Increased Intraepithelial Vα24 Invariant NKT Cells in the Celiac Duodenum

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Biopsy Samples

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Patients | n | Mean age (Range) | Gender | HLA DQ2/DQ8 | IgA anti- tTG/EMA | Marsh Criteria at Diagnosis | GFD | |
|---|---|---|---|---|---|---|---|---|
| Celiac Patients | uCD | 45 | 27.1 (4–76) | 36% males | + | + | II-III | No |
| GFD-CD | 15 | 34.2 (4–71) | 34% males | + | - | II-III | Yes | |
| Non Celiac Patients | I-controls | 15 | 42.1 (15–78) | 56% males | +/- | - (*A) | 0-I (M.M) | No |
| C-controls | 44 | 35.3 (6–81) | 41% males | - (*B) | - (*A) | 0 | No |
2.2. Quantitative PCR
| Molecule | Primers Sequence | NCBI Locus | Annealing T |
|---|---|---|---|
| βactin | fw: 5′ - ATG GGT CAG AAG GAT TCC TAT GTG - 3′ rv: 5′ - CTT CAT GAG GTA GTC AGT CAG GTC - 3′ | NM_001101.3 | 60 |
| IFNγ | fw: 5′ - TGG AAA GAG GAG AGT GAC AG - 3′ rv: 5′ - ATT CAT GTC TTC CTT GAT GG - 3′ | NM_000619.2 | 60 |
| Vα24-Jα18 | fw: 5′ - CTG GAG GGA AAG AAC TGC - 3′ rv: 5′ - TGT CAG GGA AAC AGG ACC - 3′ | NC_000014.9 | 65 |
| FoxP3 | fw: 5′ - CAG CAC ATT CCC AGA GTT CCT C - 3′ rv: 5′ - GCG TGT GAA CCA GTG GTA GAT C - 3′ | NM_014009.3 | 60 |
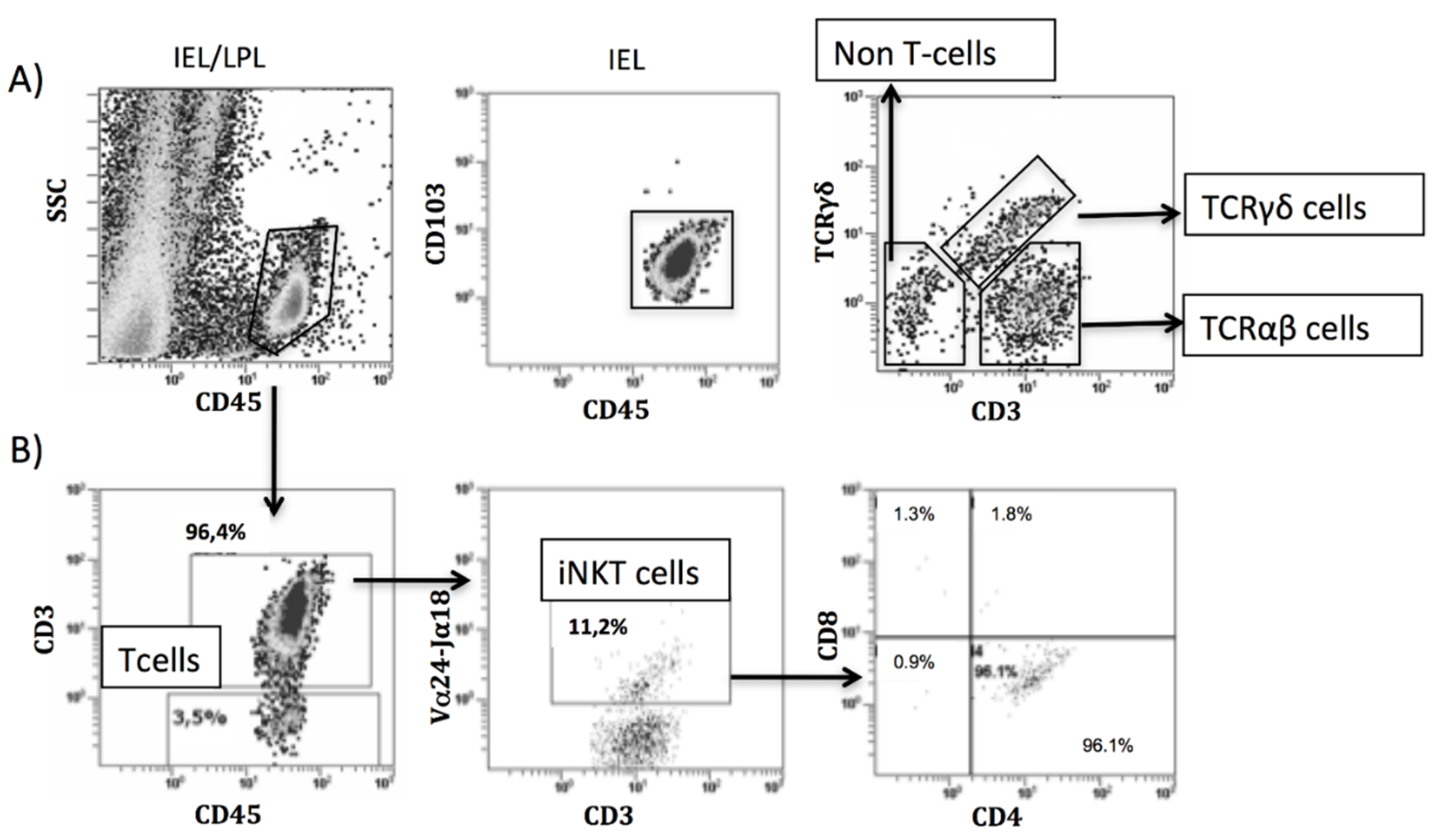
2.3. Isolation of Intraepithelial Lymphocytes and Lamina Propria Mononuclear Cells
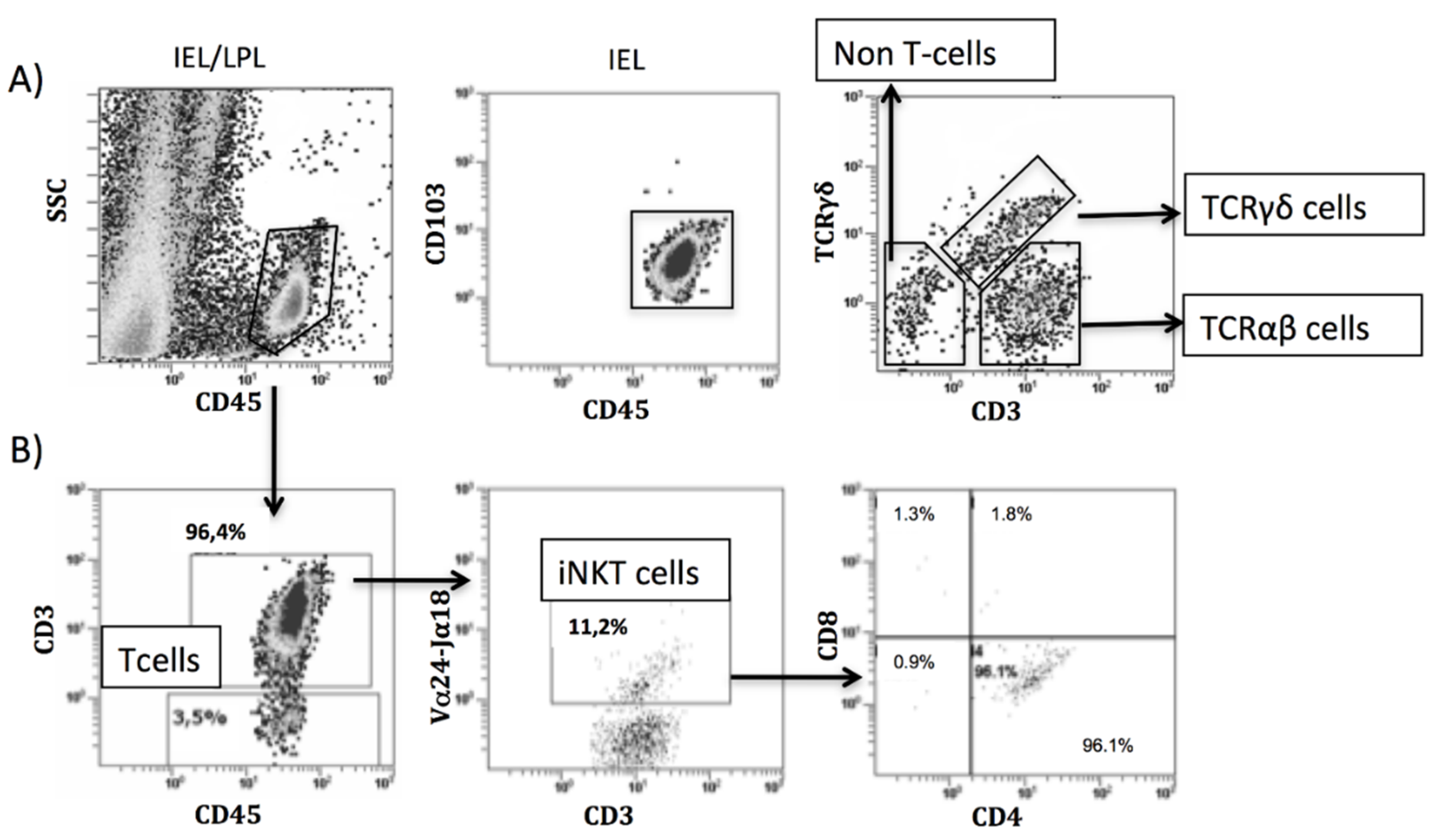
2.4. Antibody Labeling and Flow Cytometry Analysis
2.5. Statistical Analysis
3. Results
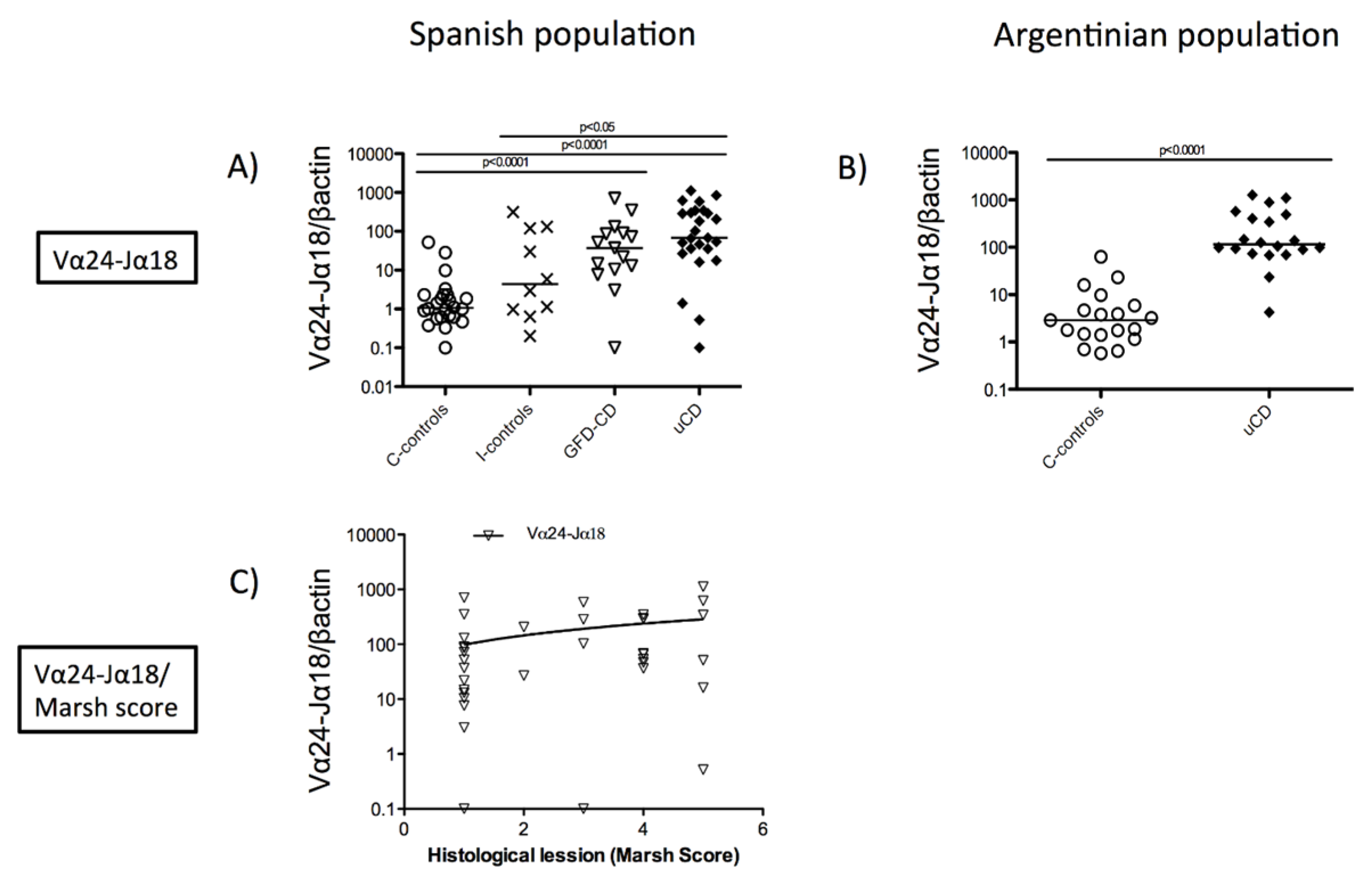
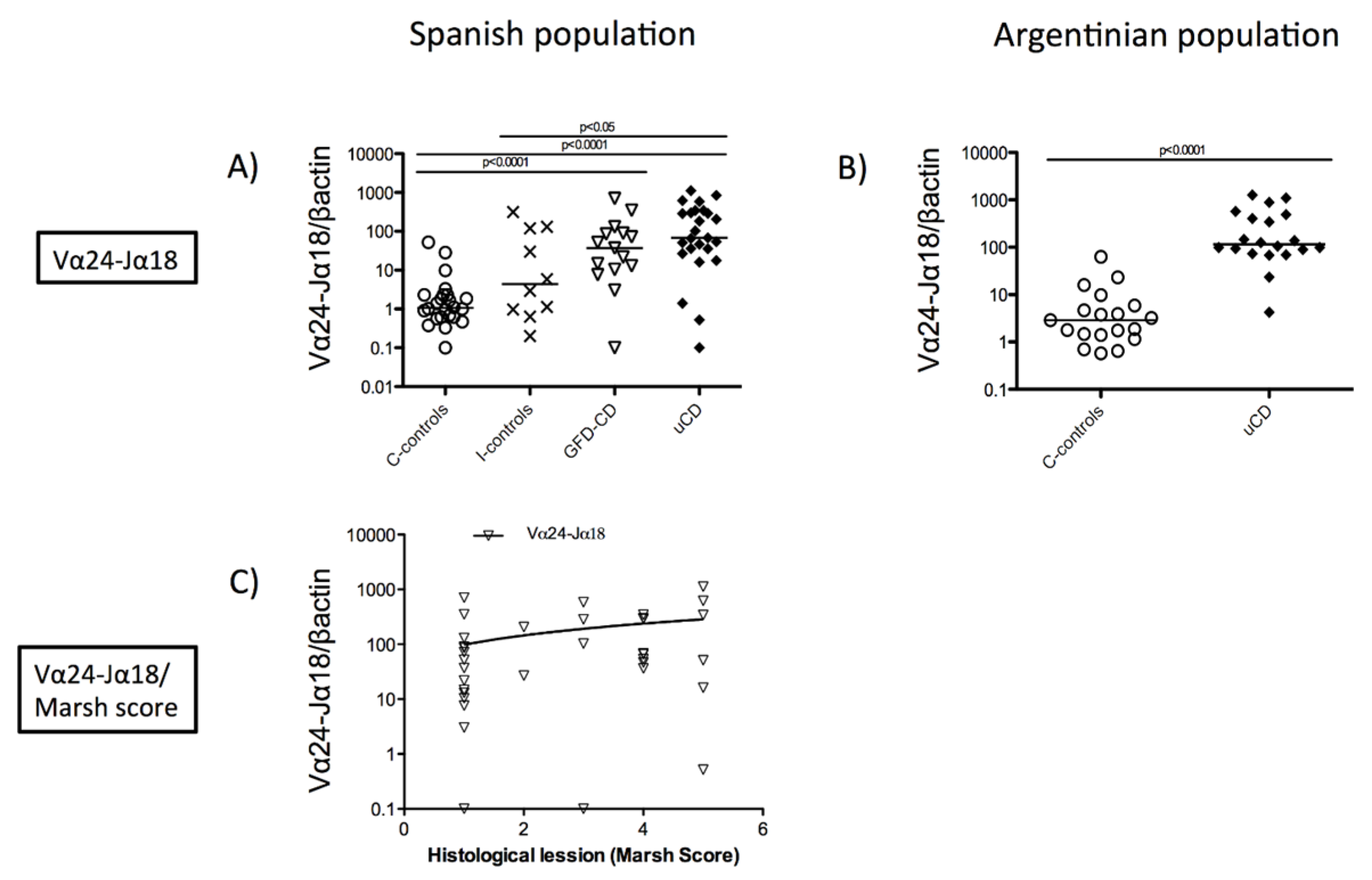
3.1. Increased Duodenal Vα24-Jα18 mRNA Expression in Celiac Disease Patients
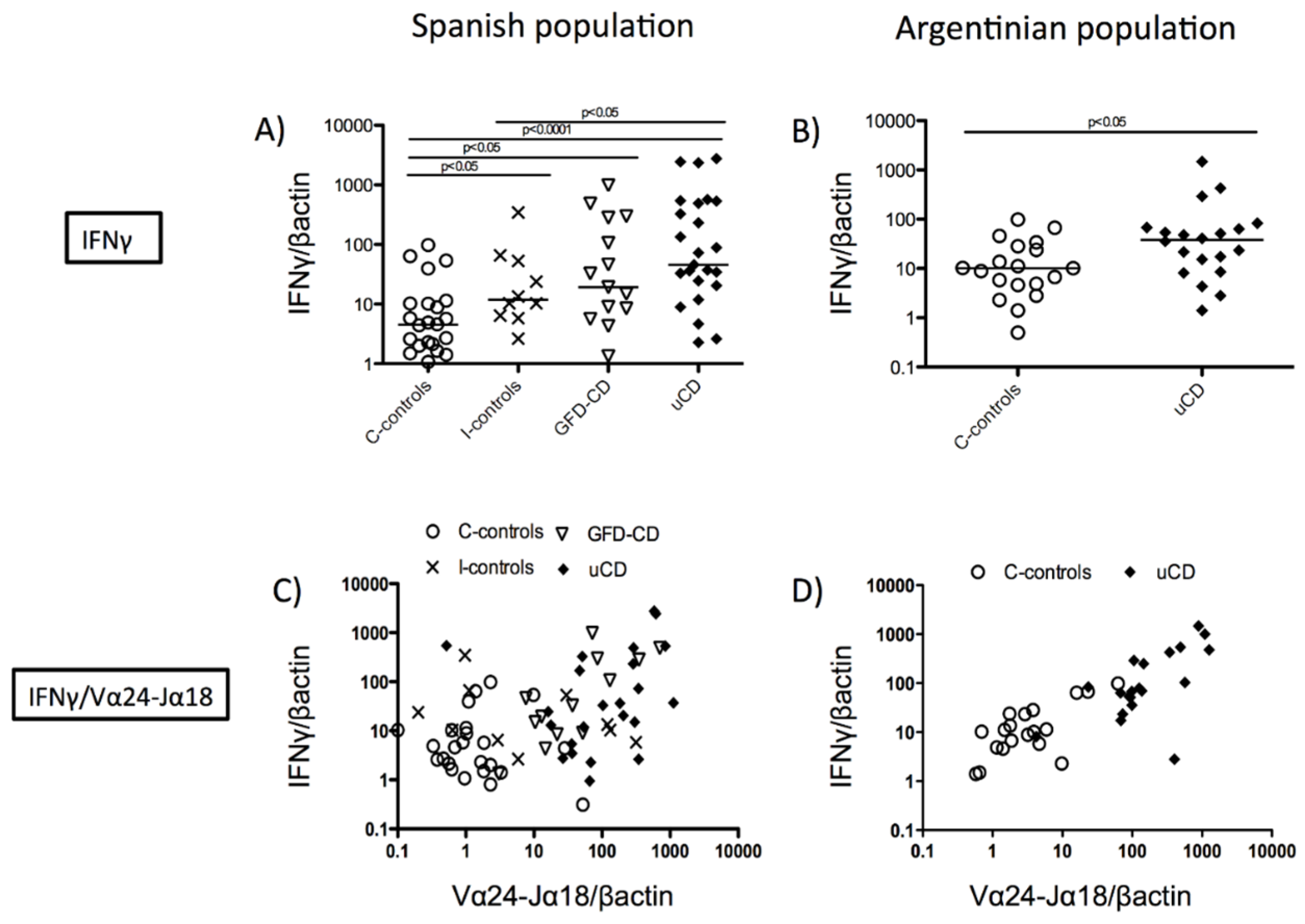
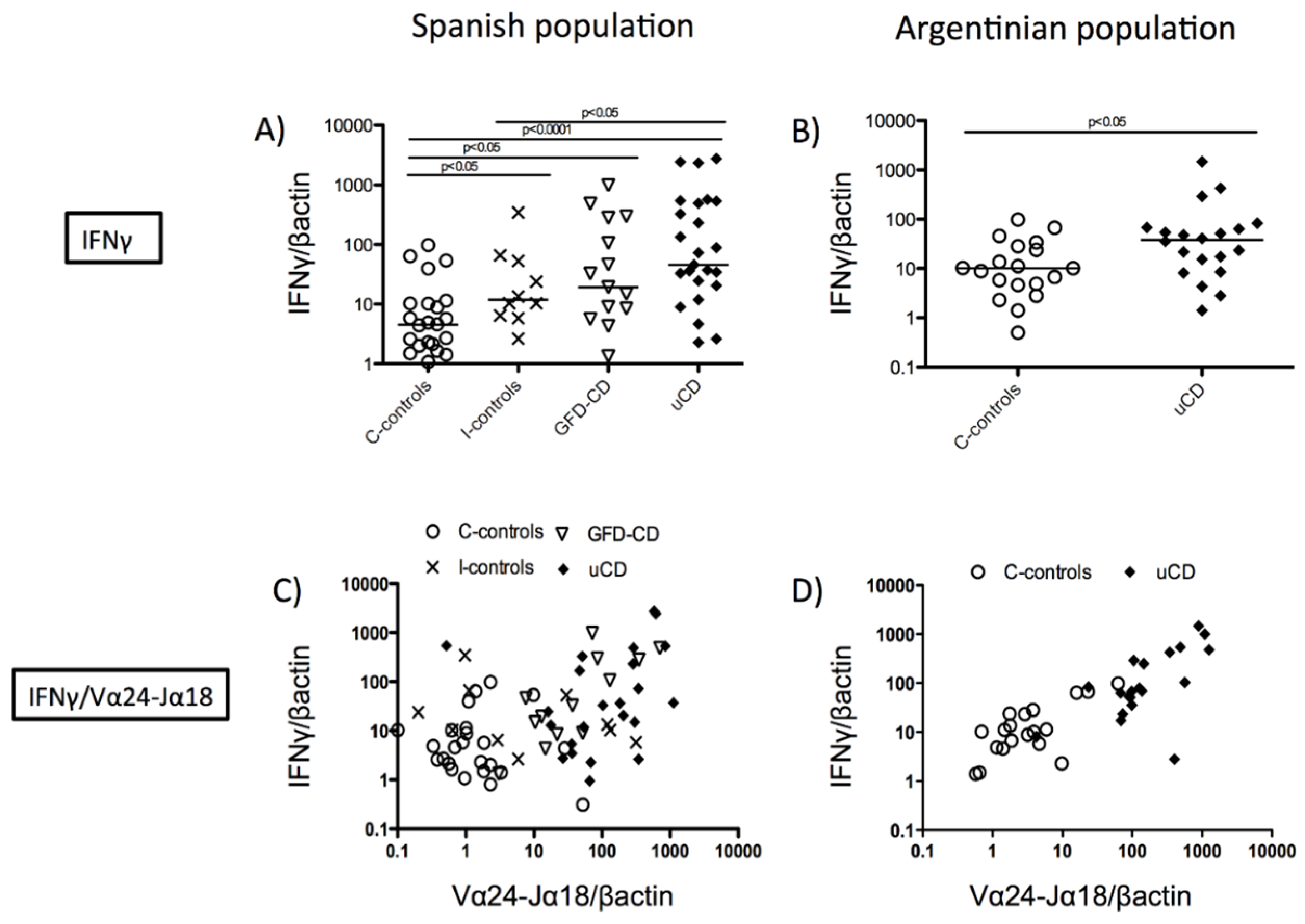
3.2. Correlation between Duodenal Vα24-Jα18 and IFNγ mRNA Expression in Celiac Disease
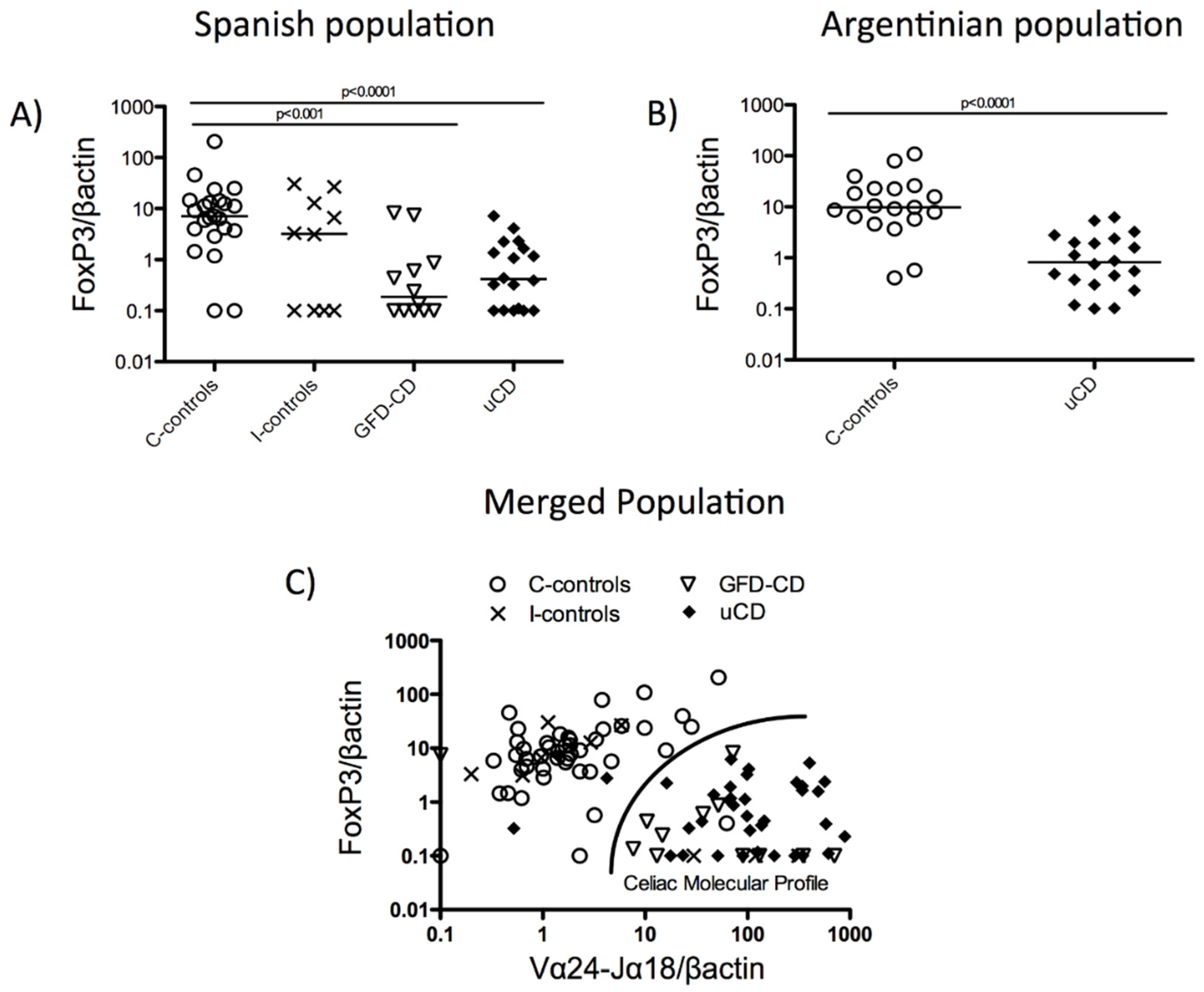
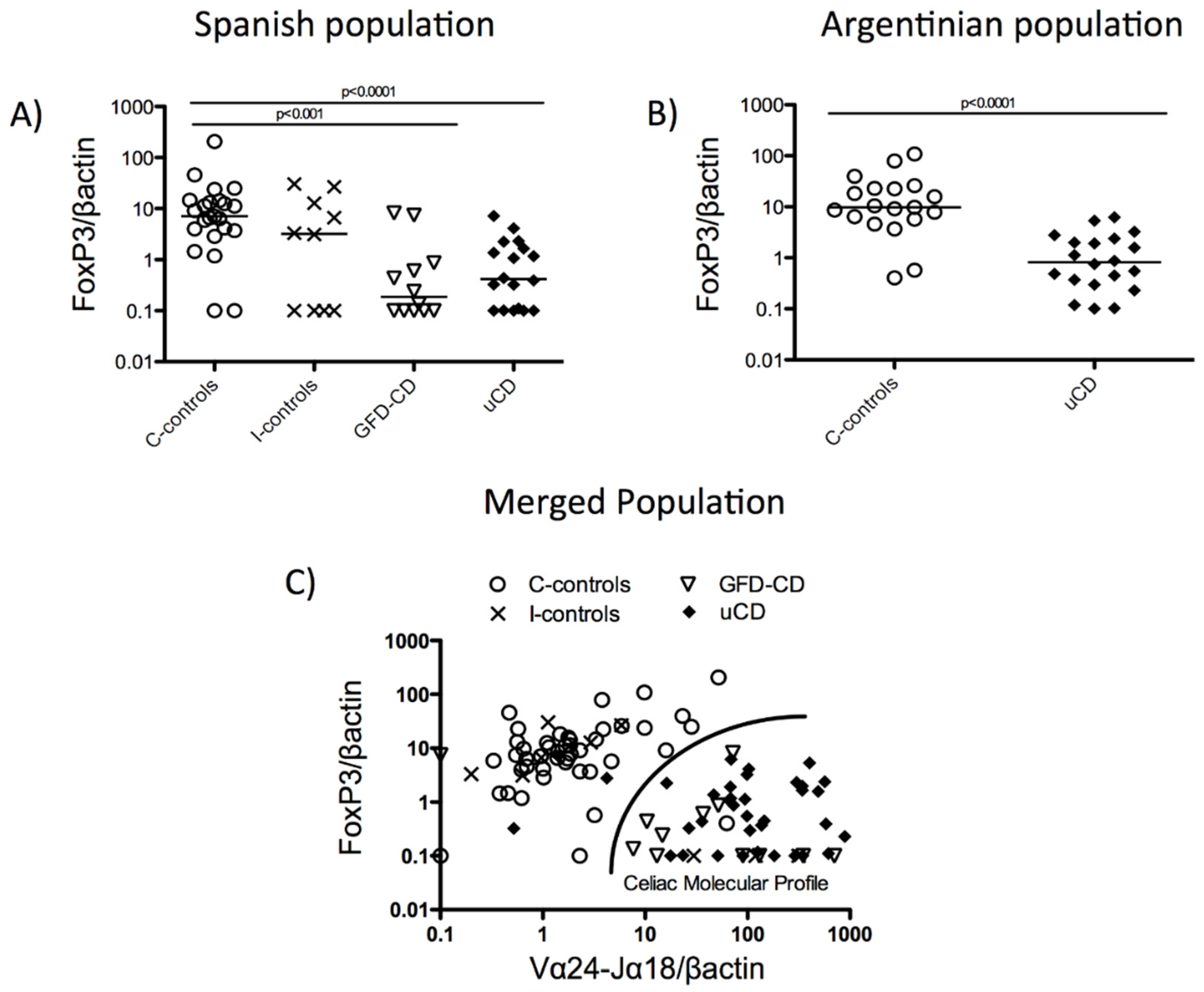
3.3. Duodenal Vα24-Jα18 and FoxP3 mRNA Levels Reveal a Celiac Disease Molecular Profile
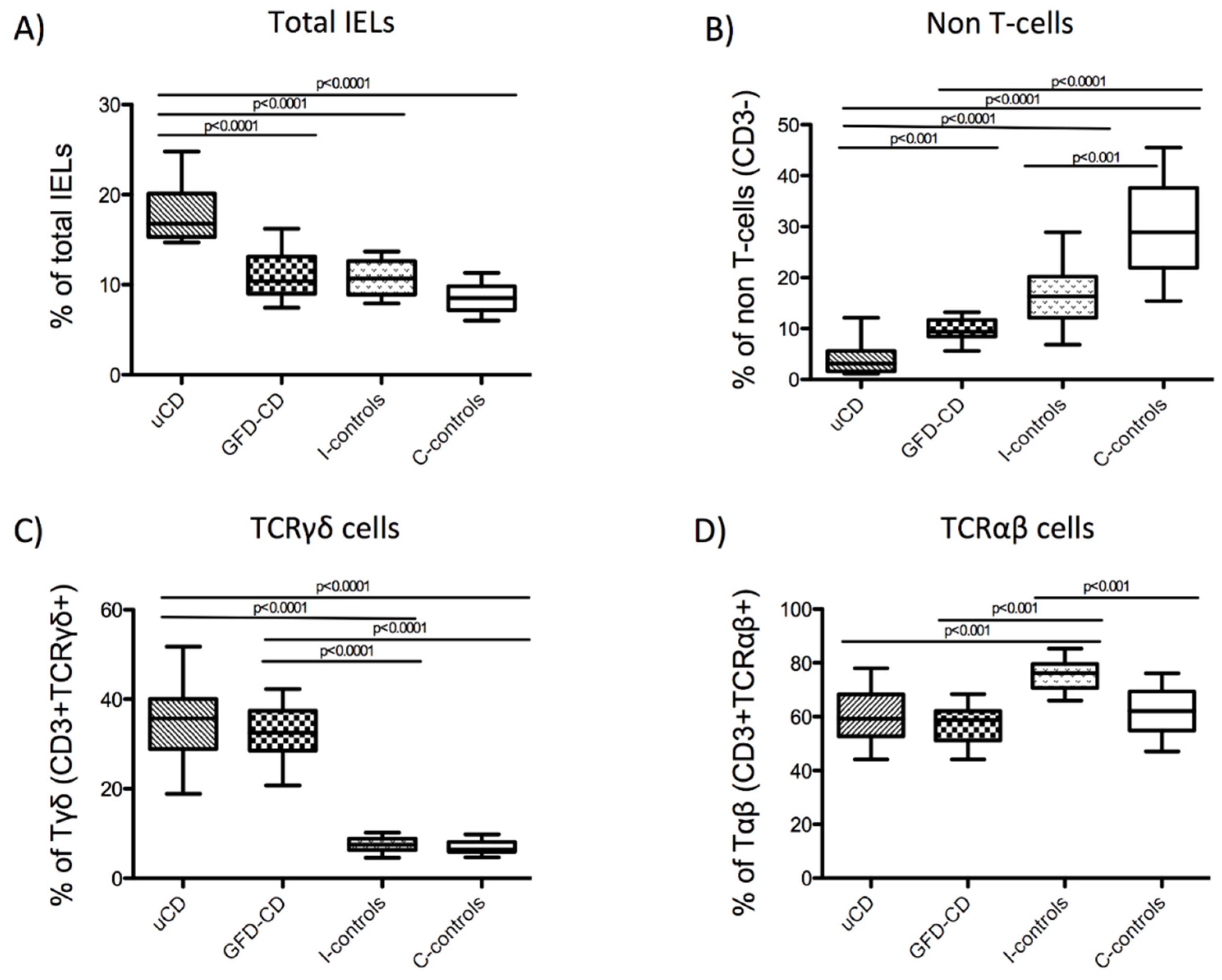
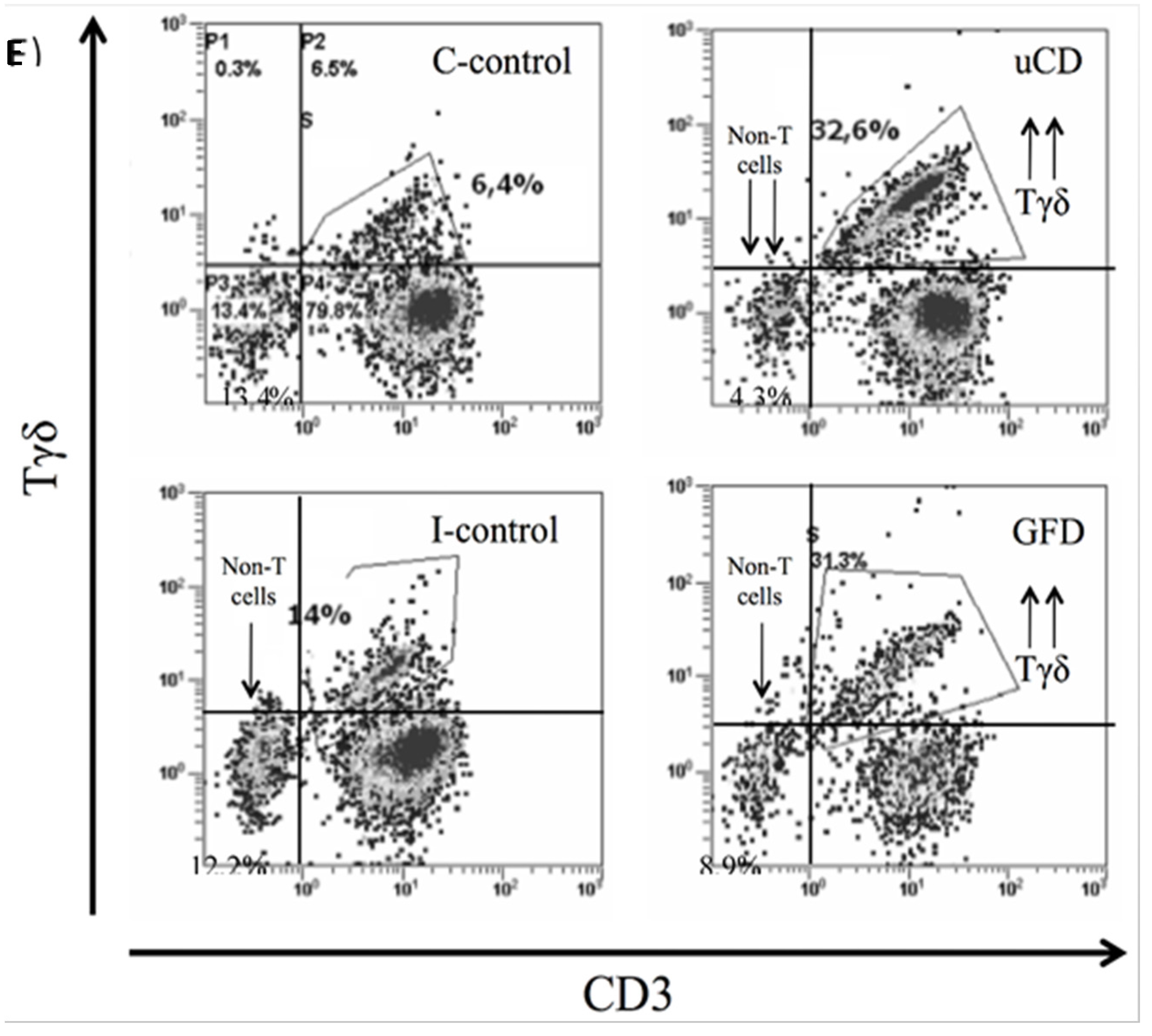
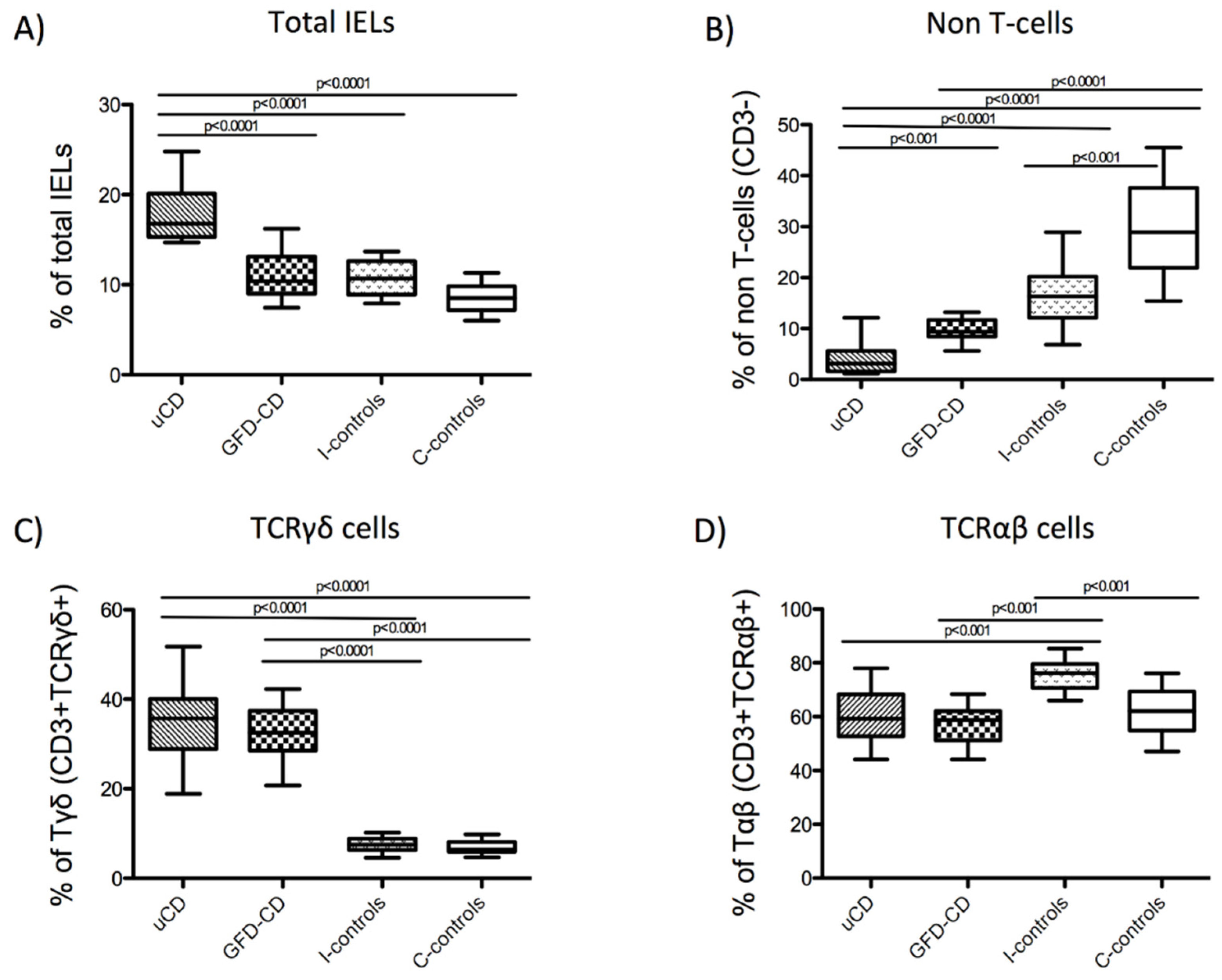
3.4. Intraepithelial Lymphocytes in the Duodenum from Celiac Disease Patients
| uCD Median (IQR) | GFD-CD Median (IQR) | I-controls Median (IQR) | C-controls Median (IQR) | |
|---|---|---|---|---|
| Total IELs | 16.80% (4.80) | 10.40% (4.11) | 10.70% (3.70) | 8.50% (2.60) |
