Allomyrina dichotoma (Arthropoda: Insecta) Larvae Confer Resistance to Obesity in Mice Fed a High-Fat Diet

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of A. dichotoma Larvae (ADL)
2.2. Animals and Diets
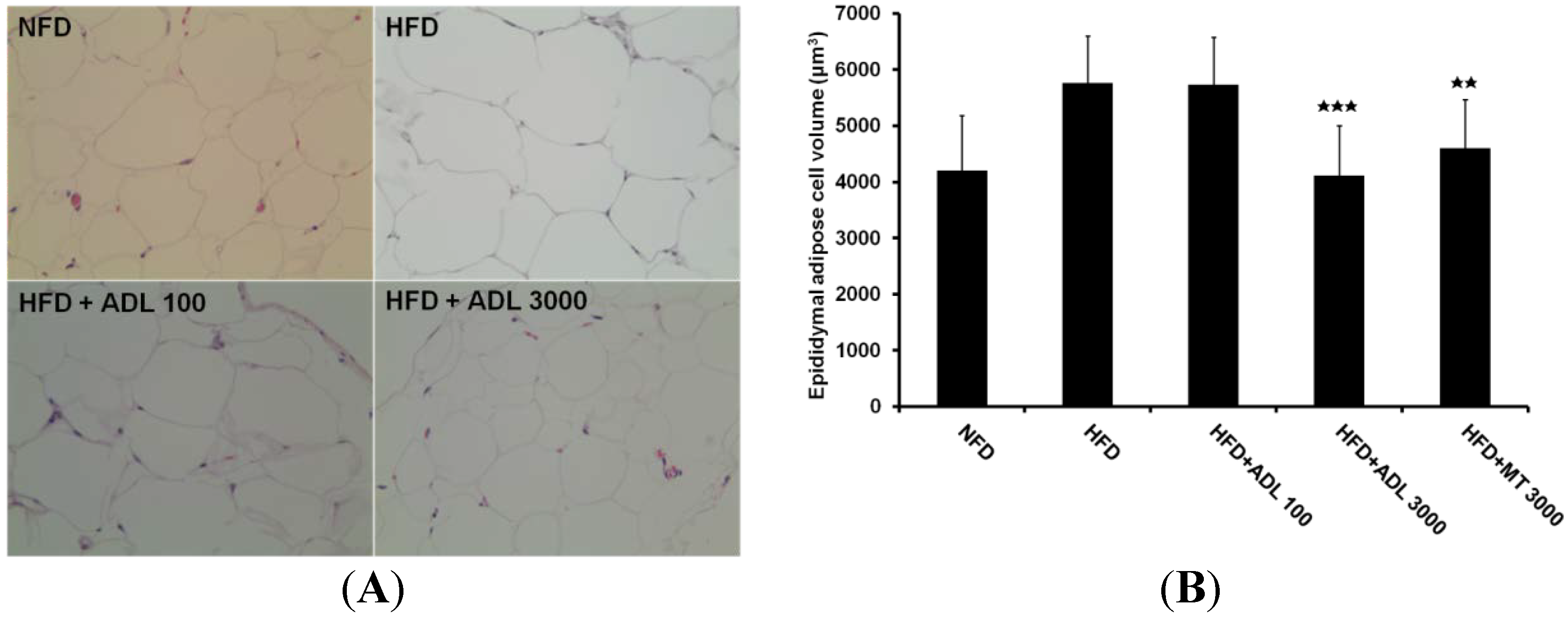
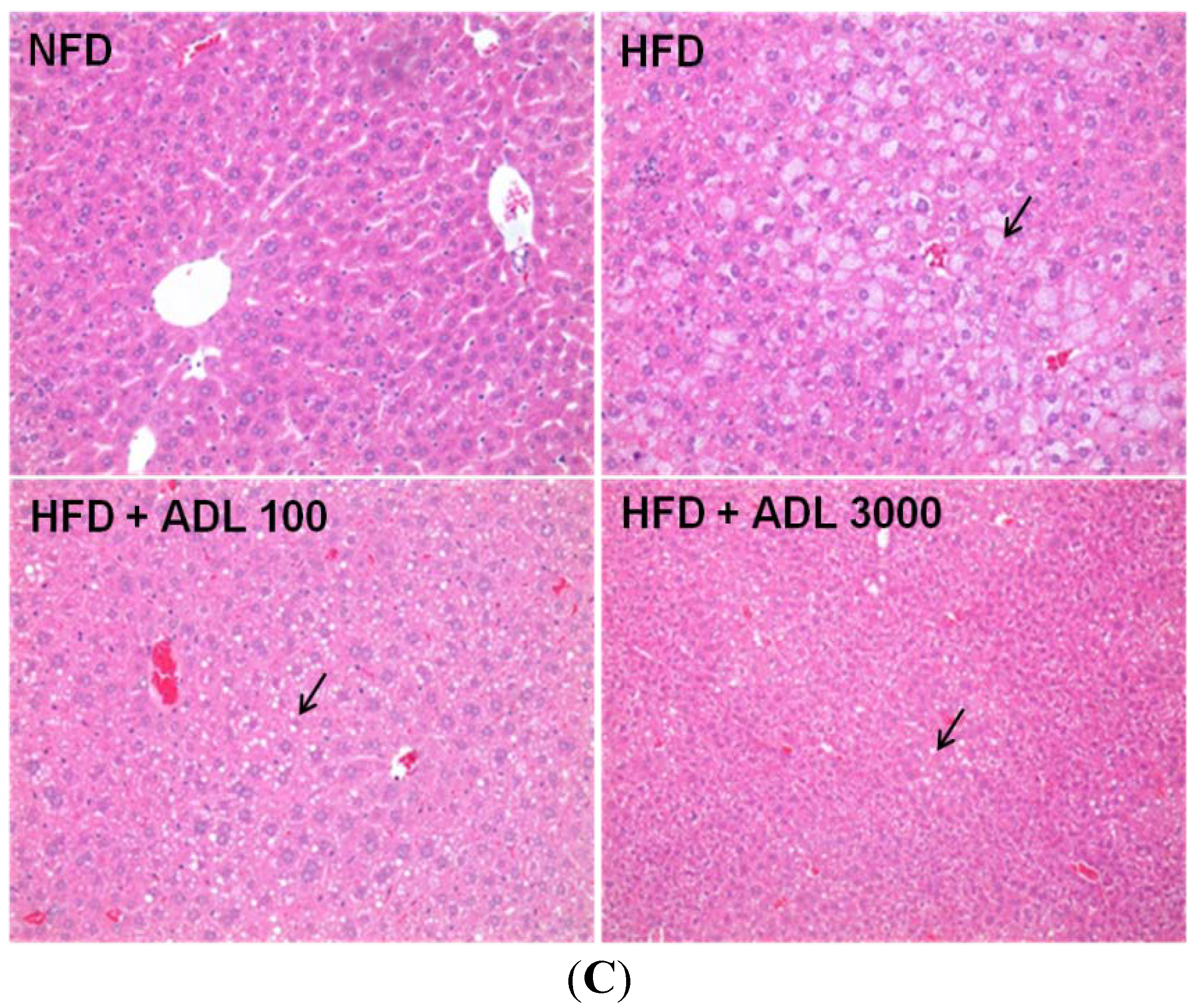
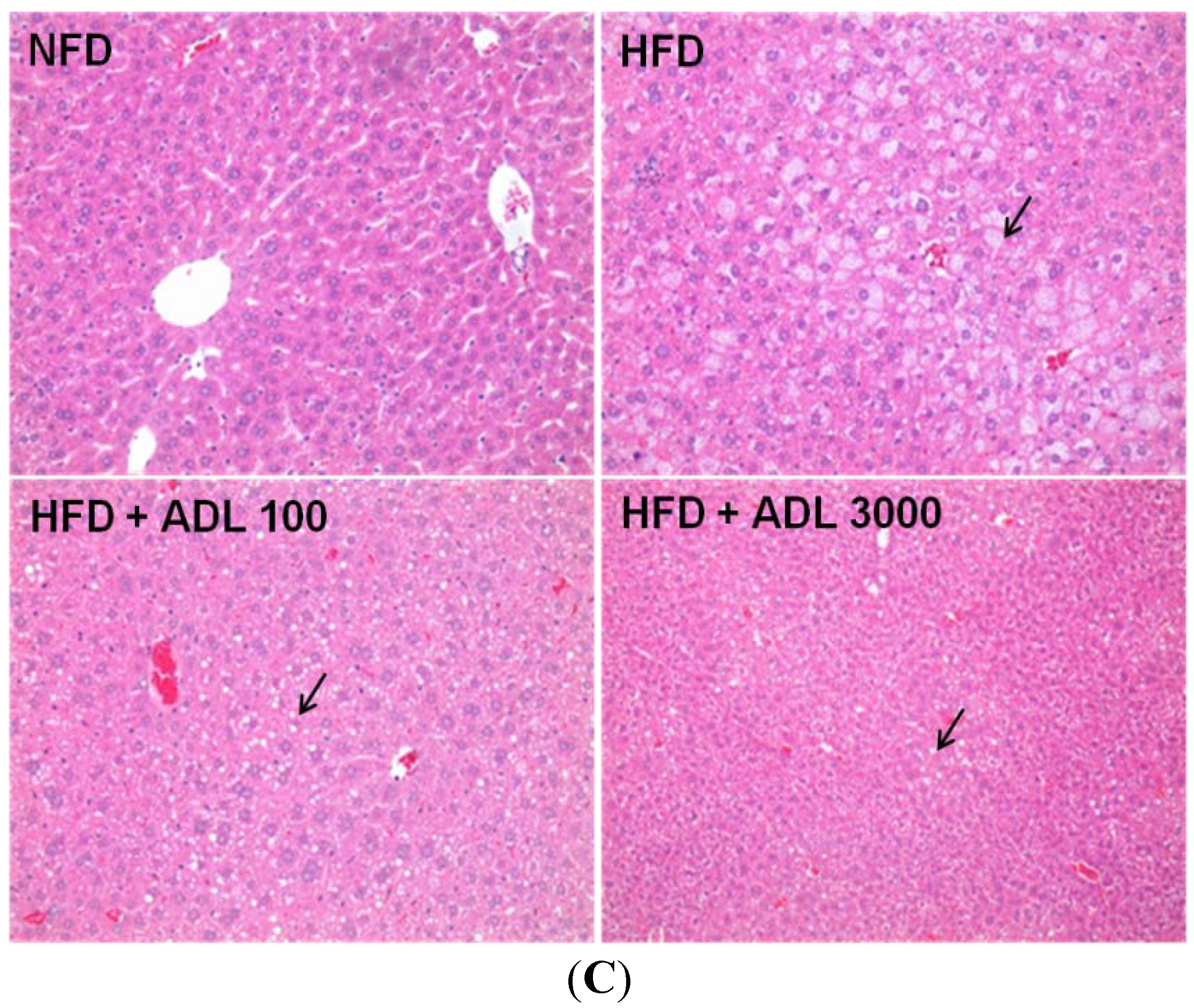
2.3. Histological Analysis
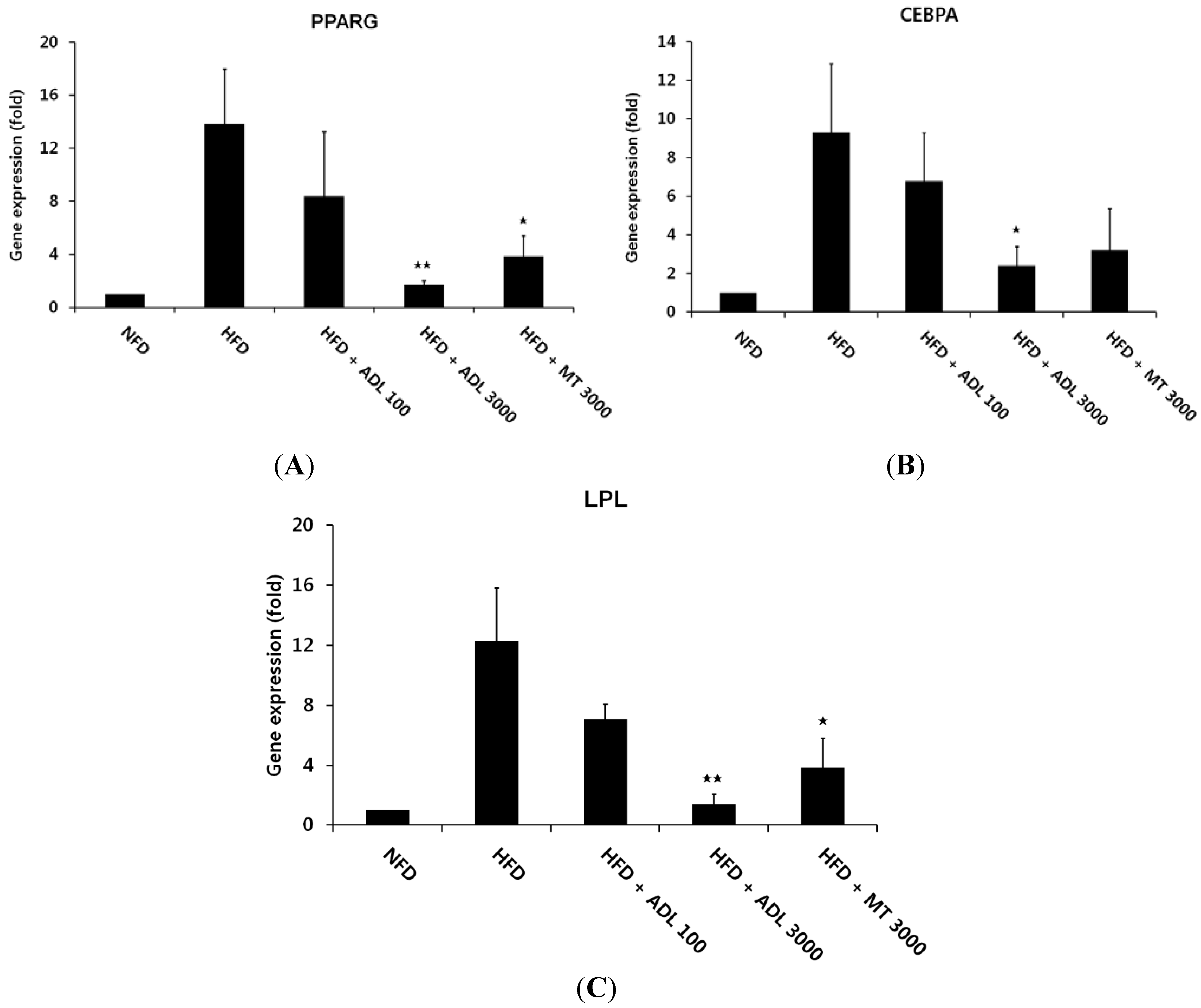
2.4. RNA Preparation and Quantitative Real-Time PCR Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Primer Sequence |
|---|---|---|
| PPARG | Peroxisome proliferator-activated receptor-γ | Forward: 5′-TGGGAACCTGGAAGCTTGTCTC-3′ |
| Reverse: 5′-TGTGGTAAAGGGCTTGATGT-3′ | ||
