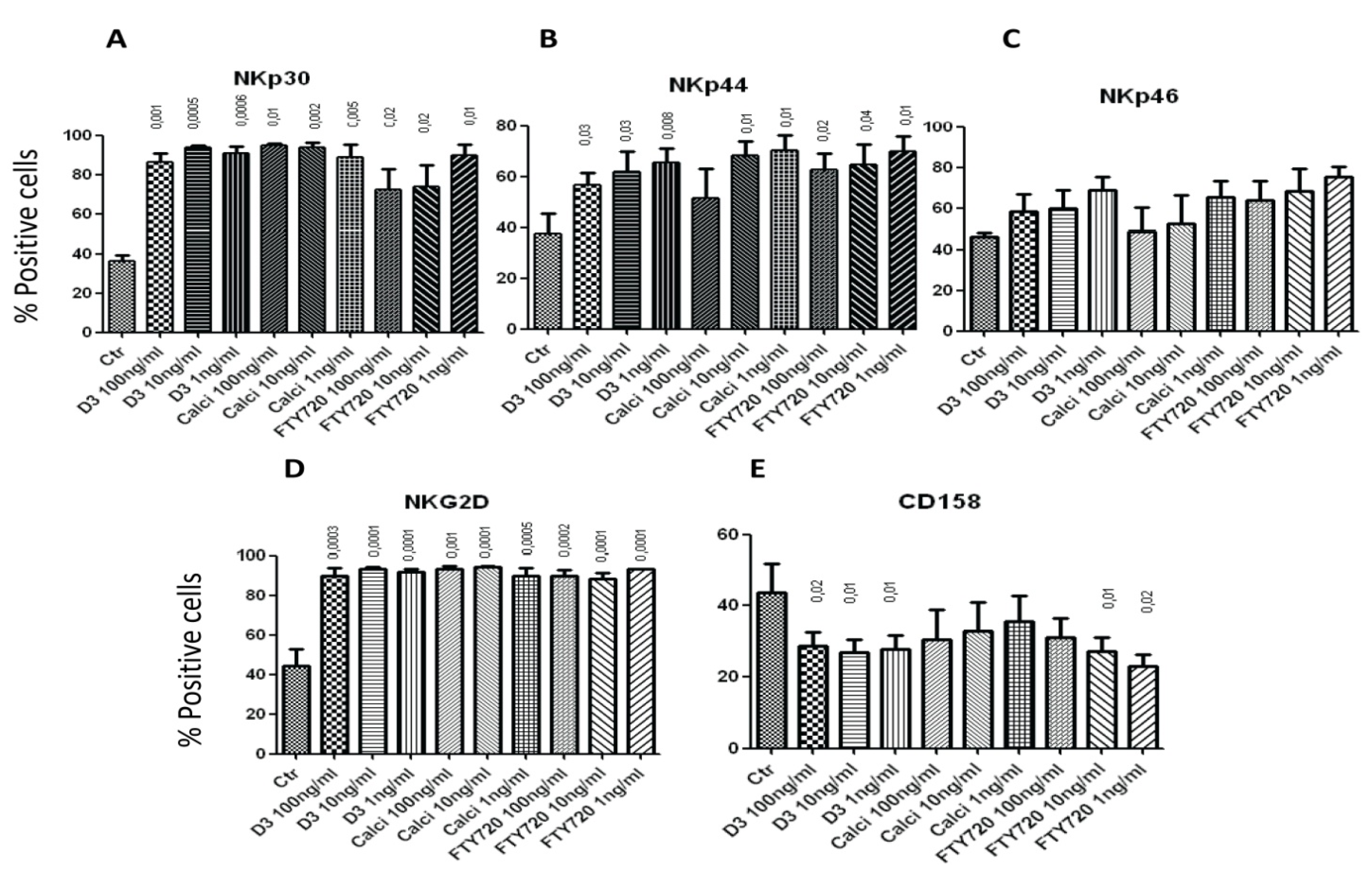
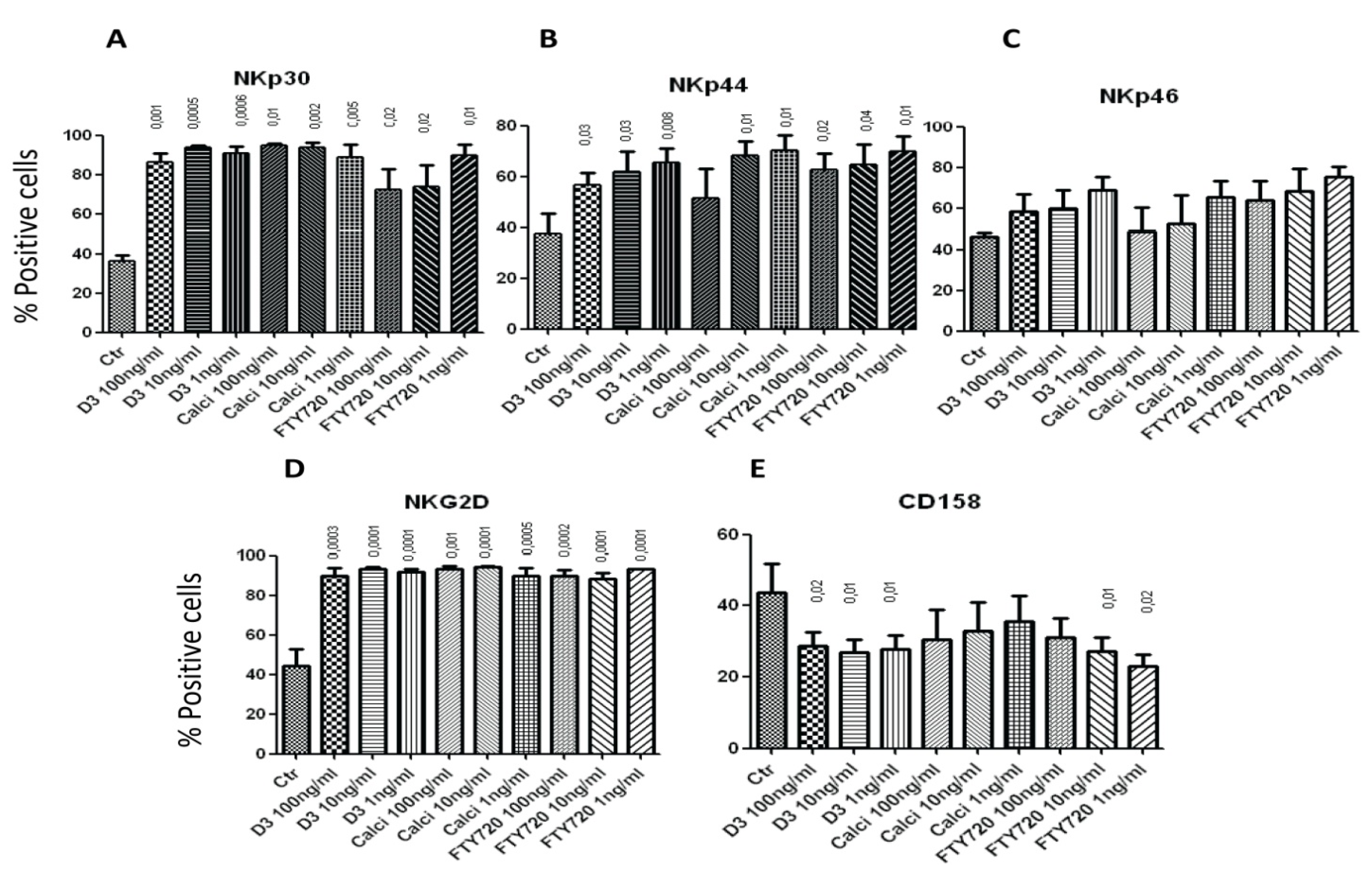
2.2. 1,25(OH)2D3, Calcipotriol and FTY720 Up-Regulate the Expression of NK Cytotoxicity Molecules on the Surface of NK Cellstitle
2.3. Effect of the Drugs on the Cytolytic Activity of NK17/NK1 Cells
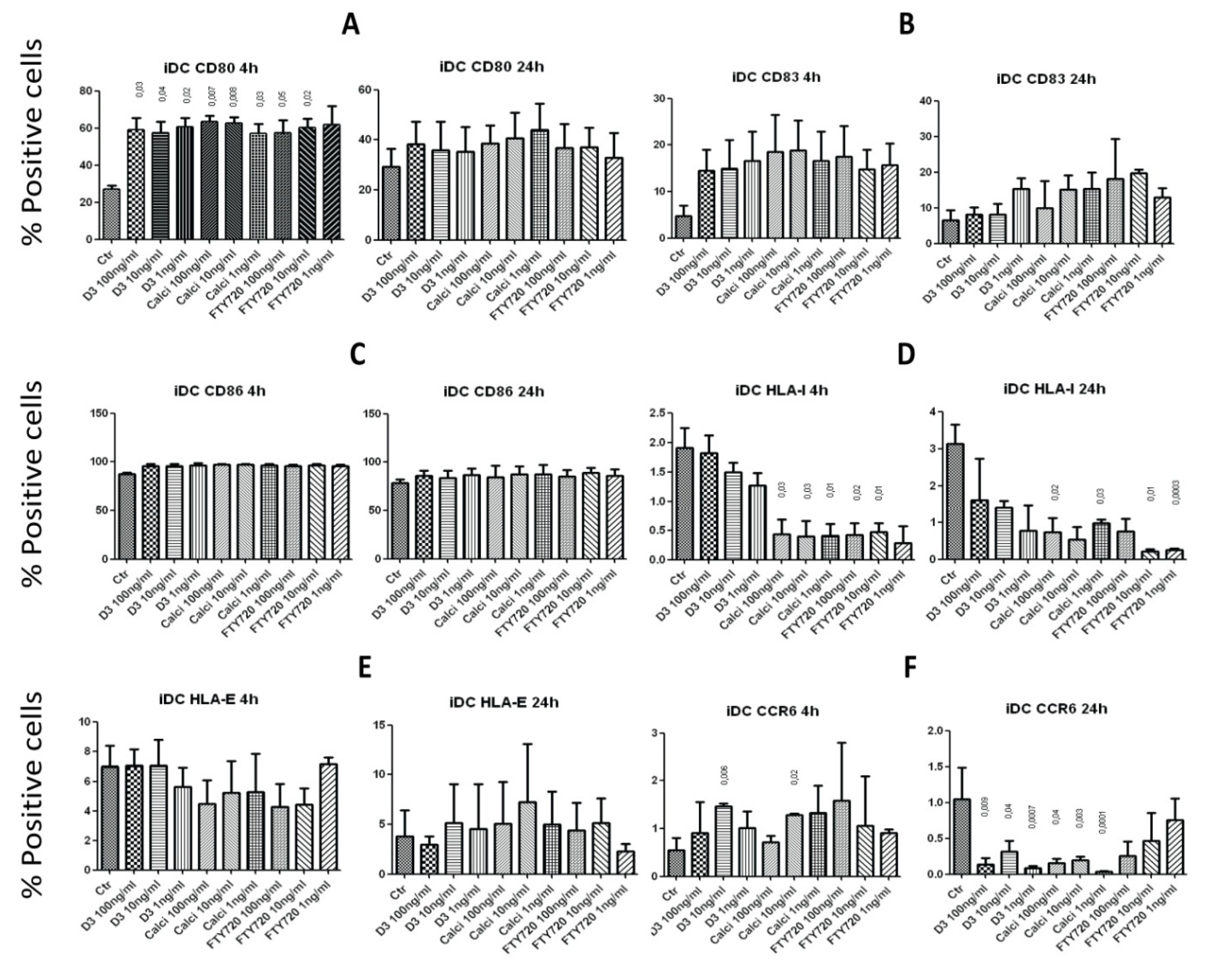
2.4. Effects of Incubating iDCs with the Drugs on the Expression of Surface Molecules
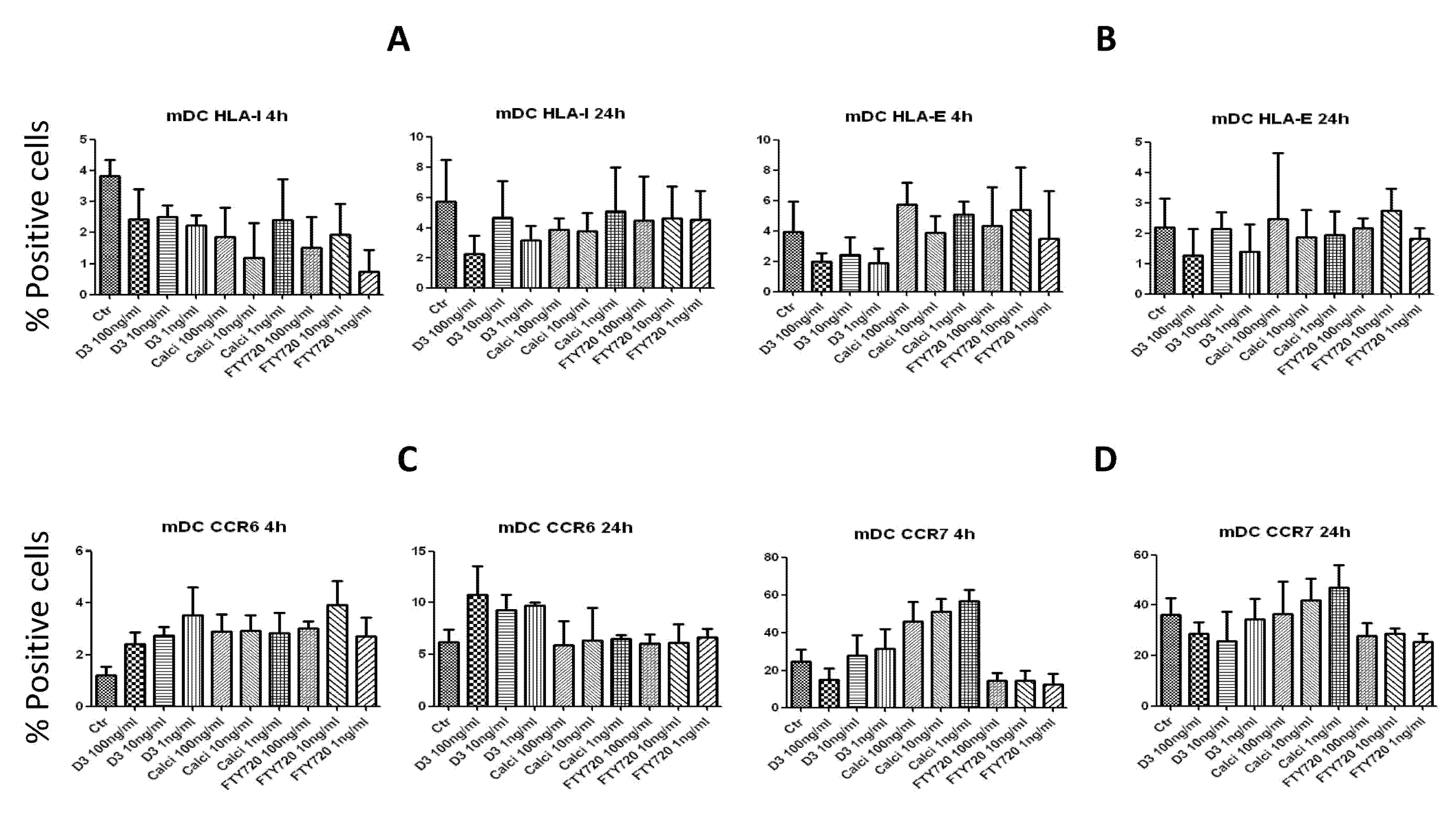
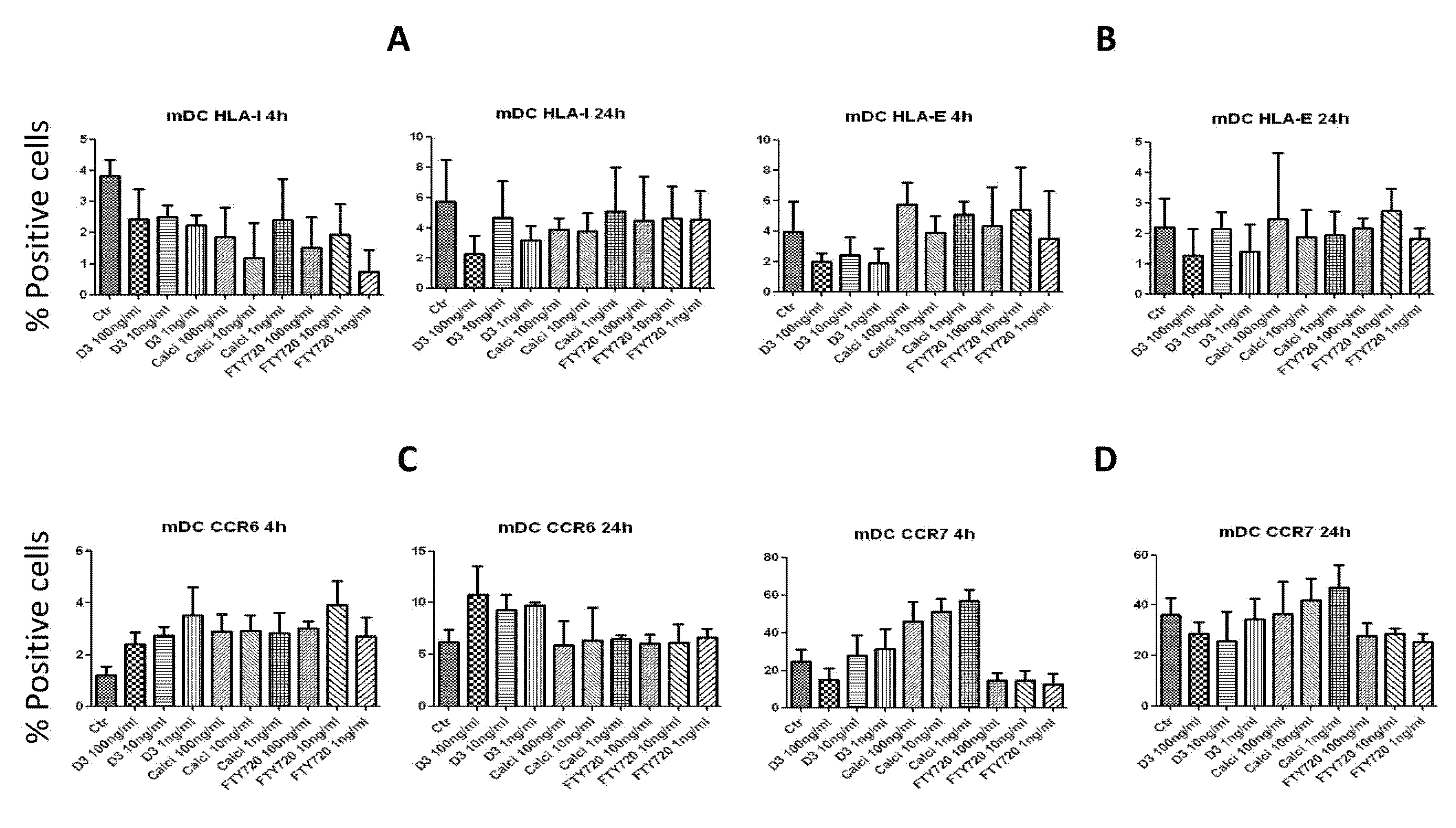
2.5. Effects on the Expression of Surface Molecules on mDCs
3. Discussion
4. Experimental Section
4.1. Cell Preparations
4.2. Isolation of NK17/NK1 Cells
4.3. Cytotoxicity Assay
4.4. Flow Cytometric Analysis
4.5. Chemotaxis Assay
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Fauriat, C.; Long, E.O.; Ljunggren, H.G.; Bryceson, Y.T. Regulation of human NK-cell cytokine and chemokie production by target cell recognition. Blood 2010, 115, 2167–2176. [Google Scholar]
- Maghazachi, A.A. Compartmentalization of human natural killer cells. Mol. Immunol. 2005, 42, 523–529. [Google Scholar]
- Wood, S.M.; Ljunggren, H.G.; Bryceson, Y.T. Insights into NK cell biology from human genetics and disease associations. Cell. Mol. Life Sci. 2011, 68, 3479–3493. [Google Scholar] [CrossRef]
- Maghazchi, A.A. On the role of natural killer cells in neurodegenerative diseases. Toxins 2013, 5, 363–375. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Role of natural killer cells in multiple sclerosis. ISRN Immunol. 2012. [Google Scholar] [CrossRef]
