Green Tea Polyphenols, Mimicking the Effects of Dietary Restriction, Ameliorate High-Fat Diet-Induced Kidney Injury via Regulating Autophagy Flux

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Cell Culture
2.4. Morphologic and Red O Oil Staining
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
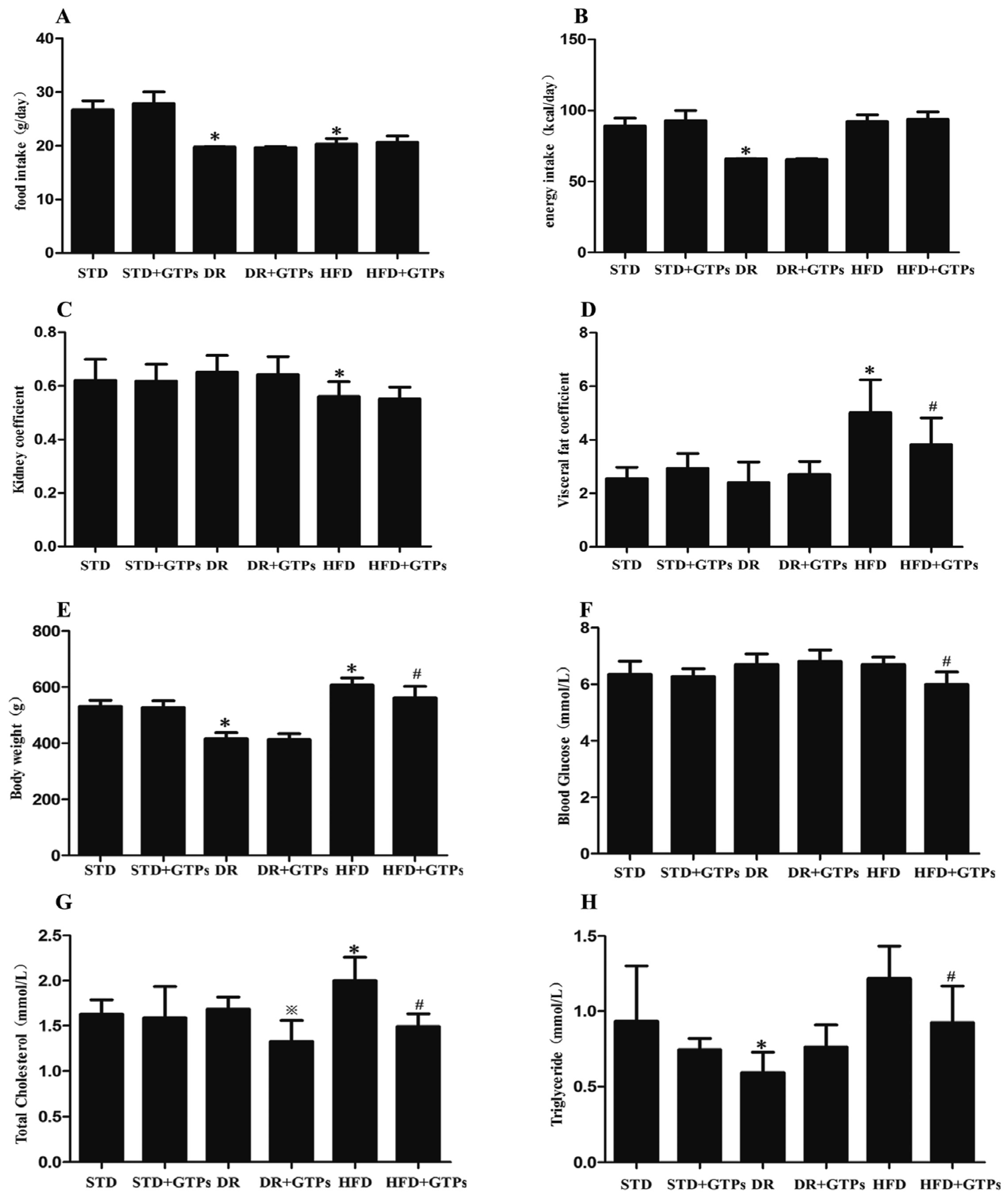
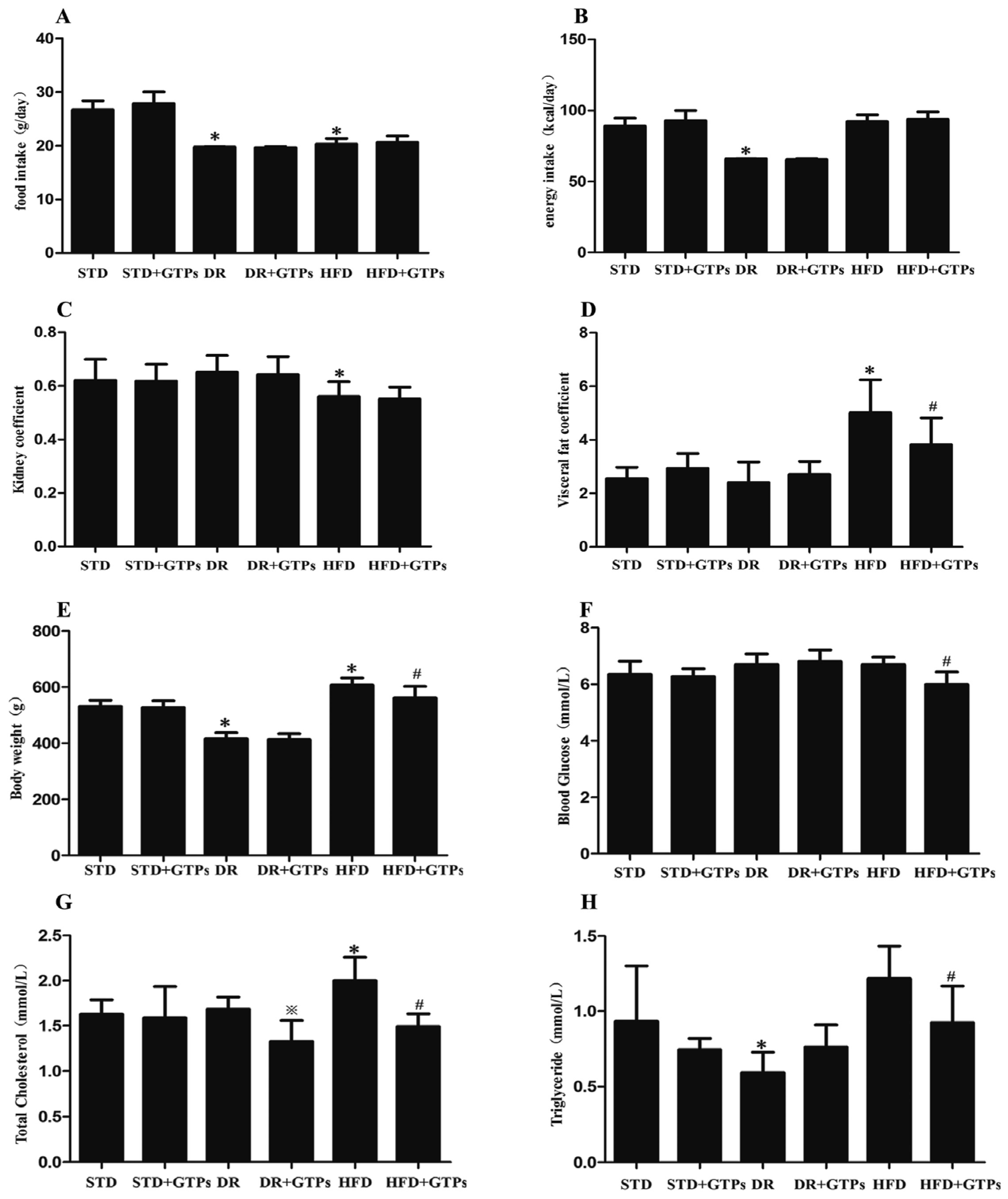
3.1. Body Weight, Blood Glucose and Serum Lipid Levels in Different Diet Groups
3.2. Renal Function in Different Diet Groups
3.3. Autophagy Activity of Kidney in Different Diet Groups
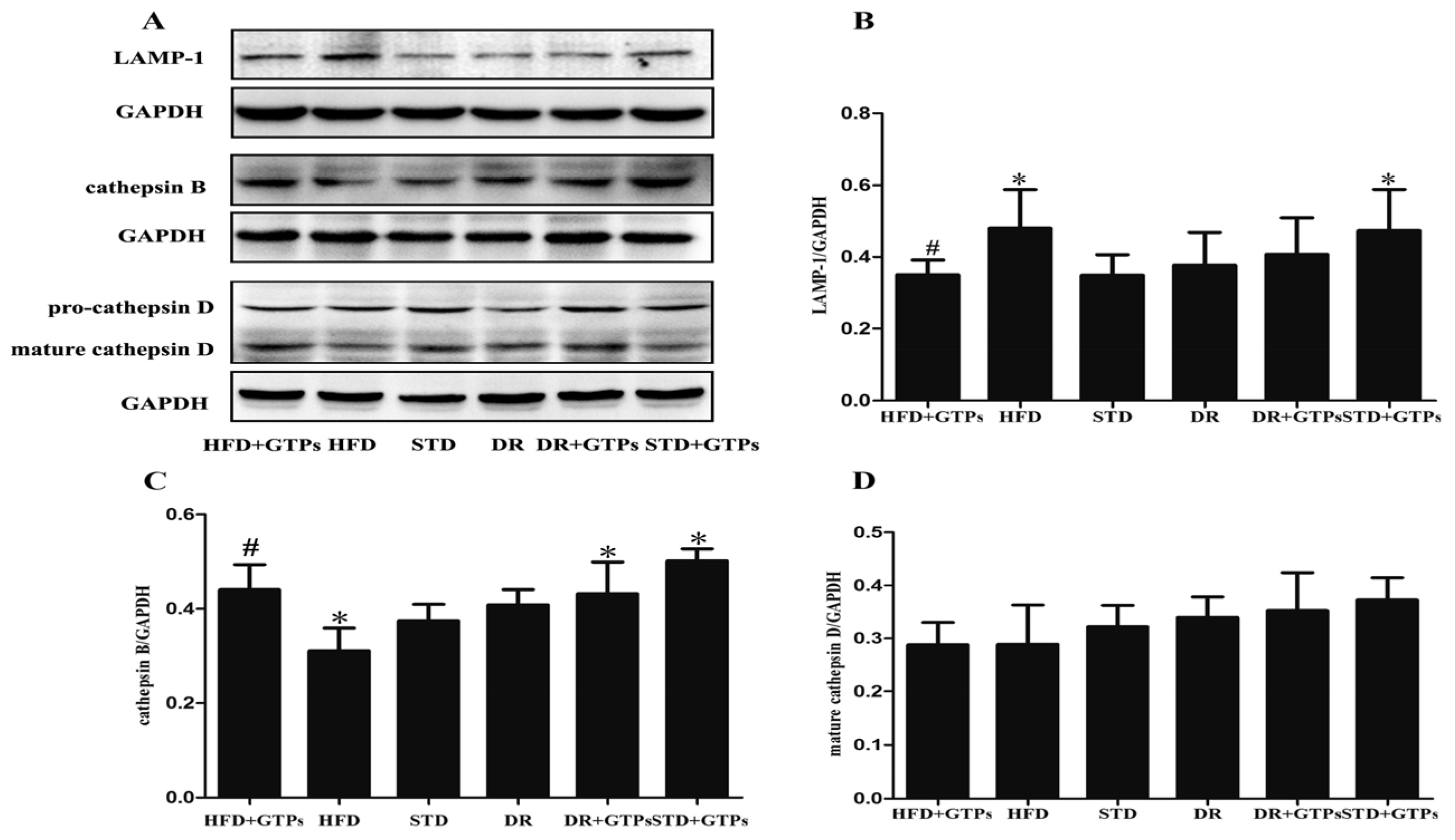
3.4. Lysosome Function of Kidney in Different Diet Groups
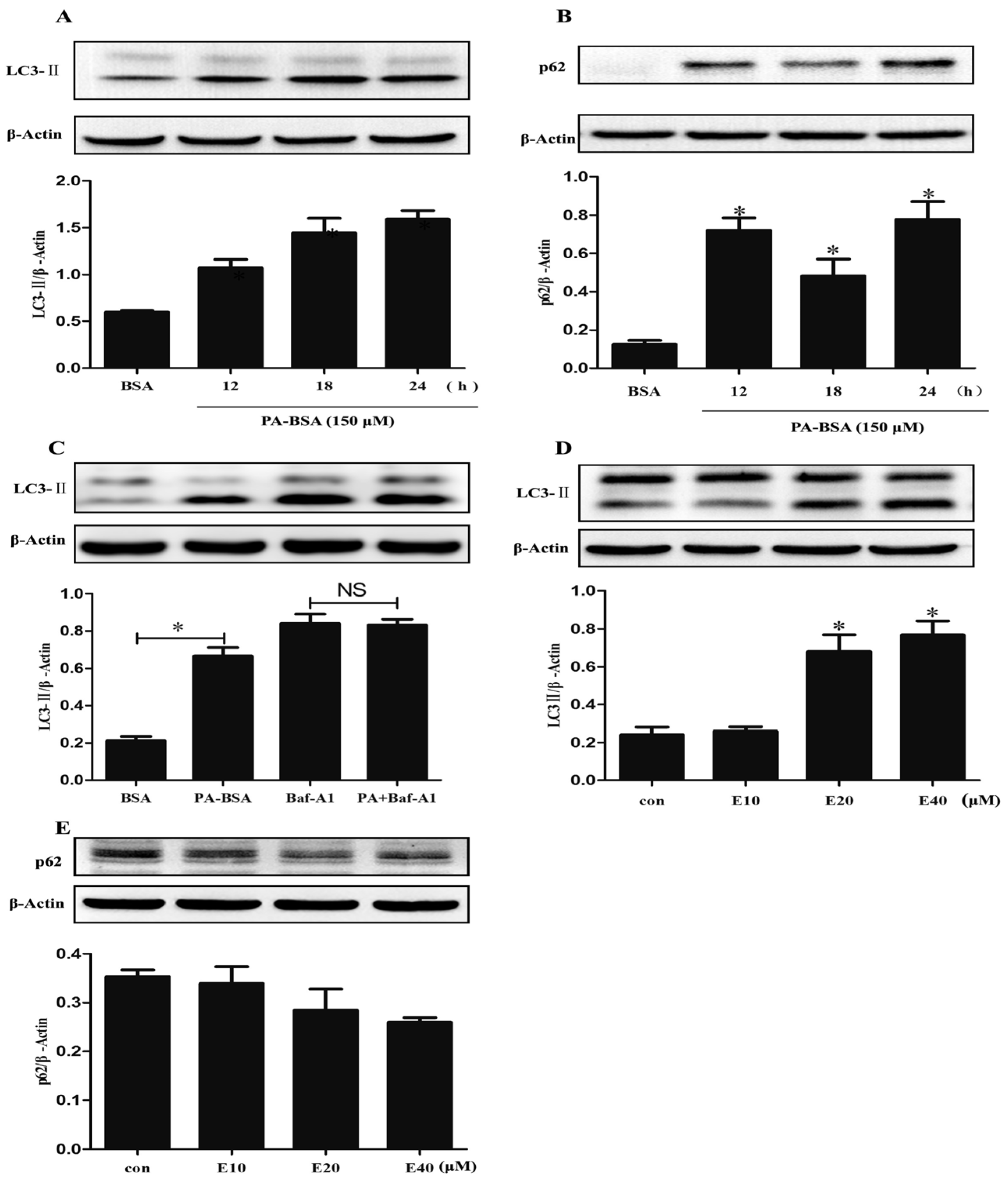
3.5. EGCG Enhanced Autophagic Flux and Ameliorated PA-Induced Autophagy Impairment in HK-2 Cells
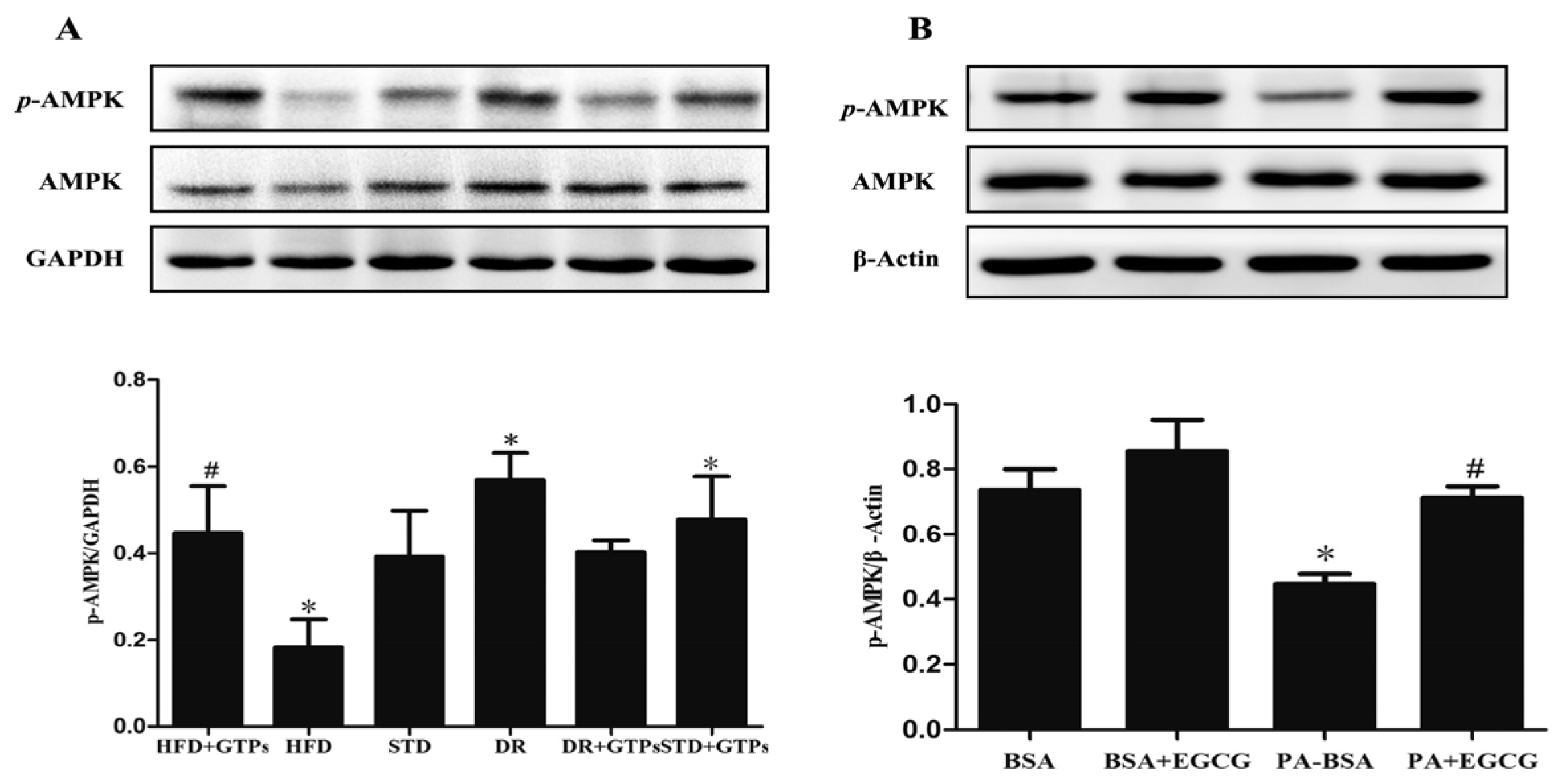
3.6. GTPs and EGCG Stimulated AMPK Activity In Vivo and In Vitro, Respectively
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, L.; Wang, F.; Wang, L.; Wang, W.; Liu, B.; Liu, J.; Chen, M.; He, Q.; Liao, Y.; Yu, X.; et al. Prevalence of chronic kidney disease in china: A cross-sectional survey. Lancet 2012, 379, 815–822. [Google Scholar] [CrossRef]
- Odermatt, A. The western-style diet: A major risk factor for impaired kidney function and chronic kidney disease. Am. J. Physiol. Renal. Physiol. 2011, 301, F919–F931. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Fung, T.T.; Hu, F.B.; Curhan, G.C. Association of dietary patterns with albuminuria and kidney function decline in older white women: A subgroup analysis from the nurses’ health study. Am. J. Kidney Dis. 2011, 57, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wildman, R.P.; Gu, D.; Kusek, J.W.; Spruill, M.; Reynolds, K.; Liu, D.; Hamm, L.L.; Whelton, P.K.; He, J. Prevalence of decreased kidney function in Chinese adults aged 35 to 74 years. Kidney Int. 2005, 68, 2837–2845. [Google Scholar] [CrossRef] [PubMed]
- Decleves, A.E.; Mathew, A.V.; Cunard, R.; Sharma, K. Ampk mediates the initiation of kidney disease induced by a high-fat diet. J. Am. Soc. Nephrol. 2011, 22, 1846–1855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yu, Y.; Deng, J.; Zhang, C.; Zhang, J.; Cheng, Y.; Luo, X.; Han, B.; Yang, H. Sesamin ameliorates high-fat diet–induced dyslipidemia and kidney injury by reducing oxidative stress. Nutrients 2016, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Aydin, C.; Ince, E.; Koparan, S.; Cangul, I.T.; Naziroglu, M.; Ak, F. Protective effects of long term dietary restriction on swimming exercise-induced oxidative stress in the liver, heart and kidney of rat. Cell Biochem. Funct. 2007, 25, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Maddox, D.A.; Alavi, F.K.; Santella, R.N.; Zawada, E.T., Jr. Prevention of obesity-linked renal disease: Age-dependent effects of dietary food restriction. Kidney Int. 2002, 62, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Kapahi, P.; Kaeberlein, M.; Hansen, M. Dietary restriction and lifespan: Lessons from invertebrate models. Ageing Res. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Choi, M.E. Autophagy in kidney health and disease. Antioxid. Redox Signal. 2014, 20, 519–537. [Google Scholar] [CrossRef] [PubMed]
