Role of Phospholipase A2 in Retrograde Transport of Ricin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and Transfection
2.3. Cytotoxicity Assay
2.4. Sulfation Assay
2.5. Mannosylation Assay
2.6. Endocytosis of 125I-ricin
2.7. Confocal Microscopy
3. Results
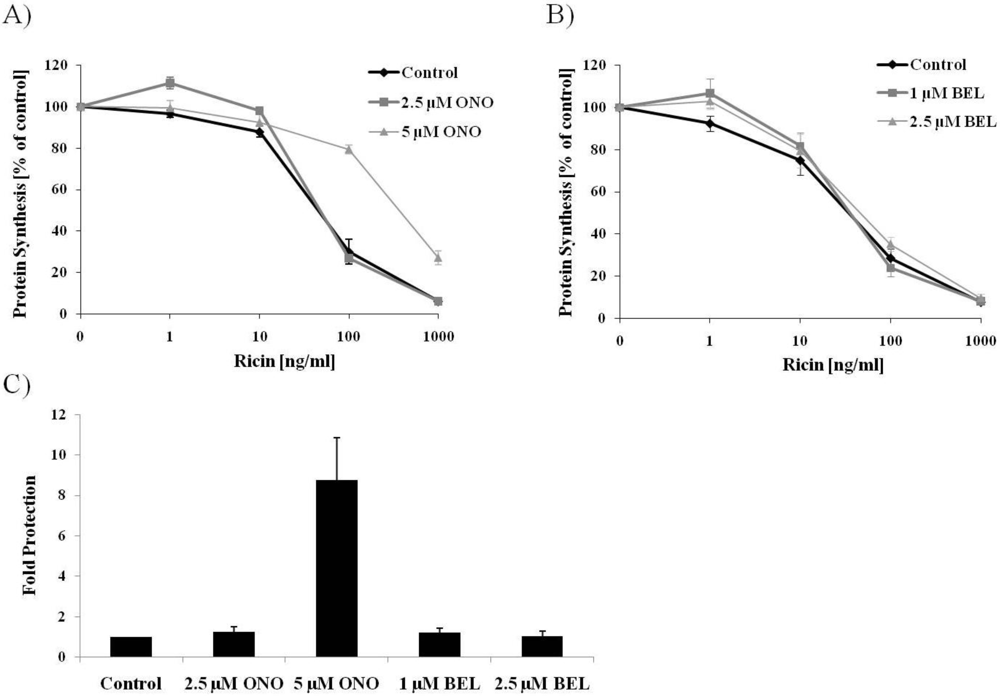
3.1. PLA2 Inhibitor Protects Cells against Ricin
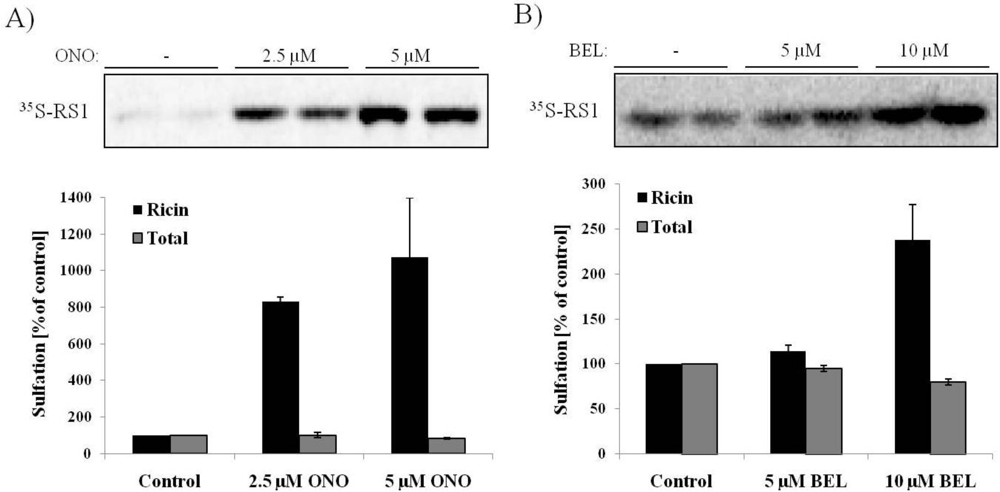
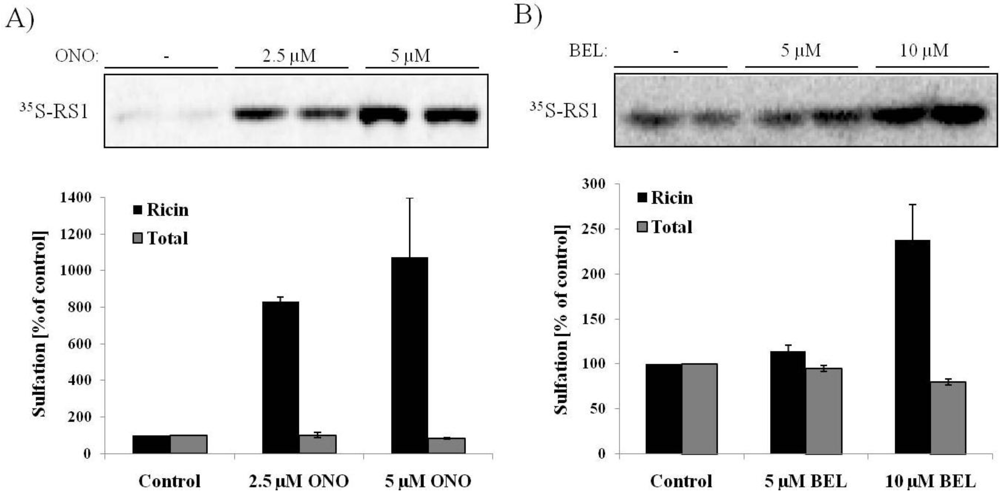
3.2. Ricin Sulfation Is Strongly Increased in Response to PLA2 Inhibition
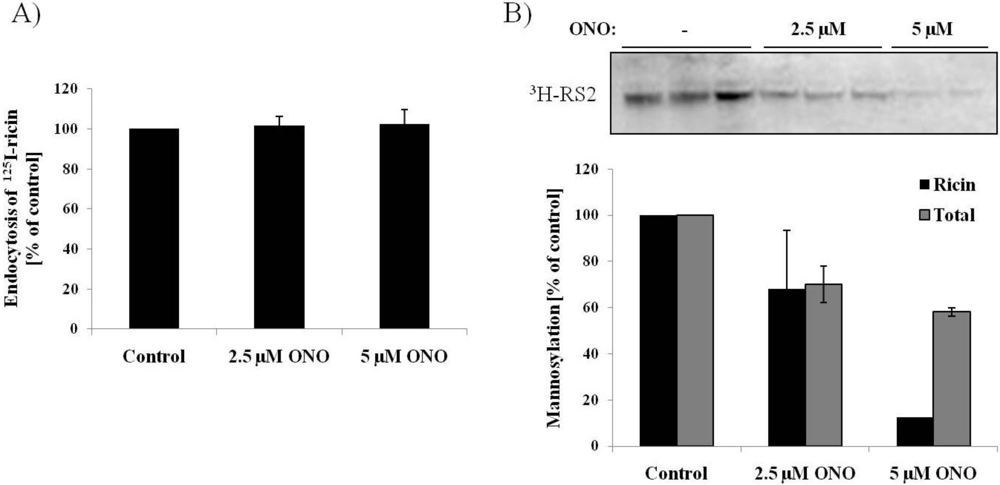
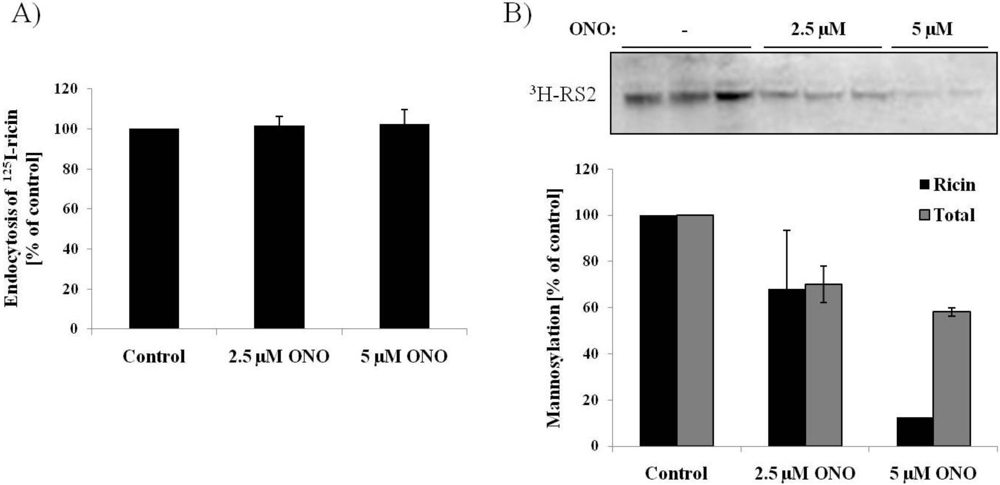
3.3. Ricin Transport to the Endoplasmic Reticulum is Dependent on PLA2 Activity
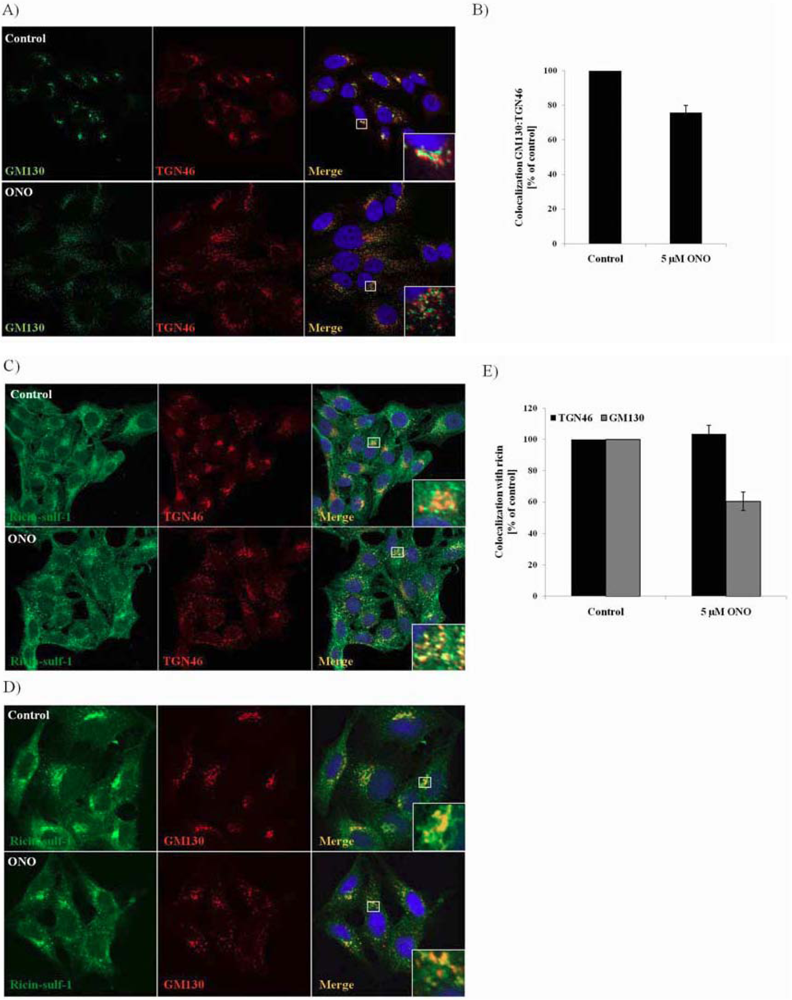
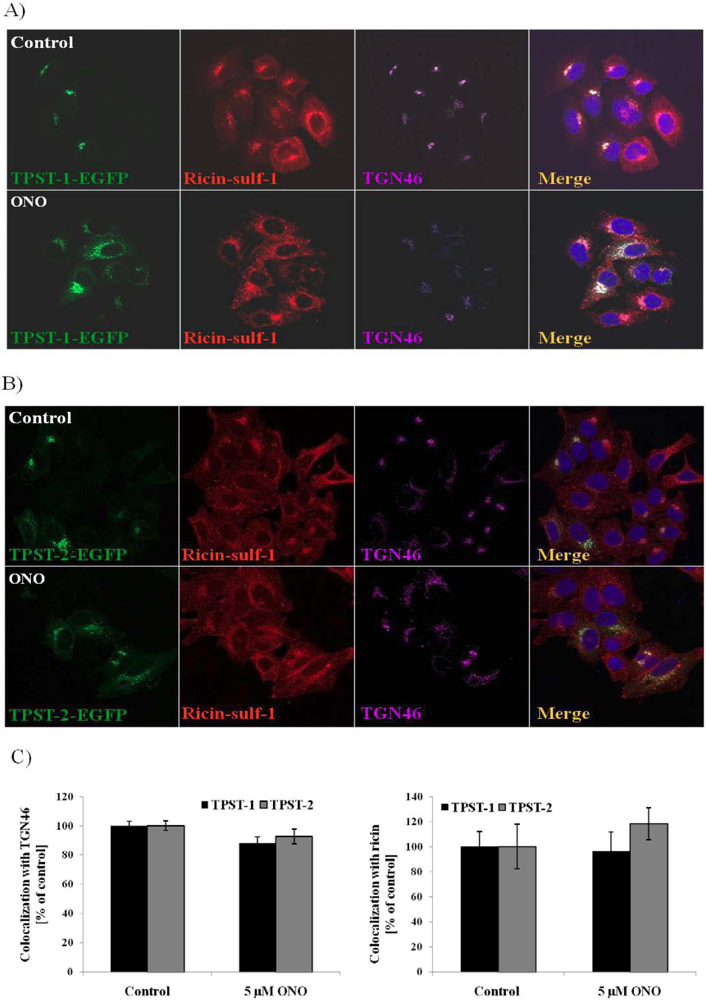
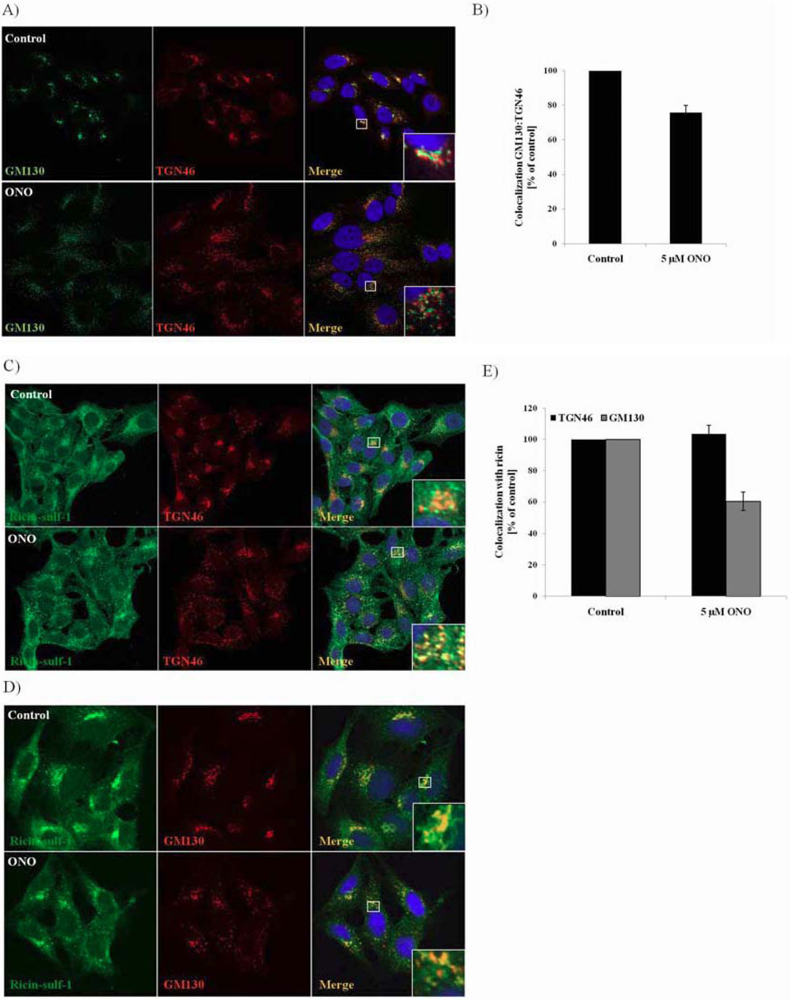
3.4. Localization of Ricin and Golgi Markers in the Presence of PLA2 Inhibitor ONO
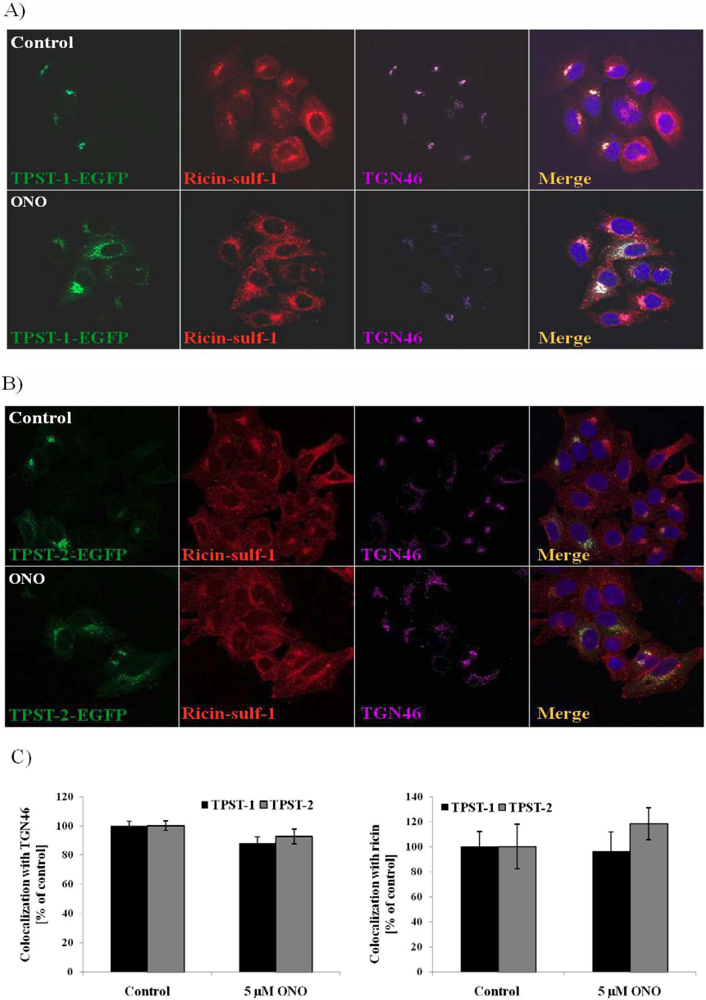
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
Supplementary Files
References
- Audi, J.; Belson, M.; Patel, M.; Schier, J.; Osterloh, J. Ricin poisoning: A comprehensive review. J. Am. Med. Assoc. 2005, 294, 2342–2351. [Google Scholar]
- Sandvig, K.; van Deurs, B. Delivery into cells: Lessons learned from plant and bacterial toxins. Gene Ther. 2005, 12, 865–872. [Google Scholar]