| C/EBPA | CCAAT/enhancer-binding protein-α | Forward: 5′-TGCCTATGAGCACTTCACAA-3′ |
| Reverse:5′-AACTCCAGCACCTTCTGTTG-3′ | ||
| LPL | Lipoprotein lipase | Forward: 5′-TCCAAGGAAGCCTTTGAGAA-3′ |
| Reverse: 5′-CCATCCTCAGTCCCAGAAAA-3′ | ||
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | Forward: 5′-CTGGAGAAACCTGCCAAGTA-3′ |
| Reverse: 5′-AGTGGGAGTTGCTGTTGAAG-3′ |
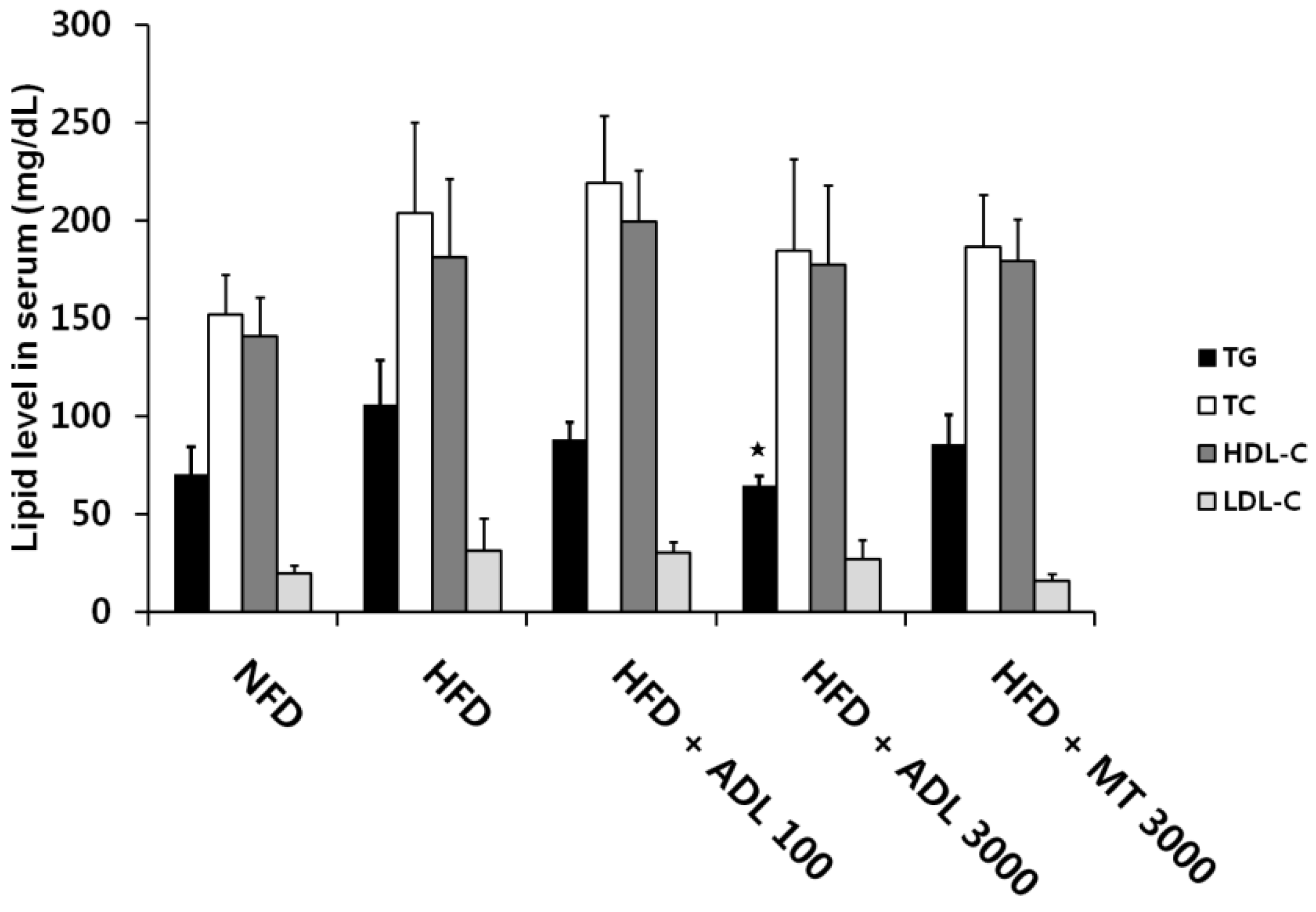
2.5. Biochemical Analysis
2.6. Statistical Analysis
3. Results and Discussion
3.1. Effect of ADL on Body Weight and Adipose Tissue Weight
| Group | Body Weight (g) | Weight Gain (g) | |
|---|---|---|---|
| Initial | Final | ||
| NFD (n = 7) | 32.95 ± 1.33 | 46.07 ± 3.28 | 0.40 ± 0.13 |