- De Rechter, S.; Decuypere, J.P.; Ivanova, E.; van den Heuvel, L.P.; De Smedt, H.; Levtchenko, E.; Mekahli, D. Autophagy in renal diseases. Pediatr. Nephrol. 2016, 31, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Yamahara, K.; Kume, S.; Koya, D.; Tanaka, Y.; Morita, Y.; Chin-Kanasaki, M.; Araki, H.; Isshiki, K.; Araki, S.; Haneda, M.; et al. Obesity-mediated autophagy insufficiency exacerbates proteinuria-induced tubulointerstitial lesions. J. Am. Soc. Nephrol. 2013, 24, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, J.; Zhen, J.; Zhang, C.; Wan, Q.; Liu, G.; Wei, X.; Zhang, Y.; Wang, Z.; Han, H.; et al. Histone deacetylase 4 selectively contributes to podocyte injury in diabetic nephropathy. Kidney Int. 2014, 86, 712–725. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Lee, J.; Seo, H.Y.; Lim, J.S.; Kim, E.K. Cathepsin inhibition-induced lysosomal dysfunction enhances pancreatic beta-cell apoptosis in high glucose. PLoS ONE 2015, 10, e0116972. [Google Scholar] [CrossRef] [PubMed]
- Lamore, S.D.; Wondrak, G.T. Autophagic-lysosomal dysregulation downstream of cathepsin b inactivation in human skin fibroblasts exposed to uva. Photochem. Photobiol. Sci. 2012, 11, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Repnik, U.; Stoka, V.; Turk, V.; Turk, B. Lysosomes and lysosomal cathepsins in cell death. Biochim. Biophys. Acta 2012, 1824, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, H.; Takemura, G.; Maruyama, R.; Goto, K.; Tsujimoto, A.; Ogino, A.; Li, L.; Kawamura, I.; Takeyama, T.; Kawaguchi, T.; et al. Functional significance and morphological characterization of starvation-induced autophagy in the adult heart. Am. J. Pathol. 