Plasma 25-Hydroxyvitamin D Is Related to Protein Signaling Involved in Glucose Homeostasis in a Tissue-Specific Manner



Abstract
:1. Introduction
2. Materials and Methods
3. Results
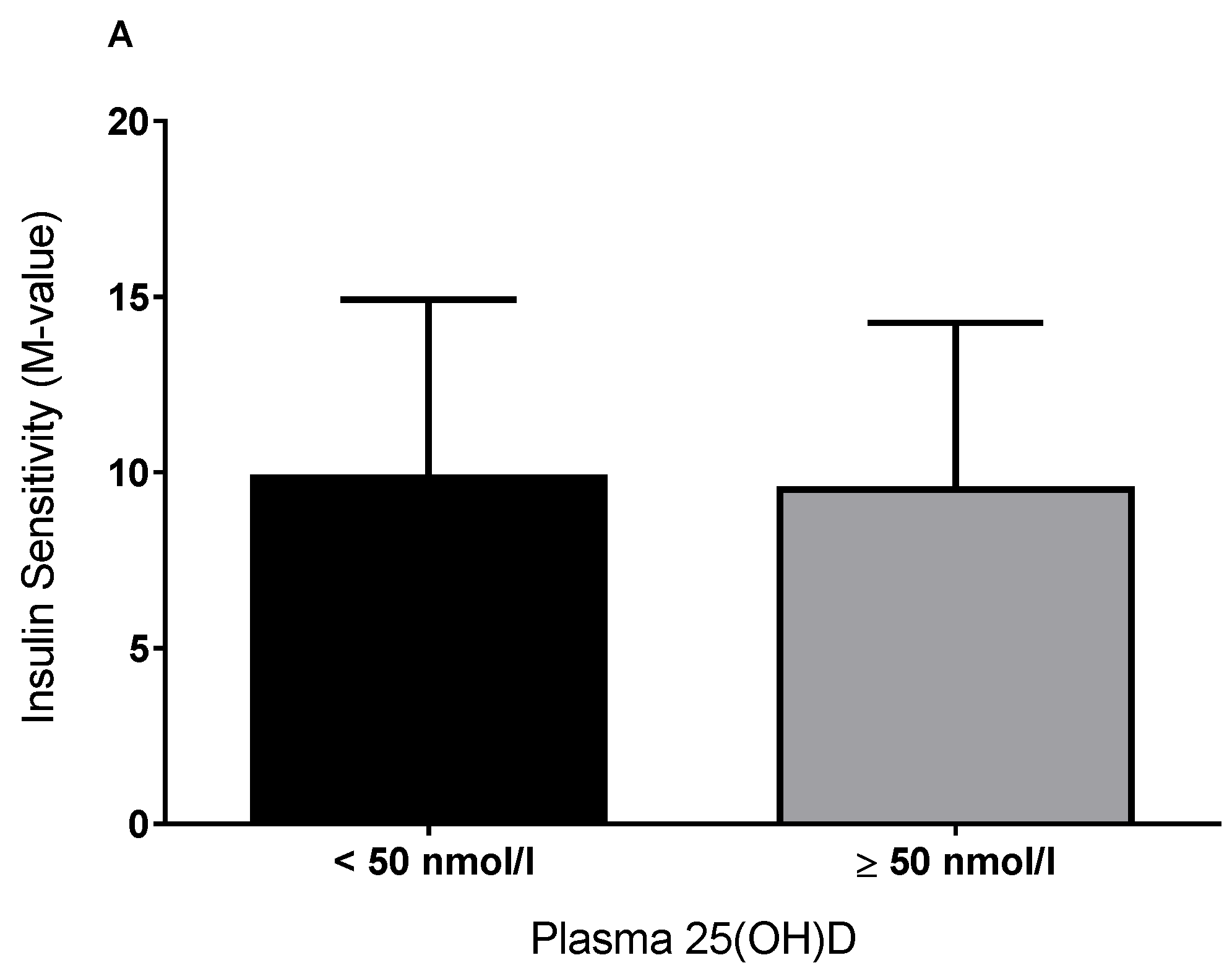
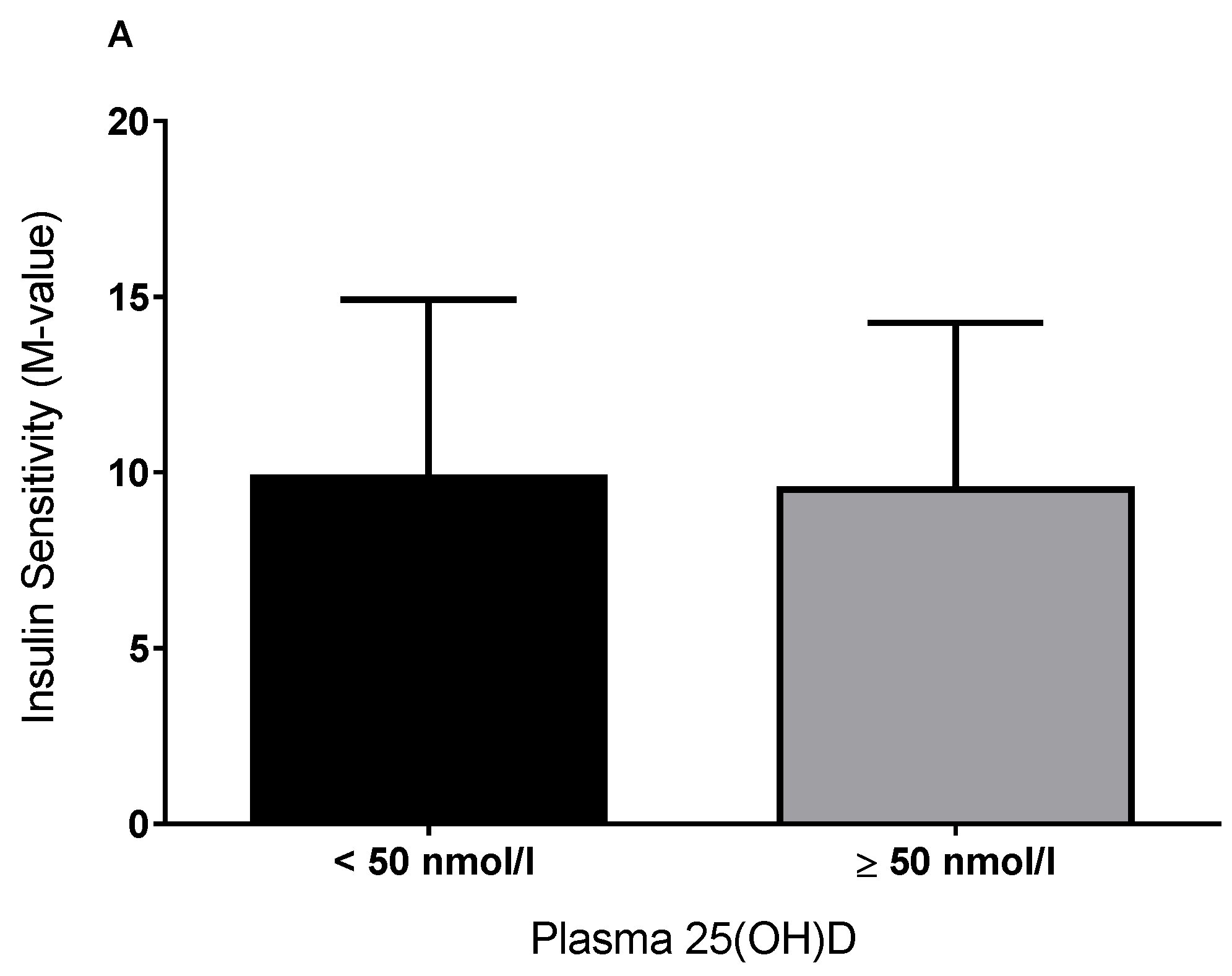
3.1. Plasma 25(OH)D and Insulin Sensitivity
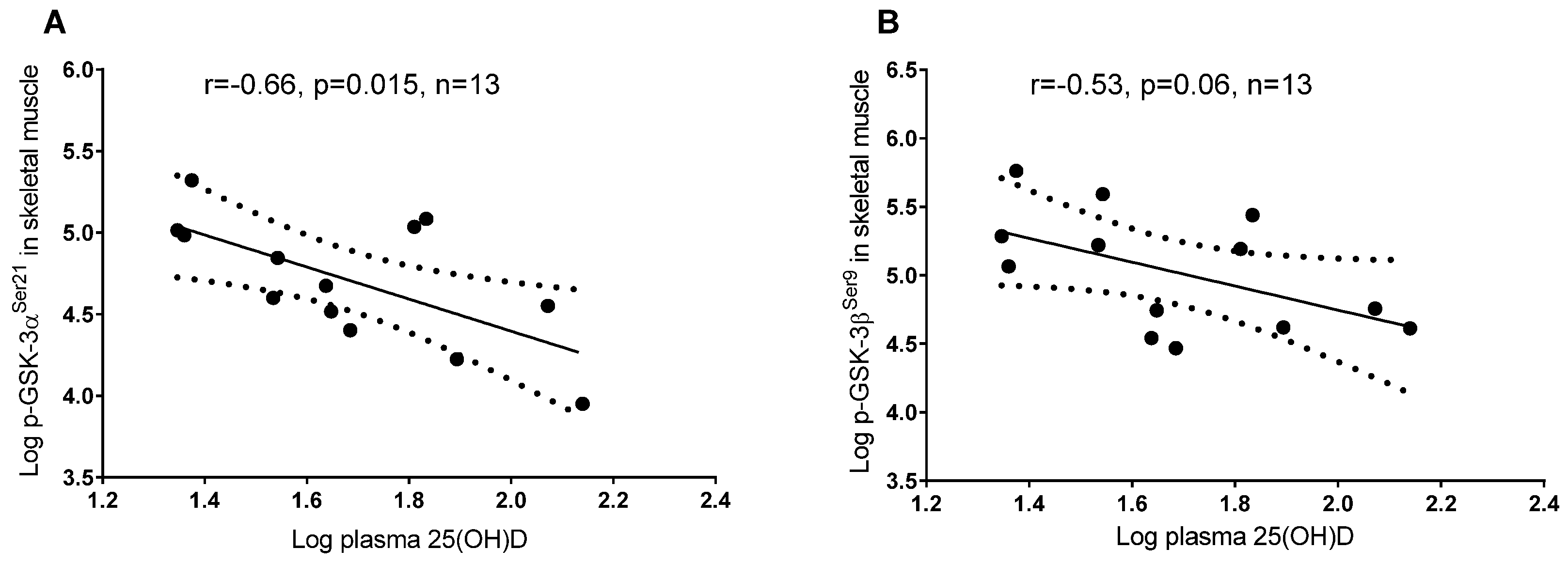
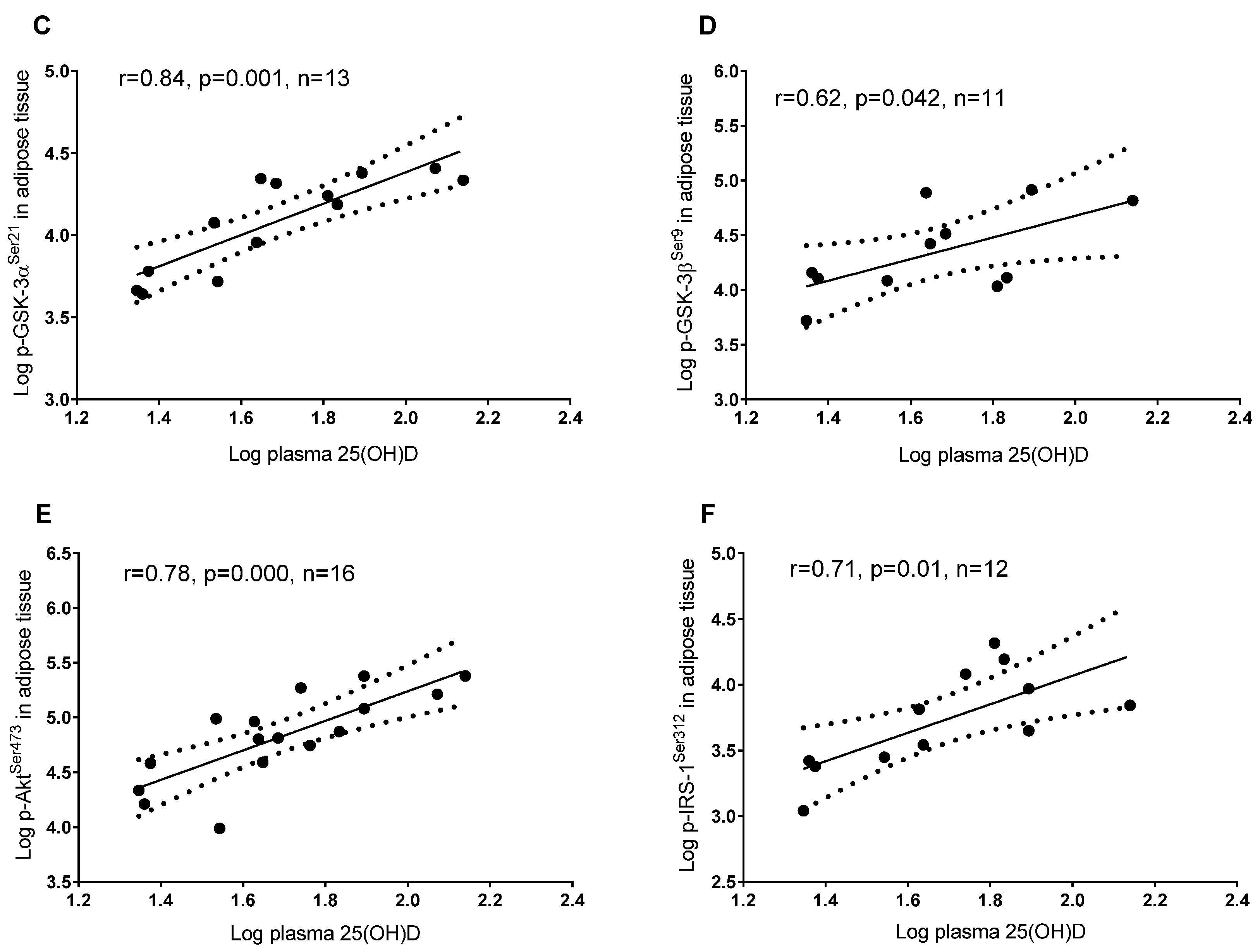
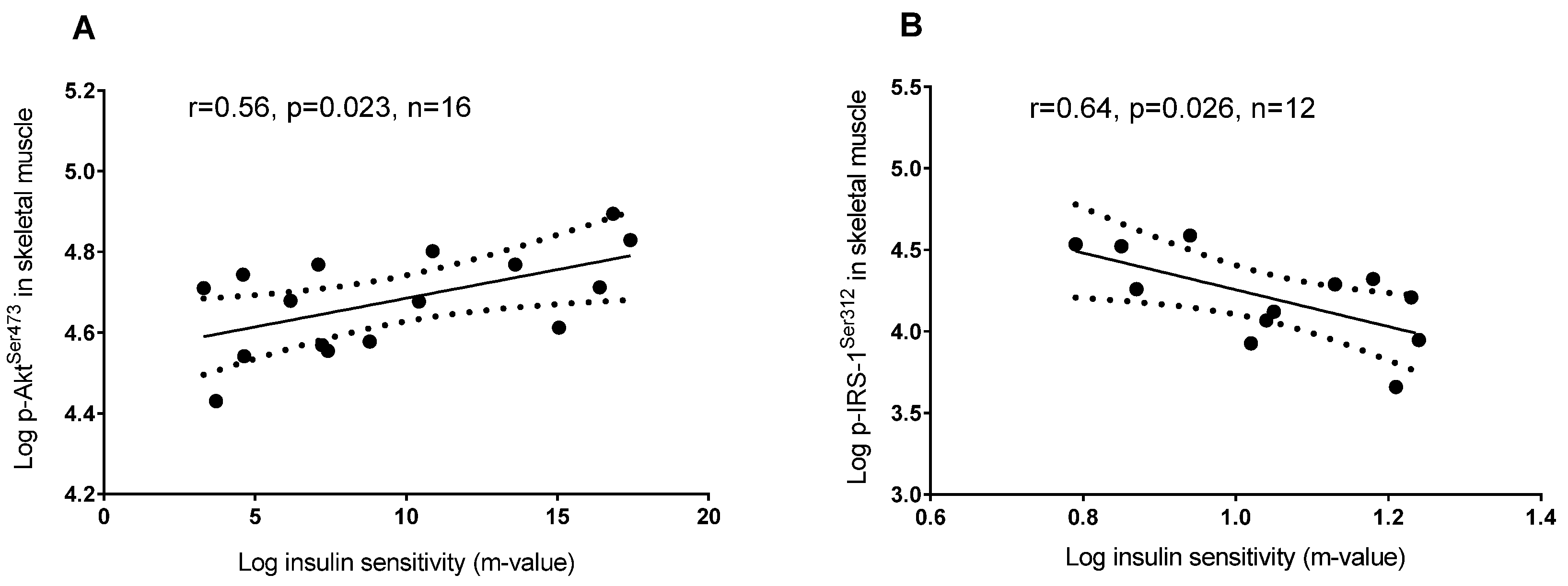
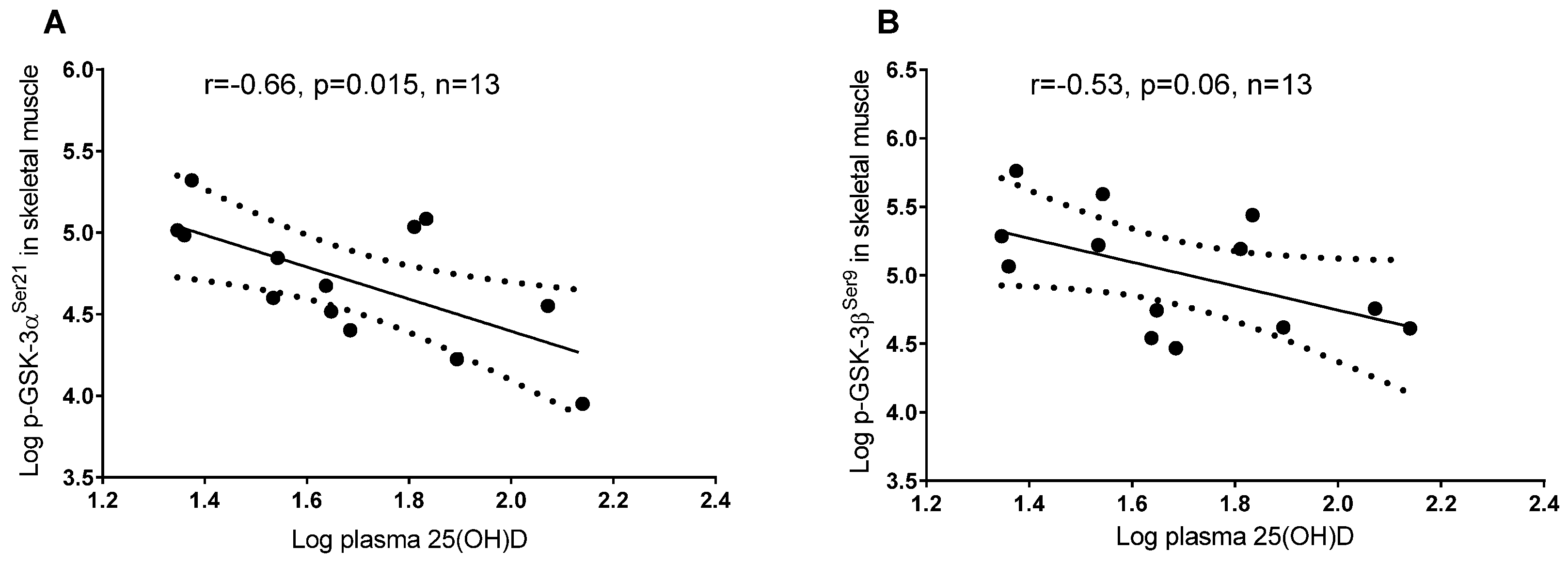
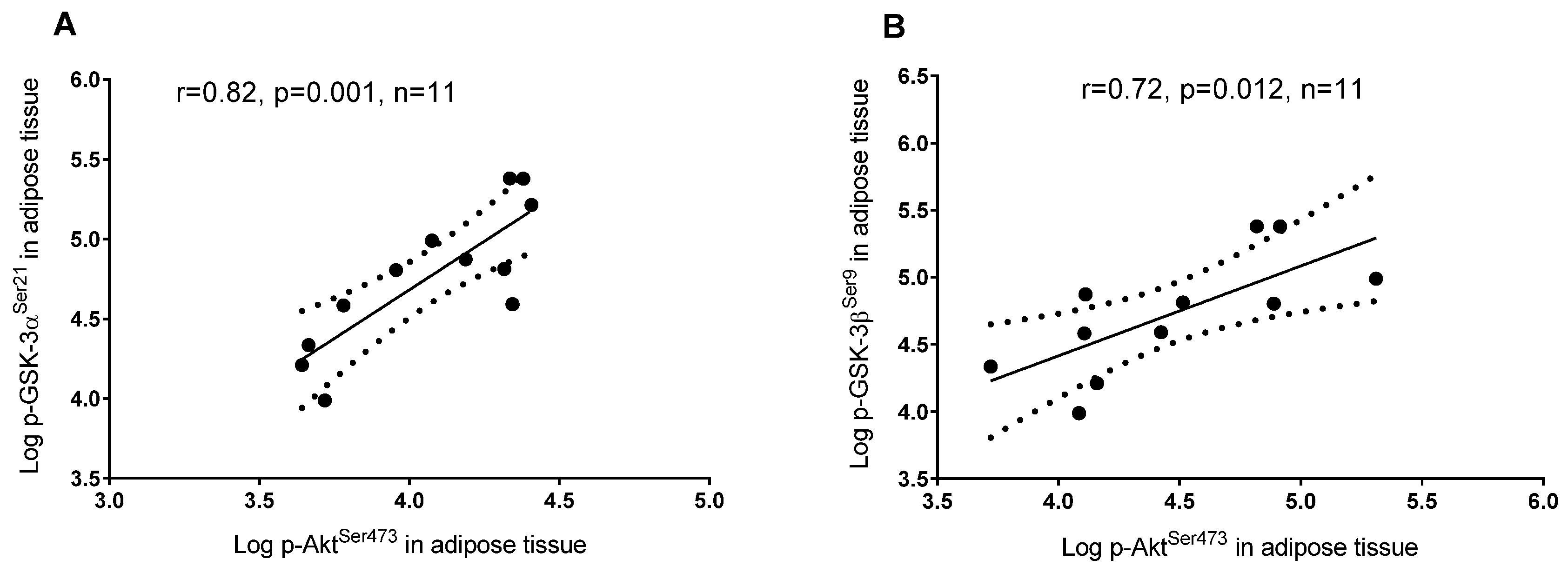
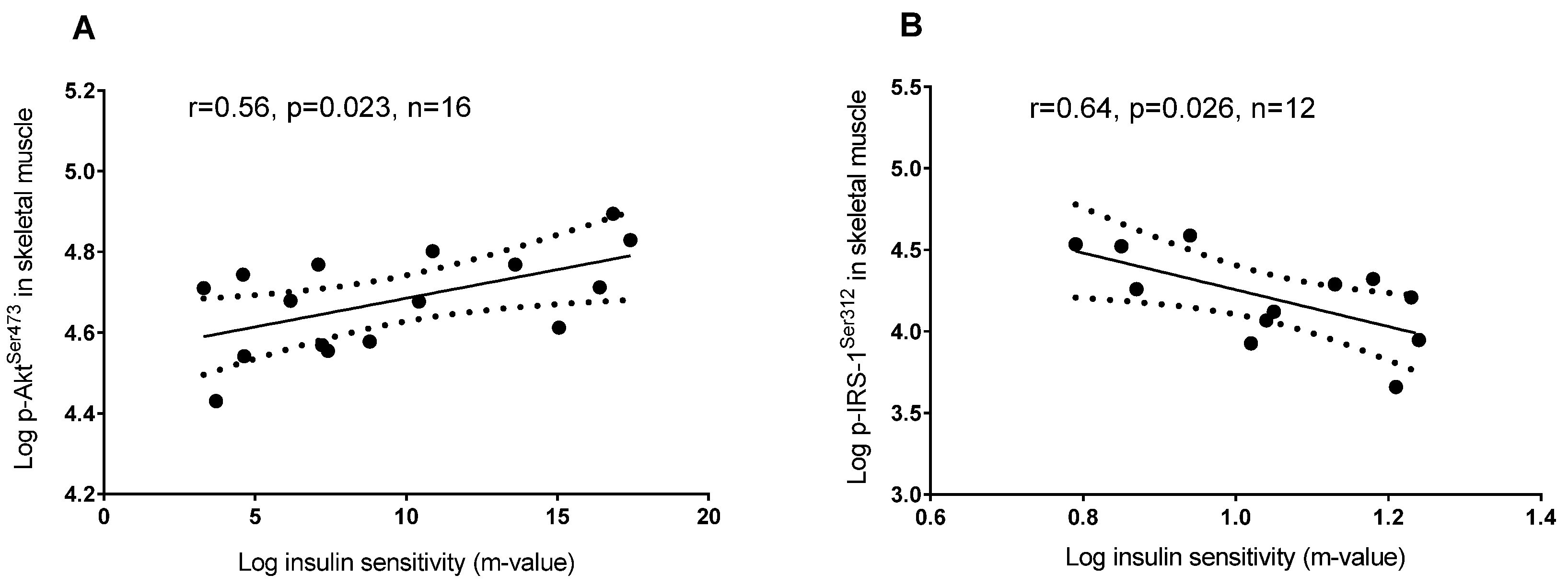
3.2. Plasma 25(OH)D and Protein Regulators of Insulin Signaling
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lips, P. Vitamin D physiology. Prog. Biophys. Mol. Biol. 2006, 92, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.S.; Whiting, S.J.; Barton, C.N. Vitamin D fortification in the United States and Canada: Current status and data needs. Am. J. Clin. Nutr. 2004, 80 (Suppl. 6), 1710S–1716S. [Google Scholar] [PubMed]
- Ovesen, L.; Brot, C.; Jakobsen, J. Food contents and biological activity of 25-hydroxyvitamin D: A vitamin D metabolite to be reckoned with? Ann. Nutr. Metab. 2003, 47, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Christakos, S.; Ajibade, D.V.; Dhawan, P.; Fechner, A.J.; Mady, L.J. Vitamin D: Metabolism. Endocrinol. Metab. Clin. N. Am. 2010, 39, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D status: Measurement, interpretation, and clinical application. Ann. Epidemiol. 