Dyslipidemia in Obesity: Mechanisms and Potential Targets

Department of Internal Medicine, Diabetes and Vascular Centre, Sint Franciscus Gasthuis, Rotterdam, P.O. Box 10900, 3004 BA, The Netherlands
*
Author to whom correspondence should be addressed.
Nutrients 2013, 5(4), 1218-1240; https://doi.org/10.3390/nu5041218
Submission received: 21 December 2012
/
Revised: 14 February 2013
/
Accepted: 27 March 2013
/
Published: 12 April 2013
(This article belongs to the Special Issue Dyslipidemia and Obesity)
{kind=link}
{kind=link}
Abstract
:Obesity has become a major worldwide health problem. In every single country in the world, the incidence of obesity is rising continuously and therefore, the associated morbidity, mortality and both medical and economical costs are expected to increase as well. The majority of these complications are related to co-morbid conditions that include coronary artery disease, hypertension, type 2 diabetes mellitus, respiratory disorders and dyslipidemia. Obesity increases cardiovascular risk through risk factors such as increased fasting plasma triglycerides, high LDL cholesterol, low HDL cholesterol, elevated blood glucose and insulin levels and high blood pressure. Novel lipid dependent, metabolic risk factors associated to obesity are the presence of the small dense LDL phenotype, postprandial hyperlipidemia with accumulation of atherogenic remnants and hepatic overproduction of apoB containing lipoproteins. All these lipid abnormalities are typical features of the metabolic syndrome and may be associated to a pro-inflammatory gradient which in part may originate in the adipose tissue itself and directly affect the endothelium. An important link between obesity, the metabolic syndrome and dyslipidemia, seems to be the development of insulin resistance in peripheral tissues leading to an enhanced hepatic flux of fatty acids from dietary sources, intravascular lipolysis and from adipose tissue resistant to the antilipolytic effects of insulin. The current review will focus on these aspects of lipid metabolism in obesity and potential interventions to treat the obesity related dyslipidemia.
1. Introduction
Obesity has turned into a worldwide epidemic. In the last decades the number of obese patients has increased considerably. It is especially alarming that in recent years the increase was most pronounced in children and that it occurs both in developed, but perhaps even more, in developing countries [1]. Visceral obesity leads to insulin resistance in part mediated by adipokines and free fatty acids (FFA). Adipokines such as resistin and retinol-binding protein 4 decrease insulin sensitivity, whereas leptin and adiponectin have the opposite effect. In addition, cytokines like TNF-α and IL-6, which originate from macrophages in adipose tissue, are involved [2]. Obesity, especially central obesity, is probably the main cause of the metabolic syndrome (MetS), which includes insulin resistance, type 2 diabetes mellitus, hypertension, the obstructive sleep apnea syndrome, non-alcoholic fatty liver disease (NAFLD) and dyslipidemia, all risk factors for cardiovascular disease [3,4]. Although doubts have arisen about the significance of the term metabolic syndrome in relation to cardiovascular complications, it has been suggested that identifying the condition will stimulate the physician to search also for the other risk factors clustering in the MetS [5].
The typical dyslipidemia of obesity consists of increased triglycerides (TG) and FFA, decreased HDL-C with HDL dysfunction and normal or slightly increased LDL-C with increased small dense LDL. The concentrations of plasma apolipoprotein (apo) B are also often increased, partly due to the hepatic overproduction of apo B containing lipoproteins [6,7]. The current review will focus on general lipid metabolism, the pathophysiological changes in lipid metabolism seen in obesity with the focus on postprandial lipemia and free fatty acid (FFA) dynamics and the potential pharmacological and non-pharmacological interventions.
2. Overview of Lipoprotein Metabolism
Numerous metabolic processes are involved in the uptake, transport and storage of lipids. After the ingestion of a meal containing fat, TG are lipolyzed in the intestinal lumen into FFA and 2-monoacylglycerols (MAG) and are taken up by the enterocytes via passive diffusion and specific transporters like CD36 [8]. Cholesterol is taken up by the enterocytes via the specific cholesterol transporter Niemann-Pick C1 Like 1 protein (NPC1L1) [9,10]. Once in the enterocyte, cholesterol is transformed into cholesterol-esters, whereas FFA and MAG are assembled into TG again. Finally, cholesterol-esters and TG are packed together with phospholipids and apolipoprotein (apo) B48 to form chylomicrons [8,11]. After assembly, the chylomicrons are secreted into the lymphatics and finally enter the circulation via the thoracic duct. The liver synthesizes TG-rich lipoproteins called very low density lipoproteins (VLDL), which increase postprandially when food derived TG and FFA reach the liver [11]. The assembly of VLDL is almost identical to the synthesis of chylomicrons, but apo B100 is the structural protein of VLDL (and its remnants, i.e., intermediate density lipoproteins (IDL) and low density lipoproteins (LDL)) [11]. The human liver lacks the editing complex necessary to change the apo B100 molecule into the smaller apoB48, by post-transcriptional modification of one base leading to a premature stop codon [12].
Chylomicrons and VLDL deliver FFA to the heart, skeletal muscle and adipose tissue for energy expenditure and storage. Adequate lipolysis of TG-rich lipoproteins is necessary for FFA to be released in the circulation. This process is regulated by several enzymes and proteins acting as co-factors. Lipoprotein lipase (LPL) is the primary enzyme for TG lipolysis in the circulation and is strongly expressed in tissues that require large amounts of FFA like the heart, skeletal muscle and adipose tissue [13]. LPL serves as the docking station for chylomicrons and VLDL for adherence to the endothelium via glycosyl-phosphatidylinositol-anchored high-density-binding protein 1 (GPIHBP1), which is present on the luminal side of the endothelium [14,15,16]. The amount of liberated FFA from chylomicrons and VLDL depends on the activity of LPL, which is stimulated by insulin [17,18]. In contrast, apo C-III is an inhibitor of LPL, but also of hepatic lipase. Plasma apo C-III concentrations correlate positively with plasma TG [19]. In addition, chylomicrons compete with endogenous VLDL for the action of LPL [20]. The liberated FFA are avidly taken up by adipocytes and re-synthesized into TG within the cytoplasm where the acylation-stimulating protein (ASP)/C3adesArg pathway plays an important role [21,22]. The scavenger receptor CD36 is the best characterized FFA transporter and is abundant in muscle, adipose tissue and the capillary endothelium [23]. Insulin and muscle contractions increase the CD36 expression thereby facilitating FFA uptake [13].
