Accumulation Kinetics of Three Scirpentriol-Based Toxins in Oats Inoculated in Vitro with Isolates of Fusarium sporotrichioides and Fusarium poae

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Substrate, Inoculation, Incubation, Sampling Device
2.2. Analysis of Mycotoxins
2.3. Curve Fitting and Statistics
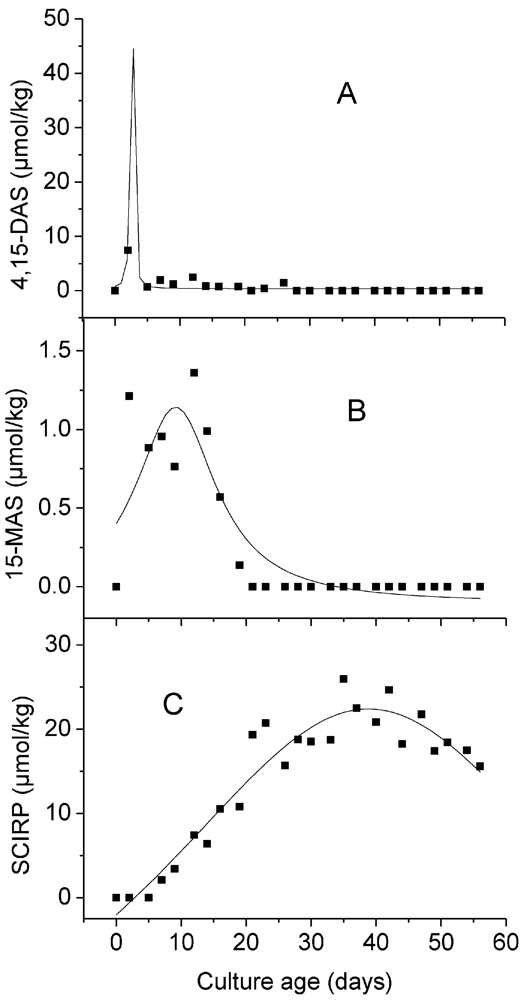
3. Results

4. Discussion
References
- DeNijs, M.; Rombouts, F.M.; Notermans, S.H.W. Fusarium molds and their mycotoxins. J. Food Safety 1996, 16, 15–58. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 2nd ed; Aspen Publishers: Gaithersburg, MD, USA, 1999. [Google Scholar]
- Schollenberger, M.; Drochner, W.; Müller, H.-M. Fusarium toxins of the scirpentriol subgroup: A review. Mycopathologia 2007, 164, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Young, J.C.; Trenholm, H.L. Fusarium toxins in field corn. I. Time course of fungal growth and production of deoxynivalenol and other mycotoxins. Can. J. Bot. 1983, 61, 3080–3087. [Google Scholar]
- Miller, J.D.; Taylor, A.; Greenhalgh, R. Production of deoxynivalenol and related compounds in liquid culture by Fusarium graminearum. Can. J. Microbiol. 1983, 29, 1171–1178. [Google Scholar] [CrossRef]
- Pestka, J.J.; El-Bahrawy, A.; Hart, L.P. Deoxynivalenol and 15-monoacetyldeoxynivalenol production by Fusarium graminearum R6576 in liquid media. Mycopathologia 1985, 91, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Abouzied, M.M.; Pestka, J.J. Fusarenon-X and nivalenol production by Gibberella zeae in liquid and rice cultures. Mycopathologia 1986, 96, 19–24. [Google Scholar] [CrossRef]
- Schuster, M.; Lepschy, J.; Biber, A.; Engelhardt, G.; Wallnöfer, P.R. Production of mycotoxins by Fusarium species isolated in Germany. 2. Time course of deoxynivalenol and 3-acetyldeoxynivalenol formation of Fusarium graminearum in different liquid media. Z. Lebensm. Unters. Forsch. 1987, 185, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.L.; Martins, H.M. Influence of water activity, temperature and incubation time on the simultaneous production of deoxynivalenol and zearalenone in corn (Zea mays) by Fusarium graminearum. Food Chem. 2002, 79, 315–318. [Google Scholar] [CrossRef]
- Bergers, W.W.A.; van der Stap, J.G.M.M.; Kientz, C.E. Trichothecene production in liquid stationary cultures of Fusarium tricinctum NRRL 3299 (Synonym: F. sporotrichioides): Comparison of quantitative brine shrimp assay with physicochemical analysis. Appl. Environ. Microb. 1985, 50, 656–662. [Google Scholar]
- Kotsonis, F.N.; Ellison, R.A. Assay and relationship of HT-2 toxin and T-2 toxin formation in liquid culture. Appl. Microbiol. 1975, 30, 33–37. [Google Scholar]
- Kokkonen, M.; Ojala, L.; Parikka, P.; Jestoi, M. Mycotoxin production of selected Fusarium species at different culture conditions. Int. J. Food Microbiol. 2010, 143, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional oats. Food Addit.Contam. 2009, 26, 1063–1069. [Google Scholar] [CrossRef]
