3.2. PA Kinetics in Untreated Rabbits
The mean (with SD error bars) observed plasma PA concentration-time profiles for the untreated animals included in the analyses are illustrated in
Figure 1. The profiles for individual animals generally followed the pattern of initial lag phase, a rising phase, a plateau, and then another rising phase, a pattern that has been reported previously in this animal model [
17]. The exceptions were that some animals died prior to attaining either the plateau phase or the second rising phase. A composite modified Gompertz/second exponential growth model, consisting of the Gompertz model with a second exponential growth phase following a second lag phase, was consistent with the observed data.
Figure 1.
Mean (SD error bars) plasma PA concentration-time profiles in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge. For the purposes of this figure, the terminal specimen results are plotted at the 84h post-challenge time, without regard to the actual collection times for those specimens. All untreated rabbits died prior to the 84h-post challenge time.
Figure 1.
Mean (SD error bars) plasma PA concentration-time profiles in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge. For the purposes of this figure, the terminal specimen results are plotted at the 84h post-challenge time, without regard to the actual collection times for those specimens. All untreated rabbits died prior to the 84h-post challenge time.
As shown in
Figure 1, there was variability in the concentrations for the terminal specimens. Due to the large variability of the terminal specimen results, and uncertainty concerning the contribution of post-mortem changes to that variability, the terminal specimen results were excluded from the PA kinetic modeling.
Modeling of inter-individual variability for all of the model parameters was attempted, but was not successful. In the final model, inter-individual variability could not be estimated for PA concentration at time 0 (N0), lag time for the first growth phase (λ), maximum specific growth rate for the first phase (μm), or lag time for the second growth phase (λ2). Given the complexity of this model, the substantial variability in the data, and the varying contribution of each animal to the model, inability to adequately model inter-individual variability for all parameters in the model could be expected. Rather than revealing any deficiency in the model, this outcome may be a qualitative reflection of the data being modeled. Since the purpose of this modeling exercise was to provide a description of the observed data, the modeling results can be considered adequate for the intended purpose, but should be interpreted with caution due to the poor definition of the variability components.
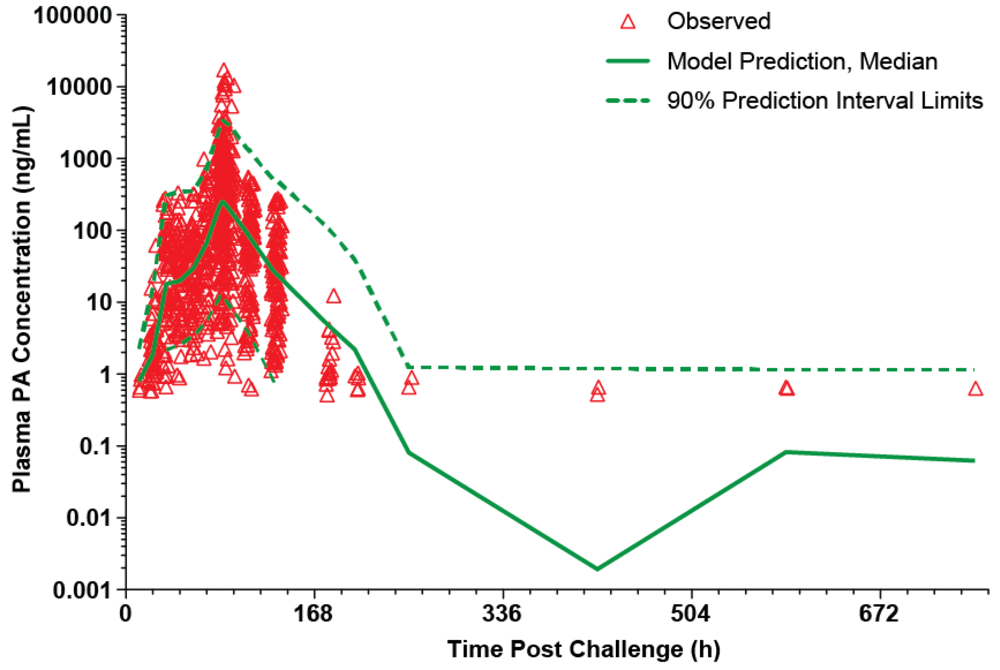
The model was evaluated using a visual predictive check. As shown in
Figure 2, the majority of the observed plasma PA concentrations are within the 90% prediction interval, suggesting that the model describes the data well. The parameter estimates for the model are summarized in
Table 2.
Figure 2.