- Sandvig, K.; Pust, S.; Skotland, T.; van Deurs, B. Clathrin-independent endocytosis: Mechanisms and function. Curr. Opin. Cell Biol. 2011, 23, 1–8. [Google Scholar]
- Sandvig, K.; Torgersen, M.L.; Engedal, N.; Skotland, T.; Iversen, T.G. Protein toxins from plants and bacteria: Probes for intracellular transport and tools in medicine. FEBS Lett. 2010, 584, 2626–2634. [Google Scholar]
- Sandvig, K.; Prydz, K.; Hansen, S.H.; van Deurs, B. Ricin transport in brefeldin A-treated cells: Correlation between Golgi structure and toxic effect. J. Cell Biol. 1991, 115, 971–981. [Google Scholar]
- Stechmann, B.; Bai, S.K.; Gobbo, E.; Lopez, R.; Merer, G.; Pinchard, S.; Panigai, L.; Tenza, D.; Raposo, G.; Beaumelle, B.; et al. Inhibition of retrograde transport protects mice from lethal ricin challenge. Cell 2010, 141, 231–242. [Google Scholar] [PubMed]
- Johannes, L.; Popoff, V. Tracing the retrograde route in protein trafficking. Cell 2008, 135, 1175–1187. [Google Scholar]
- Grimmer, S.; Iversen, T.G.; van Deurs, B.; Sandvig, K. Endosome to Golgi transport of ricin is regulated by cholesterol. Mol. Biol. Cell 2000, 11, 4205–4216. [Google Scholar]
