Post-Intake of S-Ethyl Cysteine and S-Methyl Cysteine Improved LPS-Induced Acute Lung Injury in Mice

Abstract
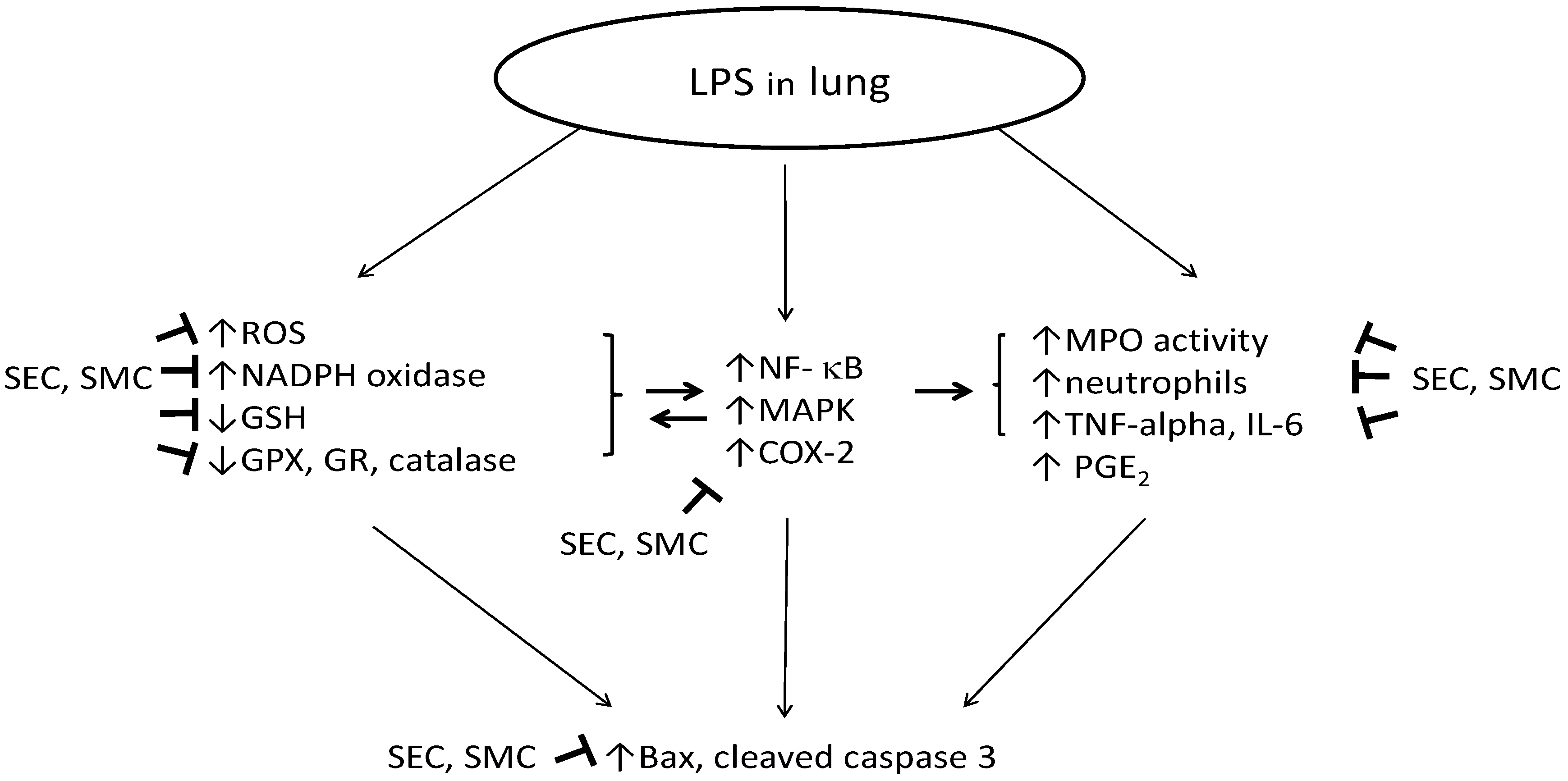
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Experimental Design
2.4. Lung Wet/Dry Weight Ratio
2.5. MPO Activity Assay
2.6. Determination of Inflammatory Factors
2.7. Measurement of Oxidative Factors
2.8. Western Blot Analyses
2.9. Statistical Analyses
3. Results
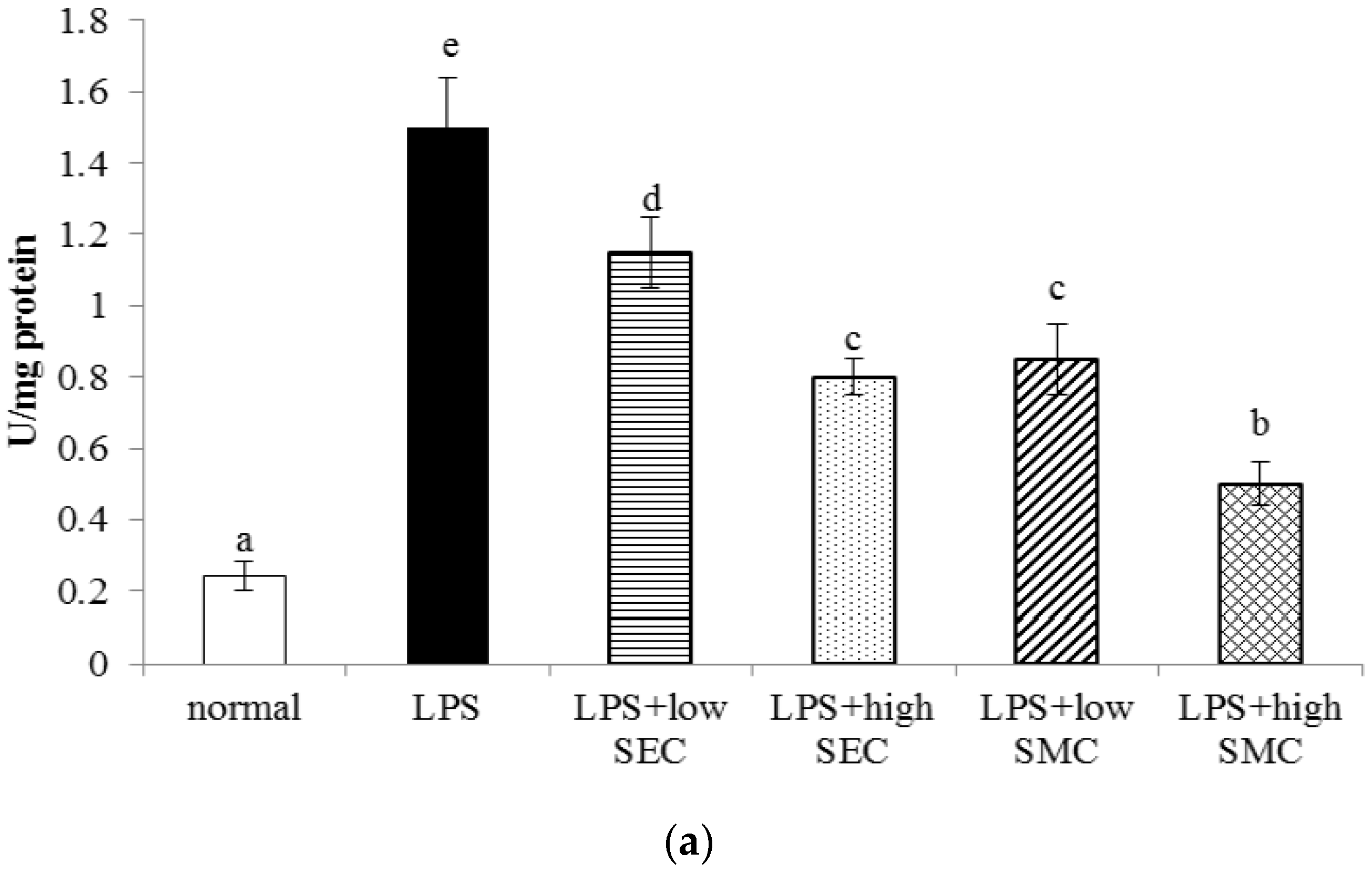
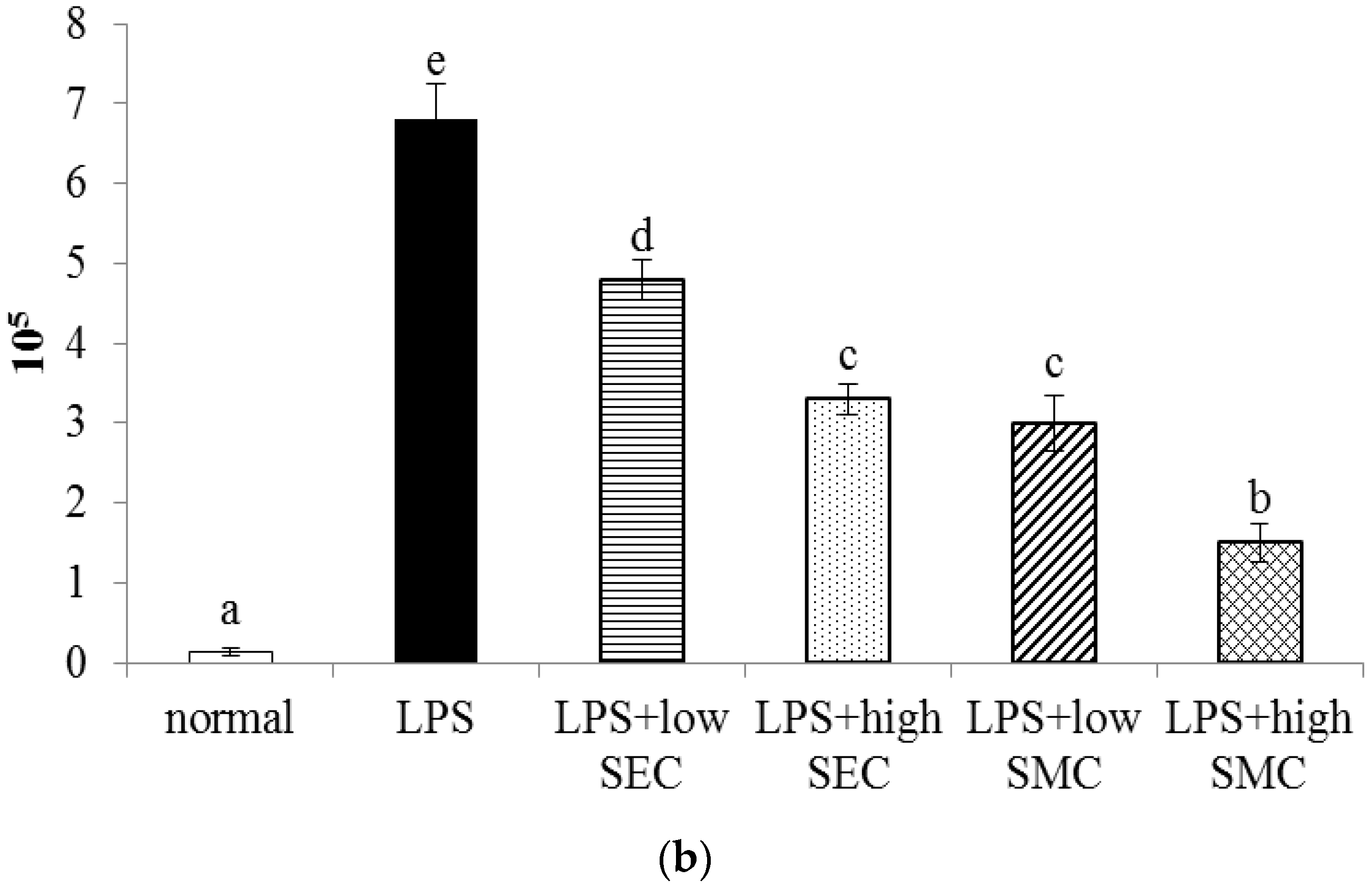
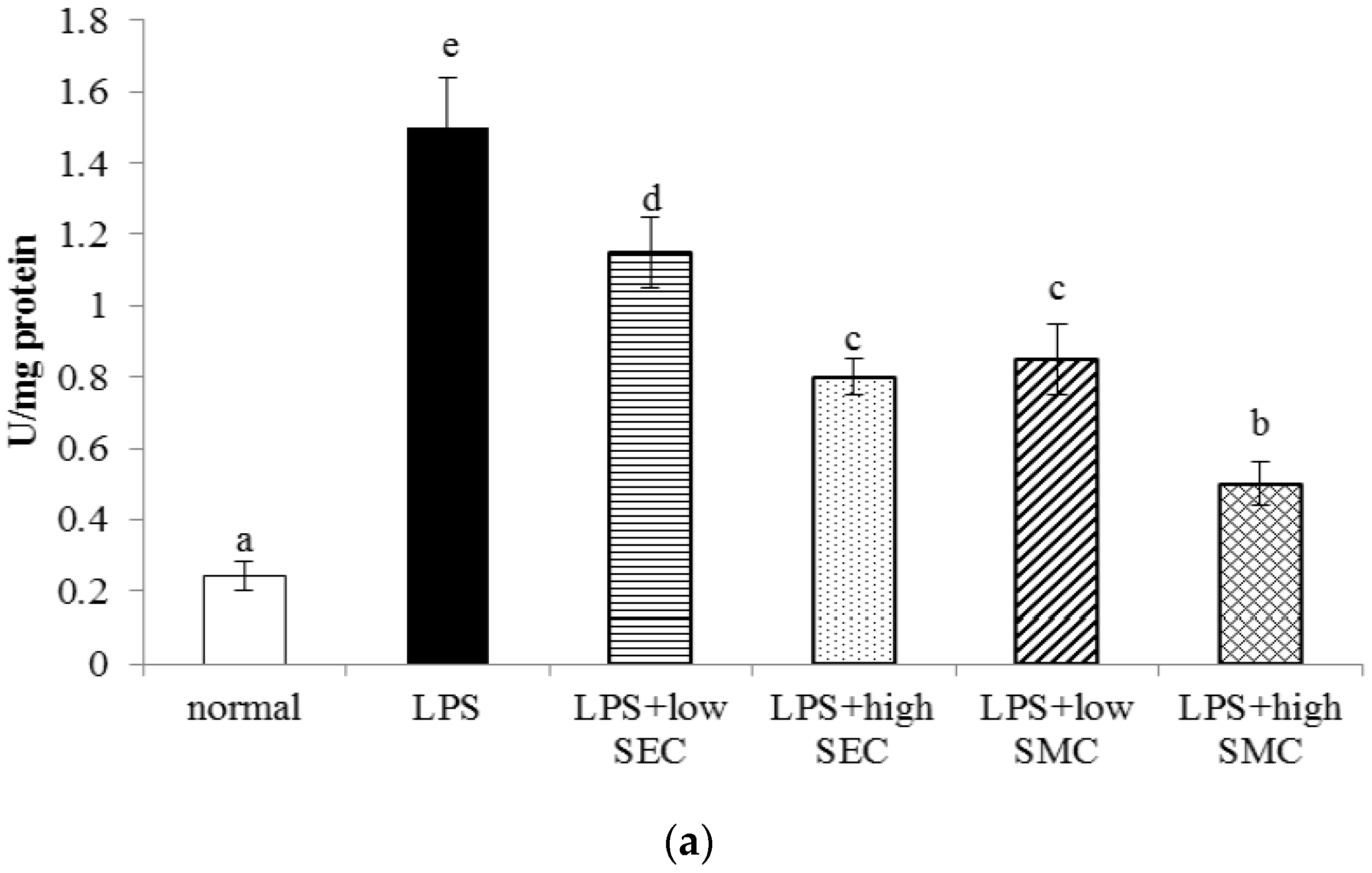
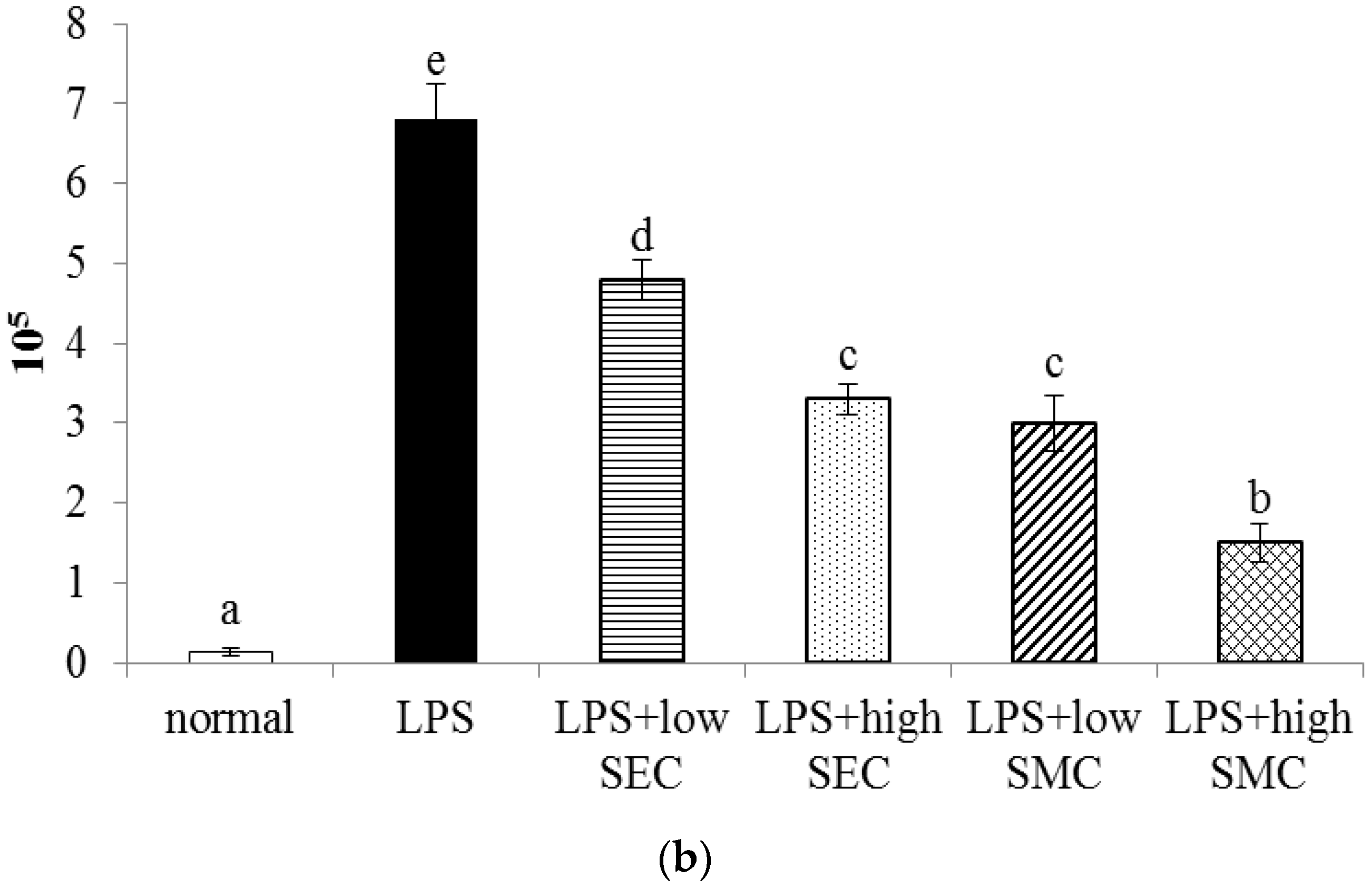
3.1. Effects of SEC and SMC on MPO and Neutrophil Counts
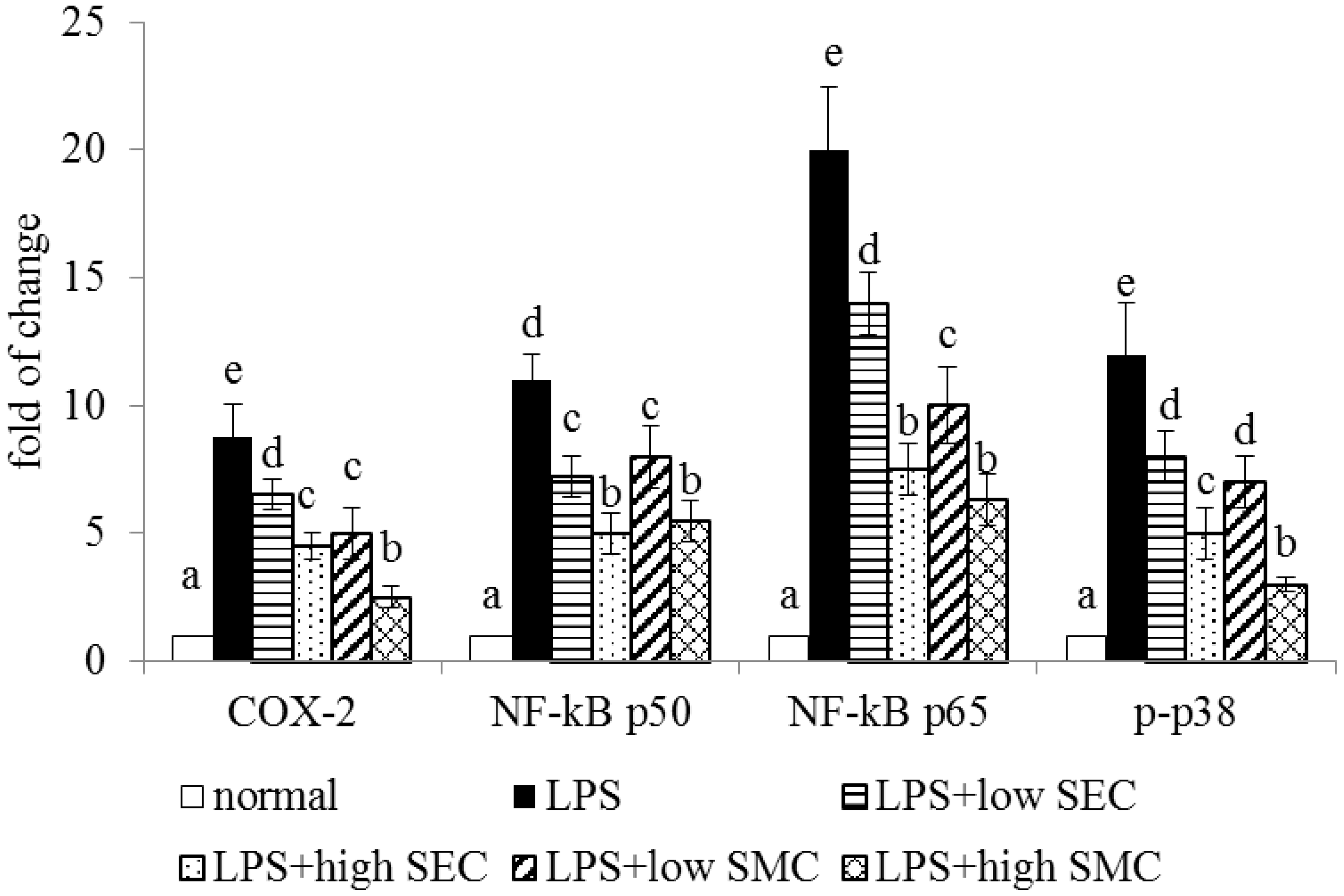
3.2. Effects of SEC and SMC on Inflammatory Factors
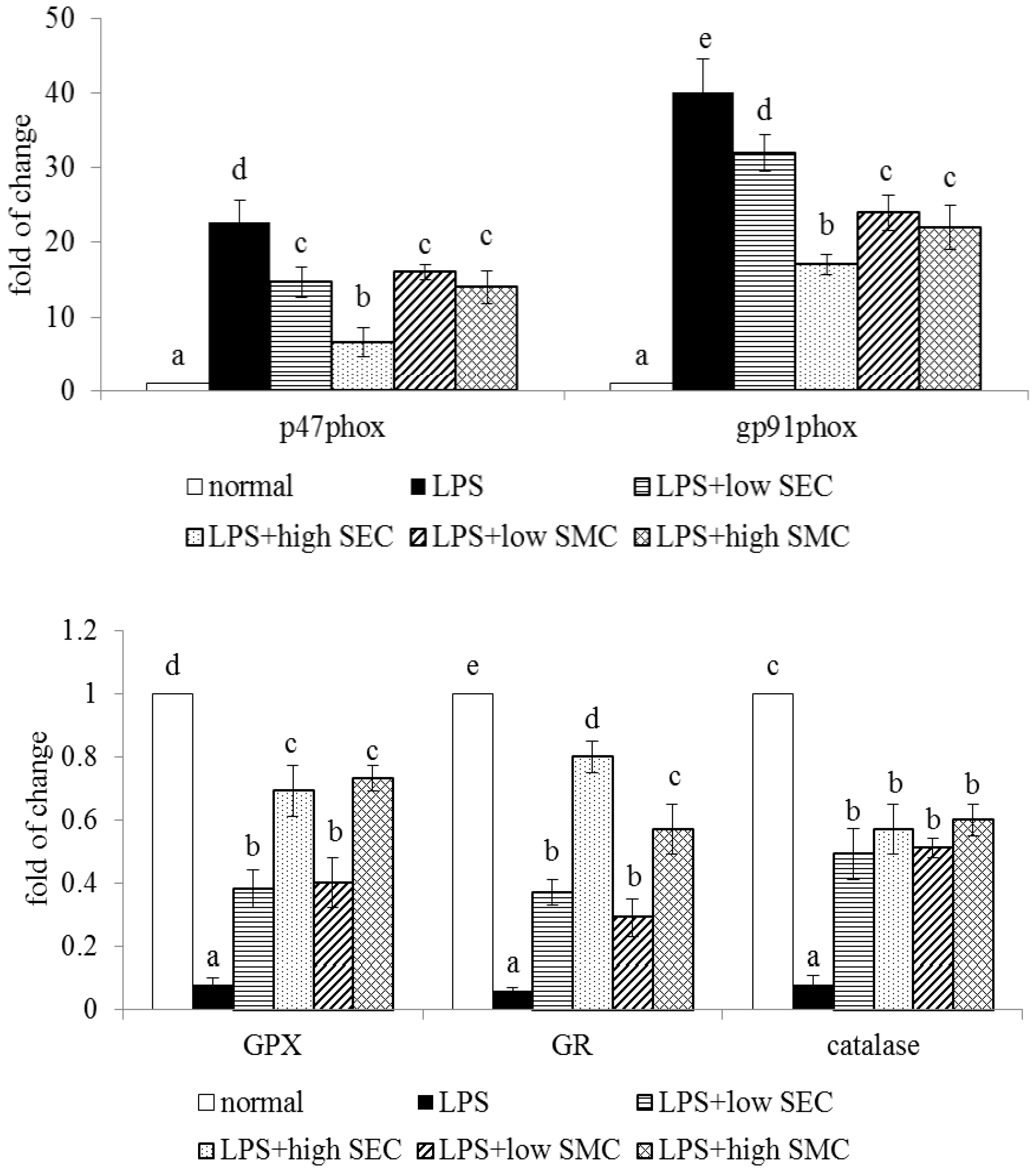
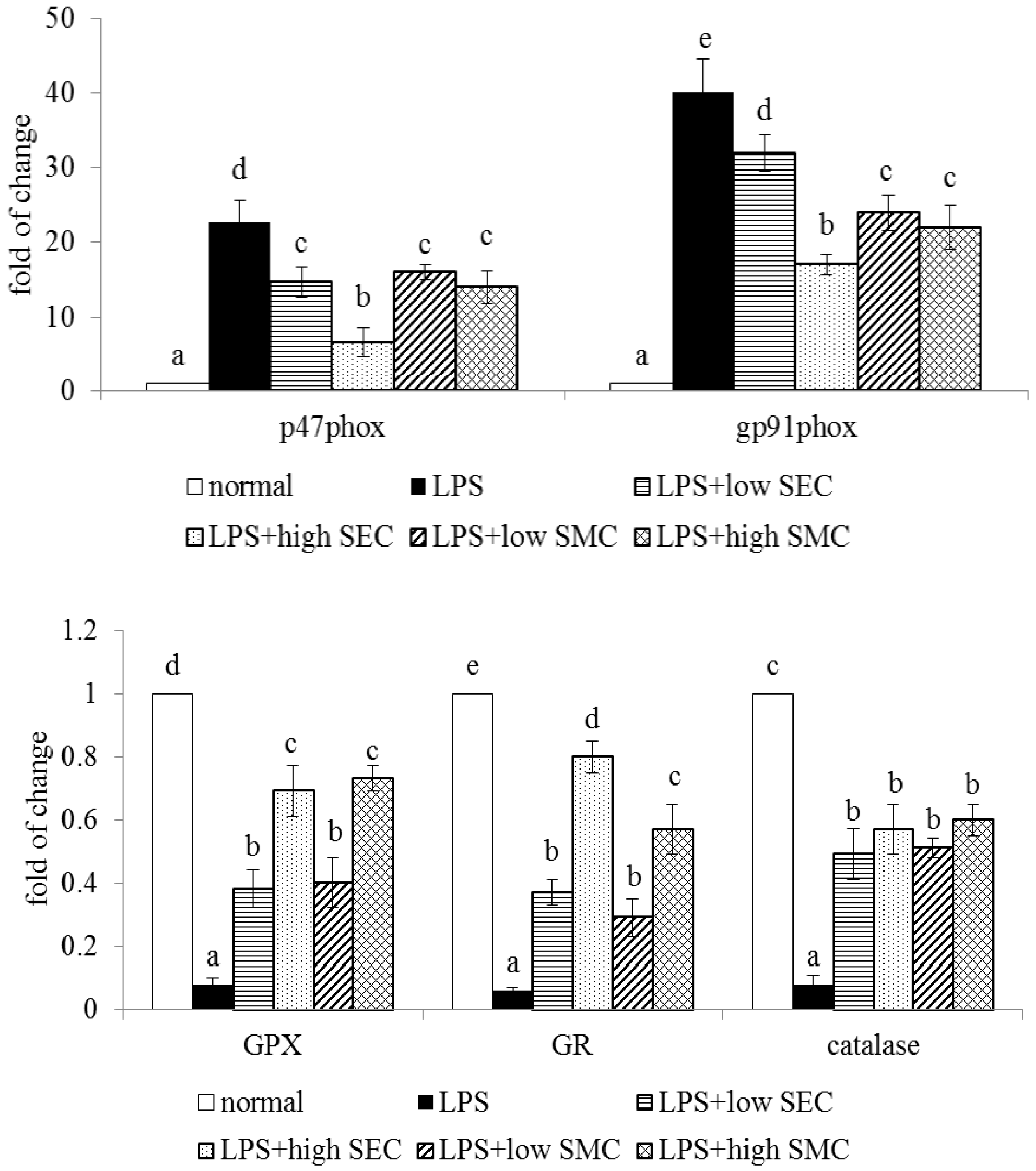
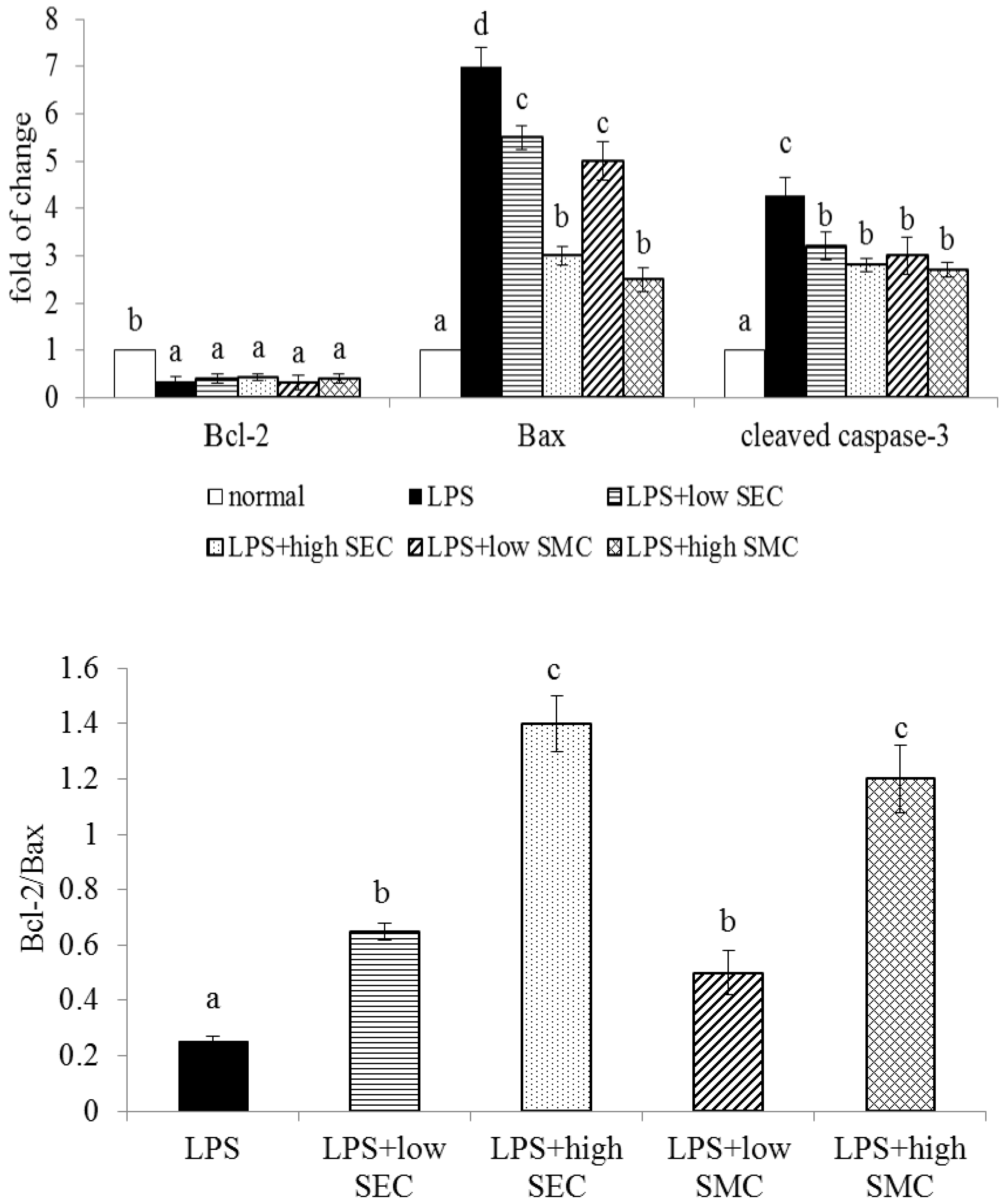
3.3. Effects of SEC and SMC upon Oxidative and Apoptotic Factors
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Johnson, E.R.; Matthay, M.A. Acute lung injury: Epidemiology, pathogenesis, and treatment. J. Aerosol. Med. Pulm. Drug Deliv. 2010, 23, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Puneet, P.; Moochhala, S.; Bhatia, M. Chemokines in acute respiratory distress syndrome. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 288, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Chi, G.; Wei, M.; Xie, X.; Soromou, L.W.; Liu, F.; Zhao, S. Suppression of MAPK and NF-κB pathways by limonene contributes to attenuation of lipopolysaccharide-induced inflammatory responses in acute lung injury. Inflammation 2013, 36, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, J.; Samanta, S.; Rajasingh, S.; Barani, B.; Xuan, Y.T.; Dawn, B.; Rajasingh, J. Epigenetic modifiers reduce inflammation and modulate macrophage phenotype during endotoxemia-induced acute lung injury. J. Cell Sci. 2015, 128, 3094–3105. [Google Scholar] [CrossRef] [PubMed]
- Ang, S.F.; Sio, S.W.; Moochhala, S.M.; MacAry, P.A.; Bhatia, M. Hydrogen sulfide upregulates cyclooxygenase-2 and prostaglandin E metabolite in sepsis-evoked acute lung injury via transient receptor potential vanilloid type 1 channel activation. J. Immunol. 2011, 187, 4778–4787. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Yu, Z.; Zheng, Y.; Wang, L.; Qin, X.; Cheng, G.; Ci, X. Isovitexin exerts anti-inflammatory and anti-oxidant activities on lipopolysaccharide-induced acute lung injury by inhibiting MAPK and NF-κB and activating HO-1/Nrf2 pathways. Int. J. Biol. Sci. 2016, 12, 72–86. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Dodia, C.; Chatterjee, S.; Feinstein, S.I.; Fisher, A.B. Protection against LPS-induced acute lung injury by a mechanism-based inhibitor of NADPH oxidase (type 2). Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Dimitropoulou, C.; Lu, Q.; Black, S.M.; Sharma, S. Glutathione supplementation attenuates lipopolysaccharide-induced mitochondrial dysfunction and apoptosis in a mouse model of acute lung injury. Front. Physiol. 2012, 3, 161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, J.; Hu, M.; Yang, Y.; Guo, L.; Xu, J.; Lei, C.; Jiao, Y.; Xu, J. Uncoupling protein 2 increases susceptibility to lipopolysaccharide-induced acute lung injury in mice. Mediators Inflamm. 2016, 2016, 9154230. [Google Scholar] [CrossRef] [PubMed]
- Mehla, K.; Balwani, S.; Agrawal, A.; Ghosh, B. Ethyl gallate attenuates acute lung injury through Nrf2 signaling. Biochimie 2013, 95, 2404–2414. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Yang, J.J.; Yang, M.L.; Li, Y.C.; Kuan, Y.H. Rutin decreases lipopolysaccharide-induced acute lung injury via inhibition of oxidative stress and the MAPK-NF-κB pathway. Free Radic. Biol. Med. 2014, 69, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bai, C.; Wang, X. The value of the lipopolysaccharide-induced acute lung