The postprandial rise in insulin is one of the most important regulatory mechanisms for fuel storage. The postprandial increase of insulin results in the effective inhibition of hormone sensitive lipase, which is the key enzyme for hydrolysis of intracellular lipids. Despite the uptake of FFA by adipocytes and myocytes, a proportion of FFA remains in the plasma compartment (“spill over”) where the FFA are bound by albumin and transported to the liver [24]. When delivery of FFA for energy expenditure is insufficient like in the fasting state, FFA can be mobilized by adipose tissue for oxidation in energy demanding tissues like cardio myocytes. Insulin is also an important regulator of FFA mobilization from adipose tissue [17]. Therefore, insulin resistance has a major impact on the metabolism of TG-rich lipoproteins and FFA.
Eventually, chylomicrons and VLDL shrink in diameter during the process of lipolysis to form chylomicron remnants and dense LDL, respectively. Chylomicron remnants are taken up by the liver via multiple pathways including apo E, hepatic lipase, the LDL receptor, the LDL receptor-related protein and heparan sulphate proteoglycans [25,26,27,28,29,30]. In contrast, LDL is primarily taken up by the liver via the LDL receptor [31,32]. The LDL receptor is recycled and re-shuttled back to the cell surface. In the last decade, many studies have extended our knowledge concerning this recycling process of the LDL receptor, which is regulated by the proprotein convertase subtilisin/kexin type 9 (PCSK9) [32,33]. The LDL receptor undergoes lysosomal degradation during the shuttling process when PCSK9 is bound to the LDL receptor, but is recycled back to the surface of the hepatocytes in the absence of PCSK9 [33]. Neutralization of PCSK9 increases the total LDL binding capacity of the hepatocytes leading to reduced LDL-C concentrations [33].
Besides the above described TG and LDL metabolism, the intestine and liver also play an important role in the reverse cholesterol transport by the synthesis of HDL particles. HDL promotes the uptake of cholesterol from peripheral tissues, including the arterial wall, and returns cholesterol to the liver. Enterocytes and hepatocytes synthesize apo A-I which is the structural protein of HDL. Nascent HDL particles acquire free cholesterol from peripheral tissues. Subsequently, the cholesterol within HDL becomes esterified into cholesterol-esters by HDL associated lecithin-cholesterol acyltransferase (LCAT) [23]. Within the circulation, the HDL particles also become enriched with cholesterol-esters by the action of cholesterylester-transfer-protein (CETP) and phospholipid transfer protein (PLTP). In this process HDL acquires TG from TG-rich lipoproteins in exchange for cholesterol-esters as a direct consequence of the CETP action [11]. In the liver, hepatic lipase hydrolyses HDL-associated TG and also phospholipids inducing the formation of smaller HDL particles which can contribute again to the reverse cholesterol transport. Therefore, lipid metabolism is highly dynamic and depends on numerous factors including the postprandial state, TG-rich lipoprotein concentrations, HDL levels and function, energy expenditure, insulin levels and sensitivity and adipose tissue function.