- Rodal, S.K.; Skretting, G.; Garred, O.; Vilhardt, F.; van Deurs, B.; Sandvig, K. Extraction of cholesterol with methyl-beta-cyclodextrin perturbs formation of clathrin-coated endocytic vesicles. Mol. Biol. Cell 1999, 10, 961–974. [Google Scholar]
- Falguieres, T.; Mallard, F.; Baron, C.; Hanau, D.; Lingwood, C.; Goud, B.; Salamero, J.; Johannes, L. Targeting of Shiga toxin B-subunit to retrograde transport route in association with detergent-resistant membranes. Mol. Biol. Cell 2001, 12, 2453–2468. [Google Scholar]
- Grimmer, S.; Spilsberg, B.; Hanada, K.; Sandvig, K. Depletion of sphingolipids facilitates endosome to Golgi transport of ricin. Traffic 2006, 7, 1243–1253. [Google Scholar]
- Raa, H.; Grimmer, S.; Schwudke, D.; Bergan, J.; Walchli, S.; Skotland, T.; Shevchenko, A.; Sandvig, K. Glycosphingolipid requirements for endosome-to-Golgi transport of Shiga toxin. Traffic 2009, 10, 868–882. [Google Scholar]
- Sandvig, K.; Garred, O.; Prydz, K.; Kozlov, J.V.; Hansen, S.H.; van Deurs, B. Retrograde transport of endocytosed Shiga toxin to the endoplasmic reticulum. Nature 1992, 358, 510–512. [Google Scholar]
- Sandvig, K.; Garred, O.; van Helvoort, A.; van Meer, G.; van Deurs, B. Importance of glycolipid synthesis for butyric acid-induced sensitization to shiga toxin and intracellular sorting of toxin in A431 cells. Mol. Biol. Cell 1996, 7, 1391–1404. [Google Scholar]
