Phytosterols and Omega 3 Supplementation Exert Novel Regulatory Effects on Metabolic and Inflammatory Pathways: A Proteomic Study


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
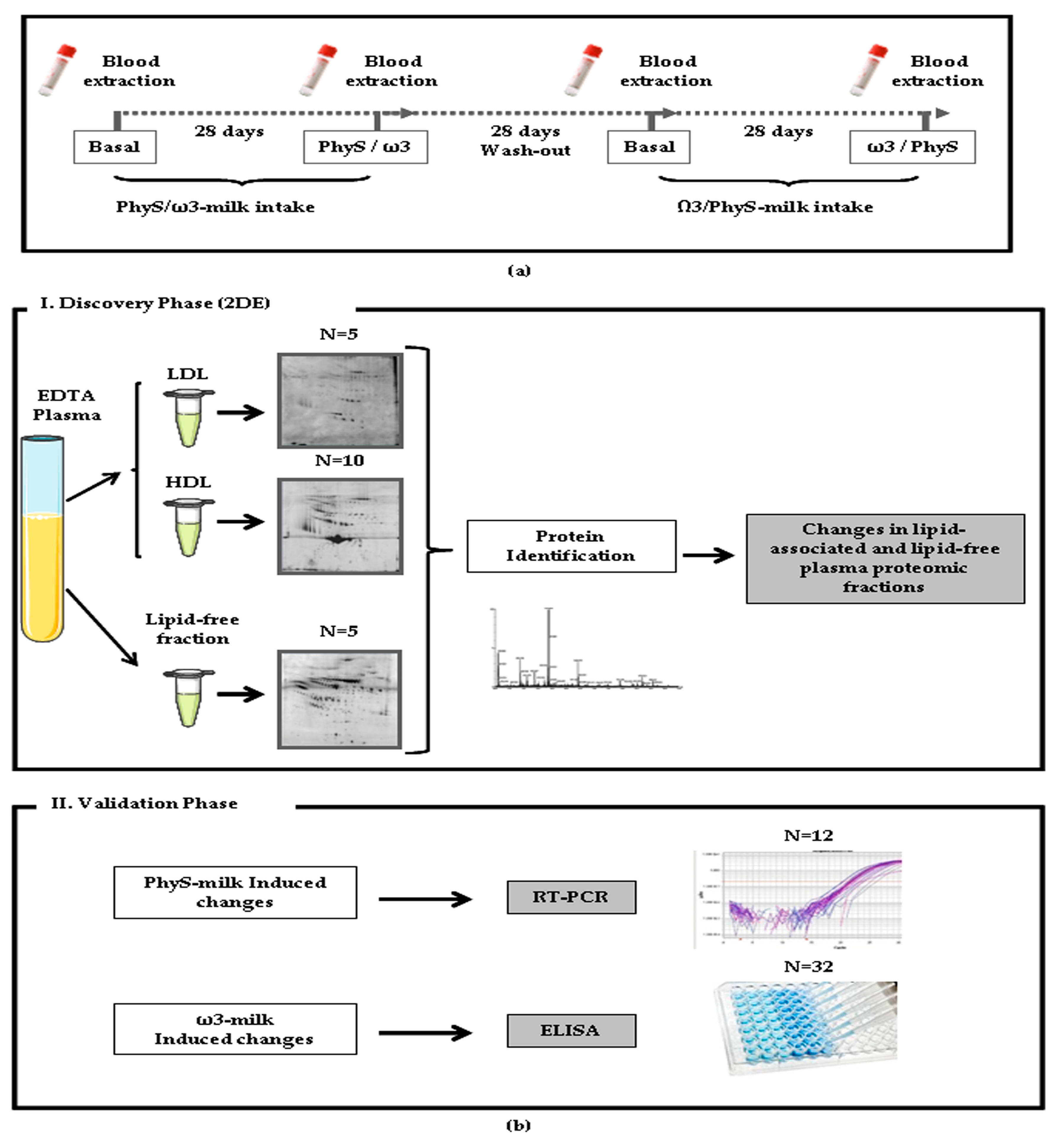
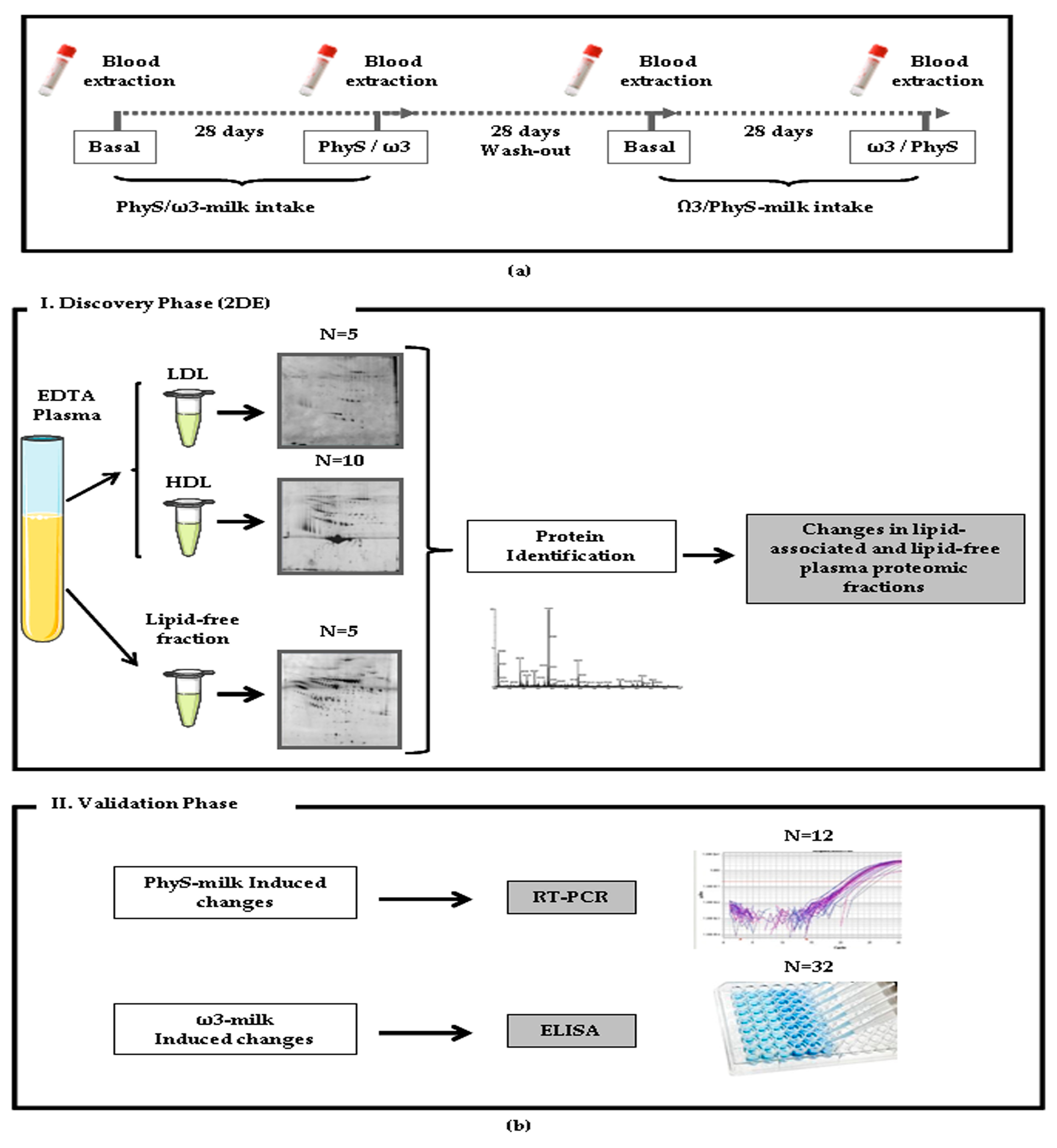
2.2. Study Phases
2.3. Sample Preparation
2.4. Differential Proteomic Profiling Analysis
2.4.1. Two-Dimensional Gel Electrophoresis (2-DE)
2.4.2. Mass Spectrometry Analysis
2.5. Western Blot Analysis
2.6. Quantification of Total Protein Systemic Levels
2.7. Gene Expression Analyses
2.8. Bioinformatic Analysis
2.8.1. Functional Analysis of a Network
2.8.2. Canonical Pathway Analysis
2.9. Statistical Analysis
3. Results
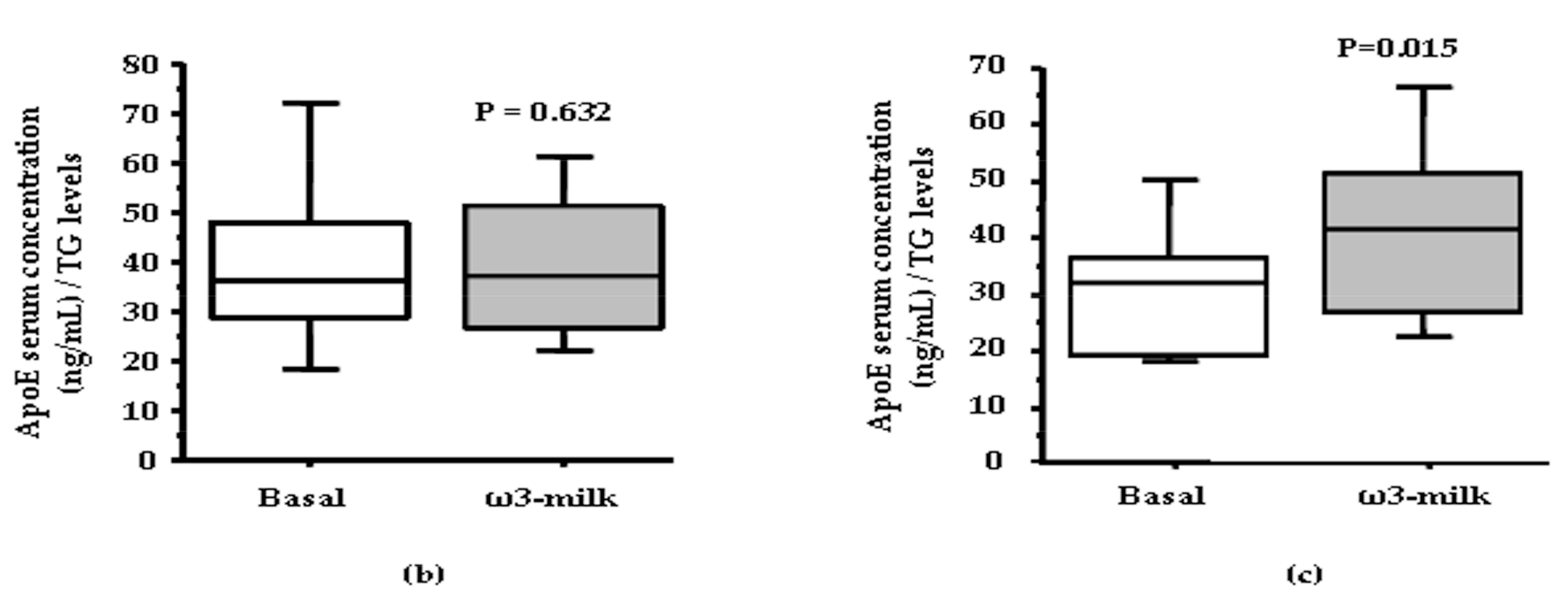
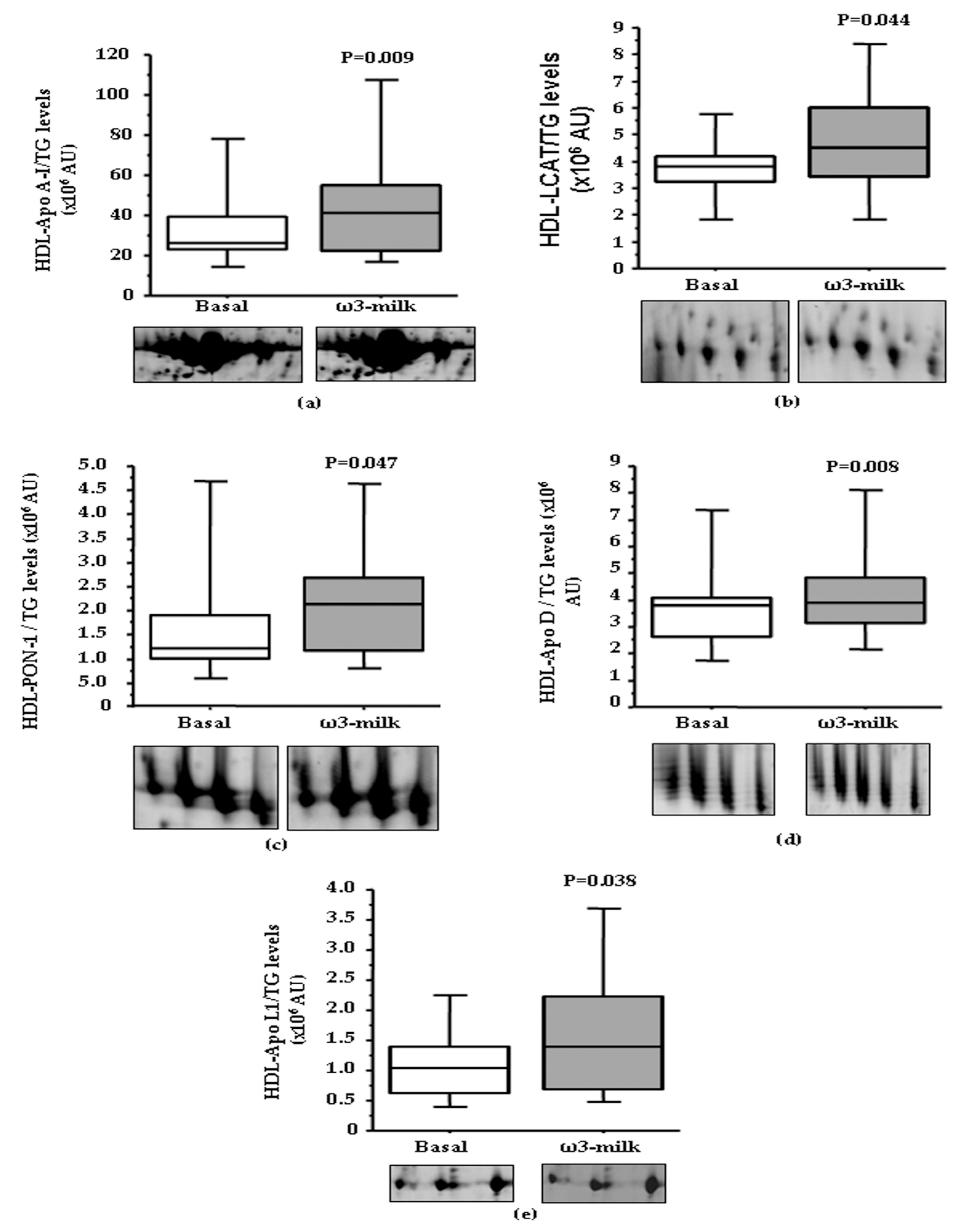
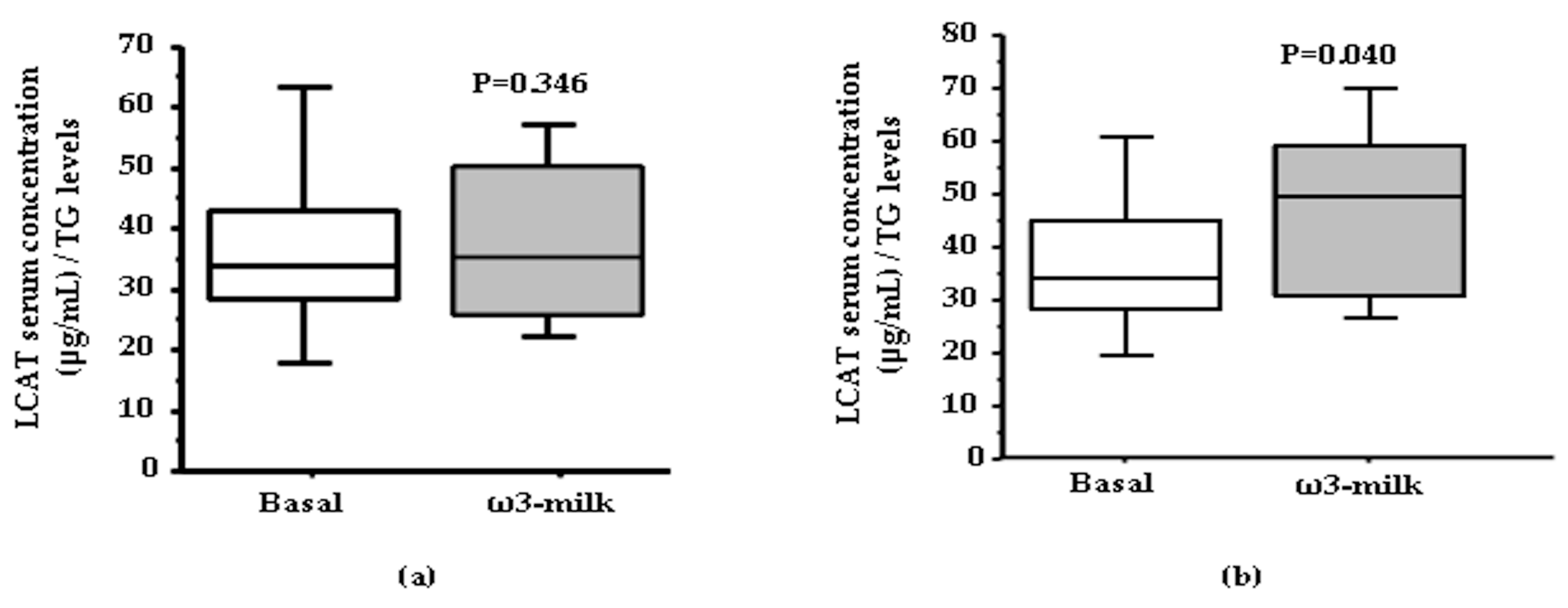
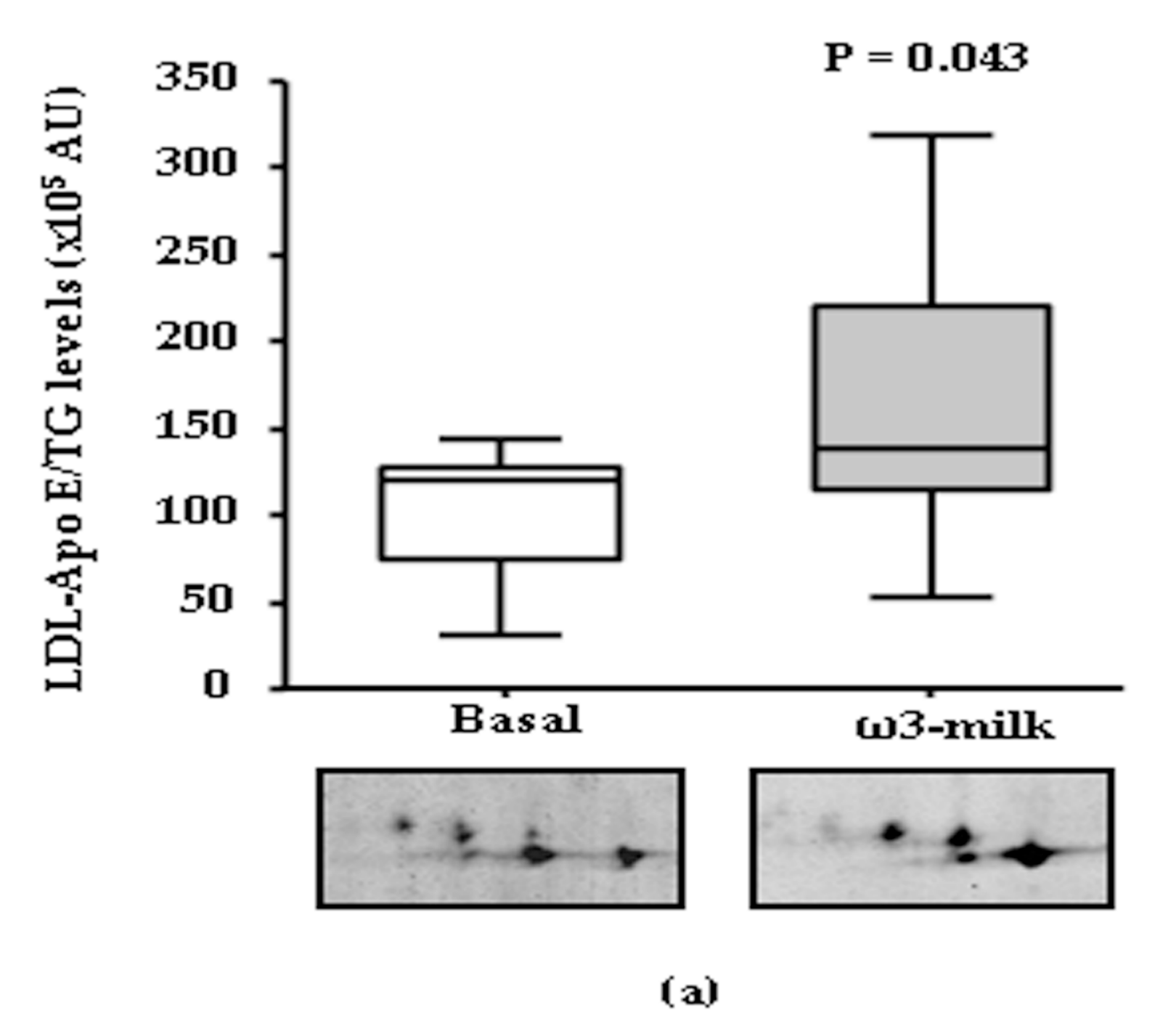
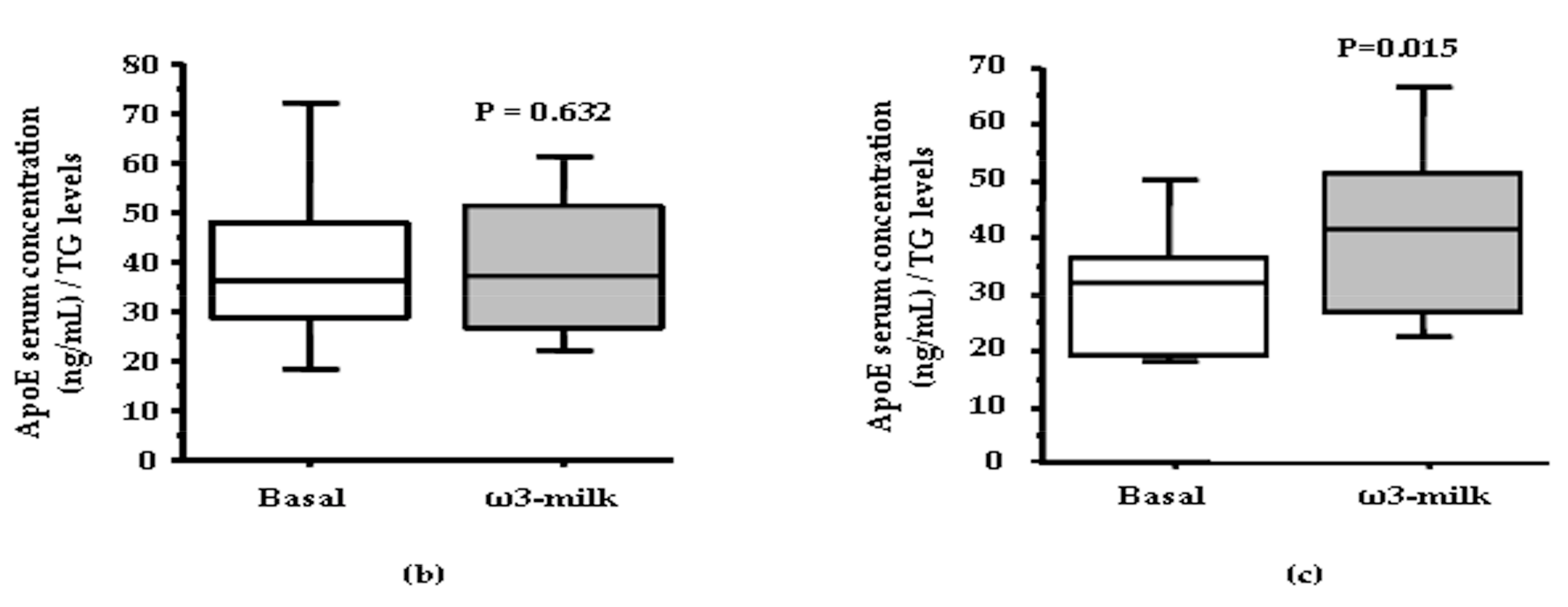
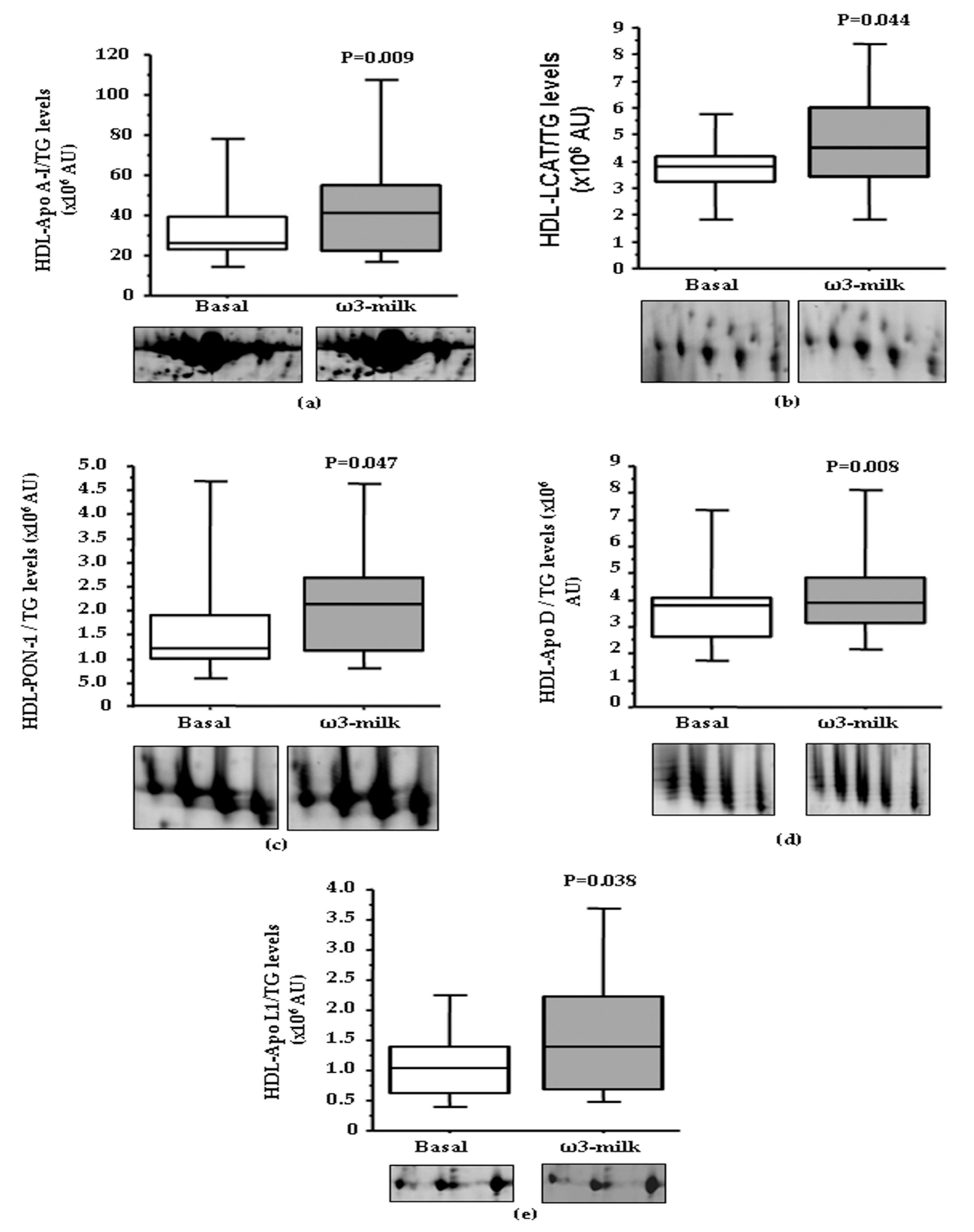
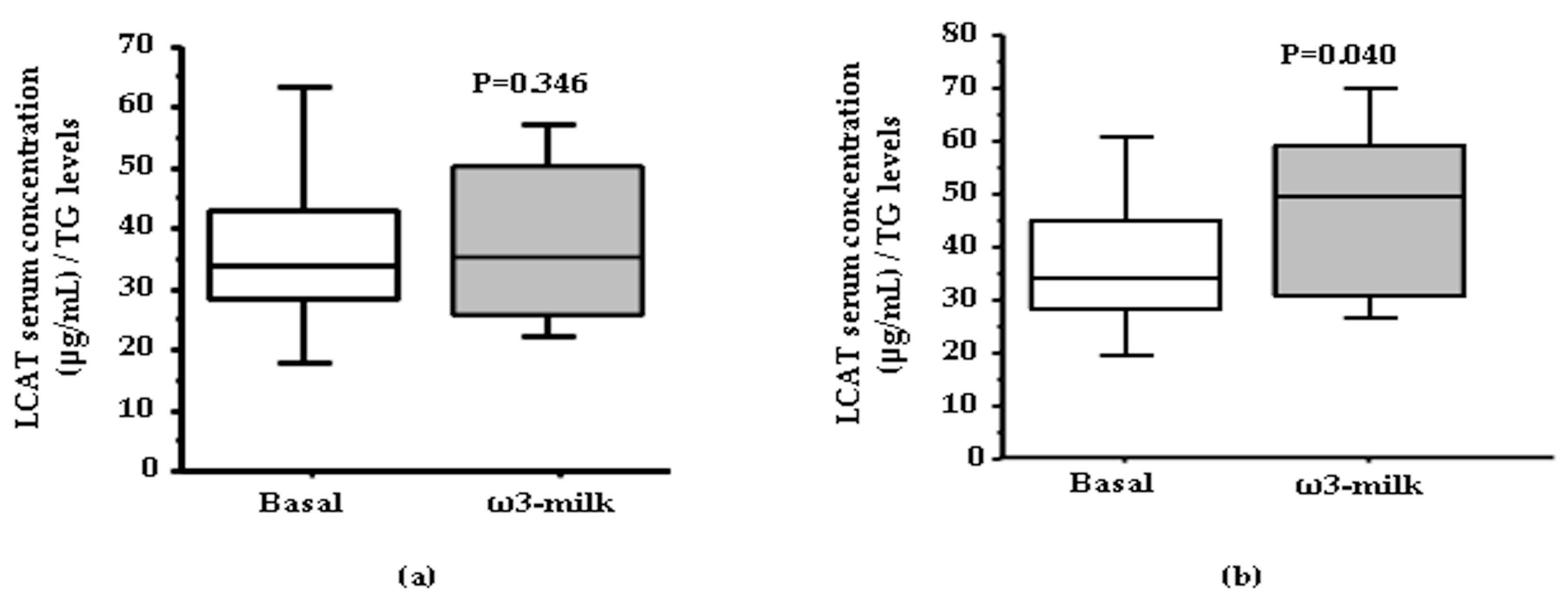
3.1. Four Weeks ω3-Milk Intake Induces Changes in the Proteomic Profile of the Lipoprotein Plasma Fraction
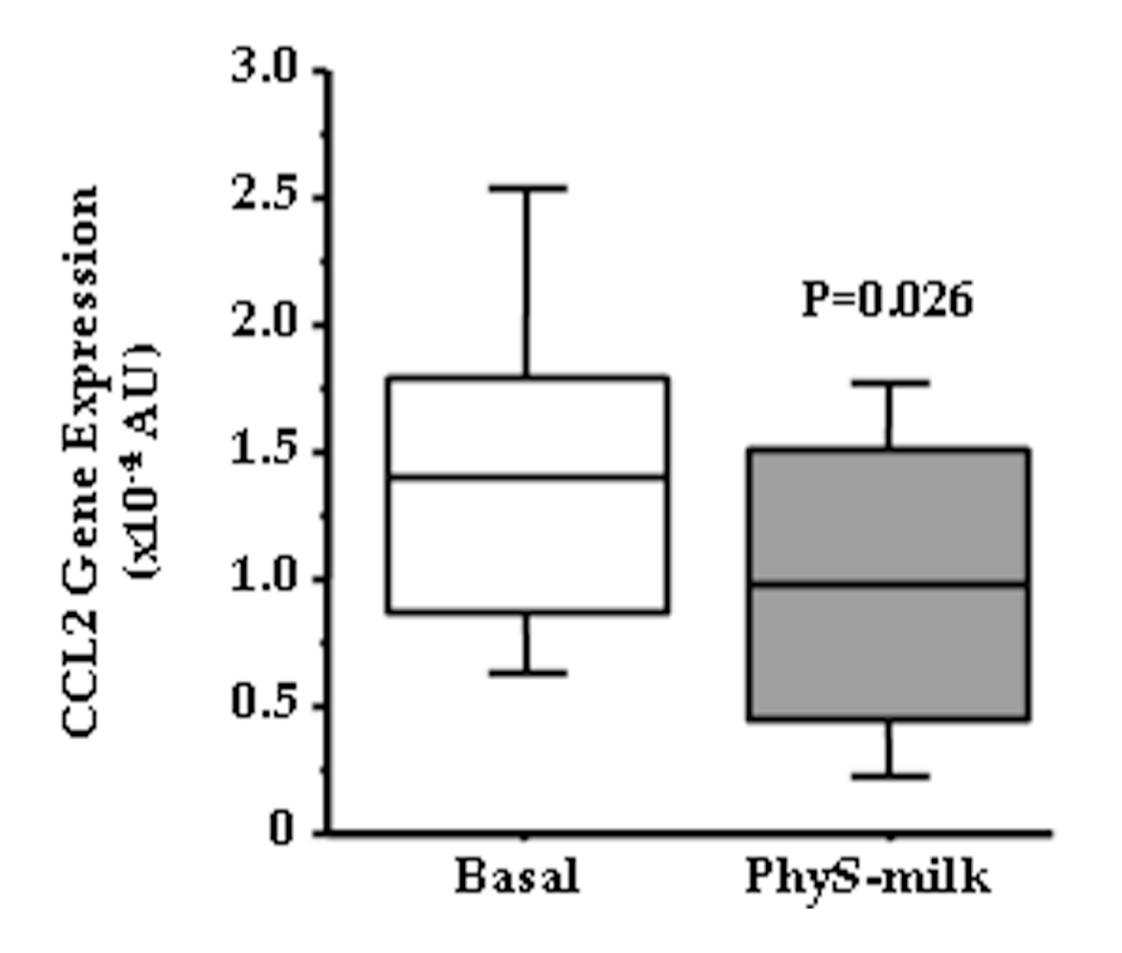
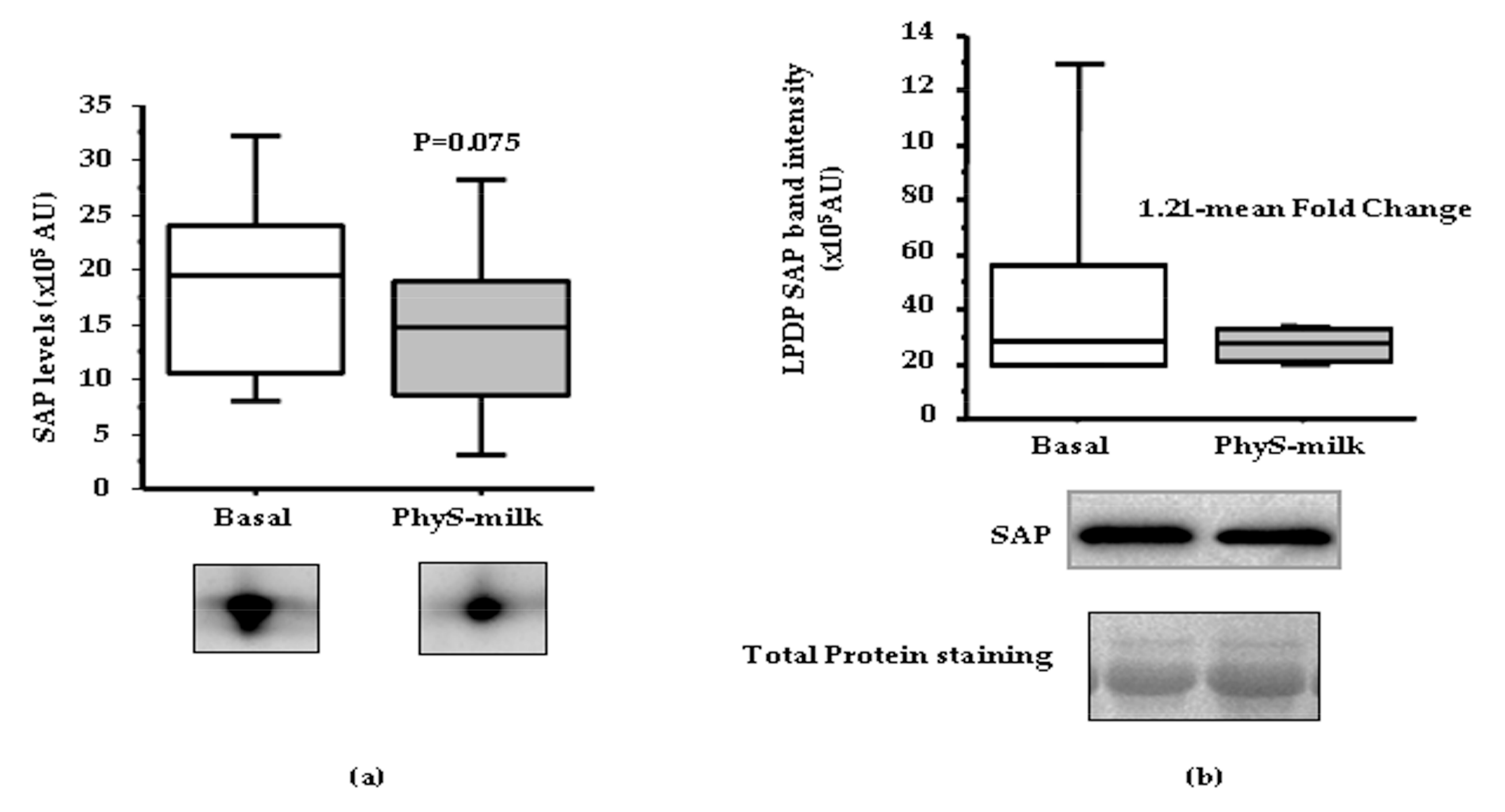
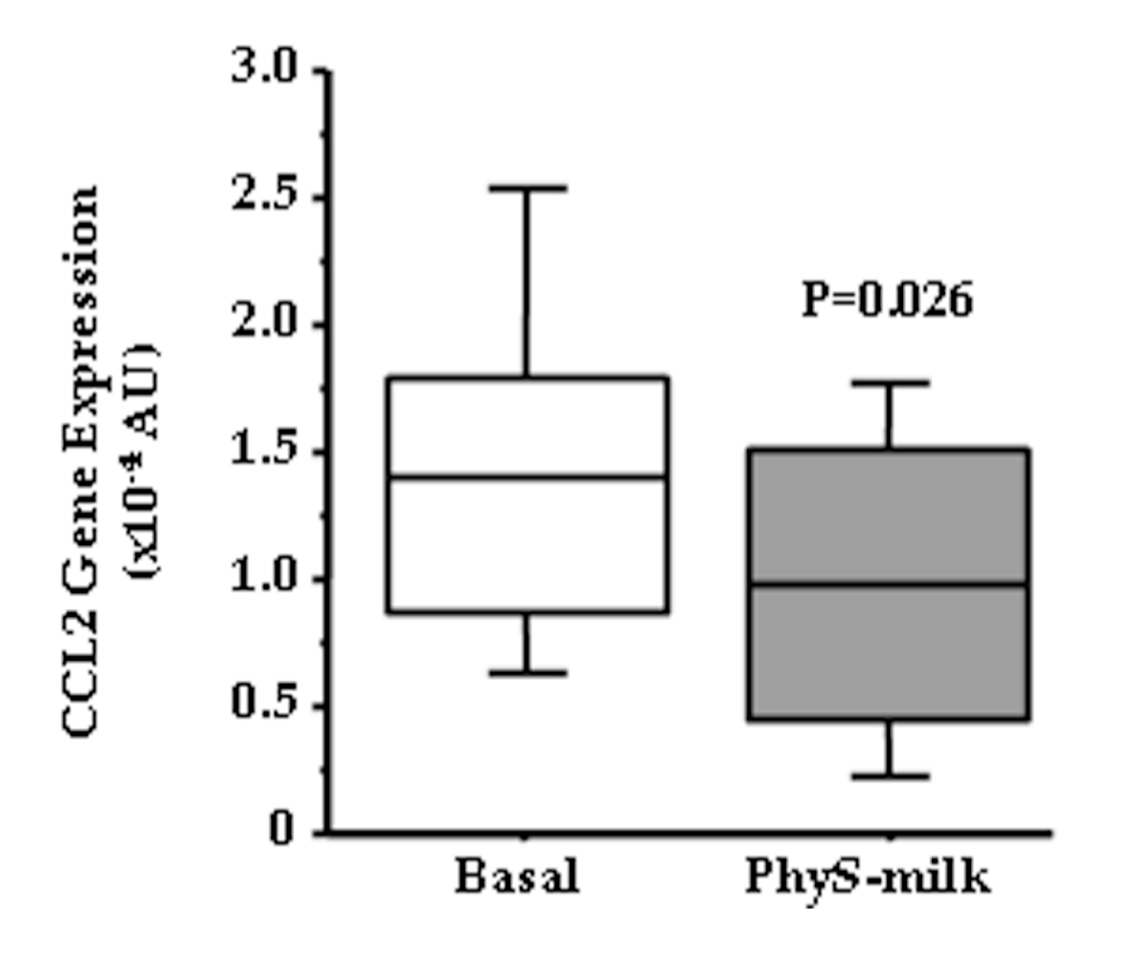
3.2. Inflammation Associated Changes after PhyS-Milk Intake
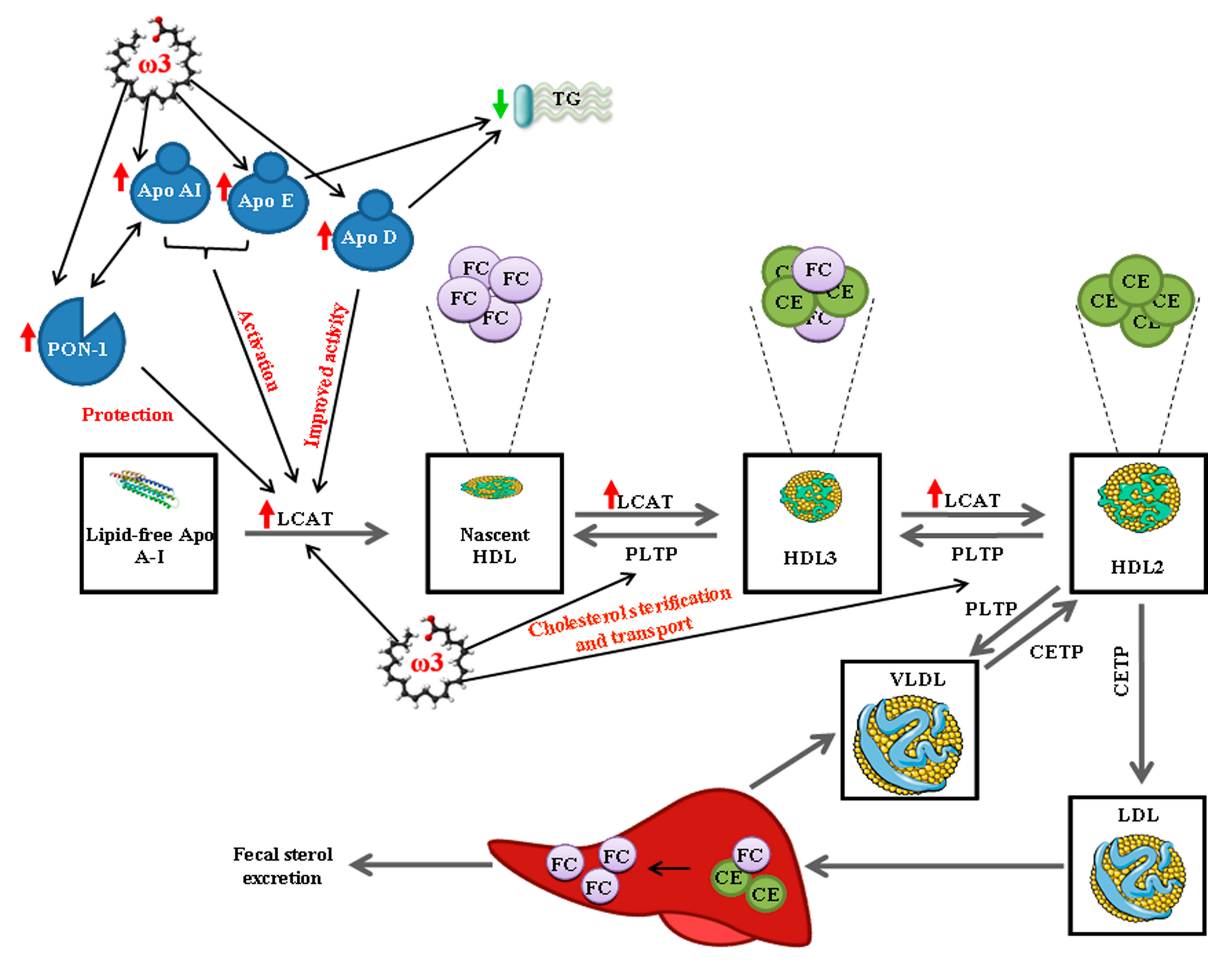
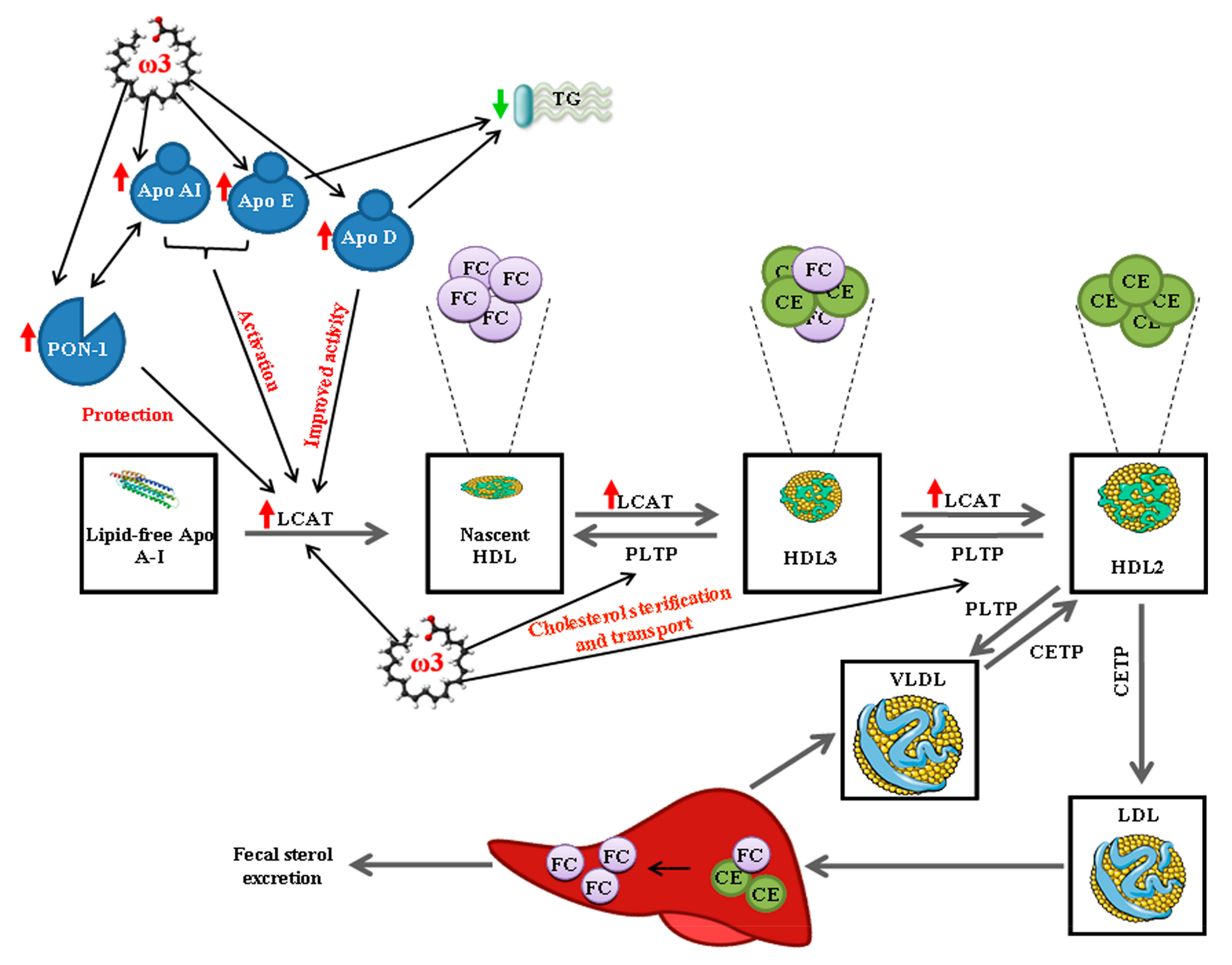
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bastien, M.; Poirier, P.; Lemieux, I.; Despres, J.P. Overview of epidemiology and contribution of obesity to cardiovascular disease. Prog. Cardiovasc. Dis. 2014, 56, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Sayon-Orea, C.; Carlos, S.; Martínez-Gonzalez, M.A. Does cooking with vegetable oils increase the risk of chronic diseases: A systematic review. Br. J. Nutr. 2015, 113, S36–S48. [Google Scholar] [CrossRef] [PubMed]