injury model in respiratory medicine. Expert. Rev. Respir. Med. 2010, 4, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.G.; Hughes, J.; Tregova, A.; Milne, J.; Tomsett, A.B.; Collin, H.A. Biosynthesis of the flavour precursors of onion and garlic. J. Exp. Bot. 2004, 55, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Mohi El-Din, M.M.; Mostafa, A.M.; Abd-Elkader, A. Experimental studies on the effect of (Lambda-Cyhalothrin) insecticide on lungs and the ameliorating effect of plant extracts (Ginseng (Panax Ginseng) and garlic (Allium sativum L.) on asthma development in albino rats. BMC Res. Notes 2014, 7, 243. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.L.; Yin, M.C. Protective and alleviative effects from 4 cysteine-containing compounds on ethanol-induced acute liver injury through suppression of oxidation and inflammation. J. Food Sci. 2007, 72, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Hsia, T.C.; Yin, M.C. S-ethyl cysteine and S-methyl cysteine protect human bronchial epithelial cells against hydrogen peroxide induced injury. J. Food Sci. 2015, 80, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.B.; Bae, G.S.; Jo, I.J.; Wang, S.; Song, H.J.; Park, S.J. Berberine inhibits inflammatory mediators and attenuates acute pancreatitis through deactivation of JNK signaling pathways. Mol. Immunol. 