3. Obesity Induced Changes in Lipoprotein Metabolism and Atherogenic Effects
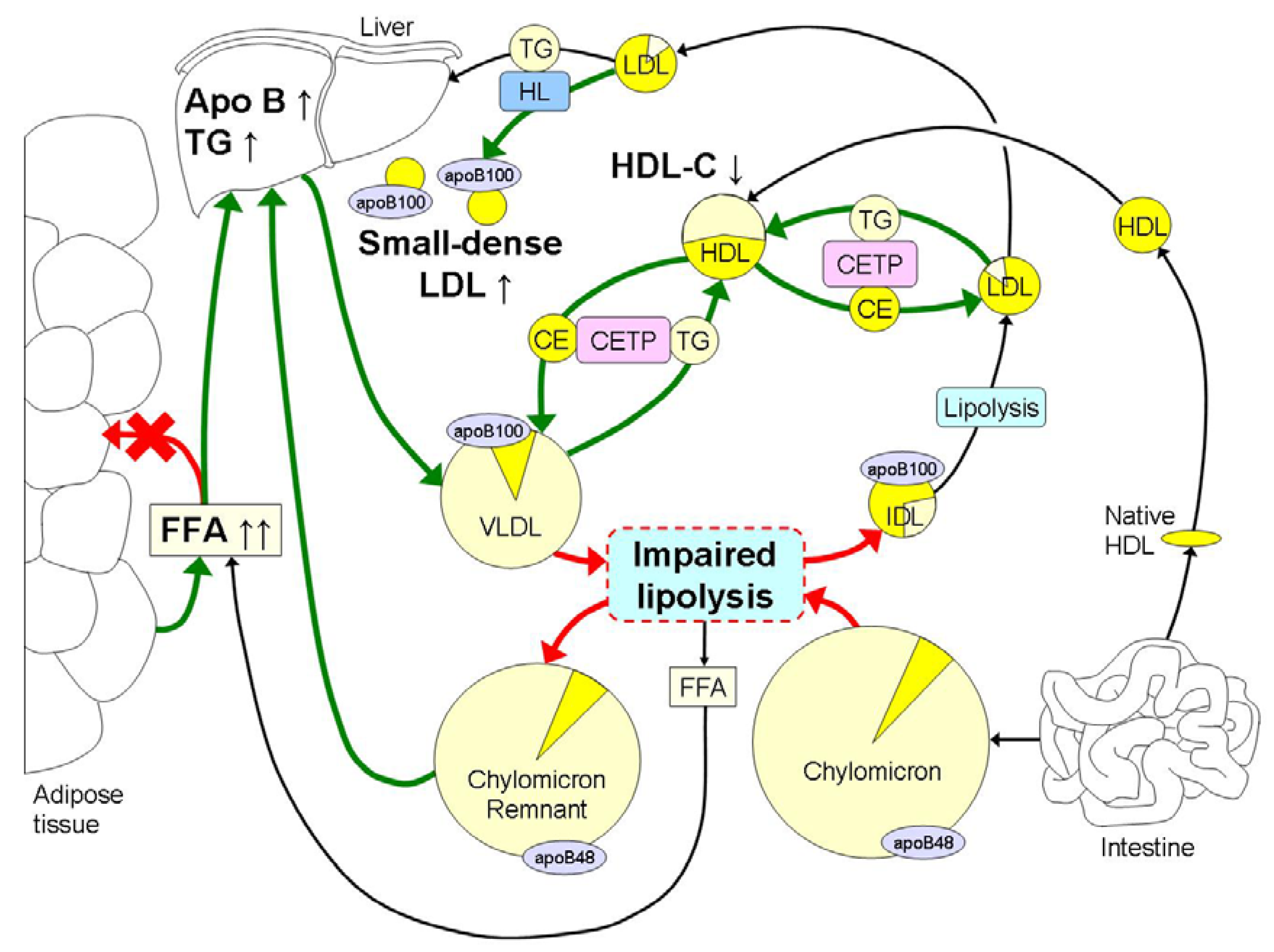
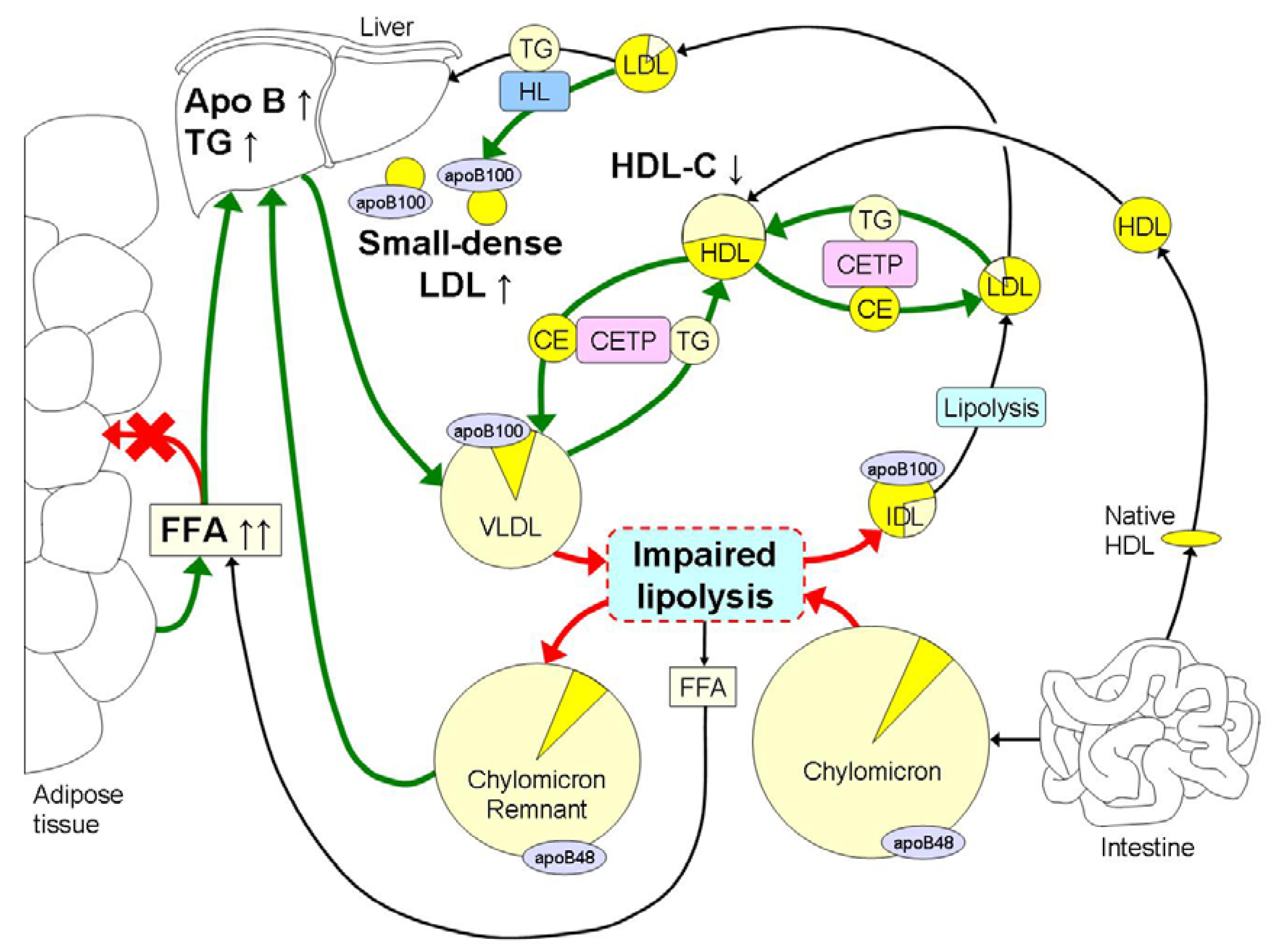
The hallmark of dyslipidemia in obesity is elevated fasting and postprandial TG in combination with the preponderance of small dense LDL and low HDL-C (Figure 1). Hypertriglyceridemia may be the major cause of the other lipid abnormalities since it will lead to delayed clearance of the TG-rich lipoproteins [34,35,36,37,38,39,40,41,42,43,44,45,46,47,48] and formation of small dense LDL [48,49].
Lipolysis of TG-rich lipoproteins is impaired in obesity by reduced mRNA expression levels of LPL in adipose tissue [50], reductions in LPL activity in skeletal muscle and competition for lipolysis between VLDL and chylomicrons [11]. Increased postprandial lipemia leads to elevated levels of FFA, resulting in detachment of LPL from its endothelial surface [51,52]. LPL may remain attached to VLDL and IDL contributing to further TG depletion. The exchange of TG from these remnants for cholesterol-esters from HDL by CETP with the concerted action of hepatic lipase, ultimately leads to the formation of small dense LDL [48,49]. In the presence of hypertriglyceridemia, the cholesterol-ester content of LDL decreases, whereas the TG content of LDL increases by the activity of CETP. However, the increased TG content within the LDL is hydrolyzed by hepatic lipase, which leads to the formation of small, dense LDL particles. The development of small dense LDL in obesity is mainly due to increased TG concentrations and does not depend on total body fat mass [53]. Small dense LDL are relatively slowly metabolized with a five day residence time, which enhances its atherogenicity [54].
Chylomicron remnants and LDL may migrate into the sub-endothelium and become trapped in the sub-endothelial space where they can be taken up by monocytes/macrophages [55,56,57]. Small dense LDL have an increased affinity for arterial proteoglycans resulting in enhanced subendothelial lipoprotein retention [58]. However, subendothelial remnants of chylomicrons and VLDL do not need to become modified to allow uptake by scavenger receptors of macrophages in contrast to native LDL [59]. It has been described that small dense LDL are more susceptible for oxidation, in part due to less free cholesterol and anti-oxidative content [60]. It should be noted that the lipoprotein size is a limiting factor for migration through the endothelium and that LDL particles migrate more easily than chylomicron remnants, but the number of migrated particles does not necessarily translate into more cholesterol deposition since chylomicron remnants contain approximately 40 times more cholesterol per particle than LDL [57]. Alternatively, LPL-enriched remnants of chylomicrons and VLDL may be transported to the tissues where interaction with proteoglycans and lipoprotein receptors lead to particle removal. This process takes place at the liver and acts as an anti-atherogenic mechanism, but it may also take place in other tissues where cholesterol can not be removed efficiently leading to cholesterol accumulation and therefore the initiation of the atherosclerotic plaque [56,57,61,62].