- Benczur, M.; Petranyl, G.G.; Palffy, G.; Varga, M.; Talas, M.; Kotsy, B.; Földes, I.; Hollán, S.R. Dysfunction of natural killer cells in multiple sclerosis: A possible pathogenetic factor. Clin. Exp. Immunol. 1980, 39, 657–662. [Google Scholar]
- Kastrukoff, L.F.; Morgan, N.G.; Aziz, T.M.; Zecchini, D.; Berkowitz, J.; Paty, D.W. Natural killer (NK) cells in chronic progressive multiple sclerosis patients treated with lymphoblastoid interferon. J. Neuroimmunol. 1998, 20, 15–23. [Google Scholar]
- Grunebaum, E.; Malatzky-Goshen, E.; Shoenfeld, Y. Natural killer cells and autoimmunity. Immunol. Res. 1989, 8, 292–304. [Google Scholar] [CrossRef]
- Zhang, B.; Yamamura, T.; Kondo, T.; Fujiwara, M.; Tabira, T. Regulation of experimental autoimmune encephalomyelitis by natural killer (NK) cells. J. Exp. Med. 1997, 186, 1677–1687. [Google Scholar] [CrossRef]
- Kaur, G.; Trowsdale, J.; Fugger, L. Natural killer cells and their receptors in multiple sclerosis. Brain 2013, 136, 2657–2676. [Google Scholar] [CrossRef]
- Winkler-Pickett, R.; Young, H.A.; Cherry, J.M.; Diehl, J.; Wine, J.; Back, T.; Bere, W.E.; Mason, A.T.; Ortaldo, J.R. In vivo regulation of experimental autoimmune encephalomyelitis by NK cells: Alteration of primary adaptive responses. J. Immunol. 2008, 180, 4495–4506. [Google Scholar]
- Vollmer, T.L.; Liu, R.; Price, M.; Rhodes, S.; La, C.A.; Shi, F.D. Differential effects of IL-21 during initiation and progression of autoimmunity against neuroantigen. J. Immunol. 2005, 174, 2696–2701. [Google Scholar]
- Sand, K.L.; Rolin, J.; Knudsen, E.; Al-Falahi, Y.; Maghazachi, A.A. Modulation of natural killer cell cytotoxicity and cytokine release by the drug glatiramer acetate. Cell. Mol. Life Sci. 2009, 66, 1446–1456. [Google Scholar] [CrossRef]
