Interaction between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and in Vitro Neutralizing Abilities

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Toxins, Chemicals, Reagents, Antibodies and Supplies
2.2. Bacterial Isolates
2.3. Stx1 and Stx2 Toxins and Toxoids
2.4. Anti-Stx1 and Anti-Stx2 Monoclonal Antibody (MAb) Production
2.5. MAb Characterization
2.5.1. MAb Isotyping and Purification
2.5.2. Interaction of MAbs with Toxin: Definition of Detection Limit, Affinity and Stability
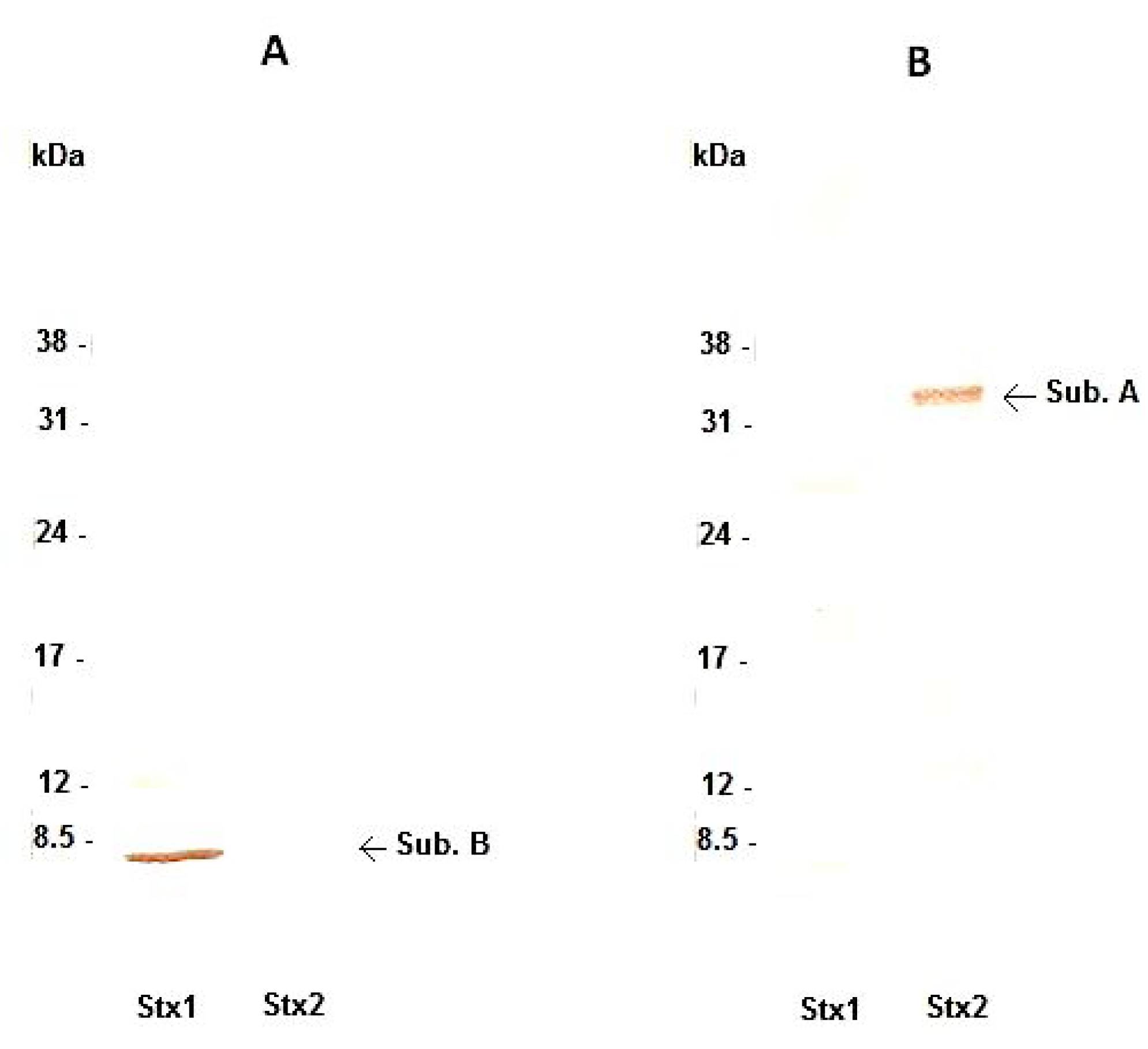
2.5.3. Stx1 and Stx2 Characterization by Immunoblotting
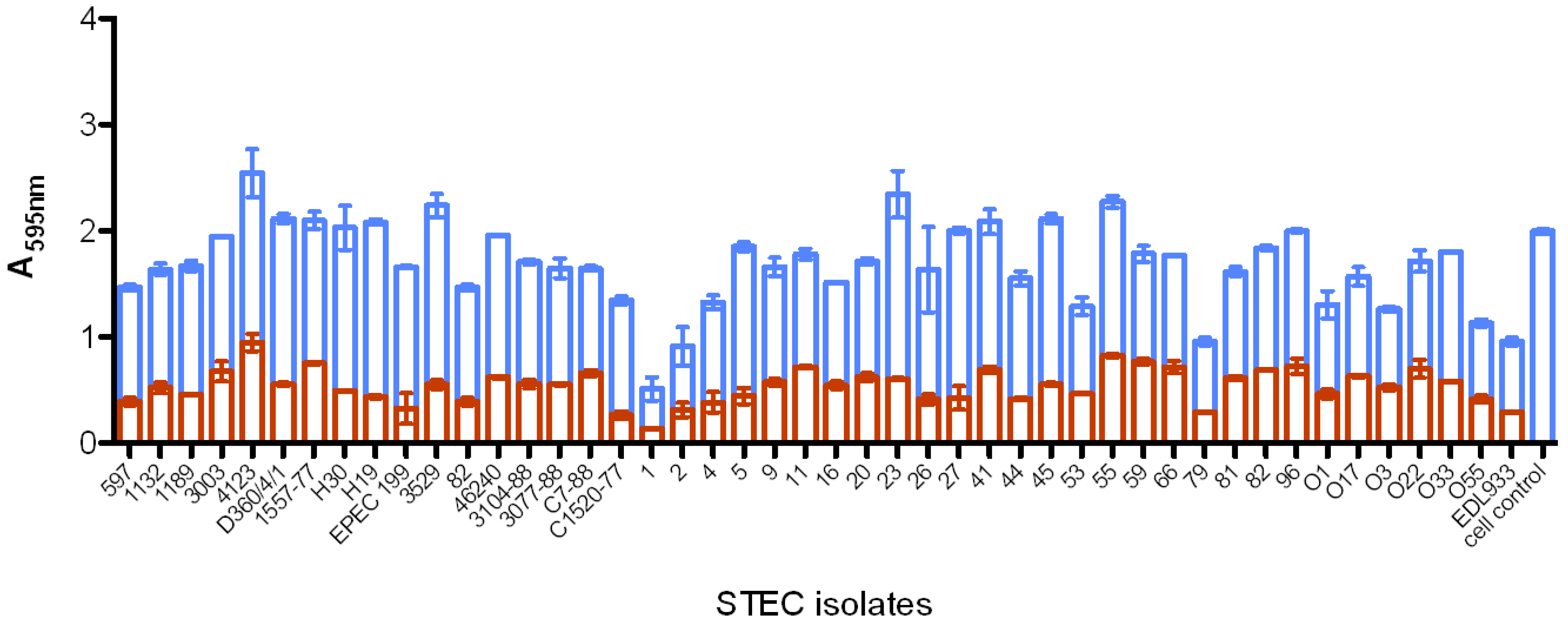
2.5.4. MAb Reactivity to Shiga Toxin-Producing E. coli
2.6. Vero Cell Toxin Assays
2.6.1. VCA
2.6.2. Monoclonal Antibody Neutralizing Assays
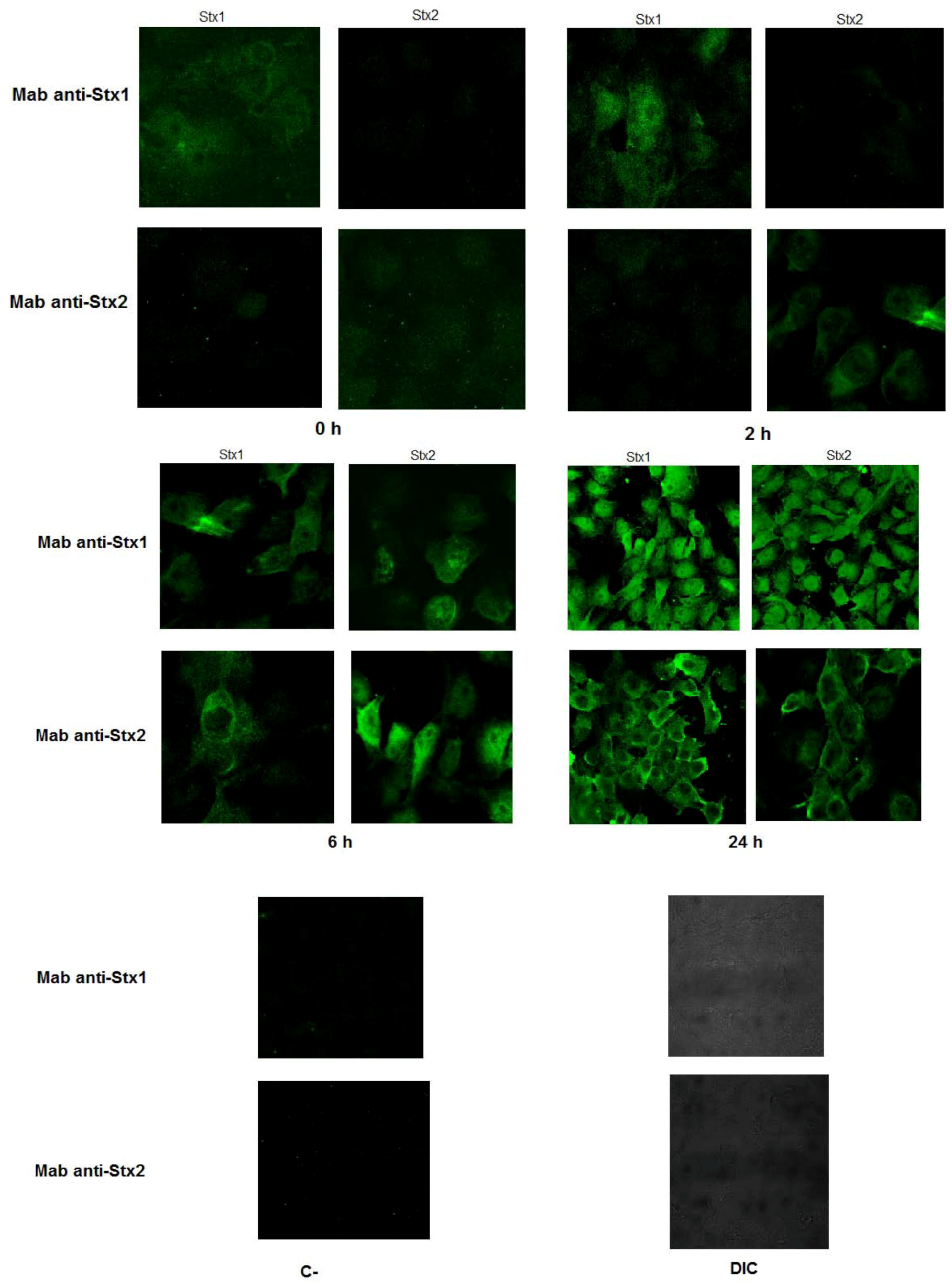
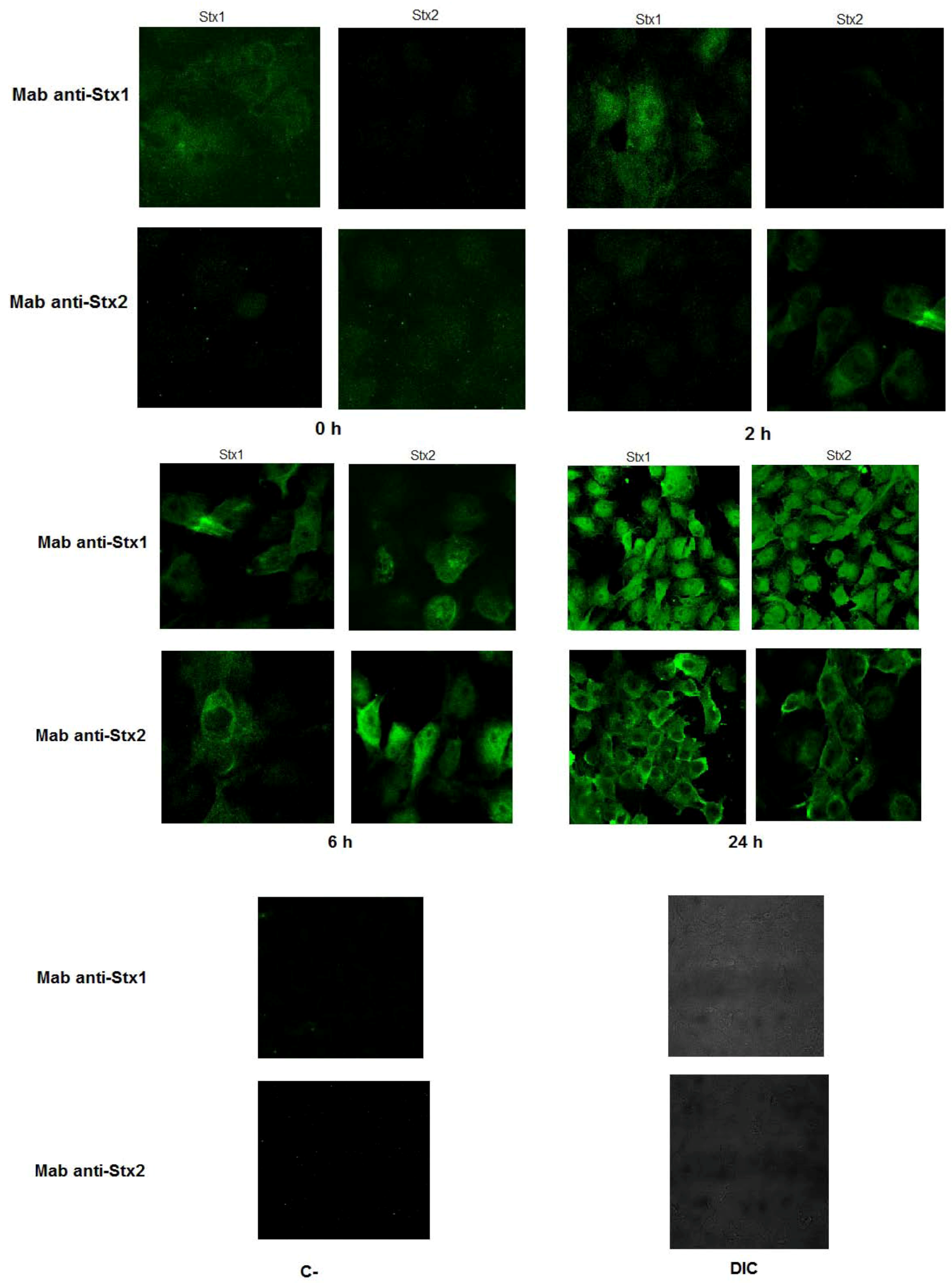
2.6.3. Immunofluorescence and Confocal Analysis
2.6.4. Statistical Analysis
3. Results
3.1. Interaction of Monoclonal Antibodies with Stx1 and Stx2

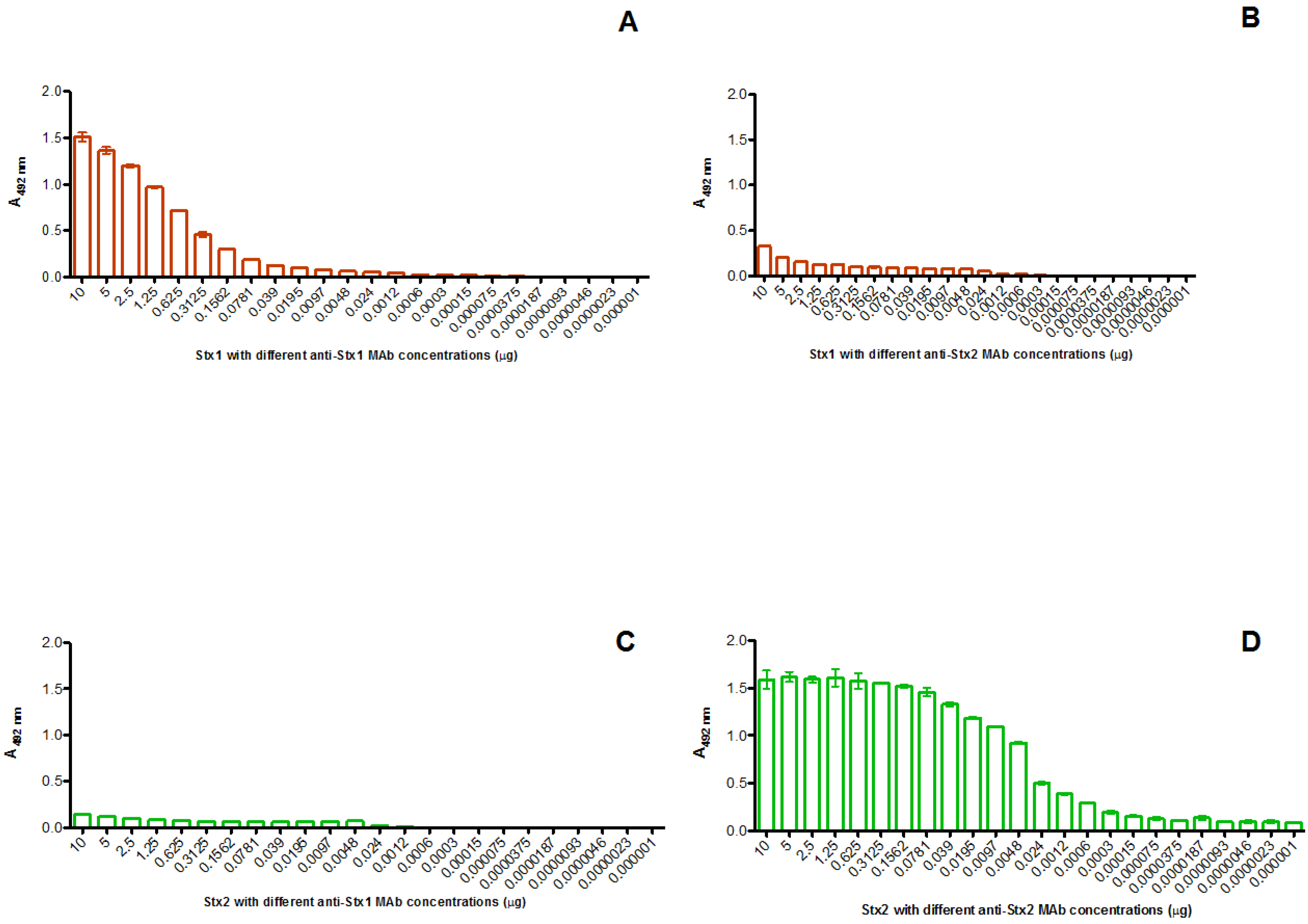

{kind=link}
{kind=link}
{kind=link}
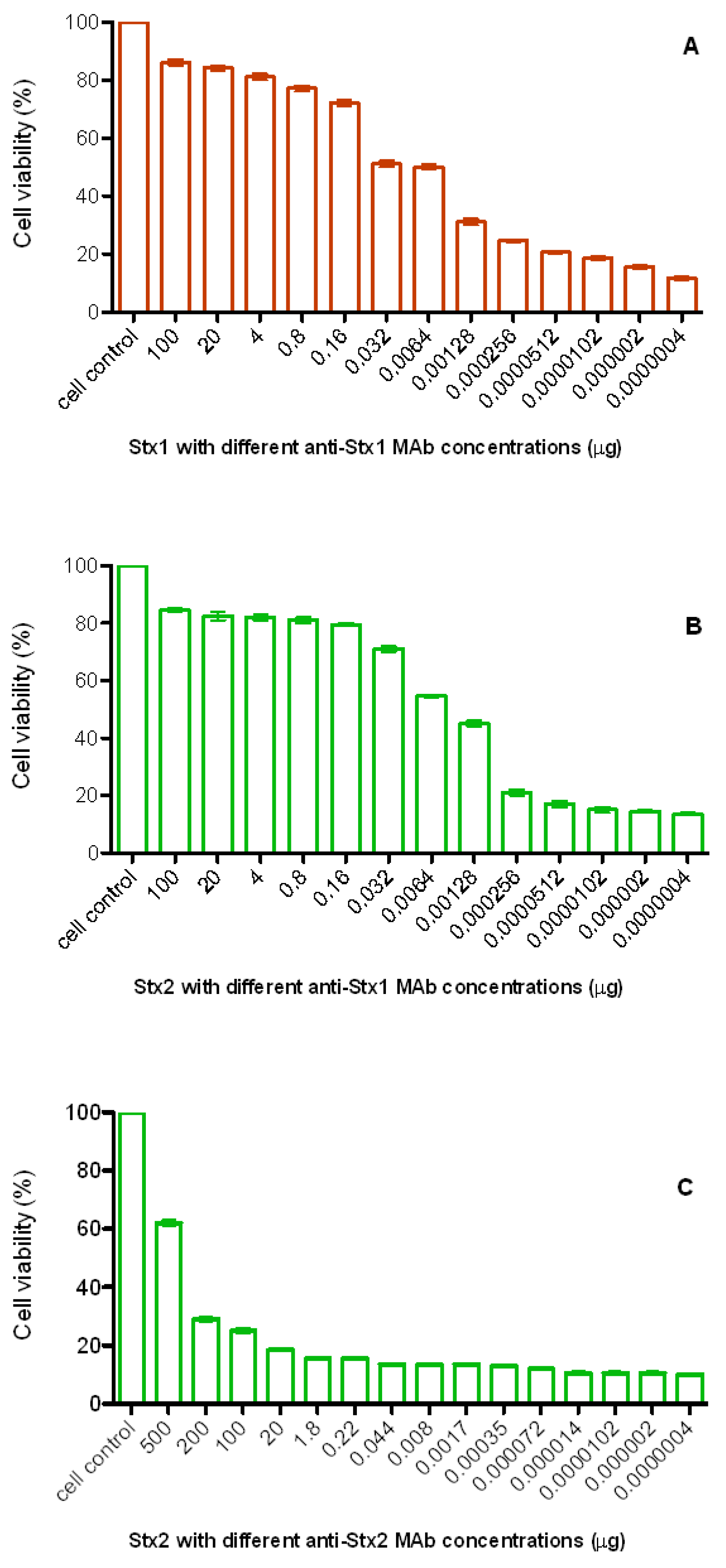{kind=link}
{kind=link}
| MAb characteristic | Anti-Stx1 | Anti-Stx2 |
|---|---|---|
| Hybridomas | 3E2 | 2E11 |
| Dissociation constant (KD) | 2.5 × 10−10 M | 6.1 × 10−10 M |
| Detection limit (200 ng) | 6.2 ng | 12.5 ng |
| Thermostability | 50º C | 70º C |
| Total loss of immunoreactivity (80 ºC) | 1 min | 5 min |
| Partial loss of immunoreactivity | 60 ºC and 70 ºC | 80 ºC and 90 ºC. |
| Strain number | Serotype | Gene presence | Reactivity to MAbs | |
|---|---|---|---|---|
| Anti-Stx1 Anti-Stx2 | ||||
| 597 | OR:NM | stx1 | H | - |
| 1132 | ONT:H49 | stx2 | - | H |
| 1189 | ONT:H49 | stx2 | - | H |
| 3003 | O48:H7 | stx1/stx2 | M | M |
| 4123 | O26:H11 | stx1 | M | - |
| D360/4/1 | O26:H11 | stx1 | M | - |
| 1557-77 | O26:H11 | stx1 | H | - |
| H30 | O26:H11 | stx1 | M | - |
| H19 | O26:H11 | stx1 | M | - |
| EPEC199 | O26:H11 | stx1 | M | - |
| 3529 | O26:H11 | stx1 | M | - |
| 82 | O157:H7 | stx1 | H | - |
| 46240 | O157:H7 | stx1/stx2 | M | H |
| 3104-88 | O157:H7 | stx1/stx2 | M | H |
| 3077-88 | O157:H7 | stx1 | M | - |
| C7-88 | O157:H7 | stx1 | L | - |
| C1520-77 | O157:H7 | stx1/stx2 | H | H |
| 1 | O157:H7 | stx2 | - | H |
| 2 | O157:H7 | stx2 | - | H |