2009, 19, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Thacher, T.D.; Clarke, B.L. Vitamin D Insufficiency. Mayo Clin. Proc. 2011, 86, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F.; Chen, T.C. Vitamin D deficiency: A worldwide problem with health consequences. Am. J. Clin. Nutr. 2008, 87, 1080s–1086s. [Google Scholar] [CrossRef] [PubMed]
- Pittas, A.G.; Lau, J.; Hu, F.B.; Dawson-Hughes, B. The role of vitamin D and calcium in type 2 diabetes. A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2007, 92, 2017–2029. [Google Scholar] [CrossRef] [PubMed]
- Al-Shoumer, K.A.; Al-Essa, T.M. Is there a relationship between vitamin D with insulin resistance and diabetes mellitus? World J. Diabetes 2015, 6, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.A.; Ashraf, A. Role of vitamin D in insulin secretion and insulin sensitivity for glucose homeostasis. Int. J. Endocrinol. 2010, 2010, 351385. [Google Scholar] [CrossRef] [PubMed]
- Grimnes, G.; Figenschau, Y.; Almås, B.; Jorde, R. Vitamin D, insulin secretion, sensitivity, and lipids: Results from a case-control study and a randomized controlled trial using hyperglycemic clamp technique. Diabetes 2011, 60, 2748–2757. [Google Scholar] [CrossRef] [PubMed]
- Ljunghall, S.; Lind, L.; Lithell, H.; Skarfors, E.; Selinus, I.; Sørensen, O.H.; Wide, L. Treatment with one-alpha-hydroxycholecalciferol in middle-aged men with impaired glucose-tolerance—A prospective randomized double-blind-study. Acta Med. Scand. 1987, 222, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Simha, V.; Mahmood, M.; Ansari, M.; Spellman, C.W.; Shah, P. Effect of vitamin D replacement on insulin sensitivity in subjects with vitamin D deficiency. J. Investig. Med. 2012, 60, 1214–1218. [Google Scholar] [CrossRef] [PubMed]