2016, 74, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.F.; LeBel, C.P.; Bondy, S.C. Reactive oxygen species formation as a biomarker of methylmercury and trimethyltin neurotoxicity. Neurotoxicology 1992, 13, 637–648. [Google Scholar] [PubMed]
- Niu, X.; Wang, Y.; Li, W.; Mu, Q.; Li, H.; Yao, H.; Zhang, H. Protective effects of Isofraxidin against lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2015, 24, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Mokra, D.; Kosutova, P. Biomarkers in acute lung injury. Respir. Physiol. Neurobiol. 2015, 209, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Grommes, J.; Soehnlein, O. Contribution of neutrophils to acute lung injury. Mol. Med. 2011, 17, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Ren, H.; Xu, N.; Xia, L.; Chen, D.; Zhang, J. Eupatorium lindleyanum DC. sesquiterpenes fraction attenuates lipopolysaccharide-induced acute lung injury in mice. J. Ethnopharmacol. 2016, 185, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Jiang, Y.W.; Fang, Q.H. Improving effect of Sivelestat on lipopolysaccharide-induced lung injury in rats. APMIS 2014, 122, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Hardy, L.A.; Booth, T.A.; Lau, E.K.; Handel, T.M.; Ali, S.; Kirby, J.A. Examination of MCP-1 (CCL2) partitioning and presentation during transendothelial leukocyte migration. Lab. Investig. 