| 4 | O93:H19 | stx1/stx2 | H | L |
| 5 | O55:H19 | stx1 | L | - |
| 9 | O103:H2 | stx1/stx2 | H | H |
| 11 | O118:H16 | stx1 | M | - |
| 16 | O26:H11 | stx1 | L | - |
| 20 | O111:H8 | stx1 | L | - |
| 23 | O111:H8 | stx1 | L | - |
| 26 | O111:NM | stx1 | M | - |
| 27 | O111:NM | stx1 | M | - |
| 41 | ONT:NM | stx2 | - | M |
| 44 | O98:H4 | stx1/stx2 | M | H |
| 45 | O181:H4 | stx1/stx2 | M | H |
| 53 | O98:H17 | stx1/stx2 | L | H |
| 55 | O98:H17 | stx1/stx2 | M | H |
| 59 | ONT:H16 | stx2 | - | L |
| 66 | O105:H18 | stx1/stx2 | H | H |
| 79 | O22:H16 | stx2 | - | H |
| 81 | ONT:H38 | stx1/stx2 | H | H |
| 82 | O112:H21 | stx2 | - | H |
| 96 | O93:H19 | stx2 | - | M |
| O1 | ONT:H8 | stx1 | L | - |
| O17 | O112:H2 | stx1 | L | - |
| O3 | O172:NM | stx2 | - | H |
| O22 | ONT:H16 | stx2 | - | L |
| O36 | O75:H8 | stx1/stx2 | M | - |
| O55 | O146:H21 | stx1/stx2 | H | L |
| EDL 933 | O157:H7 | stx1/stx2 | H | H |
3.2. Neutralizing Ability of Monoclonal Antibodies
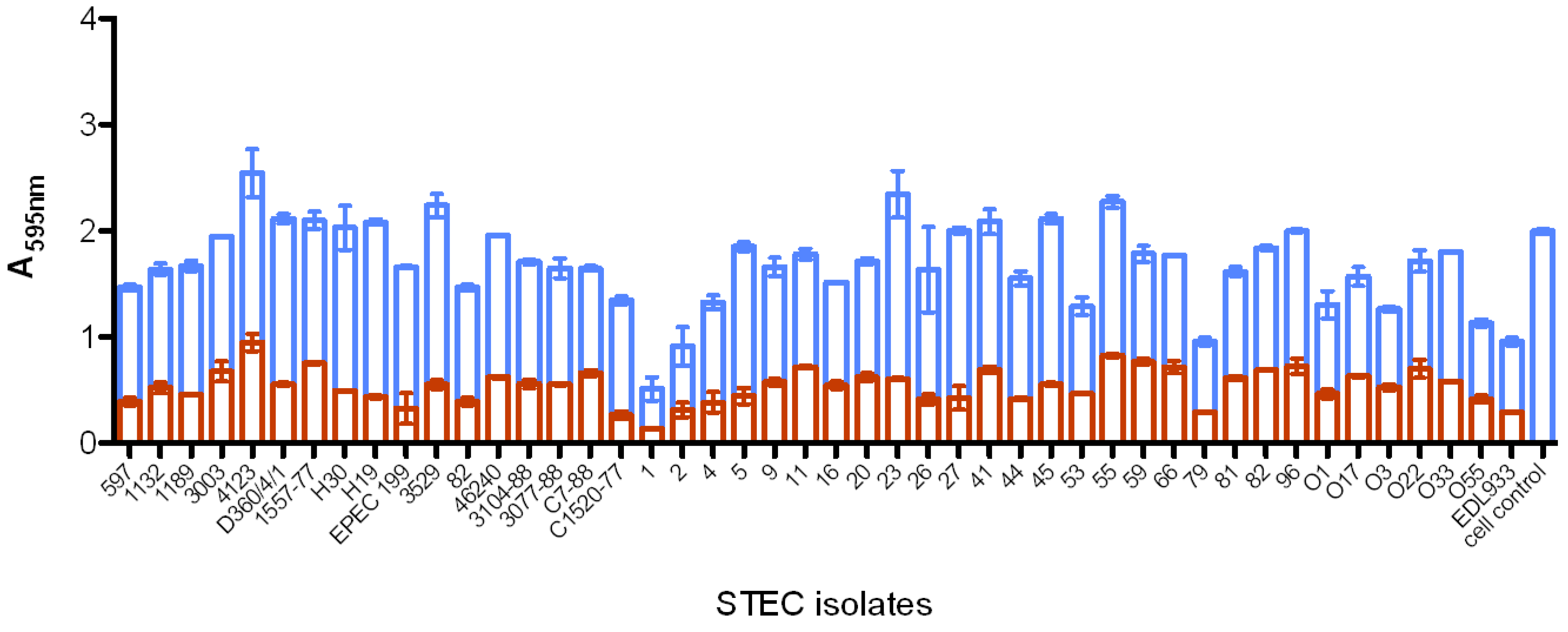
4. Discussion
Acknowledgments
Competing interests
References
- Tzipori, S.; Wachsmuth, I.K.; Chapman, C.; Birden, R.; Brittingham, J.; Jackson, C.; Hogg, J. The pathogenesis of hemorrhagic colitis caused by Escherichia coli O157:H7 in gnotobiotic piglets. J. Infect. Dis. 1986, 154, 712–716. [Google Scholar] [CrossRef]
- Tzipori, S.; Karch, H.; Wachsmuth, K.I.; Robins-Browne, R.M.; O’Brien, A.D.; Lior, H.; Cohen, M.L.; Smithers, J.; Levine, M.M. Role of a 60-megadalton plasmid and Shiga-like toxins in the pathogenesis of infection caused by enterohemorrhagic Escherichia coli O157:H7 in gnotobiotic piglets. Infect. Immun. 1987, 55, 3117–3125. [Google Scholar]
- Levine, M.M. Immunoprophylaxis of Shiga toxin-producing Escherichia coli infection and disease: strengths and weaknesses of various strategies. In Escherichia coli O157:H7 and other Shiga toxin-producing E. coli strains; Kaper, J.B., O’Brien, A.D., Eds.; ASM Press: Washington, D.C., USA, 1998; pp. 405–408. [Google Scholar]
- Fraser, M.E.M.; Chernaia, M.M.; Kozlov, Y.V.; James, M.N. Crystal structure of the holotoxin from Shigella dysenteriae at 2.5 Å resolution. Nat. Struct. Biol. 1994, 1, 59–64. [Google Scholar]