- Maestro, B.; Molero, S.; Bajo, S.; Dávila, N.; Calle, C. Transcriptional activation of the human insulin receptor gene by 1,25-dihydroxyvitamin D-3. Cell Biochem. Funct. 2002, 20, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Leitner, W.; Reusch, J.E.; Sussman, K.E.; Draznin, B. GLUT-4 phosphorylation and its intrinsic activity. Mechanism of Ca(2+)-induced inhibition of insulin-stimulated glucose transport. J. Biol. Chem. 1993, 268, 3352–3356. [Google Scholar] [PubMed]
- Begum, N.; Sussman, K.E.; Draznin, B. Calcium-induced inhibition of phosphoserine phosphatase in insulin target-cells is mediated by the phosphorylation and activation of inhibitor-1. J. Biol. Chem. 1992, 267, 5959–5963. [Google Scholar] [PubMed]
- Reusch, J.E.; Begum, N.; Sussman, K.E.; Draznin, B. Regulation of GLUT-4 phosphorylation by intracellular calcium in adipocytes. Endocrinology 1991, 129, 3269–3273. [Google Scholar] [CrossRef] [PubMed]
- Alkharfy, K.M.; Al-Daghri, N.M.; Yakout, S.M.; Hussain, T.; Mohammed, A.K.; Krishnaswamy, S. Influence of vitamin D treatment on transcriptional regulation of insulin-sensitive genes. Metab. Syndr. Relat. Disord. 