- Stefler, D.; Malyutina, S.; Kubinova, R.; Pajak, A.; Peasey, A.; Pikhart, H.; Brunner, E.J.; Bobak, M. Mediterranean diet score and total and cardiovascular mortality in Eastern Europe: The HAPIEE study. Eur. J. Nutr. 2015, 56, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G.; Padro, T. Nutraceuticals and atherosclerosis: Human trials. Cardiovasc. Ther. 2010, 28, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Balestrieri, M.L.; Napoli, C. Nutrition, physical activity, and cardiovascular disease: An update. Cardiovasc. Res. 2007, 73, 326–340. [Google Scholar] [CrossRef] [PubMed]
- Psota, T.L.; Gebauer, S.K.; Kris-Etherton, P. Dietary omega-3 fatty acid intake and cardiovascular risk. Am. J. Cardiol. 2006, 98, 3i–18i. [Google Scholar] [CrossRef] [PubMed]
- Padro, T.; Vilahur, G.; Sanchez-Hernandez, J.; Hernandez, M.; Antonijoan, R.M.; Perez, A.; Badimon, L. Lipidomic changes of LDL in overweight and moderately hypercholesterolemic subjects taking phytosterol- and omega-3-supplemented milk. J. Lipid Res. 2015, 56, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Racette, S.B.; Lin, X.; Lefevre, M.; Spearie, C.A.; Most, M.M.; Ma, L.; Ostlund, R.E. Dose effects of dietary phytosterols on cholesterol metabolism: A controlled feeding study. Am. J. Clin. Nutr. 2010, 91, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G. LDL-cholesterol versus HDL-cholesterol in the atherosclerotic plaque: Inflammatory resolution versus thrombotic chaos. Ann. N. Y. Acad. Sci. 2012, 1254, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Vilahur, G.; Gutiérrez, M.; Casaní, L.; Cubedo, J.; Capdevila, A.; Pons-Llado, G.; Carreras, F.; Hidalgo, A.; Badimon, L. Hypercholesterolemia Abolishes High-Density Lipoprotein-Related Cardioprotective Effects in the Setting of Myocardial Infarction. J. Am. Coll. Cardiol. 2015, 66, 2469–2470. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, E.J.; Anthanont, P.; Asztalos, B.F. High-density lipoprotein metabolism, composition, function, and deficiency. Curr. Opin. Lipidol. 2014, 25, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G.; Cubedo, J. High density lipoproteins and kidney function: The friend turned foe? J. Thorac. Dis. 2016, 8, 2978–2981. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G. HDL particles—More complex than we thought. Thromb. Haemost. 2014, 112, 857. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Ren, Y.; Shen, X.; Bian, Y.; Xiao, C.; Li, H. Correlation between the High Density Lipoprotein and its Subtypes in Coronary Heart Disease. Cell. Physiol. Biochem. 2016, 38, 1906–1914. [Google Scholar] [CrossRef] [PubMed]
- Navab, M. Thematic review series: The Pathogenesis of Atherosclerosis: The oxidation hypothesis of atherogenesis: The role of oxidized phospholipids and HDL. J. Lipid Res. 2004, 45, 993–1007. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.F.; Kurpinski, K.; Fang, Q.; Lee, R.J.; Li, S. Proteomic identification of biomarkers of vascular injury. Am. J. Transl. Res. 2011, 3, 139–148. [Google Scholar] [PubMed]
- Kassai, A.; Illyés, L.; Mirdamadi, H.Z.; Seres, I.; Kalmár, T.; Audikovszky, M.; Paragh, G. The effect of atorvastatin therapy on lecithin:cholesterol acyltransferase, cholesteryl ester transfer protein and the antioxidant paraoxonase. Clin. Biochem. 2007, 40, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Hine, D.; MacKness, B.; MacKness, M. Coincubation of PON1, APO A1, and LCAT increases the time HDL is able to prevent LDL oxidation. IUBMB Life 2012, 64, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Daigo, K.; Inforzato, A.; Barajon, I.; Garlanda, C.; Bottazzi, B.; Meri, S.; Mantovani, A. Pentraxins in the activation and regulation of innate immunity. Immunol. Rev. 2016, 274, 202–217. [Google Scholar] [CrossRef] [PubMed]
- Du Clos, T.W. Pentraxins: Structure, function, and role in inflammation. ISRN Inflamm. 2013, 2013, 379040. [Google Scholar] [CrossRef] [PubMed]
- Vilahur, G.; Badimon, L. Biological actions of pentraxins. Vascul. Pharmacol. 2015, 73, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Cubedo, J.; Padró, T.; García-Arguinzonis, M.; Vilahur, G.; Miñambres, I.; Pou, J.M.; Ybarra, J.; Badimon, L. A novel truncated form of apolipoprotein A–I transported by dense LDL is increased in diabetic patients. J. Lipid Res. 2015, 56, 1762–1773. [Google Scholar] [CrossRef] [PubMed]