- Fraser, M.E.M.; Fujinaga, M.M.; Cherney, A.R.; Melton-Celsa, E.M.; Twiddy, A.D.; O’Brien, M.N.; James, M.N. Structure of Shiga toxin type 2 (Stx2) from Escherichia coli O157:H7. J. Biol. Chem. 2004, 279, 27511–27517. [Google Scholar]
- Ling, H.; Boodhoo, A.; Hazes, B.; Cummings, M.D.; Armstrong, G.D.; Brunton, J.L.; Read, R.J. Structure of the Shiga-like toxin I B-pentamer complexed with an analogue of its receptor Gb3. Biochemistry 1998, 37, 1777–1788. [Google Scholar]
- Shimizu, H.R.A.; Field, S.W.; Homans, S.W.; Donohue-Rolfe, A. Solution structure of the complex between the B-subunit homopentamer of verotoxin VT-1 from Escherichia coli and the trisaccharide moiety of globotriaosylceramide. Biochemistry 1998, 37, 11078–11082. [Google Scholar]
- Thompson, G.S.; Shimizu, H.; Homans, S.W.; Donohue-Rolfe, A. Localization of the binding site for the oligosaccharide moiety of Gb3 on verotoxin 1 using NMR residual dipolar coupling measurements. Biochemistry 2000, 9, 13153–13156. [Google Scholar]
- Hunt, J.M. Shiga toxin-producing Escherichia coli (STEC). Clin. Lab. Med. 2010, 30, 21–45. [Google Scholar] [CrossRef]
- Zoja, C.; Buelli, S.; Morigi, M. Shiga toxin-associated hemolytic uremic syndrome: pathophysiology of endothelial dysfunction. Pediatr. Nephrol. 2010, 25, 2231–2240. [Google Scholar] [CrossRef]
- Bettelheim, K.A.; Beutin, L. Rapid laboratory identification and characterization of Verocytotoxigenic (Shiga toxin producing) Escherichia coli (VTEC/STEC). J. Appl. Microbiol. 2003, 95, 205–220. [Google Scholar] [CrossRef]
- Gerritzen, A.; Wittke, J.W.; Wolff, D. Rapid and sensitive detection of Shiga toxin-producing Escherichia coli directly from stool samples by real-time PCR in comparison to culture, enzyme immunoassay and Vero cell cytotoxicity assay. Clin. Lab. 2011, 57, 993–998. [Google Scholar]
- CDC U.S. Centers for Disease Control and Prevention. Recommendations for Diagnosis of Shiga Toxin-Producing Escherichia coli Infections by Clinical Laboratories. MMWR 2009, 58, 1–12.
- Kitov, I.P.; Paszkiewics, E.; Sadowska, J.M.; Deng, Z.; Ahmed, M.; Narain, R.; Griener, T.P.; Mulvey, G.L.; Armstrong, G.D.; Bundle, D.R. Impact of the nature and size of the polymeric backbone on the ability of heterobifunctional ligands to mediate shiga toxin and serum amyloid P component ternary complex formation. Toxins 2011, 3, 1065–1088. [Google Scholar] [CrossRef]