- Al-Falahi, Y.; Sand, K.L.; Knudsen, E.; Damaj, B.B.; Rolin, J.; Maghazachi, A.A. Splenic natural killer cell activity in two models of experimental neurodegenerative diseases. J. Cell. Mol. Med. 2009, 13, 2693–2703. [Google Scholar] [CrossRef]
- Høglund, R.; Harbo, H.F.; Holmøy, T.; Maghazachi, A.A. A one year follow-up study of natural killer and dendritic cells activities in multiple sclerosis patients receiving glatiramer acetate. PLoS One 2013, 8, e62237. [Google Scholar]
- Hestvick, A.L.K. The Double-edged sword of autoimmunity: Lessons from multiple sclerosis. Toxins 2010, 2, 856–877. [Google Scholar] [CrossRef]
- Ascherio, A.; Munger, K.L. Environmental risk factors for multiple sclerosis. Part II: Noninfectious factors. Ann. Neurol. 2007, 61, 504–513. [Google Scholar] [CrossRef]
- Ascherio, A.; Munger, K.L.; Simon, K.C. Vitamin D and multiple sclerosis. Lancet Neurol. 2010, 9, 599–612. [Google Scholar] [CrossRef]
- Brustad, M.; Sandanger, T.; Aksnes, L.; Lund, E. Vitamin D status in a rural population of northern Norway with high fish liver consumption. Public Health Nutr. 2004, 7, 783–789. [Google Scholar]
- Lemire, J.M.; Archer, D.C. 1,25-dihydroxyvitamin D3 prevents the in vivo induction of murine experimental autoimmune encephalomyelitis. J. Clin. Investig. 1991, 87, 1103–1107. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Hayes, C.E.; DeLuca, H.F. 1,25-Dihydroxyvitamin D3 reversibly blocks the progression of relapsing encephalomyelitis, a model of multiple sclerosis. Proc. Natl. Acad. Sci. USA 1996, 93, 7861–7864. [Google Scholar] [CrossRef]