| Non-T cells | 3.10% (4.00) | 9.34% (3.34) | 16.30% (8.12) | 28.90% (15.75) |
| TCRγδ+cells | 35.78% (11.13) | 32.50% (8.90) | 7.51% (2.60) | 6.44% (2.38) |
| TCRαβ+cells | 59.30% (15,6) | 58.80% (10,9) | 76.20% (8,9) | 62.10% (14,5) |
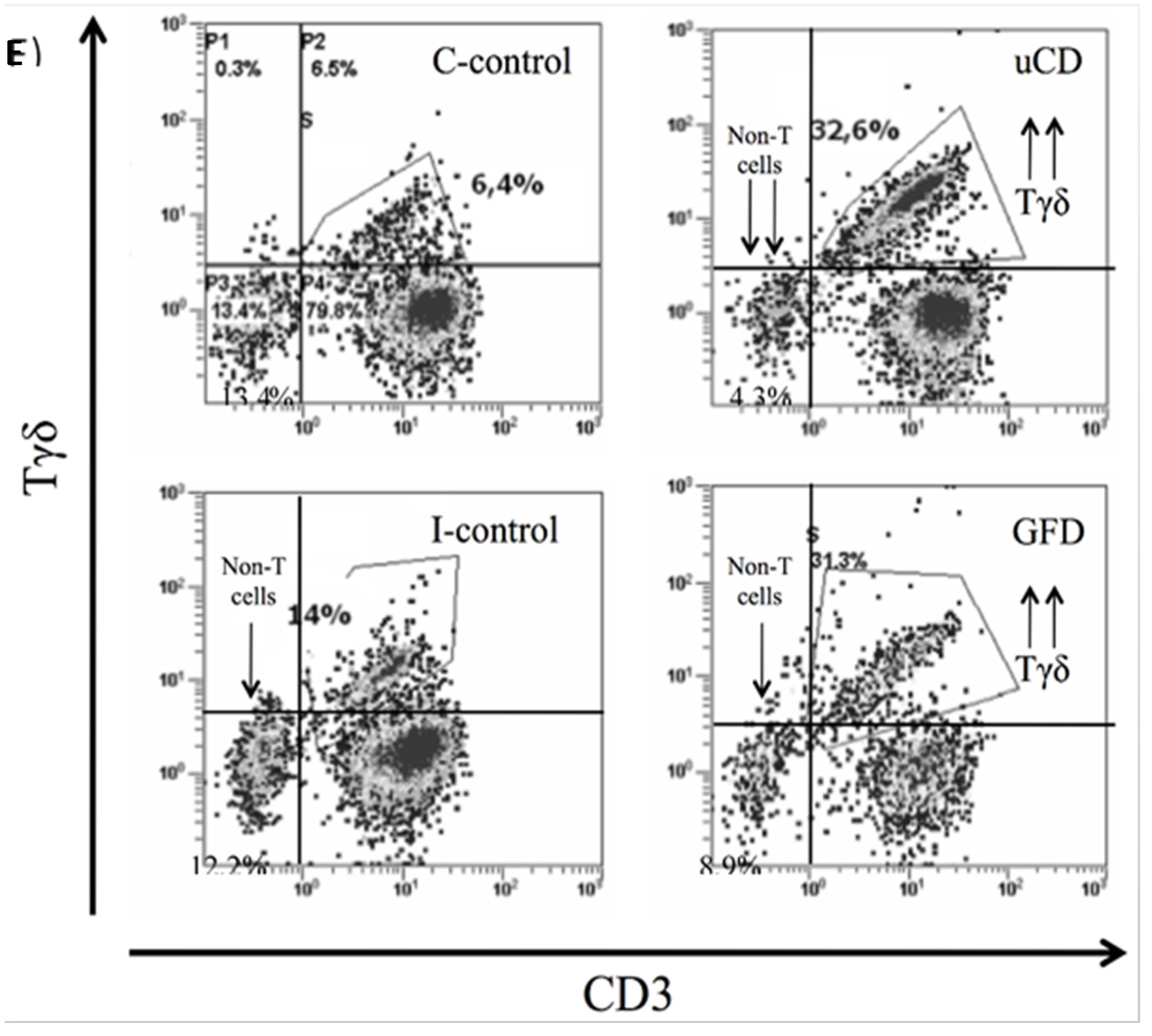
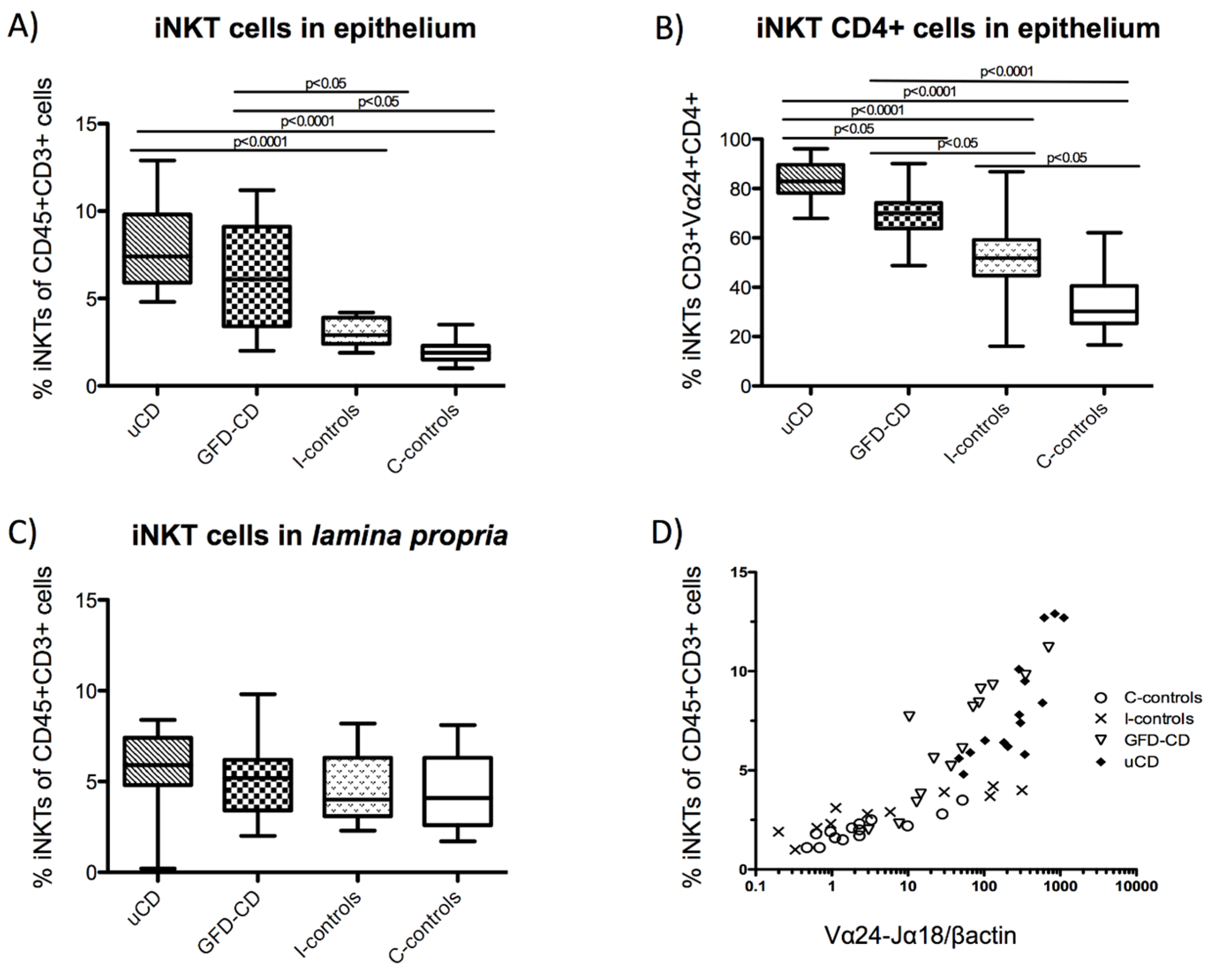
3.5. Increased Intraepithelial iNKT Cells in Celiac Disease Patients and Correlation with Vα24-Jα18 mRNA Expression

4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Comino, I.; Real, A.; de Lorenzo, L.; Cornell, H.; Lopez-Casado, M.A.; Barro, F.; Lorite, P.; Torres, M.I.; Cebolla, A.; Sousa, C. Diversity in oat potential immunogenicity: Basis for the selection of oat varieties with no toxicity in coeliac disease. Gut 2011, 60, 915–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, A.J.; Gomez, E.; Garrote, J.A.; Arranz, E. The pattern of cytokine expression determines the degree of mucosal damage. Gut 2007, 56, 441–443. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Sollid, L.M.; Barreiro, L.B.; Jabri, B. Integration of genetic and immunological insights into a model of celiac disease pathogenesis. Annu. Rev. Immunol. 2011, 29, 493–525. [Google Scholar] [CrossRef] [PubMed]
- Sollid, L.M.; Lundin, K.E. Diagnosis and treatment of celiac disease. Mucosal Immunol. 2009, 2, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Camarero, C.; Leon, F.; Sanchez, L.; Asensio, A.; Roy, G. Age-related variation of intraepithelial lymphocytes subsets in normal human duodenal mucosa. Dig. Dis. Sci. 2007, 52, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Cheroutre, H.; Lambolez, F.; Mucida, D. The light and dark sides of intestinal intraepithelial lymphocytes. Nat. Rev. Immunol. 2011, 11, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Discepolo, V.; Jabri, B. Intraepithelial lymphocytes in celiac disease immunopathology. Semin. Immunopathol. 2012, 34, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Meresse, B.; Malamut, G.; Cerf-Bensussan, N. Celiac disease: An immunological jigsaw. Immunity 2012, 36, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Camarero, C.; Eiras, P.; Asensio, A.; Leon, F.; Olivares, F.; Escobar, H.; Roy, G. Intraepithelial lymphocytes and coeliac disease: Permanent changes in CD3-/CD7+ and T cell receptor gammadelta subsets studied by flow cytometry. Acta Paediatr. 2000, 89, 285–290. [Google Scholar] [PubMed]