- Zhang, X.; McDaniel, A.D.; Wolf, L.E.; Keusch, G.T.; Waldor, M.K.; Acheson, D.W. Quinolone antibiotics induce Shiga toxin-encoding bacteriophages, toxin production, and death in mice. J. Infect. Dis. 2000, 181, 664–670. [Google Scholar] [CrossRef]
- Wong, C.S.; Jelacic, S.; Habeeb, R.L.; Watkins, S.L.; Tarr, P.I. The risk of the hemolytic-uraemic syndrome after antibiotic treatment of Escherichia coli O157:H7 infections. N. Engl. J. Med. 2000, 342, 1930–1936. [Google Scholar] [CrossRef]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar]
- Smith, K.E.; Wilker, P.R.; Reiter, P.L.; Hedican, E.B.; Bender, J.B.; Hedberg, C.W. Antibiotic treatment of Escherichia coli O157 infection and the risk of hemolytic uremic syndrome. Pediatr. Infect. Dis. J. 2012, 31, 37–41. [Google Scholar] [CrossRef]
- Smith, M.J.; Teel, L.D.; Carvalho, H.M.; Melton-Celsa, A.R.; O’Brien, A.D. Development of a hybrid Shiga holotoxoid vaccine to elicit heterologous protection against Shiga toxins types 1 and 2. Vaccine 2006, 24, 4122–4129. [Google Scholar]
- Wen, S. X.; Teel, L.D.; Judge, N.A.; O’Brien, A.D. A plant-based oral vaccine to protect against systemic intoxication by Shiga toxin type 2. Proc. Natl. Acad. Sci.USA 2006, 103, 7082–7087. [Google Scholar]
- Wen, S.X.; Teel, L.D.; Judge, N.A.; O’Brien, A.D. Genetic toxoids of Shiga toxin types 1 and 2 protect mice against homologous but not heterologous toxin challenge. Vaccine 2006, 24, 1142–1148. [Google Scholar] [CrossRef]
- Edwards, A.C.; Melton-Celsa, A.R.; Arbuthnott, K.; Stinson, J.R.; Schmitt, C.K.; Wong, H.C.; O’Brien, A.D. Vero cell neutralization and mouse protective efficacy of humanized monoclonal antibodies against Escherichia coli toxins Stx1 and Stx2. In Escherichia coli O157:H7 and other Shiga toxin-producing E. coli strains; Kaper, J.B., O’Brien, A.D., Eds.; ASM Press: Washington, D.C., USA, 1998; pp. 388–392. [Google Scholar]
- Nakao, H.; Kiyokawa, N.; Fujimoto, J.; Yamasaki, S.; Takeda, T. Monoclonal antibody to Shiga toxin 2 which blocks receptor binding and neutralizes cytotoxicity. Infect. Immun. 1999, 67, 5717–5722. [Google Scholar]
- Nakao, H.; Kataoka, C.; Kiyokawa, N.; Fujimoto, J.; Yamasaki, S.; Takeda, T. Monoclonal antibody to Shiga toxin 1, which blocks receptor binding and neutralizes cytotoxicity. Microbiol. Immunol. 2002, 46, 777–780. [Google Scholar]