2009, 174, 1705–1714. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Takeda, A.; Nagai, T.; Ito, H.; Kanasaki, K.; Koya, D. Dietary restriction ameliorates diabetic nephropathy through anti-inflammatory effects and regulation of the autophagy via restoration of sirt1 in diabetic wistar fatty (fa/fa) rats: A model of type 2 diabetes. Exp. Diabetes Res. 2011, 2011, 908185. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D. Does tea prevent cancer? Evidence from laboratory and human intervention studies. Am. J. Clin. Nutr. 2013, 98, 1667S–1675S. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Cao, C.; Cheng, Y.; Qin, X.Y. Green tea polyphenols attenuated glutamate excitotoxicity via antioxidative and antiapoptotic pathway in the primary cultured cortical neurons. Oxid. Med. Cell. Longev. 2016, 2016, 2050435. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Zhang, Y.; Zhang, L.; Li, X.; Yu, J.B.; Zhang, H.T.; Tan, B.B.; Jiang, L.H.; Wang, Y.X.; Liang, Y.; et al. Protective effect of tea polyphenols on renal ischemia/reperfusion injury via suppressing the activation of tlr4/nf-kappab p65 signal pathway. Gene 2014, 542, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Kuriyama, S.; Shimazu, T.; Ohmori, K.; Kikuchi, N.; Nakaya, N.; Nishino, Y.; Tsubono, Y.; Tsuji, I. Green tea consumption and mortality due to cardiovascular disease, cancer, and all causes in japan: The ohsaki study. JAMA 2006, 296, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, S.; Zhou, H.; Hanson, T.; Yang, L.; Chen, Z.; Zhou, M. Association of green tea consumption with mortality from all-cause, cardiovascular disease and cancer in a Chinese cohort of 165,000 adult men. Eur. J. Epidemiol. 