- Meresse, B.; Curran, S.A.; Ciszewski, C.; Orbelyan, G.; Setty, M.; Bhagat, G.; Lee, L.; Tretiakova, M.; Semrad, C.; Kistner, E.; et al. Reprogramming of CTLS into natural killer-like cells in celiac disease. J. Exp. Med. 2006, 203, 1343–1355. [Google Scholar] [CrossRef] [PubMed]
- Hue, S.; Mention, J.J.; Monteiro, R.C.; Zhang, S.; Cellier, C.; Schmitz, J.; Verkarre, V.; Fodil, N.; Bahram, S.; Cerf-Bensussan, N.; et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity 2004, 21, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, G.; Naiyer, A.J.; Shah, J.G.; Harper, J.; Jabri, B.; Wang, T.C.; Green, P.H.; Manavalan, J.S. Small intestinal CD8+TCRGAMMADELTA+NKG2a+ intraepithelial lymphocytes have attributes of regulatory cells in patients with celiac disease. J. Clin. Investig. 2008, 118, 281–293. [Google Scholar] [CrossRef] [PubMed]
- La Cava, A.; van Kaer, L.; Fu Dong, S. CD4+CD25+ TREGS and NKT cells: Regulators regulating regulators. Trends Immunol. 2006, 27, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Gianfrani, C.; Levings, M.K.; Sartirana, C.; Mazzarella, G.; Barba, G.; Zanzi, D.; Camarca, A.; Iaquinto, G.; Giardullo, N.; Auricchio, S.; et al. Gliadin-specific type 1 regulatory T cells from the intestinal mucosa of treated celiac patients inhibit pathogenic T cells. J. Immunol. 2006, 177, 4178–4186. [Google Scholar] [CrossRef] [PubMed]
- O’Garra, A.; Vieira, P. T(h)1 cells control themselves by producing interleukin-10. Nat. Rev. Immunol. 2007, 7, 425–428. [Google Scholar] [CrossRef] [PubMed]
- Middendorp, S.; Nieuwenhuis, E.E. NKT cells in mucosal immunity. Mucosal. Immunol. 2009, 2, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Brennan, P.J.; Tatituri, R.V.; Brigl, M.; Kim, E.Y.; Tuli, A.; Sanderson, J.P.; Gadola, S.D.; Hsu, F.F.; Besra, G.S.; Brenner, M.B. Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals. Nat. Immunol. 2011, 12, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, J.P.; Waldburger-Hauri, K.; Garzon, D.; Matulis, G.; Mansour, S.; Pumphrey, N.J.; Lissin, N.; Villiger, P.M.; Jakobsen, B.; Faraldo-Gomez, J.D.; et al. Natural variations at position 93 of the invariant Vα24-Jα18 α chain of human INKT-cell tcrs strongly impact on CD1D binding. Eur. J. Immunol. 