Visual predictive check for PA kinetic model with individual observed plasma PA concentrations in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge. The lower bound of the 90% prediction interval is zero at all post-challenge times, except 24 and 72h, which are marked as green squares. A composite modified Gompertz/second exponential growth phase following a second lag phase model was used for nonlinear mixed effects modeling (see Materials and Methods section), while the median prediction and prediction interval were generated by running the final model in simulation mode, generating 200 replicate simulations for each rabbit.
Figure 2.
Visual predictive check for PA kinetic model with individual observed plasma PA concentrations in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge. The lower bound of the 90% prediction interval is zero at all post-challenge times, except 24 and 72h, which are marked as green squares. A composite modified Gompertz/second exponential growth phase following a second lag phase model was used for nonlinear mixed effects modeling (see Materials and Methods section), while the median prediction and prediction interval were generated by running the final model in simulation mode, generating 200 replicate simulations for each rabbit.
Table 2.
PA kinetic parameters in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, based on fitting of a composite modified Gompertz/second exponential growth phase following a second lag phase model (see Materials and Methods section) to the data.
Table 2.
PA kinetic parameters in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, based on fitting of a composite modified Gompertz/second exponential growth phase following a second lag phase model (see Materials and Methods section) to the data.
| Parameters | Mean 1 | CV% 1 |
|---|
| N0 (ng/mL) | 0.38 (20.5%) | - 2 |
| Association of TBAC with N0 3 | N0 × (TBAC/34)−3.55 (28.2%) |
| At 10.57 h | 24.08 |
| At 20 h | 2.51 |
| At 30 h | 0.59 |
| At 34 h | 0.38 |
| At 40 h | 0.21 |
| At 44.72 h | 0.14 |
| λ (h) | 20 (2.7%) | - 2 |
| μm (h−1) | 0.86 (9.8%) | - 2 |
| A (unitless) | 5.08 (5.1%) | 24.5 (29.9%) |
| λ2 (h) | 22 (0.1%) | - 2 |
| μm,2 (h−1) | 0.16 (3.4%) | 30.4 (21.1%) |
| Residual variability (CV%) | 61.2 (7.4%) |
As shown in
Table 2, lower
N0 values were associated with longer TBAC values. That association is intuitive since higher values of
N0 would be expected to reflect earlier and more abundant spore germination and vegetative growth, leading to earlier detection of bacteremia. No other associations between PA kinetics and the covariates assessed were found.
Possible relationships between survival time and the natural log of the ratio of the PA concentration in the asymptotic (plateau) phase to
N0 (A) or the maximum specific growth rate for the second phase (μ
m,2) were assessed (
Figure 3). Survival times tended to become shorter as A increased, with A accounting for 36% of the variability in the survival time data. This is intuitive since higher plateau PA concentrations would be expected to be associated with increased morbidity. Survival time also tended to decrease as μ
m,2 increased, with μ
m,2 explaining 34% of the variability in survival times. Since higher values of μ
m,2 represent more rapid rates of increase in PA concentrations during the second phase of the profile, this relationship could be expected. Since the relationships between survival time and magnitude of PA levels or rate of increase of PA levels are not entirely predictive, it is likely that survival time was partially dependent on other factors.
Figure 3.
Relationships between survival time and natural log of the ratio of the PA concentration in the asymptotic (plateau) phase to N0 (A) and maximum specific growth rate for the second phase (B) in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge. Data points represent the post hoc PA kinetic parameter estimates for individual rabbits obtained from fitting the observed data to a composite modified Gompertz/second exponential growth phase following a second lag phase model (see Materials and Methods section).
Figure 3.
Relationships between survival time and natural log of the ratio of the PA concentration in the asymptotic (plateau) phase to N0 (A) and maximum specific growth rate for the second phase (B) in untreated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge. Data points represent the post hoc PA kinetic parameter estimates for individual rabbits obtained from fitting the observed data to a composite modified Gompertz/second exponential growth phase following a second lag phase model (see Materials and Methods section).