2016, 31, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Renno, W.M.; Abdeen, S.; Alkhalaf, M.; Asfar, S. Effect of green tea on kidney tubules of diabetic rats. Br. J. Nutr. 2008, 100, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Moon, J.H.; Kim, S.W.; Jeong, J.K.; Nazim, U.M.; Lee, Y.J.; Seol, J.W.; Park, S.Y. Egcg-mediated autophagy flux has a neuroprotection effect via a class iii histone deacetylase in primary neuron cells. Oncotarget 2015, 6, 9701–9717. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.W.; Tian, C.; Xu, F.Y.; Chen, Z.; Burnside, R.; Yi, W.J.; Xiang, S.Y.; Xie, X.; Wu, N.N.; Yang, H.; et al. Green tea polyphenols alleviate autophagy inhibition induced by high glucose in endothelial cells. Biomed. Environ. Sci. 2016, 29, 524–528. [Google Scholar] [PubMed]
- Tikoo, K.; Sharma, E.; Amara, V.R.; Pamulapati, H.; Dhawale, V.S. Metformin improves metabolic memory in high fat diet (hfd)-induced renal dysfunction. J. Biol. Chem. 2016, 291, 21848–21856. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.R.; Ren, Y.L.; Zhu, J.J.; Hu, Y.J.; Zheng, J.S.; Fan, H.; Xu, Y.; Wang, G.; Liu, W.X. Resveratrol increases nephrin and podocin expression and alleviates renal damage in rats fed a high-fat diet. Nutrients 2014, 6, 2619–2631. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto-Nonaka, K.; Koike, M.; Asanuma, K.; Takagi, M.; Oliva Trejo, J.A.; Seki, T.; Hidaka, T.; Ichimura, K.; Sakai, T.; Tada, N.; et al. Cathepsin d in podocytes is important in the pathogenesis of proteinuria and ckd. J. Am. Soc. Nephrol. 