- Lingwood, C.A.; Manis, A.; Mahfoud, R.; Khan, F.; Binnington, B.; Mylvaganam, M. New aspects of the regulation of glycosphingolipid receptor function. Chem. Phys. Lipids 2010, 163, 27–35. [Google Scholar]
- Lingwood, C.A.; Binnington, B.; Manis, A.; Branch, D.R. Globotriaosyl ceramide receptor function—where membrane structure and pathology intersect. FEBS Lett. 2010, 584, 1879–1886. [Google Scholar]
- Spilsberg, B.; Llorente, A.; Sandvig, K. Polyunsaturated fatty acids regulate Shiga toxin transport. Biochem. Biophys. Res. Commun. 2007, 364, 283–288. [Google Scholar]
- Mukherjee, S.; Maxfield, F.R. Role of membrane organization and membrane domains in endocytic lipid trafficking. Traffic 2000, 1, 203–211. [Google Scholar]
- Spilsberg, B.; van Meer, G.; Sandvig, K. Role of lipids in the retrograde pathway of ricin intoxication. Traffic 2003, 4, 544–552. [Google Scholar]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar]
- Brown, W.J.; Chambers, K.; Doody, A. Phospholipase A2 (PLA2) enzymes in membrane trafficking: Mediators of membrane shape and function. Traffic 2003, 4, 214–221. [Google Scholar]
- Choukroun, G.J.; Marshansky, V.; Gustafson, C.E.; McKee, M.; Hajjar, R.J.; Rosenzweig, A.; Brown, D.; Bonventre, J.V. Cytosolic phospholipase A(2) regulates golgi structure and modulates intracellular trafficking of membrane proteins. J. Clin. Invest 2000, 106, 983–993. [Google Scholar]