| HFD (n = 7) | 32.01 ± 1.37 | 56.53 ± 7.64 | 0.76 ± 0.29 |
| HFD + ADL 100 (n = 7) | 32.26 ± 2.11 | 54.47 ± 7.30 | 0.69 ± 0.29 |
| HFD + ADL 3000 (n = 7) | 31.66 ± 1.20 | 50.31 ± 5.85 | 0.59 ± 0.22 |
| HFD + MT 3000 (n = 7) | 32.10 ± 1.33 | 45.72 ± 4.74 * | 0.42 ± 0.18 * |
| Group | Abdominal-to-Peripheral Fat | Epididymal Fat |
|---|---|---|
| NFD (n = 3) | 3.55 ± 1.5 | 0.177 ± 0.01 |
| HFD (n = 3) | 7.52 ± 1.5 | 0.178 ± 0.03 |
| HFD + ADL 100 (n = 3) | 6.28 ± 1.8 | 0.220 ± 0.04 |
| HFD + ADL 3000 (n = 3) | 4.29 ± 1.8 * | 0.149 ± 0.02 * |
| HFD + MT 3000 (n = 3) | 4.29 ± 0.9 * | 0.185 ± 0.02 |
 p < 0.001 and
p < 0.001 and  p < 0.01, compared to the high-fat diet group.