2012, 42, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Veldt, B.J.; van der Vliet, H.J.; von Blomberg, B.M.; van Vlierberghe, H.; Gerken, G.; Nishi, N.; Hayashi, K.; Scheper, R.J.; de Knegt, R.J.; van den Eertwegh, A.J.; et al. Randomized placebo controlled phase I/II trial of α-galactosylceramide for the treatment of chronic hepatitis C. J. Hepatol. 2007, 47, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, D.; van Hoogstraten, I.M.; Verbeek, W.H.; Pena, A.S.; Mearin, M.L.; Arranz, E.; Garrote, J.A.; Scheper, R.J.; Schreurs, M.W.; Bontkes, H.J.; et al. Decreased circulating INKT cell numbers in refractory coeliac disease. Clin. Immunol. 2008, 126, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Zeissig, S.; Kaser, A.; Dougan, S.K.; Nieuwenhuis, E.E.; Blumberg, R.S. Role of NKT cells in the digestive system. III. Role of NKT cells in intestinal immunity. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G1101–G1105. [Google Scholar] [CrossRef] [PubMed]
- Molling, J.W.; Langius, J.A.; Langendijk, J.A.; Leemans, C.R.; Bontkes, H.J.; van der Vliet, H.J.; von Blomberg, B.M.; Scheper, R.J.; van den Eertwegh, A.J. Low levels of circulating invariant natural killer T cells predict poor clinical outcome in patients with head and neck squamous cell carcinoma. J. Clin. Oncol. 2007, 25, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Van der Vliet, H.J.; Molling, J.W.; von Blomberg, B.M.; Nishi, N.; Kolgen, W.; van den Eertwegh, A.J.; Pinedo, H.M.; Giaccone, G.; Scheper, R.J. The immunoregulatory role of CD1D-restricted natural killer T cells in disease. Clin. Immunol. 2004, 112, 8–23. [Google Scholar] [CrossRef] [PubMed]
- Dowds, C.M.; Blumberg, R.S.; Zeissig, S. Control of intestinal homeostasis through crosstalk between natural killer T cells and the intestinal microbiota. Clin. Immunol. 2015, 159, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Hwang, S.J.; Kim, B.K.; Jung, K.C.; Chung, D.H. NKT cells play critical roles in the induction of oral tolerance by inducing regulatory T cells producing IL-10 and transforming growth factor beta, and by clonally deleting antigen-specific t cells. Immunology 2006, 118, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; Lee, J.M.; Youn, H.J.; Lee, K.A.; Chung, Y.; Lee, A.Y.; Kweon, M.N.; Kim, H.Y.; Taniguchi, M.; Kang, C.Y. Functional maturation of lamina propria dendritic cells by activation of NKT cells mediates the abrogation of oral tolerance. Eur. J. Immunol. 