- Burton, J.M.; Kimball, S.; Vieth, R.; Bar-Or, A.; Dosch, H.M.; Cheung, R.; Gagne, D.; D’Souza, C.; Ursell, M.; O’Connor, P. A phase I/II dose-escalation trial of vitamin D3 and calcium in multiple sclerosis. Neurology 2010, 74, 1852–1859. [Google Scholar] [CrossRef]
- Smolders, J.; Thewissen, M.; Peelen, E.; Menheere, P.; Tervaert, J.W.; Damoiseaux, J.; Hupperts, R. Vitamin D status is positively correlated with regulatory T cell function in patients with multiple sclerosis. PLoS One 2009, 4, e6635. [Google Scholar]
- Smolders, J.; Peelen, E.; Thewissen, M.; Tervaert, J.W.; Menheere, P.; Hupperts, R.; Damoiseaux, J. Safety and T cell modulating effects of high hose vitamin D3 supplementation in multiple sclerosis. PLoS One 2010, 5, e15235. [Google Scholar]
- Wergeland, S.; Torkildsen, Ø.; Myhr, K.M.; Aksnes, L.; Mørk, S.J.; Bø, L. Dietary vitamin D3 supplements reduce demyelination in the cuprizone model. PLoS One 2011, 6, e26262. [Google Scholar]
- Brinkmann, V. Sphingosine 1-phosphate receptors in health and disease: Mechanistic insights from gene deletion studies and reverse pharmacology. Pharmacol. Ther. 2007, 115, 84–105. [Google Scholar] [CrossRef]
- Kveberg, L.; Bryceson, Y.; Inngjerdingen, M.; Rolstad, B.; Maghazachi, A.A. Sphingosine 1 phosphate induces the chemotaxis of human natural killer cells. Role for heterotrimeric G proteins and phosphoinositide 3 kinases. Eur. J. Immunol. 2002, 32, 1856–1864. [Google Scholar] [CrossRef]