p < 0.001 and p < 0.01, compared to the high-fat diet group.
p < 0.01, compared to the high-fat diet group.
p < 0.001 and p < 0.01, compared to the high-fat diet group.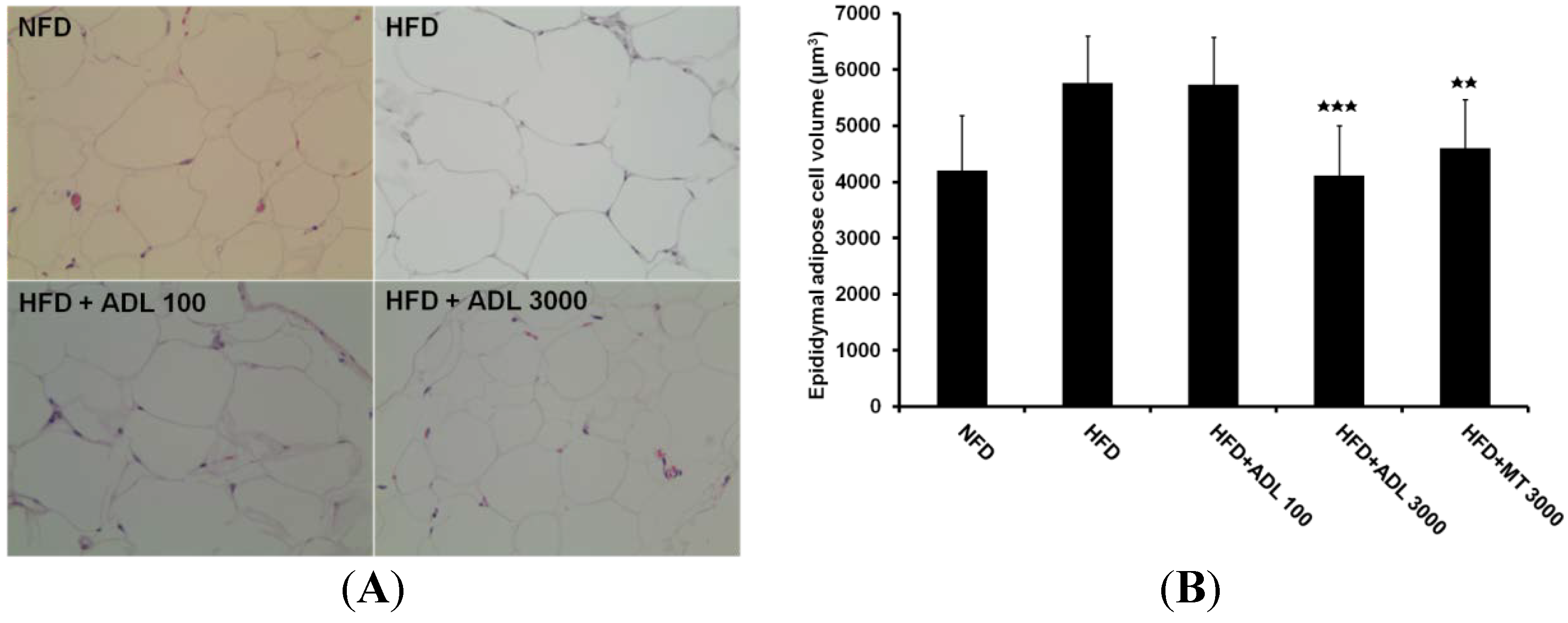
3.2. Inhibitory Effect of ADL on the Expression of Adipogenic and Lipogenic Specific Genes
p < 0.01 and  p < 0.05, compared to the high-fat diet group.
p < 0.01 and p < 0.05, compared to the high-fat diet group.
p < 0.05, compared to the high-fat diet group.