- Lombaert, G.A.; Pellaers, P.; Roscoe, V.; Monkatia, M.; Neil, R.; Scott, P.M. Mycotoxins in infant cereal foods from the Canadian retail market. Food Addit. Contam. 2003, 20, 494–504. [Google Scholar]
- Müller, H.-M.; Reimann, J.; Schumacher, U.; Schwadorf, K. Natural occurrence of Fusarium toxins in oats harvested during five years in an area of southwest Germany. Food Addit. Contam. 1998, 15, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Langseth, W.; Rundberget, T. The occurrence of HT-2 toxin and other trichothecenes in Norwegian cereals. Mycopathologia 1999, 147, 157–165. [Google Scholar]
- Schollenberger, M.; Müller, H.-M.; Rüfle, R.; Suchy, S.; Planck, S.; Drochner, W. Natural occurrence of 16 Fusarium toxins in feedstuffs and mixed feed components from Germany. Mycopathologia 2006, 161, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E. Fusarium mycotoxins. Chemistry Genetics and Biology; APS Press: St Paul, MN, USA, 2006; pp. 185–186. [Google Scholar]
- Stenglein, S.A. Fusarium poae: A pathogen that needs more attention. J. Plant Pathol. 2009, 991, 25–36. [Google Scholar]
- Liu, W.; Sundheim, L.; Langseth, W. Trichothecene production and the relationship to vegetative compatibility groups in Fusarium poae. Mycopathologia 1998, 140, 105–114. [Google Scholar]
- Clear, R.M.; Patrick, S.K.; Platford, R.G.; Desjardins, M. Occurrence and distribution of Fusarium species in barley and oat seeds from Manitoba in 1993 and 1994. Can. J. Plant Pathol. 1996, 18, 409–411. [Google Scholar] [CrossRef]
- Tekauz, A.; McCallum, B.; Ames, N.; Fetch, J.M. Fusarium head blight of oat—current status in western Canada. Can. J. Plant Pathol. 2004, 26, 473–479. [Google Scholar] [CrossRef]
- Schollenberger, M.; Lauber, U.; Terry Jara, H.; Suchy, S.; Drochner, W.; Müller, H.-M. Determination of eight trichothecenes by gas chromatography mass-spectrometry after sample clean-up by a two-stage solid phase extraction. J. Chromatogr. A 1998, 815, 123–132. [Google Scholar]
- Schollenberger, M.; Müller, H.-M.; Rüfle, R.; Suchy, S.; Planck, S.; Drochner, W. Survey of Fusarium toxins in foodstuffs of plant origin marketed in Germany. Int. J. Food Microbiol. 2005, 97, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Sundheim, L.; Langseth, W. Trichothecene production and the relationship to vegetative compatibiliy groups in Fusarium poae. Mycopathologia 1999, 147, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Thrane, U.; Adler, A.; Clasen, P.-E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.; Fog Nielsen, K.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Press, W.H.; Teukolsky, S.A.; Vetterling, W.T.; Flannery, B.P. Numerical Recipes in C++. The Art of Scientific Computing, 3rd ed; University Press: Cambridge, UK, 2007; pp. 686–694. [Google Scholar]
- Desjardins, A.E.; Hohn, T.M.; McCormick, S.P. Effect of gene disruption of trichodienesynthase on the virulence of Gibberella pulicaris. Mol. Plant Microbe 1992, 5, 214–222. [Google Scholar] [CrossRef]
- Park, J.J.; Chu, F.S. Partial purification and characterization of an esterase from Fusarium sporotrichioides. Nat. Toxins 1996, 4, 108–116. [Google Scholar] [CrossRef] [PubMed]