2013, 11, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.G.; Hou, F.F.; Guo, Z.J.; Liang, M.; Wang, G.B.; Zhang, X. 1,25-Dihydroxyvitamin D improved the free fatty-acid-induced insulin resistance in cultured C2C12 cells. Diabetes Metab. Res. Rev. 2008, 24, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Buitrago, C.G.; Arango, N.S.; Boland, R.L. 1alpha,25(OH)2D3-dependent modulation of Akt in proliferating and differentiating C2C12 skeletal muscle cells. J. Cell. Biochem. 2012, 113, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Alessi, D.R.; Downes, C.P. The role of PI 3-kinase in insulin action. Biochim. Biophys. Acta 1998, 1436, 151–164. [Google Scholar] [CrossRef]
- Mousa, A.; Naderpoor, N.; de Courten, M.P.J.; Scragg, R.; de Courten, B. 25-hydroxyvitamin D is associated with adiposity and cardiometabolic risk factors in a predominantly vitamin D-deficient and overweight/obese but otherwise healthy cohort. J. Steroid Biochem. Mol. Biol. 2016, in press. [Google Scholar]
- Howlett, K.F.; Sakamoto, K.; Yu, H.; Goodyear, L.J.; Hargreaves, M. Insulin-stimulated insulin receptor substrate-2-associated phosphatidylinositol 3-kinase activity is enhanced in human skeletal muscle after exercise. Metabolism 2006, 55, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.D. Influential Observations in Linear Regression. J. Am. Stat. Assoc. 1979, 74, 169–174. [Google Scholar] [CrossRef]
- World Medical Association. World medical association declaration of helsinki: Ethical principles for medical research involving human subjects. JAMA 2013, 310, 2191–2194. [Google Scholar]