- de Figueiredo, P.; Drecktrah, D.; Katzenellenbogen, J.A.; Strang, M.; Brown, W.J. Evidence that phospholipase A2 activity is required for Golgi complex and trans Golgi network membrane tubulation. Proc. Natl. Acad. Sci. USA 1998, 95, 8642–8647. [Google Scholar]
- de Figueiredo, P.; Polizotto, R.S.; Drecktrah, D.; Brown, W.J. Membrane tubule-mediated reassembly and maintenance of the Golgi complex is disrupted by phospholipase A2 antagonists. Mol. Biol. Cell 1999, 10, 1763–1782. [Google Scholar]
- Schmidt, J.A.; Kalkofen, D.N.; Donovan, K.W.; Brown, W.J. A role for phospholipase A2 activity in membrane tubule formation and TGN trafficking. Traffic 2010, 11, 1530–1536. [Google Scholar]
- de Figueiredo, P.; Doody, A.; Polizotto, R.S.; Drecktrah, D.; Wood, S.; Banta, M.; Strang, M.S.; Brown, W.J. Inhibition of transferrin recycling and endosome tubulation by phospholipase A2 antagonists. J. Biol. Chem. 2001, 276, 47361–47370. [Google Scholar]
- Bechler, M.E.; Doody, A.M.; Ha, K.D.; Judson, B.L.; Chen, I.; Brown, W.J. The phospholipase A2 enzyme complex PAFAH Ib mediates endosomal membrane tubule formation and trafficking. Mol. Biol. Cell 2011, 22, 2348–2359. [Google Scholar]