p < 0.01 and p < 0.05, compared to the high-fat diet group.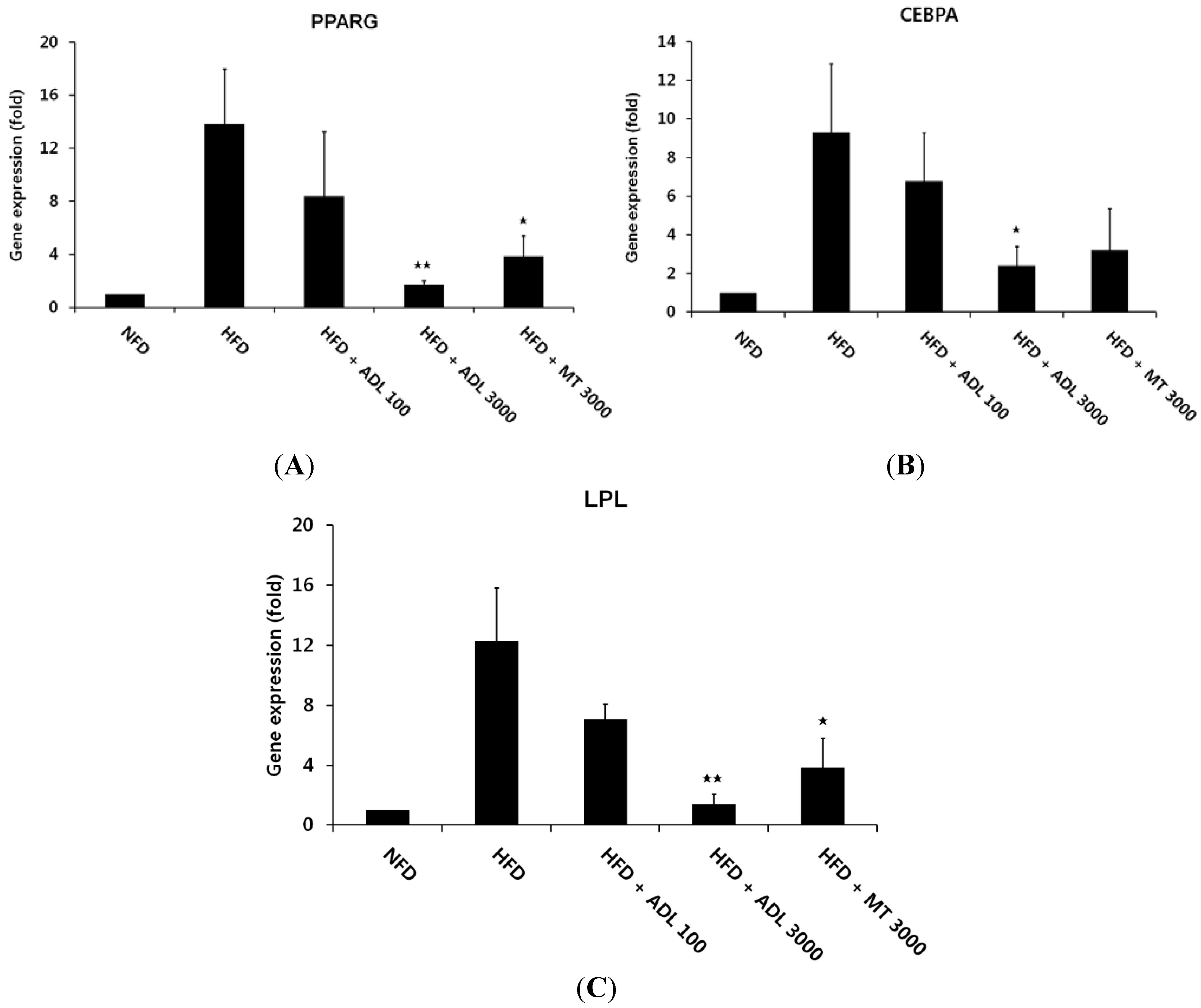
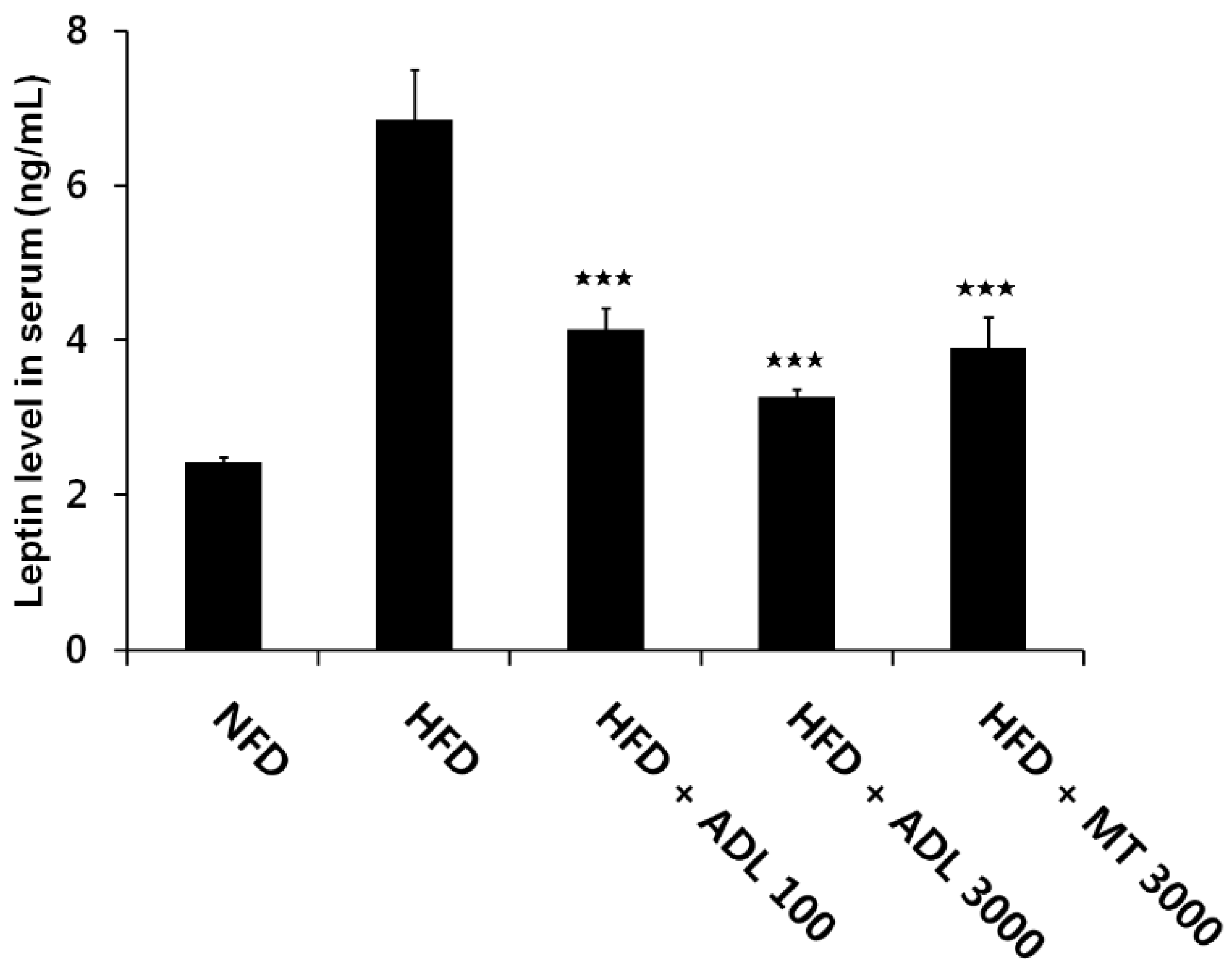
3.3. Effects of ADL on TG and Leptin Levels in Serum of HFD-Induced Obese Mice
p < 0.05, compared to the high-fat diet group.