- Fliser, D.; Stefanski, A.; Franek, E.; Fode, P.; Gudarzi, A.; Ritz, E. No effect of calcitriol on insulin-mediated glucose uptake in healthy subjects. Eur. J. Clin. Investig. 1997, 27, 629–633. [Google Scholar] [CrossRef]
- Von Hurst, P.R.; Stonehouse, W.; Coad, J. Vitamin D supplementation reduces insulin resistance in South Asian women living in New Zealand who are insulin resistant and vitamin D deficient—A randomised, placebo-controlled trial. Br. J. Nutr. 2010, 103, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R.; Figenschau, Y. Supplementation with cholecalciferol does not improve glycaemic control in diabetic subjects with normal serum 25-hydroxyvitamin D levels. Eur. J. Nutr. 2009, 48, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Orwoll, E.; Riddle, M.; Prince, M. Effects of vitamin D on insulin and glucagon secretion in non-insulin-dependent diabetes mellitus. Am. J. Clin. Nutr. 1994, 59, 1083–1087. [Google Scholar] [PubMed]
- Witham, M.D.; Dove, F.J.; Dryburgh, M.; Sugden, J.A.; Morris, A.D.; Struthers, A.D. The effect of different doses of vitamin D(3) on markers of vascular health in patients with type 2 diabetes: A randomised controlled trial. Diabetologia 2010, 53, 2112–2119. [Google Scholar] [CrossRef] [PubMed]
- Parekh, D.; Sarathi, V.; Shivane, V.K.; Bandgar, T.R.; Menon, P.S.; Shah, N.S. Pilot study to evaluate the effect of short-term improvement in vitamin D status on glucose tolerance in patients with type 2 diabetes mellitus. Endocr. Pract. 