3.3. PA Kinetics in Treated Rabbits
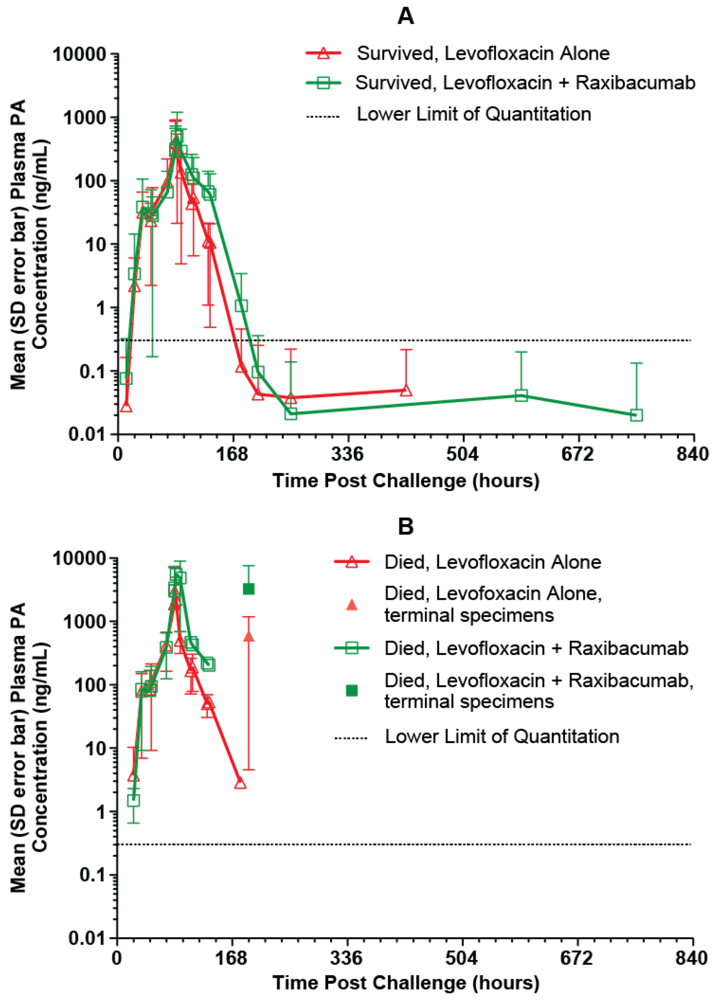
The mean (with SD error bars) observed plasma PA concentration-time profiles for the treated animals are illustrated in
Figure 4. For rabbits that survived, the mean plasma PA concentration-time profile had an initial rising phase, and after the treatment time, when bacteremia had been eradicated (data not shown), plasma PA concentrations declined. This type of PA profile was characteristic of all surviving animals. The plasma PA concentration-time profiles for treated rabbits that died were more varied. Some treated rabbits that died had plasma PA concentration-time profiles that generally exhibited increasing concentrations (4 of 20 rabbits [20%]), while others had profiles in which plasma PA concentrations were decreasing at the time of death (16 of 20 rabbits [80%]). It is possible that the rabbits whose profiles exhibited only increasing concentrations died before clearance of PA became evident. For the rabbits that died during the period when plasma PA levels were decreasing, it is likely that death occurred as a result of cumulative organ damage from the toxin or host specific response to the bacterium, despite the evidence that clearance of PA was ongoing at the time of death.
Figure 4.
Mean (SD error bars) plasma PA concentration-time profiles in rabbits that survived (A) and rabbits that died (B) after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose. For the purposes of this figure, the terminal specimen results are plotted at the 192h post challenge time, without regard to the actual collection times for those specimens.
Figure 4.
Mean (SD error bars) plasma PA concentration-time profiles in rabbits that survived (A) and rabbits that died (B) after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose. For the purposes of this figure, the terminal specimen results are plotted at the 192h post challenge time, without regard to the actual collection times for those specimens.
The data for treated animals were consistent with the composite modified Gompertz/second exponential growth model with a monoexponential elimination phase. Alternate parameterizations using clearance and volume of distribution terms and/or a delay time between treatment and the start of PA elimination were unsuccessful due to apparent over-parameterization of the model. In particular, the estimate for the delay time was very small, implying that the data did not support estimation of that parameter. The final model was parameterized with a first-order elimination rate constant (kelm) and no delay time.
Modeling of inter-individual variability for all of the model parameters was attempted, but was not successful. In the final model, inter-individual variability could not be estimated for λ, μm, N0, μm,2, and kelm. Given the complexity of this model and the substantial variability in the data, inability to adequately model inter-individual variability for all parameters in the model could be expected. Rather than revealing any deficiency in the model, this outcome may be a qualitative reflection of the data being modeled. Since the purpose of this modeling exercise was to provide a description of the observed data, the modeling results can be considered adequate for the intended purpose, but should be interpreted with caution due to the poor definition of the variability components.