- Cubedo, J.; Padro, T.; Badimon, L. Glycoproteome of human apolipoprotein A–I: N- and O-glycosylated forms are increased in patients with acute myocardial infarction. Transl. Res. 2014, 164, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Cubedo, J.; Padró, T.; Alonso, R.; Mata, P.; Badimon, L. Apo L1 levels in HDL and cardiovascular event presentation in patients with familial hypercholesterolemia. J. Lipid Res. 2016, 57, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.; Leanderson, P.; Tagesson, C.; Lindahl, M. Lipoproteomics I: Mapping of proteins in low-density lipoprotein using two-dimensional gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Cubedo, J.; Padró, T.; García-Moll, X.; Pintó, X.; Cinca, J.; Badimon, L. Proteomic signature of apolipoprotein J in the early phase of new-onset myocardial infarction. J. Proteom. Res. 2011, 10, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Juni, M.H. OBESITY: A Public Health Threats in Developing Countries. Int. J. Public Health Clin. Sci. 2015, 2, 2289–7577. [Google Scholar]
- Richard, E.; Ostlund, J. Phytosterols in Human Nutrition. Annu. Rev. Nutr. 2002, 22, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Intake, P.; Meijer, L.; Zock, P.L.; Geleijnse, J.M.; Trautwein, E.A. Continuous Dose-Response Relationship of the LDL-Cholesterol—Lowering Effect of Phytosterol Intake. J. Nutr. 2009, 139, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Glueck, C.J.; Speirs, J.; Tracy, T.; Streicher, P.; Illig, E.; Vandegrift, J. Relationships of serum plant sterols (phytosterols) and cholesterol in 595 hypercholesterolemic subjects, and familial aggregation of phytosterols, cholesterol, and premature coronary heart disease in hyperphytosterolemic probands and their first-degree relatives. Metabolism 1991, 40, 842–848. [Google Scholar] [PubMed]