Studies using stable isotopes have shown a decreased catabolism of chylomicron remnants in obese subjects with the waist/hip ratio as best predictor for the fractional catabolic rate [63]. Taskinen and co-workers showed that the defective clearance of remnant lipoproteins can be explained by elevated concentrations of apo C-III in the situation of obesity [64]. Elevated levels of apo C-III in obesity can be explained by glucose-stimulated transcription of apo C-III and it has been described that plasma apo C-III levels correlate with fasting glucose and glucose excursion after an oral glucose test in obese humans [65]. Finally, the LDL receptor expression is reduced in obesity [66].
Figure 1.
The hallmark of dyslipidemia in obesity is hypertriglyceridemia in part due to increased free fatty acid (FFA) fluxes to the liver, which leads to hepatic accumulation of triglycerides (TG). This leads to an increased hepatic synthesis of large very low density lipoproteins (VLDL) 1, which hampers the lipolysis of chylomicrons due to competition mainly at the level of lipoprotein lipase (LPL) with increased remnant TG being transported to the liver. Lipolysis is further impaired in obesity by reduced mRNA expression levels of LPL in adipose tissue and reduced LPL activity in skeletal muscle. Hypertriglyceridemia further induces an increased exchange of cholesterolesters (CE) and TG between VLDL and HDL and low density lipoproteins (LDL) by cholesterylester-transfer-protein (CETP). This leads to decreased HDL-C concentrations and a reduction in TG content in LDL. In addition, hepatic lipase (HL) removes TG and phospholipids from LDL for the final formation of TG-depleted small dense LDL. The intense yellow color represents cholesterol, whereas the light yellow color represents the TG content within the different lipoproteins. Obesity induced increases in metabolic processes are marked with green arrows, whereas reductions are marked with red arrows.
Figure 1.
The hallmark of dyslipidemia in obesity is hypertriglyceridemia in part due to increased free fatty acid (FFA) fluxes to the liver, which leads to hepatic accumulation of triglycerides (TG). This leads to an increased hepatic synthesis of large very low density lipoproteins (VLDL) 1, which hampers the lipolysis of chylomicrons due to competition mainly at the level of lipoprotein lipase (LPL) with increased remnant TG being transported to the liver. Lipolysis is further impaired in obesity by reduced mRNA expression levels of LPL in adipose tissue and reduced LPL activity in skeletal muscle. Hypertriglyceridemia further induces an increased exchange of cholesterolesters (CE) and TG between VLDL and HDL and low density lipoproteins (LDL) by cholesterylester-transfer-protein (CETP). This leads to decreased HDL-C concentrations and a reduction in TG content in LDL. In addition, hepatic lipase (HL) removes TG and phospholipids from LDL for the final formation of TG-depleted small dense LDL. The intense yellow color represents cholesterol, whereas the light yellow color represents the TG content within the different lipoproteins. Obesity induced increases in metabolic processes are marked with green arrows, whereas reductions are marked with red arrows.
Remnants of chylomicrons and VLDL are involved in the development of atherosclerosis [67]. Several investigators have demonstrated an association between TG-rich lipoproteins and remnant cholesterol levels with the presence of coronary [34,35,36,38,39,40,41,42,68], cerebral [37], and peripheral atherosclerosis [69]. In addition to a direct detrimental effect by chylomicron remnants on vessels [59], impaired endothelial function after an oral fat load [70] and after infusion of artificial TG-rich lipoproteins have been described [71]. This phenomenon may take place by elevated levels of FFA [72], which are generated by the action of LPL mediated lipolysis. Other mechanisms of remnant-mediated atherogenesis which may play a role in obesity comprise the postprandial activation of leukocytes, generation of oxidative stress and production of cytokines [55,73,74].
Postprandial hyperlipidemia with accumulation of atherogenic remnants is especially linked to visceral obesity [75,76]. Postprandial lipid metabolism has been investigated in metabolic ward studies using non-physiological high amounts of fat [77]. A more physiological method to study postprandial lipemia has been developed in our laboratory, namely the measurement of daytime capillary TG profiles using repeated capillary self-measurements in an out of hospital situation [78,79]. It has been shown that diurnal triglyceridemia in obese subjects correlates better to waist circumference than to body mass index [78,80], which is in agreement with the hypothesis that the distribution of adipose tissue modulates postprandial lipemia [81]. All these mechanisms have been related to the higher incidence of cardiovascular disease seen in obesity [82].
HDL metabolism is also strongly affected by obesity because of the increased number of remnants of chylomicrons and VLDL together with impaired lipolysis. The increased number of TG-rich lipoproteins results in increased CETP activity, which exchanges cholesterolesters from HDL for TG from VLDL and LDL [60]. Moreover, lipolysis of these TG-rich HDL occurs by hepatic lipase resulting in small HDL with a reduced affinity for apo A-I, which leads to dissociation of apo A-I from HDL. This will ultimately lead to lower levels of HDL-C and a reduction in circulating HDL particles with impairment of reversed cholesterol transport [83].
4. Interplay between FFA Metabolism and Inflammation in Obesity: Crossroad between Innate Immunity and Lipid Metabolism
There are only two sources where plasma FFA may be derived from: firstly, lipolysis of TG-rich lipoproteins within the circulation and secondly, intracellular lipolysis in adipose tissue. An excellent review from the Oxford group described the relationship between plasma concentrations of FFA and insulin resistance as seen in obesity [17]. Other reviews have been published recently by several other groups as well [3,84]. It is widely recognized that plasma FFA are elevated in obese people as a consequence of an increased fatty acid release from adipose tissue and a reduction in plasma FFA clearance [85,86,87]. The increase in FFA and obesity-induced inflammation play a crucial role in the development of insulin resistance [88].
Various fatty acids are cytotoxic and their cytotoxicity depend on the type and has been extensively reviewed elsewhere [89,90]. Saturated fatty acids (SFA), arachidonic acid and linoleic acid (both polyunsaturated fatty acids (PUFA)) can mediate a diet-induced inflammation, although the literature concerning PUFA and inflammation is not consistent [89,90]. SFA, arachidonic acid and linoleic acid can stimulate the synthesis of pro-inflammatory cytokines like IL-1, IL-6 and TNF-α, whereas eicosepantenoic acid, a fish oil, has anti-inflammatory properties [89,90,91]. Since various fatty acids are cytotoxic, an “escape mechanism” should be present in order to remove FFA from the micro-environment where they are formed. In this process both, insulin and the acylation-stimulating protein (ASP)/C3adesArg-pathway play an important role in peripheral fatty acid trapping.