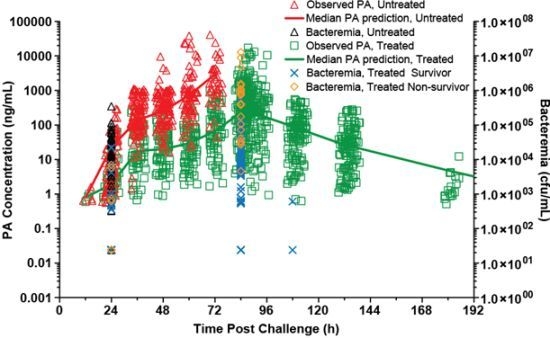
The model for the treated rabbits was evaluated using a visual predictive check. As shown in
Figure 5, the majority of the observed plasma PA concentrations are within the 90% prediction interval, suggesting that the model describes the data well. The appearance of the predicted profile at times greater than 336 h post-challenge is driven by the relatively few rabbits that had measurable PA concentrations at those times. The parameter estimates for the model are summarized in
Table 3.
Possible differences in PA kinetics for treated rabbits that died and survived were assessed (
Table 4). The 95% confidence interval (CI) of the
post hoc PA kinetic parameter estimates for the treated rabbits that survived and those that died generally overlapped, indicating no statistically significant differences, with the exceptions of A and λ
2. For the treated rabbits that died, plateau PA levels (A) were significantly higher (33%) than those for rabbits that survived, and duration of the plateau phase was significantly shorter (29%) than for rabbits that survived. These findings are consistent with the hypothesis that death due to toxemia tends to be associated with PA levels that are higher, and progress to higher levels earlier.
As shown in
Table 3, some variability in λ was associated with both sex (SEX) and treatment group (GRP), while some variability in μ
m was associated with sex, and some variability in k
elm was associated with treatment group. No other associations between PA kinetics and the covariates assessed were found. For females, λ was about 80% of that for males, and μ
m was only about 40% of that in males. Despite these differences, there was not a difference in survival rates between males and females, suggesting that the differences were not clinically meaningful. For rabbits, administered levofloxacin plus raxibacumab, λ was 7% longer than that for rabbits administered levofloxacin alone. This small difference would not be expected to be clinically meaningful. PA elimination was 45% slower in rabbits administered levofloxacin with raxibacumab than in rabbits administered levofloxacin alone. This may reflect systemic persistence of measurable PA resulting from the PA-raxibacumab complex. However, λ and k
elm were not significantly different between the survivor and non-survivor subgroups (
Table 4), implying that the effects of the treatment group on those parameters were not clinically meaningful.
Figure 5.
Visual predictive check for PA kinetic model with individual observed plasma PA concentrations in treated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose. The lower bound of the 90% prediction interval is zero at post-challenge times greater than 168h. A composite modified Gompertz/second exponential growth phase following a second lag phase/monoexponential elimination phase was used for nonlinear mixed effects modeling (see Materials and Methods section), while the median prediction and prediction interval were generated by running the final model in simulation mode, generating 200 replicate simulations for each rabbit.
Figure 5.
Visual predictive check for PA kinetic model with individual observed plasma PA concentrations in treated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose. The lower bound of the 90% prediction interval is zero at post-challenge times greater than 168h. A composite modified Gompertz/second exponential growth phase following a second lag phase/monoexponential elimination phase was used for nonlinear mixed effects modeling (see Materials and Methods section), while the median prediction and prediction interval were generated by running the final model in simulation mode, generating 200 replicate simulations for each rabbit.
Table 3.
PA kinetic parameters in treated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose, based on the fitting of a composite modified Gompertz/second exponential growth phase following a second lag phase/monoexponential elimination phase model (see Materials and Methods section) to the data.
Table 3.
PA kinetic parameters in treated rabbits after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose, based on the fitting of a composite modified Gompertz/second exponential growth phase following a second lag phase/monoexponential elimination phase model (see Materials and Methods section) to the data.
| Parameters | Mean 1 | CV% 1 |
|---|
| N0
(ng/mL) | 0.83 (11.9%) | - 2 |
| λ (h) | 24 (1.2%) | - 2 |
| Effect of treatment group on λ (h)3 | λ × 1.07GRP (1.1%) |
| | Levofloxacin alone | 24 |
| | Levofloxacin plus raxibacumab | 26 |
| Effect of sex on λ (h) 4 | λ × 0.797SEX (4.2%) |
| | Male | 24 |
| | Female | 19 |
| μm (h−1) | 1.55 (16.7%) | - 2 |
| Effect of sex on μm (h−1) 4 | μm× 0.409SEX (29.6%) |
| | Male | 1.55 |
| | Female | 0.63 |
| A (unitless) | 3.16 (6.4%) | 39.0 (16.8%) |
| λ2 (h) | 50 (4.9%) | 24.1 (20.6%) |
| μm,2 (h−1) | 0.16 (5.8%) | - 2 |
| kelm (h−1) | 0.067 (3.5%) | - 2 |
| Effect of treatment group on kelm (h−1) 3 | kelm× 0.547GRP (7.1%) |
| | Levofloxacin alone | 0.067 |
| | Levofloxacin plus raxibacumab | 0.037 |
| Residual variability (CV%) | 46.2% (7.5%) CV% for proportional error component |
| 76.7 ng/mL (34.2%) SD for additive error component |
Table 4.