- Assmann, G.; Cullen, P.; Erbey, J.; Ramey, D.R.; Kannenberg, F.; Schulte, H. Plasma sitosterol elevations are associated with an increased incidence of coronary events in men: Results of a nested case-control analysis of the Prospective Cardiovascular Munster (PROCAM) study. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Sudhop, T.; Gottwald, B.M.; von Bergmann, K. Serum plant sterols as a potential risk factor for coronary heart disease. Metabolism 2002, 51, 1519–1521. [Google Scholar] [CrossRef] [PubMed]
- Weingärtner, O.; Böhm, M.; Laufs, U. Controversial role of plant sterol esters in the management of hypercholesterolaemia. Eur. Heart J. 2009, 30, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Bottazzi, B.; Inforzato, A.; Messa, M.; Barbagallo, M.; Magrini, E.; Garlanda, C.; Mantovani, A. The pentraxins PTX3 and SAP in innate immunity, regulation of inflammation and tissue remodelling. J. Hepatol. 2016, 64, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.A.; Hatanaka, K.; Ishibashi-Ueda, H.; Yutani, C.; Yamamoto, A. Characterization of Serum Amyloid P Component From Human Aortic Atherosclerotic Lesions. Arter. Thromb. Vasc. Biol. 1995, 15, 252–257. [Google Scholar] [CrossRef]
- Cubedo, J.; Padró, T.; Badimon, L. Coordinated proteomic signature changes in immune response and complement proteins in acute myocardial infarction: The implication of serum amyloid P-component. Int. J. Cardiol. 2013, 168, 5196–5204. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Wu, T.; Zeng, C.; Li, X.; Li, X.; Wen, D.; Ji, T.; Lan, T.; Xing, L.; Li, J.; et al. SAP deficiencymitigated atherosclerotic lesions in ApoE−/− mice. Atherosclerosis 2016, 244, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Xi, D.; Luo, T.; Xiong, H.; Liu, J.; Lu, H.; Li, M.; Hou, Y.; Guo, Z. SAP: Structure, function, and its roles in immune-related diseases. Int. J. Cardiol. 2015, 187, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Kawano, Y.; Imamura, T.; Kawakita, K.; Sagara, M.; Matsuo, T.; Kakitsubata, Y.; Ishikawa, T.; Kitamura, K.; Hatakeyama, K.; et al. Reciprocal Contribution of Pentraxin 3 and C-Reactive Protein to Obesity and Metabolic Syndrome. Obesity 2010, 18, 1871–1874. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Panee, J. Monocyte Chemoattractant Protein 1 (MCP-1) in obesity and diabetes. Cytokine 2012, 60, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Cubedo, J. Adipose tissue depots and inflammation: Effects on plasticity and resident mesenchymal stem cell function. Cardiovasc. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wensveen, F.M.; Valentić, S.; Šestan, M.; Turk Wensveen, T.; Polić, B. The “Big Bang” in obese fat: Events initiating obesity-induced adipose tissue inflammation. Eur. J. Immunol. 2015, 45, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Han, M.S.; Jung, D.Y.; Morel, C.; Lakhani, S.A.; Kim, J.K.; Flavell, R.A.; Davis, R.J. JNK Expression by Macrophages Promotes Obesity-induced Insulin Resistance and Inflammation. Science 2013, 339, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.W.; De Waal Malefyt, R.; Coffman, R.L.; O’garra, A. Interleukin-10 and the Interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Steffen, L.M.; Steffen, B.T.; Guan, W.; Weir, N.L.; Rich, S.S.; Manichaikul, A.; Vargas, J.D.; Tsai, M.Y. APOE genotype modifies the association between plasma omega-3 fatty acids and plasma lipids in the Multi-Ethnic Study of Atherosclerosis (MESA). Atherosclerosis 2013, 228, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.B.; Huang, L.L.; Rong, R.; Tan, R.; Wang, J.; Kang, J.X. Endogenously decreasing tissue n-6/n-3 fatty acid ratio reduces atherosclerotic lesions in apolipoprotein E-deficient mice by inhibiting systemic and vascular inflammation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2487–2494. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Snook, J.T.; Park, S.; Williams, G.; Tsai, Y.