- Lagadari, M.; Lehmann, K.; Ziemer, M.; Truta-Feles, K.; Berod, L.; Idzko, M.; Barz, D.; Kamradt, T.; Maghazachi, A.A.; Norgauer, J. Sphingosine-1-phosphate inhibits the cytotoxic activity of NK cells via Gs protein-mediated signaling. Int. J. Oncol. 2009, 34, 287–288. [Google Scholar]
- Rolin, J.; Sand, K.L.; Knudsen, E.; Maghazachi, A.A. FTY720 and SEW2871 reverse the inhibitory effect of S1P on natural killer cell mediated lysis of K562 tumor cells and dendritic cells but not on cytokine release. Cancer Immunol. Immnuother. 2010, 59, 575–586. [Google Scholar] [CrossRef]
- Pandya, A.D.; Al-Jaderi, Z.; Høglund, R.A.; Holmøy, T.; Harbo, H.F.; Norgauer, J.; Maghazachi, A.A. Identification of human NK17/NK1 cells. PLoS One 2011, 6, e26780. [Google Scholar]
- Godiska, R.; Chantry, D.; Raport, C.J.; Sozzani, S.; Allavena, P.; Leviten, D.; Mantovani, A.; Gray, P.W. Human macrophage-derived chemokine (MDC), a novel chemoattractant for monocytes, monocyte-derived dendritic cells, and natural killer cells. J. Exp. Med. 1997, 185, 1595–1604. [Google Scholar]
- Székely, J.I.; Pataki, Á. Effects of vitamin D on immune disorders with special regard to asthma, COPD and autoimmune diseases: A short review. Experts Rev. 2012, 6, 683–704. [Google Scholar]
- Penna, G.; Amuchastegui, S.; Laverny, G.; Adorini, L. Vitamin D receptor agonists in the treatment of autoimmune diseases: Selective targeting of myeloid but not plasmacytoid dendritic cells. J. Bone Miner. Res. 2007, 22, V69–V73. [Google Scholar] [CrossRef]
- Motrich, R.D.; van Etten, E.; Depovere, J.; Riera, C.M.; Rivero, V.E.; Mathieu, C. Impact of vitamin D receptor activity on experimental autoimmune prostatitis. J. Autoimmun. 2009, 32, 140–148. [Google Scholar]