2008, 38, 2727–2739. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, L.; Lynch, S.; Feighery, C.; Weir, D.; Kelleher, D.; O’Farrelly, C. Flow cytometric analysis of surface major histocompatibility complex class II expression on human epithelial cells prepared from small intestinal biopsies. J. Immunol. Methods 1993, 158, 207–214. [Google Scholar] [CrossRef]
- Aarsaether, N.; Nilsen, B.M. Nucleotide excision repair in human cells. Biochemistry and implications in diseases. Tidsskr. Nor. Laegeforen. 1995, 115, 2786–2789. [Google Scholar] [PubMed]
- Leon, F.; Sanchez, L.; Camarero, C.; Roy, G. Cytokine production by intestinal intraepithelial lymphocyte subsets in celiac disease. Dig. Dis. Sci. 2005, 50, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Garrote, J.A.; Gomez-Gonzalez, E.; Bernardo, D.; Arranz, E.; Chirdo, F. Celiac disease pathogenesis: The proinflammatory cytokine network. J. Pediatr. Gastroenterol. Nutr. 2008, 47 (Suppl. 1), S27–S32. [Google Scholar] [CrossRef] [PubMed]
- Leon, F.; Camarero, C.; Eiras, P.; Roy, G. Specificity of IEL profiling in the diagnosis of celiac disease. Am. J. Gastroenterol. 2004, 99, 958. [Google Scholar] [CrossRef] [PubMed]
- Arranz, E.; Bode, J.; Kingstone, K.; Ferguson, A. Intestinal antibody pattern of coeliac disease: Association with gamma/delta T cell receptor expression by intraepithelial lymphocytes, and other indices of potential coeliac disease. Gut 1994, 35, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Van Dieren, J.M.; van der Woude, C.J.; Kuipers, E.J.; Escher, J.C.; Samsom, J.N.; Blumberg, R.S.; Nieuwenhuis, E.E. Roles of CD1D-restricted NKT cells in the intestine. Inflamm. Bowel Dis. 2007, 13, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Cardell, S.L. The natural killer T lymphocyte: A player in the complex regulation of autoimmune diabetes in non-obese diabetic mice. Clin. Exp. Immunol. 2006, 143, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Grose, R.H.; Thompson, F.M.; Baxter, A.G.; Pellicci, D.G.; Cummins, A.G. Deficiency of invariant NKT cells in crohn's disease and ulcerative colitis. Dig. Dis. Sci. 2007, 52, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Grose, R.H.; Cummins, A.G.; Thompson, F.M. Deficiency of invariant natural killer T cells in coeliac disease. Gut 2007, 56, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Van der Vliet, H.J.; von Blomberg, B.M.; Nishi, N.; Reijm, M.; Voskuyl, A.E.; van Bodegraven, A.A.; Polman, C.H.; Rustemeyer, T.; Lips, P.; van den Eertwegh, A.J.; et al. Circulating Vα24+ Vβ11+ NKT cell numbers are decreased in a wide variety of diseases that are characterized by autoreactive tissue damage. Clin. Immunol. 