2016, 27, 2685–2700. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Liu, L.; Xin, W.; Zhao, X.; Chen, L.; Zhen, J.; Wan, Q. The renoprotective role of autophagy activation in proximal tubular epithelial cells in diabetic nephropathy. J. Diabetes Complicat. 2015, 29, 976–983. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Shen, T.T.; Chen, R.H.; Wu, H.L.; Wang, Y.J.; Deng, J.K.; Chen, Q.H.; Pan, Q.; Huang Fu, C.M.; Tao, J.L.; et al. Autophagy-lysosome pathway in renal tubular epithelial cells is disrupted by advanced glycation end products in diabetic nephropathy. J. Biol. Chem. 2015, 290, 20499–20510. [Google Scholar] [CrossRef] [PubMed]
- Devika, P.T.; Prince, P.S. Preventive effect of (-)epigallocatechin-gallate (egcg) on lysosomal enzymes in heart and subcellular fractions in isoproterenol-induced myocardial infarcted wistar rats. Chem. Biol. Interact. 2008, 172, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, S.E.; Julian, D.; Akin, D.E.; Fried, J.; Toscano, K.; Leeuwenburgh, C.; Dunn, W.A., Jr. Autophagy in the heart and liver during normal aging and calorie restriction. Rejuvenation Res. 2007, 10, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.C.; Cai, G.Y.; Zhuo, L.; Gao, J.J.; Dong, D.; Cui, S.; Feng, Z.; Shi, S.Z.; Bai, X.Y.; Sun, X.F.; et al. Short-term calorie restriction protects against renal senescence of aged rats by increasing autophagic activity and reducing oxidative damage. Mech. Ageing Dev. 2013, 134, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Farah, B.L.; Sinha, R.A.; Wu, Y.; Singh, B.K.; Bay, B.H.; Yang, C.S.; Yen, P.M. Epigallocatechin-3-gallate (egcg), a green tea polyphenol, stimulates hepatic autophagy and lipid clearance. PLoS ONE 2014, 9, e87161. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Montana, V.; Jang, H.J.; Parpura, V.; Kim, J.A. Epigallocatechin gallate (egcg) stimulates autophagy in vascular endothelial cells: A potential role for reducing lipid accumulation. J. Biol. Chem. 2013, 288, 22693–22705. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Obesity and diabetic kidney disease: Role of oxidant stress and redox balance. Antioxid. Redox Signal 2016, 25, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Declèves, A.-E.; Zolkipli, Z.; Satriano, J.; Wang, L.; Nakayama, T.; Rogac, M.; Le, T.P.; Nortier, J.L.; Farquhar, M.G.; Naviaux, R.K.; et al. Regulation of lipid accumulation by amk-activated kinase in high fat diet–induced kidney injury. Kidney Int. 2014, 85, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Takemura, G.; Kanamori, H.; Goto, K.; Tsujimoto, A.; Okada, H.; Kawamura, I.; Ogino, A.; Takeyama, T.; Kawaguchi, T.; et al. Restriction of food intake prevents postinfarction heart failure by enhancing autophagy in the surviving cardiomyocytes. Am. J. Pathol. 2014, 184, 1384–1394. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.J.; Li, Y.F.; Liu, F.L.; Deng, J.; Wu, Y.P.; Yuan, W.L.; Tsoi, B.; Chen, J.L.; Wang, Q.; Cai, S.H.; et al. A sirt3/ampk/autophagy network orchestrates the protective effects of trans-resveratrol in stressed peritoneal macrophages and raw 264.7 macrophages. Free Radic. Biol. Med. 2016, 95, 230–242. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
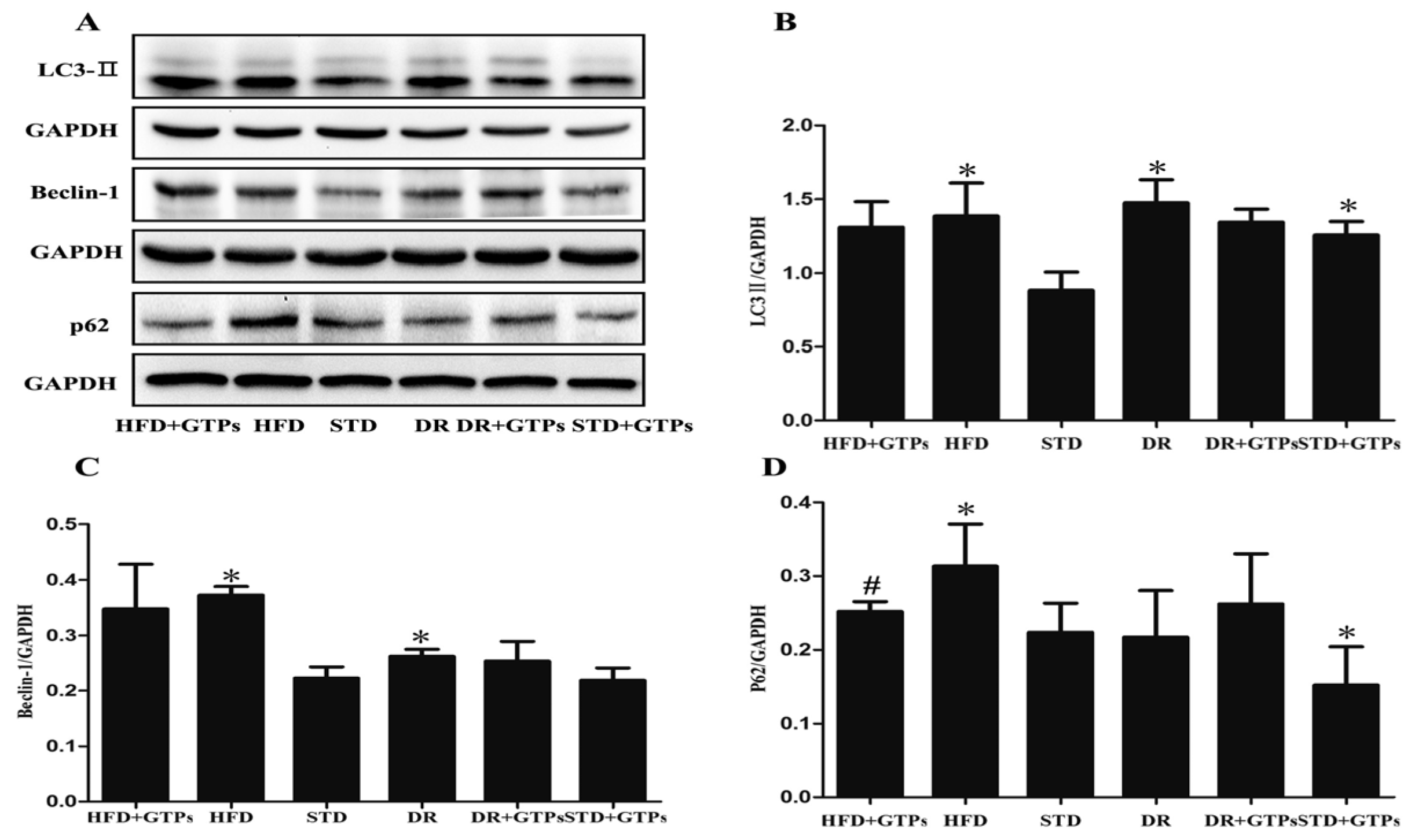{kind=link}