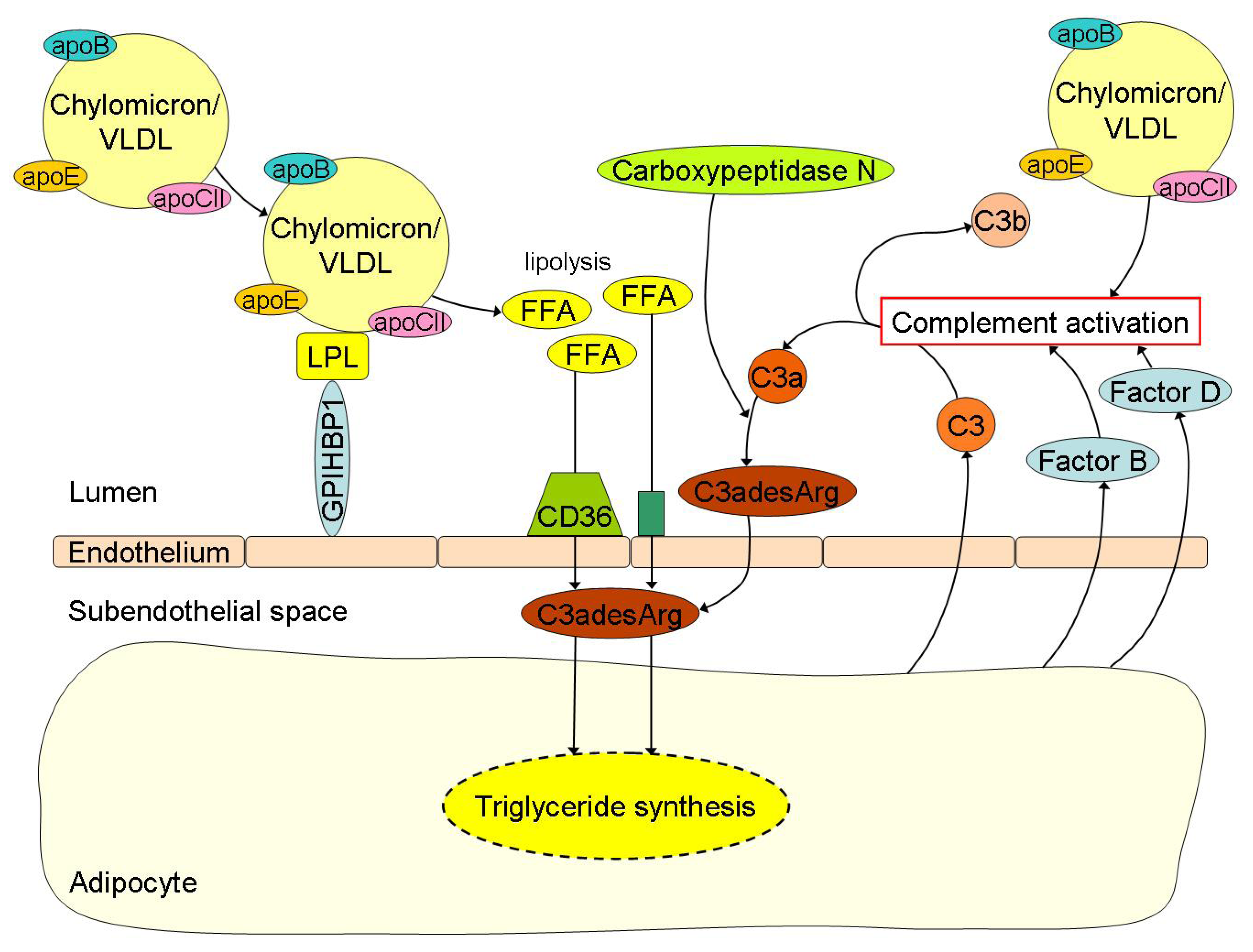
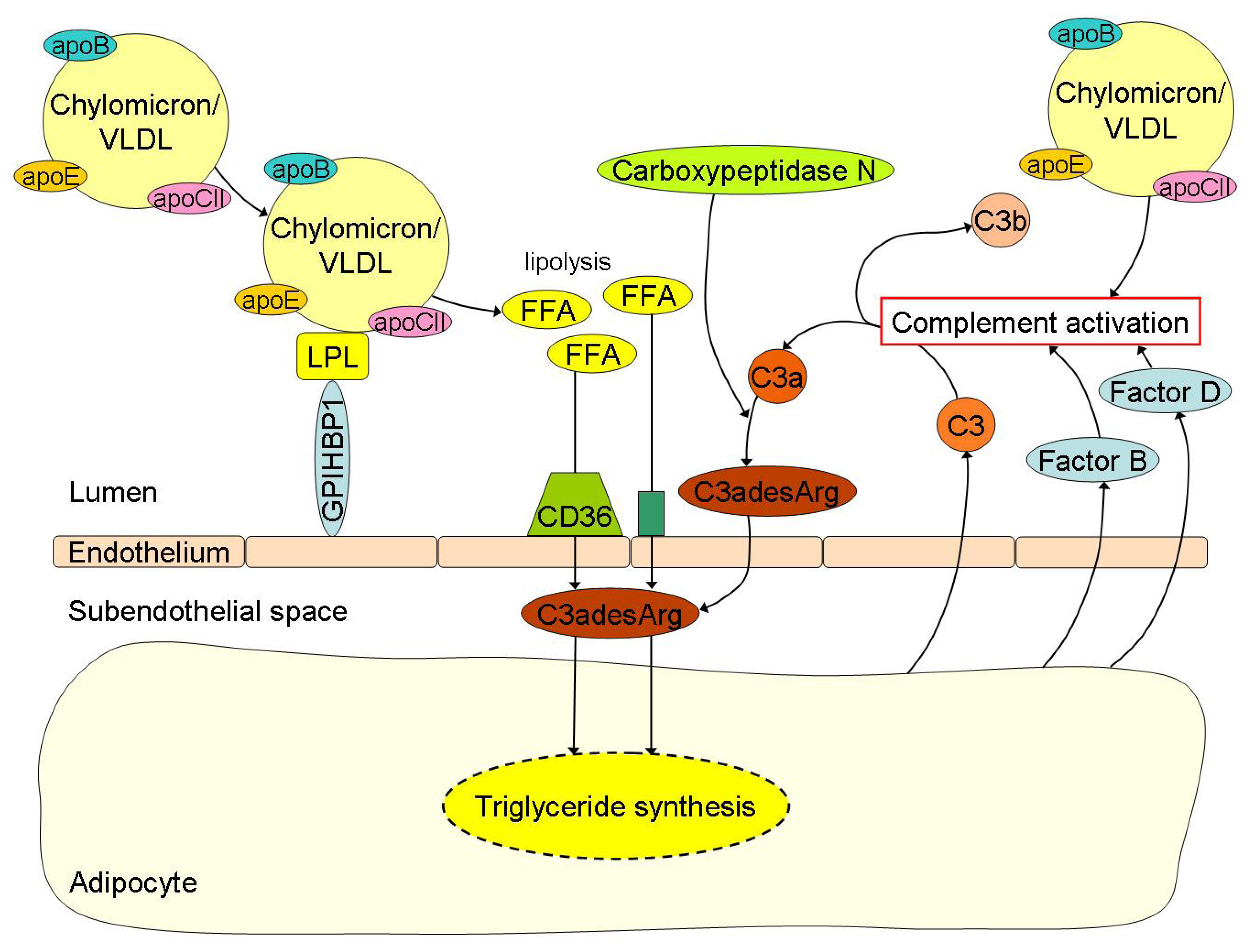
ASP in relation to peripheral fatty acid trapping was first described by Sniderman and collaborators [92]. In reaction to fatty acid delivery, adipocytes and fibroblasts secrete complement component 3 (C3) [93,94,95,96]. By the action of factor B and factor D (also secreted by adipocytes and fibroblasts) a small active fragment is split (C3a) from C3, which is readily converted into C3adesArg (also known as ASP) by carboxypeptidase N (Figure 2) [97]. C3adesArg, while not immunological active, has an important physiological role in the storage of fatty acids in adipocytes and other peripheral cells. Besides insulin, C3adesArg induces trans-membrane transport of fatty acids and their intracellular esterification into TG [21,22]. Recently, it has been described that ASP mRNA expression in visceral adipose tissue is reduced by approximately 40% in obese and morbidly obese subjects with or without insulin resistance when compared to lean controls [50]. In addition, C3adesArg mediates insulin-independent trans-membrane glucose transport [98]. It should be mentioned that these ASP-mediated processes only take place at peripheral cells and not in the liver. Fatty acid and glucose uptake by hepatocytes is ASP-independent.