- Yamagami, S.; Motoki, M.; Kimura, T.; Izumi, H.; Takeda, T.; Katsuura, Y.; Matsumoto, Y. Efficacy of postinfection treatment with anti-Shiga toxin (Stx) 2 humanized monoclonal antibody TMA-15 in mice lethally challenged with Stx-producing Escherichia coli. J. Infect. Dis. 2001, 184, 738–742. [Google Scholar] [CrossRef]
- Kimura, T.; Co, M.S.; Vasquez, M.; Wei, S.; Xu, H.; Tani, S.; Sakai, Y.; Kawamura, T.; Matsumoto, Y.; Nakao, H.; Takeda, T. Development of humanized monoclonal antibody TMA-15 which neutralizes Shiga toxin 2. Hybrid Hybridomics 2002, 21, 161–168. [Google Scholar] [CrossRef]
- Mukherjee, J.; Chios, K.; Fishwild, D.; Hudson, D.; O’Donnell, S.; Rich, S.M.; Donohue-Rolfe, A.; Tzipori, S. Human Stx2-specific monoclonal antibodies prevent systemic complications of Escherichia coli O157:H7 infection. Infect. Immun. 2002, 70, 612–619. [Google Scholar] [CrossRef]
- Mukherjee, J.; Chios, K.; Fishwild, D.; Hudson, D.; O’Donnell, S.; Rich, S.M.; Donohue-Rolfe, A.; Tzipori, S. Production and characterization of protective human antibodies against Shiga toxin 1. Infect. Immun. 2002, 70, 5896–5899. [Google Scholar] [CrossRef]
- Dowling, T.C.; Chavaillaz, P.A.; Young, D.G.; Melton-Celsa, A.; O’Brien, A.; Thuning-Roberson, C.; Edelman, R.; Tacket, C.O. Phase 1 safety and pharmacokinetic study of chimeric murine-human monoclonal antibody c alpha Stx2 administered intravenously to healthy adult volunteers. Antimicrob. Agents Chemother. 2005, 49, 1808–1812. [Google Scholar] [CrossRef]
- Smith, M.J.; Carvalho, H.M.; Melton-Celsa, A.R.; O’Brien, A.D. The 13C4 antibody that neutralizes Shiga toxin type 1 (Stx1) recognizes three regions on the Stx1 B subunit and prevents Stx1 from binding to its eukaryotic receptor globotriaosylceramide. Infect. Immun. 2006, 74, 6992–6998. [Google Scholar]
- Lopez, E.L.; Contrini, M.M.; Glatstein, E.; Ayala, S.G.; Santoro, R.; Allende, D.; Ezcurra, G.; Teplitz, E.; Koyama, T.; Matsumoto, Y.; Sato, H.; Sakai, K.; Hoshide, S.; Komoriya, K.; Morita, T.; Harning, R.; Brookman, S. Safety and pharmacokinetics of urtoxazumab, a humanized monoclonal antibody, against Shiga-like toxin 2 in healthy adults and in pediatric patients infected with Shiga-like toxin-producing Escherichia coli. Antimicrob. Agents Chemother. 2010, 54, 239–243. [Google Scholar]