2001, 100, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Calleja, S.; Vivas, S.; Santiuste, M.; Arias, L.; Hernando, M.; Nistal, E.; Casqueiro, J.; Ruiz de Morales, J.G. Dynamics of non-conventional intraepithelial lymphocytes-NK, NKT, and gammadelta T-in celiac disease: Relationship with age, diet, and histopathology. Dig. Dis. Sci. 2011, 56, 2042–2049. [Google Scholar] [CrossRef] [PubMed]
- Dunne, M.R.; Elliott, L.; Hussey, S.; Mahmud, N.; Kelly, J.; Doherty, D.G.; Feighery, C.F. Persistent changes in circulating and intestinal gammadelta T cell subsets, invariant natural killer T cells and mucosal-associated invariant T cells in children and adults with coeliac disease. PLoS ONE 2013, 8, e76008. [Google Scholar] [CrossRef] [PubMed]
- Leon, F.; Eiras, P.; Roy, G.; Camarero, C. Intestinal intraepithelial lymphocytes and anti-transglutaminase in a screening algorithm for coeliac disease. Gut 2002, 50, 740–741. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Banares, F.; Carrasco, A.; Garcia-Puig, R.; Rosinach, M.; Gonzalez, C.; Alsina, M.; Loras, C.; Salas, A.; Viver, J.M.; Esteve, M. Intestinal intraepithelial lymphocyte cytometric pattern is more accurate than subepithelial deposits of anti-tissue transglutaminase iga for the diagnosis of celiac disease in lymphocytic enteritis. PLoS ONE 2014, 9, e101249. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.M.; Murray, J.A. An update in the diagnosis of coeliac disease. Histopathology 2011, 59, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Grose, R.H.; Thompson, F.M.; Cummins, A.G. Deficiency of 6b11+ invariant NKT-cells in celiac disease. Dig. Dis. Sci. 2008, 53, 1846–1851. [Google Scholar] [CrossRef] [PubMed]
- Wingender, G.; Kronenberg, M. Role of NKT cells in the digestive system. Iv. The role of canonical natural killer T cells in mucosal immunity and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1–G8. [Google Scholar] [CrossRef] [PubMed]