2004, 84, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Maijó, M.; Miró, L.; Polo, J.; Campbell, J.; Russell, L.; Crenshaw, J.; Weaver, E.; Moretó, M.; Pérez-Bosque, A. Dietary plasma proteins attenuate the innate immunity response in a mouse model of acute lung injury. Br. J. Nutr. 2012, 107, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Britt, R.D., Jr.; Locy, M.L.; Tipple, T.E.; Nelin, L.D.; Rogers, L.K. Lipopolysaccharide-induced cyclooxygenase-2 expression in mouse transformed Clara cells. Cell. Physiol. Biochem. 2012, 29, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Park, G.Y.; Christman, J.W. Involvement of cyclooxygenase-2 and prostaglandins in the molecular pathogenesis of inflammatory lung diseases. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase isozymes: The biology of prostaglandin synthesis and inhibition. Pharmacol. Rev. 2004, 56, 387–437. [Google Scholar] [CrossRef] [PubMed]
- Carnesecchi, S.; Pache, J.C.; Barazzone-Argiroffo, C. NOX enzymes: Potential target for the treatment of acute lung injury. Cell Mol. Life Sci. 2012, 69, 2373–2385. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal | LPS | LPS + SEC, Low | LPS + SEC, High | LPS + SMC, Low | LPS + SMC, High | |