2010, 16, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Pollare, T.; Hvarfner, A.; Lithell, H.; Sørensen, O.H.; Ljunghall, S. Long-term treatment with active vitamin D (alphacalcidol) in middle-aged men with impaired glucose tolerance. Effects on insulin secretion and sensitivity, glucose tolerance and blood pressure. Diabetes Res. 1989, 11, 141–147. [Google Scholar] [PubMed]
- Hoseini, S.A.; Aminorroaya, A.; Iraj, B.; Amini, M. The effects of oral vitamin D on insulin resistance in pre-diabetic patients. J. Res. Med. Sci. 2013, 18, 47–51. [Google Scholar] [PubMed]
- Chang, E.; Donkin, S.S.; Teegarden, D. Parathyroid hormone suppresses insulin signaling in adipocytes. Mol. Cell. Endocrinol. 2009, 307, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; O’Keefe, J.H.; Bell, D.; Hensrud, D.D.; Holick, M.F. Vitamin D deficiency: An important, common, and easily treatable cardiovascular risk factor? J. Am. Coll. Cardiol. 2008, 52, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, C.; Leighton, I.A.; Cohen, P. Inactivation of glycogen synthase kinase-3βby phosphorylation: New kinase connections in insulin and growth-factor signalling. Biochem. J. 1993, 296, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.A.E.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of Glycogen-Synthase Kinase-3 by Insulin-Mediated by Protein-Kinase-B. Nature 1995, 378, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Ciaraldi, T.P.; Oh, D.K.; Christiansen, L.; Nikoulina, S.E.; Kong, A.P.; Baxi, S.; Mudaliar, S.; Henry, R.R. Tissue-specific expression and regulation of GSK-3 in human skeletal muscle and adipose tissue. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E891–E898. [Google Scholar] [CrossRef] [PubMed]