In line with this ASP/C3adesArg concept are several studies, which investigated the role of the complement system in lipoprotein metabolism. Our group and others were able to demonstrate that the complement component 3 (C3) is one of the major determinants of the MetS [99,100,101] and postprandial lipemia in insulin resistant subjects, but also in insulin sensitive subjects [87,102,103]. C3 has also been genetically linked to the MetS in a recent meta-analysis of multiple genome wide association studies [104]. It was also demonstrated that a different component of the complement system, mannose binding lectin, may be involved in normal handling of postprandial lipoproteins [105]. Therefore, there is sufficient evidence supporting the notion that the complement system is an important regulator of postprandial fatty acid and TG metabolism and substantiates the concept that ASP/C3adesArg resistance plays a role in adequate peripheral fatty acid handling [102,106,107].
One of the difficulties in the evaluation of fatty acid metabolism is the determination of exact kinetics and trafficking of fatty acids between different tissues. Recent work from the Oxford group using arterio-venous blood sampling in adipose tissue with labeled palmitate, elegantly demonstrated impaired fatty acid trapping in vivo in obese men [18]. In addition, treatment of insulin resistance with metformin has been shown to reduce plasma FFA concentrations by lowering fasting FFA levels but without any effect on catecholamine mediated lipolysis of adipocytes [108]. Moreover, obese men also showed decreased uptake of dietary fat by adipose tissue, which results in a higher delivery of chylomicron remnants to the liver with consequently enhanced VLDL-TG being delivered to peripheral adipocytes [18]. The authors referred to this situation as “a seemingly unnecessary loop of fatty acid trafficking to the liver” and associated that to increased liver fat content.
Figure 2.
Free fatty acid (FFA) uptake and its related triglyceride (TG) synthesis in adipocytes are highly depended of C3adesArg or acylation-stimulation protein (ASP). Chylomicrons and VLDL undergo lipolysis by lipoprotein lipase (LPL) with subsequent release of FFA into the circulation. The FFA are then transported into the subendothelial space by the scavenger receptor CD36 and other transporters where C3adesArg plays an important role in the subsequent TG synthesis for storage of lipids in the adipocytes. C3adesArg is the most potent molecule known, which induces transmembrane transport of FFA and its intracellular esterification into TG within adipocytes. C3adesArg is metabolized from complement component (C) 3a by carboxypeptidase N and C3a is again the splice product from C3, which is formed in case of complement activation. Postprandial lipemia is directly linked to complement activation. For example, adipocytes secrete C3 when incubated with TG-rich lipoproteins like chylomicrons or very low density lipoproteins (VLDL), but also Factor B and Factor D, thereby causing activation of the complement cascade.
Figure 2.
Free fatty acid (FFA) uptake and its related triglyceride (TG) synthesis in adipocytes are highly depended of C3adesArg or acylation-stimulation protein (ASP). Chylomicrons and VLDL undergo lipolysis by lipoprotein lipase (LPL) with subsequent release of FFA into the circulation. The FFA are then transported into the subendothelial space by the scavenger receptor CD36 and other transporters where C3adesArg plays an important role in the subsequent TG synthesis for storage of lipids in the adipocytes. C3adesArg is the most potent molecule known, which induces transmembrane transport of FFA and its intracellular esterification into TG within adipocytes. C3adesArg is metabolized from complement component (C) 3a by carboxypeptidase N and C3a is again the splice product from C3, which is formed in case of complement activation. Postprandial lipemia is directly linked to complement activation. For example, adipocytes secrete C3 when incubated with TG-rich lipoproteins like chylomicrons or very low density lipoproteins (VLDL), but also Factor B and Factor D, thereby causing activation of the complement cascade.
5. Lifestyle Interventions for Dyslipidemia in Obesity
Treatment of obesity-associated dyslipidemia should be focused on lifestyle changes including weight loss, physical exercise and a healthy diet. Lifestyle changes synergistically improve insulin resistance and dyslipidemia [59]. The amount of ingested fat and total calories are the most important dietary factors to induce obesity and its related postprandial lipemia [109]. This has already been demonstrated in early childhood [110]. Weight loss has been demonstrated to markedly reduce fasting and non-fasting TG concentrations, which can be attributed to an increase in LPL activity with a concomitant reduction in apo C-III levels [111], a decrease in CETP activity [112,113] and an increased catabolism of TG-rich lipoproteins [114]. Besides reductions in fasting and non-fasting TG concentrations, a small reduction in LDL-C can be expected upon weight loss, which may be attributed to increased LDL receptor activity. A weight loss of 4–10 kg in obese subjects resulted in a 12% reduction in LDL-C and a 27% increase in LDL receptor mRNA levels [111,115].