|---|---|---|---|---|---|---|
| WI | 2.3 ± 0.8 a | 1.9 ± 0.4 a | 2.1 ± 0.5 a | 2.0 ± 0.7 a | 1.8 ± 0.6 a | 2.1 ± 0.7 a |
| FI | 2.2 ± 0.5 a | 1.8 ± 0.6 a | 2.0 ± 0.4 a | 1.9 ± 0.6 a | 1.8 ± 0.4 a | 2.0 ± 0.5 a |
| BW | 26.1 ± 1.0 b | 22.9 ± 0.4 a | 24.9 ± 0.6 b | 25.5 ± 0.5 b | 24.7 ± 0.3 b | 25.3 ± 0.8 b |
| W/D | 2.8 ± 0.2 a | 6.4 ± 0.4 d | 5.3 ± 0.2 c | 3.9 ± 0.3 b | 5.0 ± 0.2 c | 3.7 ± 0.4 b |
| ALT | 27 ± 2 a | 35 ± 5 a | 32 ± 3 a | 28 ± 4 a | 29 ± 3 a | 25 ± 4 a |
| AST | 24 ± 4 a | 31 ± 2 a | 30 ± 4 a | 26 ± 5 a | 32 ± 4 a | 27 ± 3 a |
| Normal | LPS | LPS + SEC, Low | LPS + SEC, High | LPS + SMC, Low | LPS + SMC, High | |
|---|---|---|---|---|---|---|
| IL-1beta | 12 ± 3 a | 195 ± 14 e | 149 ± 9 d | 92 ± 11 c | 138 ± 13 d | 68 ± 6 b |
| IL-6 | 10 ± 2 a | 218 ± 10 e | 161 ± 12 d | 101 ± 7 c | 153 ± 9 d | 57 ± 5 b |
| TNF-alpha | 13 ± 4 a | 253 ± 16 d | 192 ± 8 c | 112 ± 13 b | 188 ± 14 c | 97 ± 10 b |
| MCP-1 | 15 ± 3 a | 231 ± 12 e | 174 ± 15 d | 116 ± 9 c | 165 ± 11 d | 82 ± 5 b |
| PGE2 | 558 ± 49 a | 2271 ± 124 d | 1645 ± 110 c | 1082 ± 96 b | 1431 ± 108 c | 952 ± 73 b |
| Normal | LPS | LPS + SEC, Low | LPS + SEC, High | LPS + SMC, Low | LPS + SMC, High | |