- McCormick, S.P.; Alexander, N.J. Fusarium Tri8 encodes a trichothecene C-3 esterase. Appl. Environ. Microbiol. 2002, 68, 2959–2964. [Google Scholar] [CrossRef] [PubMed]
- Reinbrecht, C.; Miedaner, T.; Geiger, H.H. Changes in deoxynivalenol and nivalenol contents during grain filling and maturity of wheat after inoculation with different isolates of Fusarium culmorum. J. Appl. Genet. 2002, 43, 311–318. [Google Scholar]
- Brinkmeyer, U.; Dänicke, S.; Lehmann, M.; Valenta, H.; Lebzien, P.; Schollenberger, M.; Südekum, K.H.; Weinert, J.; Flachowsky, G. Influence of a Fusarium culmorum inoculation of wheat on the progression of mycotoxin accumulation, ingredient concentrations and ruminal in sacco dry matter degradation of wheat residues. Arch. Anim. Nutr. 2006, 60, 141–157. [Google Scholar] [PubMed]
- Udell, M.N.; Dewick, P.M. Metabolic conversions of trichothecene mycotoxins: De-esterification reactions using cell-free extracts of Fusarium. Z. Naturforsch. C 1989, 44, 660–668. [Google Scholar] [PubMed]
- Yoshizawa, T.; Morooka, N. Comparative studies on microbial and chemical modifications of trichothecene mycotoxins. Appl. Microbiol. 1975, 30, 38–43. [Google Scholar]
- Engelhardt, G.; Ruhland, M.; Wallnöfer, P.R. Metabolism of mycotoxins in plants. Adv. Food Sci. 1999, 21, 71–78. [Google Scholar]
- Alexander, N.J.; McCormick, S.P.; Ziegenhorn, S.L. Phytotoxicity of selected trichothecenes using Chlamydomonas reinhardtii as a model system. Nat. Toxins 1999, 7, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.E.; McCormick, S.P.; Appell, M. Structure-activity relationships of trichothecene toxins in an Arabidopsis thaliana leaf assay. J. Agric. Food. Chem. 2007, 55, 6487–6492. [Google Scholar] [PubMed]
- Thompson, W.L.; Wannemacher, R.W. Structure—Function relationships of 12,13-epoxytrichothecenemycotoxins in cell culture: Comparison to whole animal lethality. Toxicon 1986, 24, 985–994. [Google Scholar]
- Richardson, K.E.; Hamilton, P.B. Comparative toxicity of scirpentriol and its acetylated derivatives. Poultry Sci. 1990, 69, 397–402. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Schollenberger, M.; Müller, H.-M.; Liebscher, M.; Schlecker, C.; Berger, M.; Hermann, W. Accumulation Kinetics of Three Scirpentriol-Based Toxins in Oats Inoculated in Vitro with Isolates of Fusarium sporotrichioides and Fusarium poae. Toxins 2011, 3, 442-452. https://doi.org/10.3390/toxins3050442
Schollenberger M, Müller H-M, Liebscher M, Schlecker C, Berger M, Hermann W. Accumulation Kinetics of Three Scirpentriol-Based Toxins in Oats Inoculated in Vitro with Isolates of Fusarium sporotrichioides and Fusarium poae. Toxins. 2011; 3(5):442-452. https://doi.org/10.3390/toxins3050442
Chicago/Turabian StyleSchollenberger, Margit, Hans-Martin Müller, Melanie Liebscher, Claudia Schlecker, Melanie Berger, and Wilfried Hermann. 2011. "Accumulation Kinetics of Three Scirpentriol-Based Toxins in Oats Inoculated in Vitro with Isolates of Fusarium sporotrichioides and Fusarium poae" Toxins 3, no. 5: 442-452. https://doi.org/10.3390/toxins3050442