- Doody, A.M.; Antosh, A.L.; Brown, W.J. Cytoplasmic phospholipase A2 antagonists inhibit multiple endocytic membrane trafficking pathways. Biochem. Biophys. Res. Commun. 2009, 388, 695–699. [Google Scholar]
- de Figueiredo, P.; Drecktrah, D.; Polizotto, R.S.; Cole, N.B.; Lippincott-Schwartz, J.; Brown, W.J. Phospholipase A2 antagonists inhibit constitutive retrograde membrane traffic to the endoplasmic reticulum. Traffic 2000, 1, 504–511. [Google Scholar]
- Polizotto, R.S.; de, F.P.; Brown, W.J. Stimulation of Golgi membrane tubulation and retrograde trafficking to the ER by phospholipase A(2) activating protein (PLAP) peptide. J. Cell Biochem. 1999, 74, 670–683. [Google Scholar]
- Drecktrah, D.; Chambers, K.; Racoosin, E.L.; Cluett, E.B.; Gucwa, A.; Jackson, B.; Brown, W.J. Inhibition of a Golgi complex lysophospholipid acyltransferase induces membrane tubule formation and retrograde trafficking. Mol. Biol. Cell 2003, 14, 3459–3469. [Google Scholar]
- Grimmer, S.; Ying, M.; Walchli, S.; van Deurs, B.; Sandvig, K. Golgi vesiculation induced by cholesterol occurs by a dynamin- and cPLA2-dependent mechanism. Traffic 2005, 6, 144–156. [Google Scholar]
- Rapak, A.; Falnes, P.O.; Olsnes, S. Retrograde transport of mutant ricin to the endoplasmic reticulum with subsequent translocation to cytosol. Proc. Natl. Acad. Sci. USA 1997, 94, 3783–3788. [Google Scholar]