- Vaz, T.M.; Irino, K.; Kato, M.A.; Dias, A.M.; Gomes, T.A.; Medeiros, M.L.; Rocha, M.M.; Guth, B.E. Virulence properties and characteristics of Shiga toxin-producing Escherichia coli in São Paulo, Brazil, from 1976 through 1999. J. Clin. Microbiol. 2004, 42, 903–905. [Google Scholar]
- Rocha, L.B.; Piazza, R.M. Production of Shiga toxin by Shiga toxin-expressing Escherichia coli (STEC) in broth media: from divergence to definition. Lett. Appl. Microbiol. 2007, 45, 411–417. [Google Scholar] [CrossRef]
- Mendes-Ledesma, M.R.; Rocha, L.B.; Bueris, V.; Krause, G.; Beutin, L.; Franzolin, M.R.; Trabulsi, L.R.; Elias, W.P.; Piazza, R.M. Production and characterization of rabbit polyclonal sera against Shiga toxins Stx1 and Stx2 for detection of Shiga toxin-producing Escherichia coli. Microbiol. Immunol. 2008, 52, 484–491. [Google Scholar] [CrossRef]
- Donohue-Rolfe, A.; Keusch, G.T.; Edson, C.; Thorley-Lawson, D.; Jacewicz, M. Pathogenesis of Shigella diarrhea. IX. Simplified high yield purification of Shigella toxin and characterization of subunit composition and function by the use of subunit-specific monoclonal and polyclonal antibodies. J. Exp. Med. 1984, 160, 1767–1781. [Google Scholar]
- Brown, J.E.; Griffin, D.E.; Rothman, S.W.; Proctor, B.P. Purification and biological characterization of Shiga toxin from Shigella dysenteriae. Infect. Immun. 1982, 36, 996–1005. [Google Scholar]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Biotechnology 1975, 24, 524–526. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Studier, F.W. Analysis of bacteriophage T7 early RNAs and proteins on slab gels. J. Mol. Biol. 1973, 79, 237–248. [Google Scholar] [CrossRef]
- Friguet, B.; Chaffotte, A.F.; Djavadi-Ohaniance, L.; Goldberg, M.E. Measurements of the true affinity constant in solution of antigen-antibody complexes by enzyme-linked immunosorbent assay. J. Immunol. Methods 1985, 2, 305–319. [Google Scholar]
- Gentry, M.K.; Dalrymple, J.M. Quantitative microtiter cytotoxicity assay for Shigella toxin. J. Clin. Microbiol. 1980, 12, 361–366. [Google Scholar]
- Beutin, L.; Zimmermann, S.; Gleier, K. Evaluation of the VTEC-screen 'Seiken' test for detection of different types of Shiga toxin (verotoxin)-producing Escherichia coli (STEC) in human stool samples. Diagn. Microbiol. Infect. Dis. 2002, 42, 1–8. [Google Scholar] [CrossRef]