|---|---|---|---|---|---|---|
| GSH | 19.1 ± 0.5 e | 8.1 ± 0.2 a | 11.3 ± 0.4 b | 16.0 ± 0.7 d | 11.1 ± 0.4 b | 13.9 ± 0.3 c |
| GSSG | 0.27 ± 0.05 a | 1.50 ± 0.16 d | 0.98 ± 0.09 c | 0.59 ± 0.06 b | 1.03 ± 0.08 c | 0.67 ± 0.07 b |
| ROS | 0.19 ± 0.04 a | 1.71 ± 0.14 e | 1.19 ± 0.11 d | 0.58 ± 0.09 b | 1.25 ± 0.12 d | 0.92 ± 0.1 c |
| GPX | 29.4 ± 2.3 d | 16.3 ± 1.4 a | 19.0 ± 1.2 b | 23.3 ± 1.6 c | 20.1 ± 0.8 b | 24.5 ± 1.3 c |
| GR | 1.82 ± 0.12 d | 0.67 ± 0.06 a | 1.02 ± 0.1 b | 1.38 ± 0.08 c | 0.95 ± 0.07 b | 1.31 ± 0.11 c |
| catalase | 22.8 ± 1.5 d | 12.7 ± 0.6 a | 15.9 ± 1.4 b | 19.0 ± 0.7 c | 15.1 ± 1.0 b | 18.6 ± 1.3 c |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsia, T.-c.; Yin, M.-c. Post-Intake of S-Ethyl Cysteine and S-Methyl Cysteine Improved LPS-Induced Acute Lung Injury in Mice. Nutrients 2016, 8, 507. https://doi.org/10.3390/nu8080507
Hsia T-c, Yin M-c. Post-Intake of S-Ethyl Cysteine and S-Methyl Cysteine Improved LPS-Induced Acute Lung Injury in Mice. Nutrients. 2016; 8(8):507. https://doi.org/10.3390/nu8080507
Chicago/Turabian StyleHsia, Te-chun, and Mei-chin Yin. 2016. "Post-Intake of S-Ethyl Cysteine and S-Methyl Cysteine Improved LPS-Induced Acute Lung Injury in Mice" Nutrients 8, no. 8: 507. https://doi.org/10.3390/nu8080507