p < 0.05, compared to the high-fat diet group.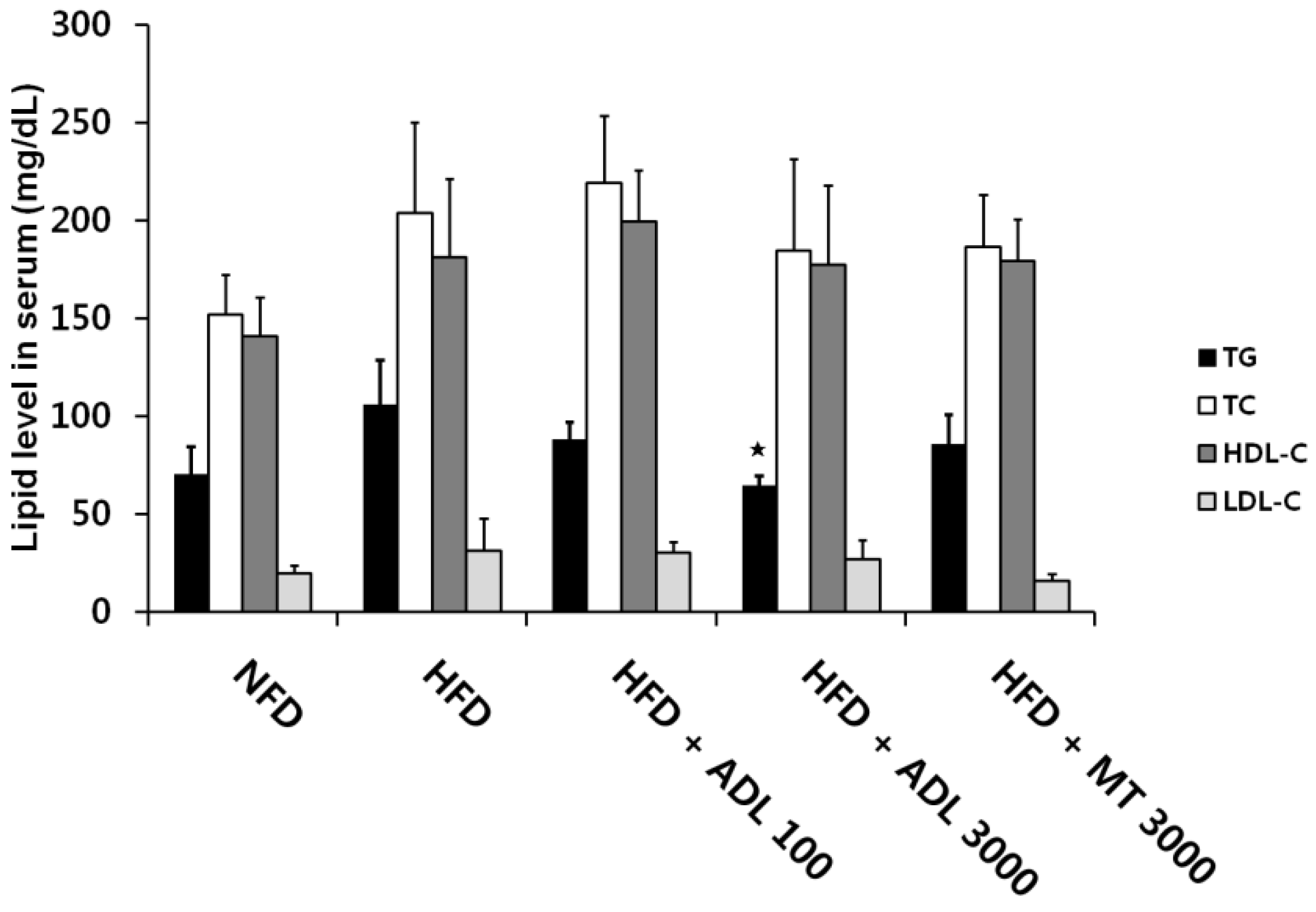 p < 0.001, compared to the high-fat diet group.
p < 0.001, compared to the high-fat diet group.
p < 0.001, compared to the high-fat diet group.
p < 0.001, compared to the high-fat diet group.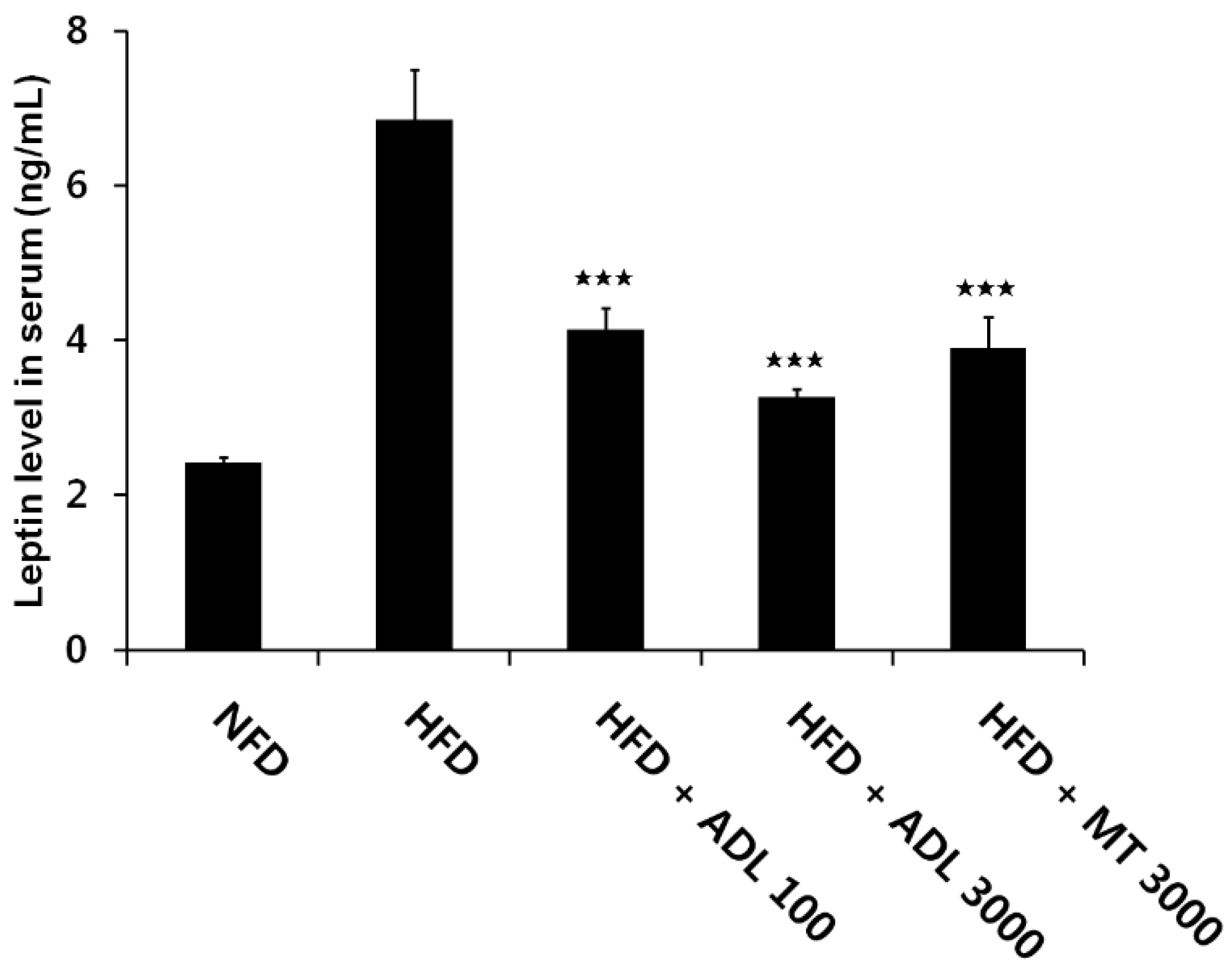
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flier, J.S. Obesity wars: Molecular progress confronts an expanding epidemic. Cell 2004, 116, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Lei, F.; Zhang, X.N.; Wang, W.; Xing, D.M.; Xie, W.D.; Su, H.; Du, L.J. Evidence of anti-obesity effects of the pomegranate leaf extract in high-fat induced obese mice. Int. J. Obes. 2007, 31, 1023–1029. [Google Scholar] [CrossRef]
- Spiegelman, B.M.; Flier, J.S. Adipogenesis and obesity: Rounding out the big picture. Cell 1996, 87, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Cha, B.Y.; Saito, K.; Choi, S.S.; Wang, X.X.; Choi, B.K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.T. Effects of a Citrus depressa Hayata (shiikuwasa) extract on obesity in high-fat diet-induced obese mice. Phytomedicine 2011, 18, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, K.G.; Nicholson, J.R.; Boss, O. The obesity epidemic: Current and future pharmacological treatments. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 565–592. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Nozaki, Y.; Nishizuka, M.; Ikawa, M.; Osada, S.; Imagawa, M. Factor for adipocyte differentiation 158 gene disruption prevents the body weight gain and insulin resistance induced by a high-fat diet. Biol. Pharm. Bull. 2011, 34, 1257–1263. [Google Scholar] [CrossRef]
- Kishino, E.; Ito, T.; Fujita, K.; Kiuchi, Y. A mixture of the Salacia reticulate (Kotala himbutu) aqueous extract and cyclodextrin reduces the accumulation of visceral fat mass in mice and rats with high-fat diet-induced obesity. J. Nutr. 2006, 136, 433–439. [Google Scholar] [PubMed]
- Moro, C.O.; Basile, G. Obesity and medicinal plants. Fitoterapia 2000, 71, 73–82. [Google Scholar] [CrossRef]
- Rayalam, S.; Della-Fera, M.A.; Baile, C.A. Phytochemicals and regulation of the adipocyte life cycle. J. Nutr. Biochem. 2008, 19, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Birgit, A.R.; Oliver, K.S. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A.; van ltterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Sah, L.P.; Jung, C.E. Global perspective of edible insects as human food. Korean J. Soil Zool. 2012, 16, 1–8. [Google Scholar]
- Yoo, J.M.; Hwang, J.S.; Goo, T.W.; Yun, E.Y. Comparative analysis of nutritional and harmful components in Korean and Chinese mealworms (Tenebrio molitor). J. Korean Soc. Food Sci. Nutr. 2013, 42, 249–254. [Google Scholar] [CrossRef]
- Chung, M.Y.; Hwang, J.S.; Goo, T.W.; Yun, E.Y. Analysis of general composition and harmful material of Protaetia brevitarsis. J. Life Sci. 2013, 23, 664–668. [Google Scholar] [CrossRef]
- Miyanoshita, A.; Hara, S.; Sugiyama, M.; Asaoka, A.; Taniai, K.; Yukuhiro, F.; Yamakawa, M. Isolation and characterization of a new member of the insect defensin family from a beetle, Allomyrina dichotoma. Biochem. Biophys. Res. Commun. 1996, 220, 526–531. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, K.; Umetsu, K.; Shinzawa, H.; Yuasa, I.; Maruyama, K.; Ohkura, T.; Yamashita, K.; Suzuki, T. Determination of carbohydrate-deficient transferring separated by lectin affinity chromatography for detecting chronic alcohol abuse. FEBS Lett. 1999, 458, 112–116. [Google Scholar] [CrossRef]
- Sagisaka, A.; Miyanoshita, A.; Ishibashi, J.; Yamakawa, M. Purification, characterization and gene expression of a glycine and proline-rich antibacterial protein family from larvae of a beetle, Allomyrina dichotoma. Insect Mol. Biol. 2001, 10, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.Y.; Yoon, Y.I.; Hwang, J.S.; Goo, T.W.; Yoon, E.Y. Anti-obesity effect of Allomyrinal dichotoma (Arthropoda: Insects) larvae ethanol extract on 27. 3T3-L1 adipocyte differentiation. Entomol. Res. 2014, 44, 9–16. [Google Scholar]