- Dorsey, F.C.; Fischer, J.F.; Fleckenstein, J.M. Directed delivery of heat-labile enterotoxin by enterotoxigenic Escherichia coli. Cell Microbiol. 2006, 8, 1516–1527. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Marques, L.R.; Holmes, R.K.; O'Brien, A.D. Characterization of monoclonal antibodies against Shiga-like toxin from Escherichia coli. Infect. Immun. 1985, 50, 695–700. [Google Scholar]
- Perera, L.P.; Marques, L.R.; O'Brien, A.D. Isolation and characterization of monoclonal antibodies to Shiga-like toxin II of enterohemorrhagic Escherichia coli and use of the monoclonal antibodies in a colony enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1988, 26, 2127–2131. [Google Scholar]
- Downes, F.P.; Barrett, T.J.; Green, J.H.; Aloisio, C.H.; Spika, J.S.; Strockbine, N.A.; Wachsmuth, I.K. Affinity purification and characterization of Shiga-like toxin II and production of toxin-specific monoclonal antibodies. Infect. Immun. 1988, 56, 1926–1933. [Google Scholar]
- Sheoran, A.S.; Chapman, S.; Singh, P.; Donohue-Rolfe, A.; Tzipori, S. Stx2-specific human monoclonal antibodies protect mice against lethal infection with Escherichia coli expressing Stx2 variants. Infect. Immun. 2003, 71, 3125–3130. [Google Scholar] [CrossRef]
- Mohawk, K.L.; Melton-Celsa, A.R.; Robinson, C.M.; O'Brien, A.D. Neutralizing antibodies to Shiga toxin type 2 (Stx2) reduce colonization of mice by Stx2-expressing Escherichia coli O157:H7. Vaccine 2010, 28, 4777–4785. [Google Scholar] [CrossRef]
- Li, G.; Hong, J.; Huo, G.; Ren, X. Monoclonal antibodies against Stx1B subunit of Escherichia coli O157:H7 distinguish the bacterium form other bacteria. Lett. App. Microbiol. 2010, 51, 499–503. [Google Scholar] [CrossRef]
- Menezes, C.A.; Gonçalves, D.S.; Amianti, J.; Fernandes, I.; Taddei, C.R.; Koga, P.C.M.; Trabulsi, L.R.; Martinez, M.B.; Piazza, R.M. Capture immunoassay for LT detection produced by enterotoxigenic Escherichia coli in bacterial isolates. Braz. J. Microbiol. 2003, 34, 11–13. [Google Scholar] [CrossRef]
- Menezes, C.A.; Imamura, S.Y.; Trabulsi, L.R.; Fernandes-Filho, A.; Martinez, M.B.; Guth, B.E.; Girão, D.M.; Piazza, R.M. Production, characterization, and application of antibodies against heat-labile type-I toxin for detection of enterotoxigenic Escherichia coli. Mem. Inst. Oswaldo Cruz 2006, 101, 875–880. [Google Scholar]