-H.; Lee, N. Effect of synthetic triglycerides of myristic, palmitic, and stearic acid on serum lipoprotein metabolism. Eur. J. Clin. Nutr. 1999, 53, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.L.; Zhu, X.; Rong, S.; Shewale, S.; Seo, J.; Boudyguina, E.; Gebre, A.K.; Alexander-Miller, M.A.; Parks, J.S. Omega-3 fatty acids ameliorate atherosclerosis by favorably altering monocyte subsets and limiting monocyte recruitment to aortic lesions. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Conway, V.; Allard, M.; Minihane, A.; Jackson, K.G.; Lovegrove, J.A.; Plourde, M. Postprandial enrichment of triacylglycerol-rich lipoproteins with omega-3 fatty acids: Lack of an interaction with apolipoprotein E genotype? Lipids Health Dis. 2014, 13, 148. [Google Scholar] [CrossRef] [PubMed]
- Reilly, M.; Rader, D.J. Apolipoprotein E and coronary disease: A puzzling paradox. PLoS Med. 2006, 3, 736–738. [Google Scholar] [CrossRef] [PubMed]
- Chouinard-watkins, R.; Plourde, M. Fatty Acid Metabolism in Carriers of Apolipoprotein E Epsilon 4 Allele: Is It Contributing to Higher Risk of Cognitive Decline and Coronary Heart Disease? Nutrients 2014, 6, 4452–4471. [Google Scholar] [CrossRef] [PubMed]
- Kaneva, A.M.; Bojko, E.R.; Potolitsyna, N.N.; Odland, J.O. Plasma levels of apolipoprotein-E in residents of the European North of Russia. Lipids Health Dis. 2013, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Daniels, J.-A.; Mulligan, C.; McCance, D.; Woodside, J.V.; Patterson, C.; Young, I.S.; McEneny, J. A randomised controlled trial of increasing fruit and vegetable intake and how this influences the carotenoid concentration and activities of PON-1 and LCAT in HDL from subjects with type 2 diabetes. Cardiovasc. Diabetol. 2014, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- White, C.R.; Garber, D.W.; Anantharamaiah, G.M. Anti-inflammatory and cholesterol-reducing properties of apolipoprotein mimetics: A review. J. Lipid Res. 2014, 55, 2007–2021. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Menini, T. Paraoxonase 1 and HDL maturation. Clin. Chim. Acta 2015, 439, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Rousset, X.; Vaisman, B.; Amar, M.; Sethi, A.A.; Remaley, A.T. Lecithin: cholesterol acyltransferase: From biochemistry to role in cardiovascular disease. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Ali, K.; Abo-Ali, E.M.; Kabir, M.D.; Riggins, B.; Nguy, S.; Li, L.; Srivastava, U.; Thinn, S.M.M. A Western-fed diet increases plasma HDL and LDL-cholesterol levels in ApoD−/− mice. PLoS ONE 2014, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Product | PhyS-Milk | ω3-Milk | ||
|---|---|---|---|---|
| Women/Men | 19/13 | 19/13 | ||
| Age (Years) | 50.5 ± 1.6 | 50.5 ± 1.6 | ||
| Parameter | Baseline | After PhyS-milk | Baseline | After ω3-milk |
| BMI | 28.2 ± 0.7 | 28.1 ± 0.7 | 28.3 ± 0.7 | 28.2 ± 0.7 |
| Ch (mg/dL) | 216.0 ± 6.0 | 204.5 ± 5.6 * | 216.4 ± 6.1 | 213.78 ± 5.9 |
| TG (mg/dL) | 110.2 ± 10.3 | 115.2 ±15.1 | 116.3 ± 14.3 | 99.5 ± 8.7 * |
| HDL-C (mg/dL) | 54.5 ± 3.1 | 54.5 ± 3.0 | 57.0 ± 2.9 | 56.4 ± 2.8 |
| LDL-C (mg/dL) | 137.7 ± 4.9 | 127.2 ± 4.7 * | 136.4 ± 5.0 | 137.7 ± 5.0 |
| VLDL-C (mg/dL) | 22.0 ± 2.0 | 23.0 ± 3.0 | 23.2 ± 2.9 | 19.8 ± 1.7 * |
| Non-HDL-C (mg/dL) | 159.5 ± 5.8 | 150.0 ± 5.8 * | 159.4 ± 6.0 | 157.3 ± 6.0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lambert, C.; Cubedo, J.; Padró, T.; Sánchez-Hernández, J.; Antonijoan, R.M.; Perez, A.; Badimon, L. Phytosterols and Omega 3 Supplementation Exert Novel Regulatory Effects on Metabolic and Inflammatory Pathways: A Proteomic Study. Nutrients 2017, 9, 599. https://doi.org/10.3390/nu9060599
Lambert C, Cubedo J, Padró T, Sánchez-Hernández J, Antonijoan RM, Perez A, Badimon L. Phytosterols and Omega 3 Supplementation Exert Novel Regulatory Effects on Metabolic and Inflammatory Pathways: A Proteomic Study. Nutrients. 2017; 9(6):599. https://doi.org/10.3390/nu9060599
Chicago/Turabian StyleLambert, Carmen, Judit Cubedo, Teresa Padró, Joan Sánchez-Hernández, Rosa M. Antonijoan, Antonio Perez, and Lina Badimon. 2017. "Phytosterols and Omega 3 Supplementation Exert Novel Regulatory Effects on Metabolic and Inflammatory Pathways: A Proteomic Study" Nutrients 9, no. 6: 599. https://doi.org/10.3390/nu9060599