The type of dietary fat also affects postprandial lipemia [109]. A study in rats showed that a diet high in saturated fats reduced LPL protein levels and LPL activity in skeletal muscle, whereas LPL activity was increased in adipose tissue favoring shunting of lipids from skeletal muscle to adipose tissue [116]. Moderate weight loss (approximately 10%) in obese, but otherwise healthy men, which was induced by a diet low on carbohydrates and SFA and high on mono-unsaturated fatty acids (MUFA) resulted in a 27%–46% reduction in postprandial TG levels [117]. Long term intervention with MUFA resulted in a reduction in postprandial inflammation when compared to a diet rich in SFA in patients with the MetS [118].
Recent genome wide association studies have found more than 95 loci associated with lipid levels, but together they explain less than 10% of the variation in lipids. Interactions between genes, obesity and lipid levels but also with the type of dietary fat consumed have recently been described [119,120,121,122]. Homozygosity for the C allele of the APOA2 −265T > C polymorphism was associated with an increased obesity prevalence compared to the TT + TC genotype in those subjects with high SFA consumption (OR 1.84 95% CI 1.38–2.47) [120]. In a Spanish population with a relatively high MUFA intake, carriers of the minor C allele of the APOA5 −1131T > C polymorphism, which is associated with increased plasma TG, appear to be more resistant to weight gain by fat consumption and showed an inverse relationship between fat intake and plasma TG [122]. However, high PUFA consumption was associated with increased plasma TG and decreased LDL particle size in carriers of the C allele in a U.S. population [121]. These results suggest the potential usefulness of a nutrigenomic approach for dietary interventions to prevent or treat obesity and its related dyslipidemia.
Physical exercise has been shown to increase LPL and hepatic lipase activity, which stimulates TG lipolysis [123,124]. The mechanism of exercise-induced LPL activity remains unclear, but it was hypothesized that exercise stimulates especially muscular LPL activity, although this could not be confirmed in a recent study [125]. A 12-week walking program supplemented with fish oil (1000 mg eicosepantenoic acid and 700 mg docosahexaenoic acid daily) in subjects with the MetS resulted in lower fasting TG and decreased the postprandial response of TG and apoB48 [126]. Exercise training for 16 weeks in obese subjects with NAFLD resulted in a small reduction in intra-hepatic TG content, although no changes in VLDL-TG or apoB100 secretion were observed [127]. Exercise induced reductions in intra-hepatic TG content have also been reported even in the absence of weight loss [128]. Moreover, intra-hepatic TG content was reduced in overweight men after a low fat diet for three weeks, whereas a high fat diet increased intra-hepatic TG [129]. The plasma TG lowering effect of exercise and weight loss is the most consistent finding in studies concerning blood lipids [130], whereas increasing HDL-C levels by exercise remains controversial, especially in those subjects with high TG and low HDL-C levels [131].
Other dietary factors besides calorie restriction and the type of dietary fat have also been shown to have beneficial effects on dyslipidemia. Dietary intake of resistant starch, a dietary fiber, has been shown to improve nutrient absorption and has also been linked to insulin metabolism. Daily intake of resistant starch from bread, cereals, vegetables and pastas is approximately 5 g/day in the Western world, which is highly insufficient for potential health benefits [132]. Recently, a randomized study in 15 insulin resistant subjects has shown that 8 weeks of resistant starch supplementation (40 g/day) improved insulin resistance and subsequently FFA metabolism. Resistant starch ingestion resulted in lower fasting FFA concentrations, increased TG lipolysis by enhanced expression of related genes like LPL together with increased FFA uptake by skeletal muscle [133]. However, no effect from resistant starch supplementation was observed on TG and cholesterol concentrations [132,133].
Unfortunately, lifestyle modifications are often insufficient to achieve weight loss and improvement of the dyslipidemia. A recent meta-analysis concerning anti-obesity drugs reported a mean weight loss of 3.13 kg, but marked improvements in dyslipidemia were absent [134]. Orlistat, which reduces the lipolysis of TG within the gastrointestinal system and thus prevents absorption of intestinal fat by 30%, showed only a modest reduction in LDL-C of 0.21 mmol/L. Sibutramine, which increases the sensation of satiety by modulating the central nervous system, showed a 0.13 mmol/L reduction in TG, whereas rimonabant did not show any lipid improvements [134]. Finally, bariatric surgery-induced weight loss has been associated with decreased TG and increased HDL-C levels [135].
6. Lipid Targets and the Pharmacological Treatment of Dyslipidemia in Obesity
The EAS/ESC guidelines recommend to profile lipids in obese subjects in order to assess cardiovascular risk [136]. However, the necessity to initiate pharmacological treatment next to lifestyle intervention in obese subjects with dyslipidemia depends on co-morbidity, the potential underlying primary lipid disorders and the calculated cardiovascular risk [11,136]. High risk subjects with primary lipid disorders like familial hypercholesterolemia or familial combined hyperlipidemia as well as subjects with known diabetes mellitus or cardiovascular disease all require appropriate pharmacological treatment independent from obesity [136,137]. Nevertheless, the presence of obesity can affect treatment targets since obesity may contribute to increased remnant cholesterol, higher TG levels and lower HDL-C concentrations. Therefore, apo B or non-HDL-C levels are recommended as secondary treatment targets next to LDL-C levels in the presence of the hypertriglyceridemic waist [11,136,138]. Apo B represents the total number of atherogenic particles (chylomicrons, chylomicron remnants, VLDL, IDL and LDL), whereas non-HDL-C represents the amount of cholesterol in both the TG-rich lipoproteins and LDL. Recently, a meta-analysis has shown that implementation of non-HDL-C or apo B as treatment target over LDL-C would prevent an additional 300,000–500,000 cardiovascular events in the US population over a 10-year period [139]. Although others did not describe any benefit of apo B or non-HDL-C over LDL-C levels to assess cardiovascular risk [140,141,142]. The treatment target for non-HDL-C should be 0.8 mmol/L higher than the target for LDL-C, which corresponds with non-HDL-C levels of 3.8 mmol/L and 3.3 mmol/L for subjects at moderate and high risk, respectively. Treatment targets for apo B are approximately 0.80–1.00 g/L [136]. Specific treatment targets for TG levels are unavailable, especially since TG are highly variable and increase during the day [143]. However, pharmacological interventions to lower specifically TG should be initiated when TG exceed 10 mmol/L to reduce the risk for pancreatitis [11,144]. In addition, additional diagnostic tests are warranted to test for the presence of familial hypertriglyceridemia or familial dysbetalipoproteinemia [11,136,138,144].