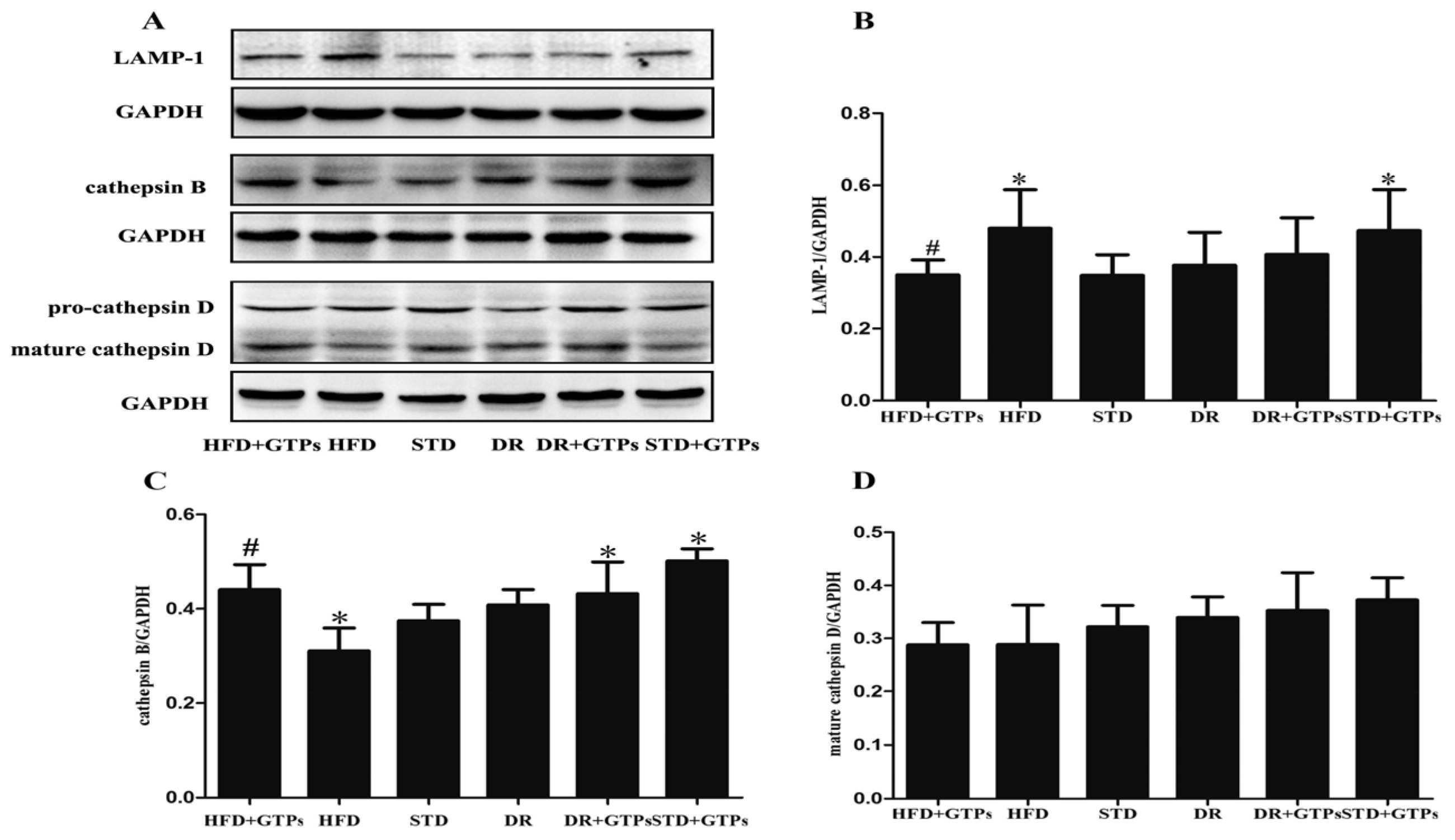{kind=link}
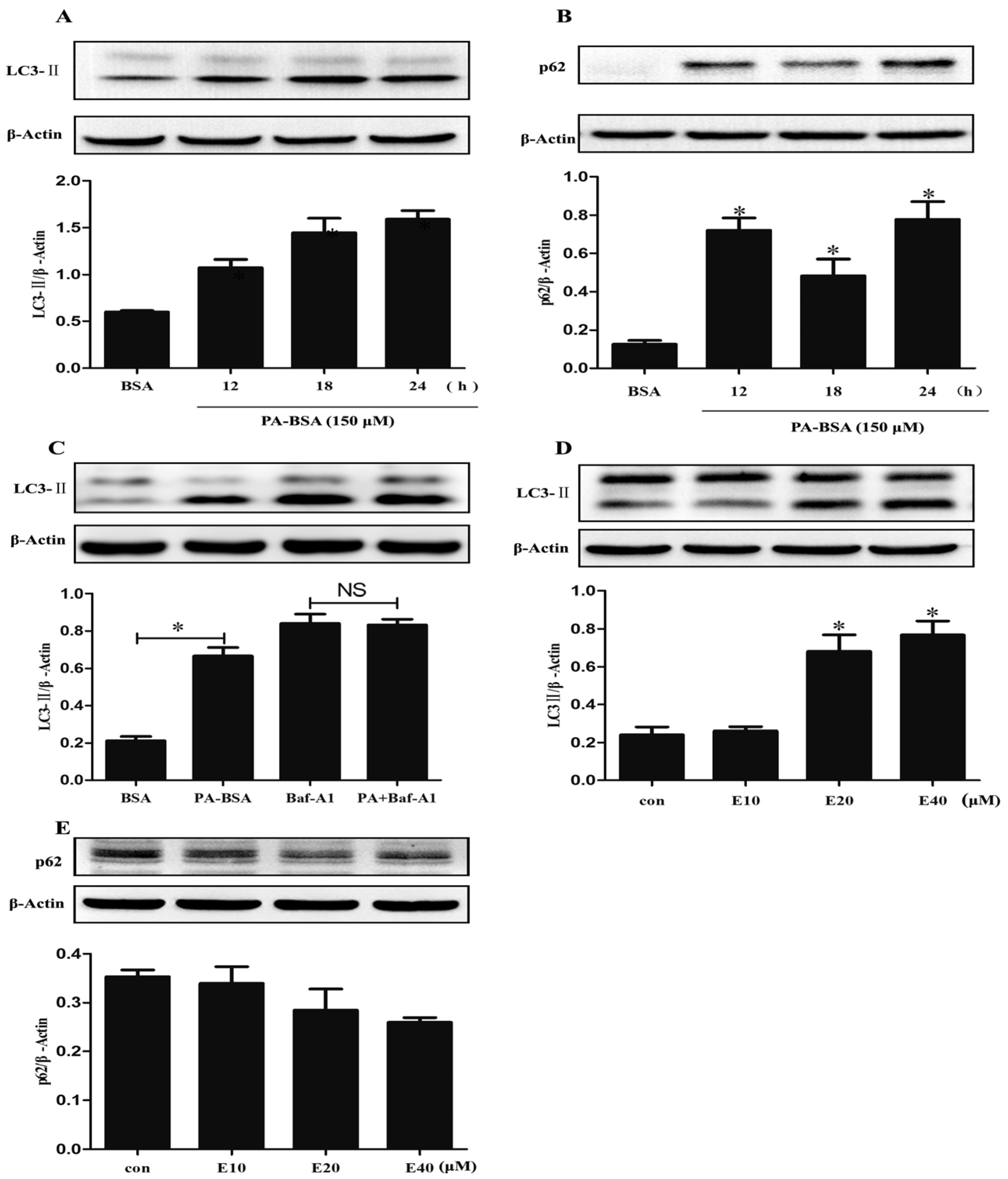{kind=link}
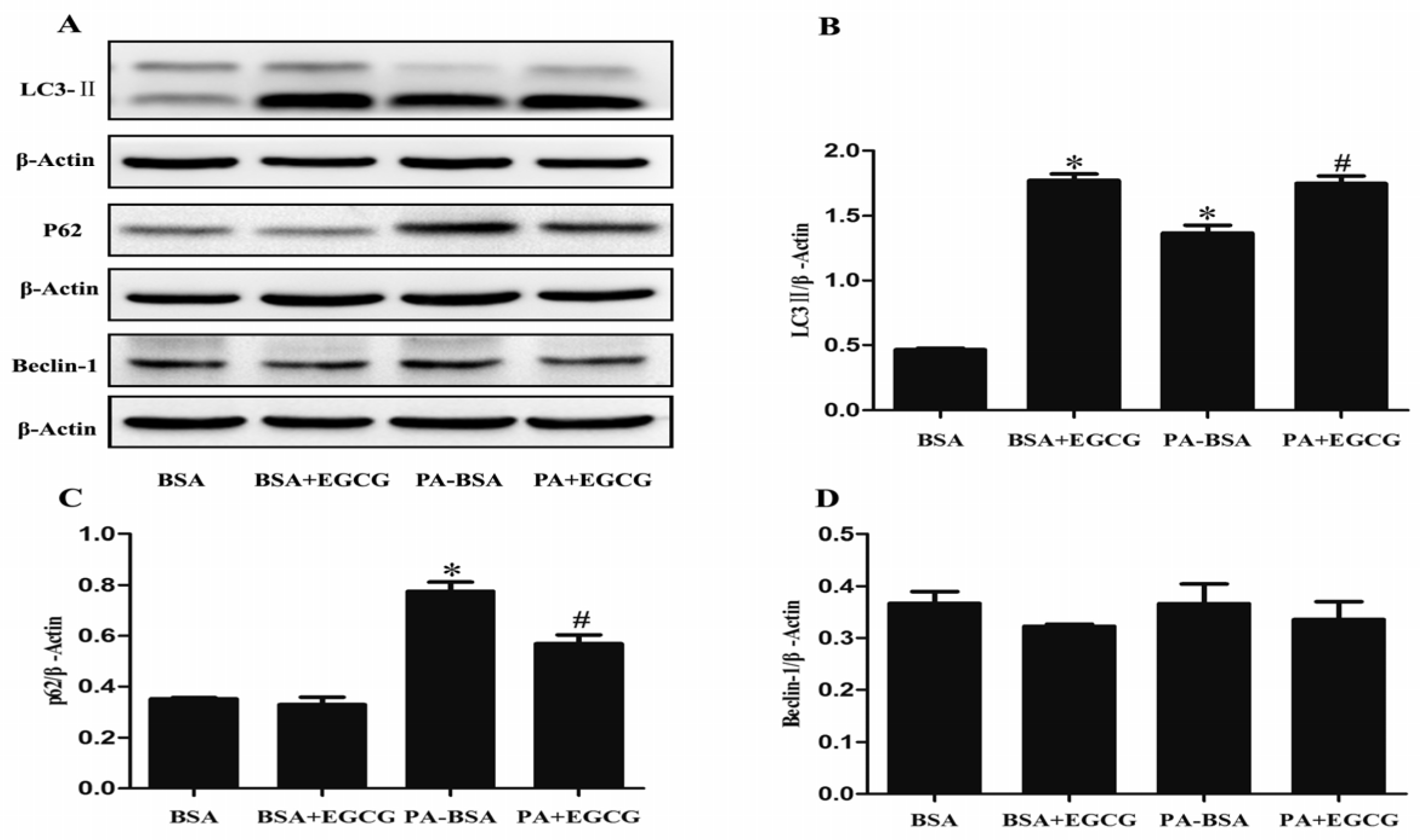{kind=link}
{kind=link}
| Groups | Ccr (mL/h/kg(bw)) | Serum Cys C (μg/mL) | Urinary NAG (U/L) |
|---|---|---|---|
| STD | 68.35 ± 26.11 | 1.42 ± 0.39 | 20.51 ± 4.03 |
| STD+GTPs | 61.66 ± 20.69 | 1.51 ± 0.50 | 19.35 ± 6.07 |
| DR | 109.03 ± 33.02 * | 1.34 ± 0.10 | 13.84 ± 5.28 * |
| DR+GTPs | 102.32 ± 43.40 | 1.45 ± 0.35 | 13.74 ± 4.90 |
| HFD | 46.77 ± 28.76 | 1.99 ± 0.34 * | 28.54 ± 5.13 * |
| HFD+GTPs | 52.05 ± 18.35 | 1.45 ± 0.49 # | 22.19 ± 2.91 # |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, X.; Yi, W.; Zhang, P.; Wu, N.; Yan, Q.; Yang, H.; Tian, C.; Xiang, S.; Du, M.; Getachew Assefa, E.; et al. Green Tea Polyphenols, Mimicking the Effects of Dietary Restriction, Ameliorate High-Fat Diet-Induced Kidney Injury via Regulating Autophagy Flux. Nutrients 2017, 9, 497. https://doi.org/10.3390/nu9050497
Xie X, Yi W, Zhang P, Wu N, Yan Q, Yang H, Tian C, Xiang S, Du M, Getachew Assefa E, et al. Green Tea Polyphenols, Mimicking the Effects of Dietary Restriction, Ameliorate High-Fat Diet-Induced Kidney Injury via Regulating Autophagy Flux. Nutrients. 2017; 9(5):497. https://doi.org/10.3390/nu9050497
Chicago/Turabian StyleXie, Xiao, Weijie Yi, Piwei Zhang, Nannan Wu, Qiaoqiao Yan, Hui Yang, Chong Tian, Siyun Xiang, Miying Du, Eskedar Getachew Assefa, and et al. 2017. "Green Tea Polyphenols, Mimicking the Effects of Dietary Restriction, Ameliorate High-Fat Diet-Induced Kidney Injury via Regulating Autophagy Flux" Nutrients 9, no. 5: 497. https://doi.org/10.3390/nu9050497