- Grimes, C.A.; Jope, R.S. The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling. Prog. Neurobiol. 2001, 65, 391–426. [Google Scholar] [CrossRef]
- Tanti, J.F.; Jager, J. Cellular mechanisms of insulin resistance: Role of stress-regulated serine kinases and insulin receptor substrates (IRS) serine phosphorylation. Curr. Opin. Pharmacol. 2009, 9, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.L.; Li, Y.; Cama, A.; Quon, M.J. Tyr(612) and Tyr(632) in human insulin receptor substrate-1 are important for full activation of insulin-stimulated phosphatidylinositol 3-kinase activity and translocation of GLUT4 in adipose cells. Endocrinology 2001, 142, 2833–2840. [Google Scholar] [PubMed]
- Prada, P.O.; Zecchin, H.G.; Gasparetti, A.L.; Torsoni, M.A.; Ueno, M.; Hirata, A.E.; Corezola do Amaral, M.E.; Höer, N.F.; Boschero, A.C.; Saad, M.J. Western diet modulates insulin signaling, c-Jun N-Terminal kinase activity, and insulin receptor substrate-1ser307phosphorylation in a tissue-specific fashion. Endocrinology 2005, 146, 1576–1587. [Google Scholar] [CrossRef] [PubMed]
- Krul-Poel, Y.H.M.; Westra, S.; ten Boekel, E.; ter Wee, M.M.; van Schoor, N.M.; van Wijland, H.; Stam, F.; Lips, P.T.; Simsek, S. Effect of vitamin D supplementation on glycemic control in patients with type 2 diabetes (SUNNY Trial): A randomized placebo-controlled trial. Diabetes Care 2015, 38, 1420–1426. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | All | Males | Females |