- Sandvig, K.; Bergan, J.; Dyve, A.B.; Skotland, T.; Torgersen, M.L. Endocytosis and retrograde transport of Shiga toxin. Toxicon 2009, 56, 1181–1185. [Google Scholar]
- Huttner, W.B. Tyrosine sulfation and the secretory pathway. Annu. Rev. Physiol 1988, 50, 363–376. [Google Scholar]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta 2006, 1761, 1246–1259. [Google Scholar]
- Banga, H.S.; Simons, E.R.; Brass, L.F.; Rittenhouse, S.E. Activation of phospholipases A and C in human platelets exposed to epinephrine: Role of glycoproteins IIb/IIIa and dual role of epinephrine. Proc. Natl. Acad. Sci. USA 1986, 83, 9197–9201. [Google Scholar]
- Ackermann, E.J.; Conde-Frieboes, K.; Dennis, E.A. Inhibition of macrophage Ca(2+)-independent phospholipase A2 by bromoenol lactone and trifluoromethyl ketones. J. Biol. Chem. 1995, 270, 445–450. [Google Scholar]
- Hazen, S.L.; Zupan, L.A.; Weiss, R.H.; Getman, D.P.; Gross, R.W. Suicide inhibition of canine myocardial cytosolic calcium-independent phospholipase A2. Mechanism-based discrimination between calcium-dependent and -independent phospholipases A2. J. Biol. Chem. 1991, 266, 7227–7232. [Google Scholar] [PubMed]