- Eiras, P.; Roldan, E.; Camarero, C.; Olivares, F.; Bootello, A.; Roy, G. Flow cytometry description of a novel CD3-/CD7+ intraepithelial lymphocyte subset in human duodenal biopsies: Potential diagnostic value in coeliac disease. Cytometry 1998, 34, 95–102. [Google Scholar] [CrossRef]
- Eiras, P.; Leon, F.; Camarero, C.; Lombardia, M.; Roldan, E.; Bootello, A.; Roy, G. Intestinal intraepithelial lymphocytes contain a CD3- CD7+ subset expressing natural killer markers and a singular pattern of adhesion molecules. Scand. J. Immunol. 2000, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.O.; Porcelli, S.A. The diverse functions of CD1D-restricted NKT cells and their potential for immunotherapy. Immunol. Lett. 2005, 100, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Shinga, J.; Yamasaki, S.; Kawamura, M.; Dorrie, J.; Schaft, N.; Sato, Y.; Iyoda, T.; Fujii, S. Transfer of mRNA encoding invariant NKT cell receptors imparts glycolipid specific responses to T cells and gammadeltat cells. PLoS ONE 2015, 10, e0131477. [Google Scholar] [CrossRef] [PubMed]
- O’Keeffe, J.; Podbielska, M.; Hogan, E.L. Invariant natural killer T cells and their ligands: Focus on multiple sclerosis. Immunology 2015, 145, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Schrumpf, E.; Tan, C.; Karlsen, T.H.; Sponheim, J.; Bjorkstrom, N.K.; Sundnes, O.; Alfsnes, K.; Kaser, A.; Jefferson, D.M.; Ueno, Y.; et al. The biliary epithelium presents antigens to and activates natural killer T cells. Hepatology 2015, 62, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Sarra, M.; Cupi, M.L.; Monteleone, I.; Franze, E.; Ronchetti, G.; Di Sabatino, A.; Gentileschi, P.; Franceschilli, L.; Sileri, P.; Sica, G.; et al. IL-15 positively regulates IL-21 production in celiac disease mucosa. Mucosal Immunol. 2013, 6, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Van Bergen, J.; Mulder, C.J.; Mearin, M.L.; Koning, F. Local communication among mucosal immune cells in patients with celiac disease. Gastroenterology 2015, 148, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Gill, N.; Rosenthal, K.L.; Ashkar, A.A. Nk and nkt cell-independent contribution of interleukin-15 to innate protection against mucosal viral infection. J. Virol. 2005, 79, 4470–4478. [Google Scholar] [CrossRef] [PubMed]