- Gauzzi, M.C.; Purificato, C.; Donato, K.; Jin, Y.; Wang, L.; Daniel, K.C.; Maghazachi, A.A.; Belardelli, F.; Adorini, L.; Gessani, S. Suppressive effect of 1alpha,25-dihydroxyvitamin D3 on type I IFN-mediated monocyte differentiation into dendritic cells: Impairment of functional activities and chemotaxis. J. Immunol. 2005, 174, 270–276. [Google Scholar]
- Lee, J.H.; Park, S.; Cheon, S.; Lee, J.H.; Kim, S.; Hur, D.Y.; Kim, T.S.; Yoon, S.R.; Yang, Y.; Bang, S.I.; et al. 1,25-Dihydroxyvitamin D3 enhances NK susceptibility of human melanoma cells via Hsp60-mediated FAS expression. Eur. J. Immunol. 2011, 41, 2937–2946. [Google Scholar] [CrossRef]
- El-Shazly, A.E.; Lefebvre, P.P. Modulation of NK cell autocrine-induced eosinophil chemotaxis by interleukin-15 and vitamin D3: A possible NK-eosinophil crosstalk via IL-8 in the pathophysiology of allergic rhinitis. Mediat. Inflamm. 2011. [Google Scholar] [CrossRef]
- Brandt, C.S.; Baratin, M.; Yi, E.C.; Kennedy, J.; Gao, Z.; Fox, B.; Haldeman, B.; Ostrander, C.D.; Kaifu, T.; Chabannon, C.; et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J. Exp. Med. 2009, 206, 1495–1503. [Google Scholar] [CrossRef]
- Simhadri, V.R.; Reiners, K.S.; Hansen, H.P.; Topolar, D.; Simhadri, V.L.; Nohroudi, K.; Kufer, T.A.; Engert, A.; Pogge von Strandmann, E. Dendritic cells release HLA-B-associated transcript-3 positive exosomes to regulate natural killer function. PLoS One 2008, 3, e3377. [Google Scholar]