|---|---|---|---|
| Age (years) | 30 ± 12 (18–49) | 28 ± 10 (18–46) | 33 ± 13 (21–49) |
| Height (cm) | 172 ± 6 (173–199) | 175 ± 5 (167–185) | 167 ± 6 (159–176) |
| Weight (kg) | 78 ± 11 (53–99) | 81 ± 10 (66–99) | 72 ± 12 (53–91) |
| BMI (kg∙m−2) | 26 ± 4 (21–34) | 27 ± 4 (22–34) | 26 ± 4 (21–31) |
| Plasma 25(OH)D (nmol/L) | 57 ± 31 (22–138) | 65 ± 35 (23–138) | 47 ± 20 (22–78) |
| Vitamin D status (25(OH)D) | |||
| Deficient: <25 nmol/L | 3 | 1 | 2 |
| Insufficient: 25–49 nmol/L | 6 | 4 | 2 |
| Sufficient: ≥50 nmol/L | 8 | 5 | 3 |
| GIR per unit of insulin (M-value; mg/kg/min) | 10 ± 5 (3–17) | 9 ± 3 (4–15) | 10 ± 6 (3–17) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parker, L.; Levinger, I.; Mousa, A.; Howlett, K.; De Courten, B. Plasma 25-Hydroxyvitamin D Is Related to Protein Signaling Involved in Glucose Homeostasis in a Tissue-Specific Manner. Nutrients 2016, 8, 631. https://doi.org/10.3390/nu8100631
Parker L, Levinger I, Mousa A, Howlett K, De Courten B. Plasma 25-Hydroxyvitamin D Is Related to Protein Signaling Involved in Glucose Homeostasis in a Tissue-Specific Manner. Nutrients. 2016; 8(10):631. https://doi.org/10.3390/nu8100631
Chicago/Turabian StyleParker, Lewan, Itamar Levinger, Aya Mousa, Kirsten Howlett, and Barbora De Courten. 2016. "Plasma 25-Hydroxyvitamin D Is Related to Protein Signaling Involved in Glucose Homeostasis in a Tissue-Specific Manner" Nutrients 8, no. 10: 631. https://doi.org/10.3390/nu8100631