- San, P.E.; Capestrano, M.; Polishchuk, E.V.; DiPentima, A.; Trucco, A.; Zizza, P.; Mariggio, S.; Pulvirenti, T.; Sallese, M.; Tete, S.; et al. Group IV phospholipase A(2)alpha controls the formation of inter-cisternal continuities involved in intra-Golgi transport. PLoS. Biol. 2009, 7, e1000194. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, R.K.; Aoki, J.; Kano, F.; Murata, M.; Yamamoto, A.; Tsujimoto, M.; Arai, H. Intracellular phospholipase A1gamma (iPLA1gamma) is a novel factor involved in coat protein complex I- and Rab6-independent retrograde transport between the endoplasmic reticulum and the Golgi complex. J. Biol. Chem. 2009, 284, 26620–26630. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Klokk, T.I.; Lingelem, A.B.D.; Myrann, A.-G.; Sandvig, K. Role of Phospholipase A2 in Retrograde Transport of Ricin. Toxins 2011, 3, 1203-1219. https://doi.org/10.3390/toxins3091203
Klokk TI, Lingelem ABD, Myrann A-G, Sandvig K. Role of Phospholipase A2 in Retrograde Transport of Ricin. Toxins. 2011; 3(9):1203-1219. https://doi.org/10.3390/toxins3091203
Chicago/Turabian StyleKlokk, Tove Irene, Anne Berit Dyve Lingelem, Anne-Grethe Myrann, and Kirsten Sandvig. 2011. "Role of Phospholipase A2 in Retrograde Transport of Ricin" Toxins 3, no. 9: 1203-1219. https://doi.org/10.3390/toxins3091203