- Marras, F.; Nicco, E.; Bozzano, F.; Di Biagio, A.; Dentone, C.; Pontali, E.; Boni, S.; Setti, M.; Orofino, G.; Mantia, E.; et al. Natural killer cells in HIV controller patients express an activated effector phenotype and do not up-regulate NKp44 on IL-2 stimulation. Proc. Natl. Acad. Sci. USA 2013, 110, 11970–11975. [Google Scholar] [CrossRef]
- De Maria, A.; Biassoni, R.; Fogli, M.; Rizzi, M.; Cantoni, C.; Costa, P.; Conte, R.; Mavilio, D.; Ensoli, B.; Cafaro, A.; et al. Indentification, molecular cloning and functional characterization of NKp46 and NKp30 natural cytotoxicity receptors in Macaca fascicularis NK cells. Eur. J. Immunol. 2001, 31, 3546–3556. [Google Scholar] [CrossRef]
- Arnon, T.I.; Achdout, H.; Lieberman, N.; Gazit, R.; Gonen-Gross, T.; Katz, G.; Bar-Ilan, A.; Bloushtain, N.; Lev, M.; Joseph, A.; et al. The mechanisms controlling the recognition of tumor- and virus-infected cells by NKp46. Blood 2004, 103, 664–672. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Insights into seven and single transmembrane-spanning domain receptors and their signaling pathways in human natural killer cells. Pharmacol. Rev. 2005, 57, 339–357. [Google Scholar] [CrossRef]
- Raϊch-Regué, D.; Grau-López, L.; Naranjo-Gómez, M.; Ramo-Tello, C.; Pujol-Borrell, R.; Martínez-Cáceres, E.; Borràs, F.E. Stable antigen-specific T-cell hyporesponsiveness induced by tolerogenic dendritic cells from multiple sclerosis patients. Eur. J. Immunol. 2012, 42, 771–782. [Google Scholar] [CrossRef]
- Kappos, L.; Antel, J.; Comi, G.; Montalban, X.; O’Connor, P.; Polman, C.H.; Haas, T.; Korn, A.A.; Karlsson, G.; Radue, E.W. Oral fingolimod (FTY720) for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 355, 1124–1140. [Google Scholar] [CrossRef]