- Menezes, M.A.; Rocha, L.B.; Koga, P.C.; Fernandes, I.; Nara, J.M.; Magalhães, C.A.; Abe, C.M.; Ayala, C.O.; Burgos, Y.K.; Elias, W.P.; Castro, A.F.; Piazza, R.M. Identification of enteropathogenic and enterohaemorrhagic Escherichia coli strains by immunoserological detection of intimin. J. Appl. Microbiol. 2010, 108, 878–887. [Google Scholar] [CrossRef]
- Tanikawa, T.; Ishikawa, T.; Maekawa, T.; Kuronane, K.; Imai, Y. Characterization of monoclonal immunoglobulin A and G against Shiga toxin binding subunits produced by intranasal immunization. Scand. J. Immunol. 2008, 68, 414–422. [Google Scholar] [CrossRef]
- Sheoran, A.S.; Chapman-Bonofiglio, S.; Harvey, B.R.; Mukherjee, J.; Georgiou, G.; Donohue-Rolfe, A.; Tzipori, S. Human antibody against Shiga toxin 2 administered to piglets after the onset of diarrhea due to Escherichia coli O157:H7 prevents fatal systemic complications. Infect. Immun. 2005, 73, 4607–4613. [Google Scholar] [CrossRef]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef]
- Jeong, K.I.; Chapman-Bonofiglio, S.; Singh, P.; Lee, J.; Tzipori, S.; Sheoran, A.S. In vitro and in vivo protective efficacies of antibodies that neutralize the RNA N-glycosidase activity of Shiga toxin 2. BMC Immunol. 2010, 24, 11–16. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rocha, L.B.; Luz, D.E.; Moraes, C.T.P.; Caravelli, A.; Fernandes, I.; Guth, B.E.C.; Horton, D.S.P.Q.; Piazza, R.M.F. Interaction between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and in Vitro Neutralizing Abilities. Toxins 2012, 4, 729-747. https://doi.org/10.3390/toxins4090729
Rocha LB, Luz DE, Moraes CTP, Caravelli A, Fernandes I, Guth BEC, Horton DSPQ, Piazza RMF. Interaction between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and in Vitro Neutralizing Abilities. Toxins. 2012; 4(9):729-747. https://doi.org/10.3390/toxins4090729
Chicago/Turabian StyleRocha, Letícia B., Daniela E. Luz, Claudia T. P. Moraes, Andressa Caravelli, Irene Fernandes, Beatriz E. C. Guth, Denise S. P. Q. Horton, and Roxane M. F. Piazza. 2012. "Interaction between Shiga Toxin and Monoclonal Antibodies: Binding Characteristics and in Vitro Neutralizing Abilities" Toxins 4, no. 9: 729-747. https://doi.org/10.3390/toxins4090729