Summary of post hoc PA kinetic parameter estimates for treated rabbits that survived and died after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose. Mean and 95% CI of the post hoc PA kinetic parameter estimates for individual rabbits in each subgroup are displayed, based on fitting the observed data to a composite modified Gompertz/second exponential growth phase following a second lag phase/monoexponential elimination phase model (see Materials and Methods section).
Table 4.
Summary of post hoc PA kinetic parameter estimates for treated rabbits that survived and died after a targeted 200 × LD50 (Ames strain) inhaled B. anthracis spore challenge, followed by treatment with IG 50mg/kg qd×3 levofloxacin doses with a single intravenous placebo or 40mg/kg raxibacumab dose after the first levofloxacin dose. Mean and 95% CI of the post hoc PA kinetic parameter estimates for individual rabbits in each subgroup are displayed, based on fitting the observed data to a composite modified Gompertz/second exponential growth phase following a second lag phase/monoexponential elimination phase model (see Materials and Methods section).
| | λ | μm | A | N0 | λ2 | μm,2 | kelm |
|---|
| (h) | (h−1) | Unitless | (ng/mL) | (h) | (h−1) | (h−1) |
|---|
| Rabbits That Died |
| N | 20 | 20 | 20 | 20 | 20 | 20 | 20 |
| Mean | 22 | 1.09 | 4.14 | 0.83 | 39 | 0.16 | 0.057 |
| 95% CI | (21, 23) | (0.87, 1.31) | (3.60, 4.68) | (0.83, 0.83) | (36, 42) | (0.16,0.16) | (0.050, 0.063) |
| Rabbits That Survived |
| N | 56 | 56 | 56 | 56 | 56 | 56 | 56 |
| Mean | 22 | 1.06 | 3.12 | 0.83 | 55 | 0.16 | 0.050 |
| 95% CI | (22, 23) | (0.94, 1.18) | (2.83, 3.40) | (0.83, 0.83) | (52, 58) | (0.16, 0.16) | (0.046, 0.054) |
Based on the kelm of PA, the half-lives of PA elimination (t1/2.elm) in this study were 10 and 19 h for rabbits administered levofloxacin alone and levofloxacin with raxibacumab, respectively, with the difference likely reflecting persistence of the PA-raxibacumab complex in circulation. That is, when raxibacumab is not present to bind PA, the PA t1/2.elm is 10 h. However, when raxibacumab is present (and raxibacumab has a half-life of ~19 days), PA is bound in PA-raxibacumab complexes. Although the t1/2.elm of the complex has not been measured, it is very likely extremely long (many days) relative to that for PA alone (less than 0.5 day). In essence, the PA-raxibacumab complex probably acts as a sink which slows PA elimination. The formation of the complex also clears plasma of the toxin, leading to efficacy.
There was similarity in PA kinetics between the untreated rabbits (
i.e., those that died prior to the 84 h post-spore challenge treatment time;
Table 2) and the treated rabbits (
Table 3), with the exception of λ
2, which essentially represents the duration of the plateau phase of the serum PA profile. This suggests that although an inhaled
B. anthracis spore challenge can be lethal, survival to the 84 h post-challenge treatment time was dependent on the duration of the plateau phase, rather than other PA kinetics. Since λ
2 was not associated with the covariates assessed in this study, survival to the 84 h post-challenge treatment time was likely dependent on other factors. Most important, for those animals that were treated in the late stages of toxemia, both levofloxacin alone and levofloxacin plus raxibacumab were highly effective in attaining high survival rates.
Overall, this study provides a detailed examination of PA kinetics in a large number of untreated rabbits, as well as in rabbits treated with antibiotic alone or antibiotic plus the antitoxin raxibacumab, when treatment was initiated late in the course of anthrax disease. Despite the limitations of predicting survival time, the assessment of PA kinetics in untreated rabbits does reveal that death is associated with the highest plasma PA concentration attained for an animal, and that survival time is a function of how rapidly PA levels increase and how high the levels become, although survival time is influenced by other factors not identified in this study. Relative to this, the PA kinetics observed in this study were consistent with those observed in a smaller prior study of untreated rabbits [
17]. The PA kinetics model for treated rabbits extends the model for untreated rabbits to account for the disappearance of systemic PA that occurs once bacteremia has been eradicated. These models may be useful for design of studies to further probe the factors that influence survival time or responses to alternate treatment regimens.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}