- O’Connor, P.; Comi, G.; Montalban, X.; Antel, J.; Radue, E.W.; de Vera, A.; Pohlmann, H.; Kappos, L. FTY720 D2201 Study Group: Oral fingolimod (FTY720) in multiple sclerosis: Two-year results of a phase II extension study. Neurology 2009, 72, 73–79. [Google Scholar] [CrossRef]
- Pyne, S.; Pyne, N.J. New perspectives on the role of sphingosine 1-phosphate in cancer. Handb. Exp. Pharmacol. 2013, 216, 55–71. [Google Scholar]
- Jin, Y.; Knudsen, E.; Wang, L.; Bryceson, Y.; Damaj, B.; Gessani, S.; Maghazachi, A.A. Sphingosine 1-phosphate is a novel inhibitor of T-cell proliferation. Blood 2003, 101, 4909–4915. [Google Scholar] [CrossRef] [Green Version]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Al-Jaderi, Z.; Maghazachi, A.A. Effects of Vitamin D3, Calcipotriol and FTY720 on the Expression of Surface Molecules and Cytolytic Activities of Human Natural Killer Cells and Dendritic Cells. Toxins 2013, 5, 1932-1947. https://doi.org/10.3390/toxins5111932
Al-Jaderi Z, Maghazachi AA. Effects of Vitamin D3, Calcipotriol and FTY720 on the Expression of Surface Molecules and Cytolytic Activities of Human Natural Killer Cells and Dendritic Cells. Toxins. 2013; 5(11):1932-1947. https://doi.org/10.3390/toxins5111932
Chicago/Turabian StyleAl-Jaderi, Zaidoon, and Azzam A. Maghazachi. 2013. "Effects of Vitamin D3, Calcipotriol and FTY720 on the Expression of Surface Molecules and Cytolytic Activities of Human Natural Killer Cells and Dendritic Cells" Toxins 5, no. 11: 1932-1947. https://doi.org/10.3390/toxins5111932