- Pillai, V.; Ortega, S.B.; Wang, C.K.; Karandikar, N.J. Transient regulatory T-cells: A state attained by all activated human T-cells. Clin. Immunol. 2007, 123, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, D.; Al-Hassi, H.O.; Mann, E.R.; Tee, C.T.; Murugananthan, A.U.; Peake, S.T.; Hart, A.L.; Knight, S.C. T-cell proliferation and forkhead box p3 expression in human T cells are dependent on T-cell density: Physics of a confined space? Hum. Immunol. 2012, 73, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, D.L.; Spencer, J. Mouse and human intestinal immunity: Same ballpark, different players; different rules, same score. Mucosal Immunol. 2011, 4, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Ebert, L.M.; Tan, B.S.; Browning, J.; Svobodova, S.; Russell, S.E.; Kirkpatrick, N.; Gedye, C.; Moss, D.; Ng, S.P.; MacGregor, D.; et al. The regulatory T cell-associated transcription factor FoxP3 is expressed by tumor cells. Cancer Res. 2008, 68, 3001–3009. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.E.; van Bilsen, J.H.; Bakker, A.M.; Heemskerk, B.; Schilham, M.W.; Hartgers, F.C.; Elferink, B.G.; van der Zanden, L.; de Vries, R.R.; Huizinga, T.W.; et al. Expression of FoxP3 mRNA is not confined to CD4+CD25+ T regulatory cells in humans. Hum. Immunol. 2005, 66, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Chen, C.; Wang, L.; Chang, X.; Zheng, P.; Liu, Y. Cutting edge: Broad expression of the FoxP3 locus in epithelial cells: A caution against early interpretation of fatal inflammatory diseases following in vivo depletion of FoxP3-expressing cells. J. Immunol. 2008, 180, 5163–5166. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montalvillo, E.; Bernardo, D.; Martínez-Abad, B.; Allegretti, Y.; Fernández-Salazar, L.; Calvo, C.; Chirdo, F.G.; Garrote, J.A.; Arranz, E. Increased Intraepithelial Vα24 Invariant NKT Cells in the Celiac Duodenum. Nutrients 2015, 7, 8960-8976. https://doi.org/10.3390/nu7115444
Montalvillo E, Bernardo D, Martínez-Abad B, Allegretti Y, Fernández-Salazar L, Calvo C, Chirdo FG, Garrote JA, Arranz E. Increased Intraepithelial Vα24 Invariant NKT Cells in the Celiac Duodenum. Nutrients. 2015; 7(11):8960-8976. https://doi.org/10.3390/nu7115444
Chicago/Turabian StyleMontalvillo, Enrique, David Bernardo, Beatriz Martínez-Abad, Yessica Allegretti, Luis Fernández-Salazar, Carmen Calvo, Fernando G. Chirdo, José A. Garrote, and Eduardo Arranz. 2015. "Increased Intraepithelial Vα24 Invariant NKT Cells in the Celiac Duodenum" Nutrients 7, no. 11: 8960-8976. https://doi.org/10.3390/nu7115444