- Green, H.; Kehinde, O. Spontaneous heritable changes leading to increased adipose conversion in 3T3-L1 cells. Cell 1976, 7, 105–113. [Google Scholar] [CrossRef]
- Hansen, P.A.; Han, D.H.; Nolte, L.A.; Chen, M.; Holloszy, J.O. DHEA protects against visceral obesity and muscle insulin resistance in rats fed a high-fat diet. Am. J. Physiol. 1997, 273, R1704–R1708. [Google Scholar]
- Katagiri, K.; Arakawa, S.; Kurahasi, R.; Hatano, Y. Impaired contact hypersensitivity in diet-induced obese mice. J. Dermatol. Sci. 2007, 46, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Supriya, K.; Kotagiri, S.; Vrushabendra Swamy, B.M.; Archana Swamy, P.; Vishwanath, K.M. Anti-obesity activity of Shorea robusta G. leaves extract on high fat diet induced obesity in albino rats. Res. J. Pharm. Biol. Chem. Sci. 2012, 3, 542–554. [Google Scholar]
- Arçari, D.P.; Bartchewsky, W.; dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzoli, J.; de Souza, M.F.; Saad, M.J.; Bastos, D.H.; Gambero, A.; et al. Antiobesity effects of yerba maté extract (Ilex paraguariensis) in high-fat diet-induced obese mice. Obesity 2009, 17, 2127–2133. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Park, T. Genes are differentially expressed in the epididymal fat of rats rendered obese by a high-fat diet. Nutr. Res. 2008, 28, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.D.; Bueno, A.L.; Gallon, C.W.; Gomes, L.F.; Kaiser, S.; Pavei, C.; Ortega, G.G.; Kucharski, L.C.; Jahn, M.P. The effect of aqueous extract of gross and commercial yerba mate (Ilex paraguariensis) on intra-abdominal and epididymal fat and glucose levels in male Wistar rats. Fitoterapia 2011, 82, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Resende, P.E.; Verza, S.G.; Kaiser, S.; Gomes, L.F.; Kucharski, L.C.; Ortega, G.G. The activity of mate saponins (Ilex paraguariensis) in intra-abdominal and epididymal fat, and glucose oxidation in male Wistar rats. J. Ethnopharmacol. 2012, 144, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Flier, J.S. The adipocyte: Storage depot or node on the energy information superhighway? Cell 1995, 80, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Mohamed-Ali, V.; Pinkney, J.H.; Coppack, S.W. Adipose tissue as an endocrine and paracrine organ. Int. Obes. Relat. Metab. Disord. 1998, 22, 1145–1158. [Google Scholar] [CrossRef]
- Dalmas, E.; Clément, K.; Guerre-Millo, M. Defining macrophage phenotype and function in adipose tissue. Trends Immunol. 2011, 32, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Weyer, C.; Foley, J.E.; Bogardus, C.; Tataranni, P.A.; Pratley, R.E. Enlarged subcutaneous abdominal adipocyte size, predicts type II diabetes independent of insulin resistance. Diabetologia 2000, 43, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M. Nonalcoholic steatohepatitis. Semin. Liver Dis. 2004, 24, 3–20. [Google Scholar] [PubMed]
- Brookes, M.J.; Cooper, B.T. Hypertension and fatty liver: Guilty by association? J. Hum. Hypertens. 2007, 2, 264–270. [Google Scholar]
- Youn, K.; Kim, J.Y.; Yeo, H.; Yun, E.Y.; Hwang, J.S.; Jun, M. Fatty acid and volatile oil compositions of Allomyrina dichotoma larvae. Prev. Nutr. Food Sci. 2012, 17, 310–314. [Google Scholar] [CrossRef]
- Pagliosa, C.M.; de Simas, K.N.; Amboni, R.D.M.C.; Murakami, A.N.N.; Petkowicz, C.L.O.; Medeiros, J.D.D.; Rodrigues, A.C.; Amante, E.R. Characterization of the bark from residues from mate tree harvesting (Ilex paraguariensis St. Hil.). Ind. Crops Prod. 2010, 32, 428–433. [Google Scholar] [CrossRef]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [PubMed]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Gwon, S.Y.; Ahn, J.Y.; Kim, T.W.; Ha, T.Y. Zanthoxylum piperitum DC ethanol extract suppresses fat accumulation in adipocytes and high fat diet-induced obese mice by regulating adipogenesis. J. Nutr. Sci. Vitaminol. 2012, 58, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Choi, H.S.; Jeong, E.; Choi, B.D.; Lee, M.Y. Effect of meal pattern on lipogenesis and lipogenic enzyme activity in rat adipose tissue fed high fat diet. J. Korean. Soc. Food Sci. Nutr. 2006, 35, 335–343. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, D.; Yang, G.; Bae, D.K.; Yang, Y.H.; Kim, T.K.; Kim, D.; Kyung, J.; Yeon, S.; Koo, K.C.; et al. Silk and silkworm pupa peptides suppress adipogenesis in preadipocytes and fat accumulation in rats fed a high-fat diet. Eur. J. Nutr. 2012, 51, 1011–1019. [Google Scholar]
- Fasshauer, M.; Paschke, R. Regulation of adipocytokines and insulin resistance. Diabetologia 2003, 46, 1594–1603. [Google Scholar] [CrossRef] [PubMed]
- Mapfei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S.; et al. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Halaas, J.L. Leptin and the regulation of body weight in mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, H.K. Perilla leaf extract ameliorates obesity and dyslipidemia induced by high-fat diet. Phytother. Res. 2009, 23, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.P.; Wang, M.L.; Chan, M.H.; Chiu, Y.S.; Chen, Y.H. Antiobesity activities of indole-3-carbinol in high-fat-diet–induced obese mice. Nutrition 2011, 27, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.W.; Song, M.; Park, J.; Park, S.W.; Kim, N.H.; Gaire, B.P.; Choi, H.Y.; Kim, H. Anti-obesity effect of HT048, a herbal combination, in high fat diet-induced obese rats. Molecules 2012, 17, 14765–14777. [Google Scholar] [CrossRef] [PubMed]
- Ferrannini, E.; Natali, A.; Capaldo, B.; Lehtovirta, M.; Jacob, S.; Yki-Jarvinen, H. Insulin resistance, hyperinsulinemia, and blood pressure: Role of age and obesity. European Group for the Study of Insulin Resistance (EGIR). Hypertension 1997, 30, 1144–1149. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, W.Y.; Bergstrom, R.W.; Boyko, E.J.; Chen, K.W.; Leonetti, D.L.; Newell-Morris, L.; Shofer, J.B.; Wahl, P.W. Visceral adiposity and incident coronary heart disease in Japanese-American men. The 10-year follow-upresults of the Seattle Japanese-American Community Diabetes Study. Diabetes Care 1999, 22, 1808–1812. [Google Scholar] [CrossRef] [PubMed]
- Pitombo, C.; Araújo, E.P.; de Souza, C.T.; Pareja, J.C.; Geloneze, B.; Velloso, L.A. Amelioration of diet-induced diabetes mellitus by removal of visceral fat. J. Endocrinol. 2006, 191, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Barzilai, N.; She, L.; Liu, B.Q.; Vuguin, P.; Cohen, P.; Wang, J.; Rossetti, L. Surgical removal of visceral fat reverses hepatic insulin resistance. Diabetes 1999, 48, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Kim, J.Y.; Lee, S.K. Surgical removal of visceral fat decreases plasma free fatty acid and increases insulin sensitivity on liver and peripheral tissue in monosodium glutamate (MSG)-obese rats. J. Korean Med. Sci. 1999, 14, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Gabriely, I.; Ma, X.H.; Yang, X.M.; Atzmon, G.; Rajala, M.W.; Berg, A.H.; Scherer, P.; Rossetti, L.; Barzilai, N. Removal of visceral fat prevents insulin resistance and glucose intolerance of aging: An adipokine-mediated process? Diabetes 2002, 51, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, Y.-I.; Chung, M.Y.; Hwang, J.-S.; Han, M.S.; Goo, T.-W.; Yun, E.-Y. Allomyrina dichotoma (Arthropoda: Insecta) Larvae Confer Resistance to Obesity in Mice Fed a High-Fat Diet. Nutrients 2015, 7, 1978-1991. https://doi.org/10.3390/nu7031978
Yoon Y-I, Chung MY, Hwang J-S, Han MS, Goo T-W, Yun E-Y. Allomyrina dichotoma (Arthropoda: Insecta) Larvae Confer Resistance to Obesity in Mice Fed a High-Fat Diet. Nutrients. 2015; 7(3):1978-1991. https://doi.org/10.3390/nu7031978
Chicago/Turabian StyleYoon, Young-Il, Mi Yeon Chung, Jae-Sam Hwang, Myung Sae Han, Tae-Won Goo, and Eun-Young Yun. 2015. "Allomyrina dichotoma (Arthropoda: Insecta) Larvae Confer Resistance to Obesity in Mice Fed a High-Fat Diet" Nutrients 7, no. 3: 1978-1991. https://doi.org/10.3390/nu7031978