Statins are the first choice drug of all pharmacological agents to reduce LDL-C, non-HDL-C and/or apo B. However, statins lower TG only marginally and do not fully correct the characteristic dyslipidemia seen in obesity, which may contribute to the residual risk after initiating statin therapy [145]. Statins inhibit the enzyme 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA), which is the rate limiting step in the hepatic cholesterol synthesis. This efficiently increases the fractional catabolic rate of VLDL and LDL together with a slight reduction in hepatic secretion of VLDL. Therefore, statins lower both remnant cholesterol and LDL-C levels [146].
Recently, strategies for combination therapies with statins to achieve even lower cholesterol levels have been reviewed [145,146,147,148,149,150]. Combinations can be made with ezetimibe, which inhibits the intestinal cholesterol absorption by interaction with NPC1L1, which results in an additional 20% lowering effect on LDL-C, but without affecting TG or HDL-C concentrations. On the contrary, fibrates are primarily indicated in the case of hypertriglyceridemia and they reduce TG by approximately 30% and LDL-C by 8%, whereas HDL-C is increased by an average of 9% [149]. Fibrates (fibric acid derivatives) are peroxisome proliferator-activated receptor-α agonists, which transcriptionally regulate lipid metabolism related genes. Fibrates as monotherapy have been shown to reduce cardiovascular mortality, especially in subjects with characteristics of the MetS with TG levels > 2.20 mmol/L [151,152,153,154,155]. However, there is controversy about the effectiveness of fibrate therapy on top of statin therapy since the ACCORD trial was unable to confirm a beneficial effect on cardiovascular endpoints by fenofibrate combined with statins in diabetic patients [156]. Although subgroup analyses suggested a beneficial effect from combination therapy of fibrates with statins in patients with diabetes and the characteristic dyslipidemia with high TG and low HDL-C [156]. Therefore, fenofibrate may be used to treat residual dyslipidemia in diabetic patients on top of statin therapy [145].
Nicotinic acid inhibits the lipolysis of adipocytes, which results in decreased FFA levels, reduced VLDL synthesis, a slight increase in HDL production rate and decreased catabolism of HDL [146]. These changes by niacin subsequently lead to 15%–35% lower TG levels and 10%–25% higher HDL-C concentrations [11,146]. Recently, it has been shown that the addition of niacin to patients with a known history of cardiovascular disease, typical dyslipidemia and intensively controlled LDL-C levels with statin therapy did not lead to clinical benefit despite a reduction in fasting TG and increase in HDL-C [157]. However, specific data concerning combination therapy of niacin with statins in obesity remains scarce. Omega-3 fatty acids, which decrease the hepatic synthesis and accumulation of TG [158], have been shown to reduce plasma TG by 25%–30% by effectively reducing the hepatic secretion of VLDL in insulin resistant subjects [146,159]. Omega-3 fatty acids have also been shown to increase the conversion of VLDL into IDL, which suggests an additional benefit for combining omega-3 fatty acids with statins by increased catabolism of VLDL, IDL and LDL [159].
Drugs that increase insulin sensitivity like metformin or thiazolidinedione derivatives, have no [108] or minimal effects on the lipoprotein profile in obesity [160]. In the case of thiazolidinedione derivatives, their mode of action causes an increase of body weight, due to expansion of the subcutaneous fat compartment, which makes these drugs less appropriate in the case of obesity [160].
7. Conclusions
The pathophysiology of the typical dyslipidemia observed in obesity is multifactorial and includes hepatic overproduction of VLDL, decreased circulating TG lipolysis and impaired peripheral FFA trapping, increased FFA fluxes from adipocytes to the liver and other tissues and the formation of small dense LDL. Impairment of the ASP/C3adesArg pathway probably contributes to the typical dyslipidemia as well. Treatment should be aimed at weight loss by increased exercise and improved dietary habits with a reduction in total calorie intake and reduced SFA intake. Medical therapy can be initiated if lifestyle changes are insufficient. Statins are the primary lipid lowering drugs with effective reductions in LDL and remnant cholesterol levels. Moreover, the addition of fibrates may be considered in case of residual dyslipidemia in subjects with diabetes mellitus, elevated TG and reduced HDL-C levels. ApoB and/or non-HDL-C concentrations reflect the atherogenic lipid burden more accurately than LDL-C alone in obesity and should be used as treatment targets.
Conflict of Interests
The authors declare not to have any conflicts of interest.
References
- Knight, J.A. Diseases and disorders associated with excess body weight. Ann. Clin. Lab Sci. 2011, 41, 107–121. [Google Scholar]
- Flock, M.R.; Green, M.H.; Kris-Etherton, P.M. Effects of adiposity on plasma lipid response to reductions in dietary saturated fatty acids and cholesterol. Adv. Nutr. 2011, 2, 261–274. [Google Scholar] [CrossRef]
- Boden, G. Obesity, insulin resistance and free fatty acids. Curr. Opin. Endocrinol. Diabetes Obes. 2011, 18, 139–143. [Google Scholar] [CrossRef]
- Zalesin, K.C.; Franklin, B.A.; Miller, W.M.; Peterson, E.D.; McCullough, P.A. Impact of obesity on cardiovascular disease. Med. Clin. North. Am. 2011, 95, 919–937. [Google Scholar] [CrossRef]
- Castro Cabezas, M.; Elte, J.W. Farewell to the metabolic syndrome? Not too soon. Atherosclerosis 2009, 204, 348–349; author reply 350–351. [Google Scholar]
- Franssen, R.; Monajemi, H.; Stroes, E.S.; Kastelein, J.J. Obesity and dyslipidemia. Med. Clin. North. Am. 2011, 95, 893–902. [Google Scholar] [CrossRef]
- Wang, H.; Peng, D.Q. New insights into the mechanism of low high-density lipoprotein cholesterol in obesity. Lipids Health Dis. 2011, 10. [Google Scholar] [CrossRef]
- Pan, X.; Hussain, M.M. Gut triglyceride production. Biochim. Biophys. Acta 2011, 1821, 727–735. [Google Scholar]
- Altmann, S.W.; Davis, H.R., Jr.; Zhu, L.J.; Yao, X.; Hoos, L.M.; Tetzloff, G.; Iyer, S.P.; Maguire, M.; Golovko, A.; Zeng, M.; et al. Niemann-Pick C1 Like 1 protein is critical for intestinal cholesterol absorption. Science 2004, 303, 1201–1204. [Google Scholar] [CrossRef]
- Davis, H.R., Jr.; Zhu, L.J.; Hoos, L.M.; Tetzloff, G.; Maguire, M.; Liu, J.; Yao, X.; Iyer, S.P.; Lam, M.H.; Lund, E.G.; et al. Niemann-Pick C1 Like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 2004, 279, 33586–33592. [Google Scholar] [CrossRef]
- Klop, B.; Jukema, J.W.; Rabelink, T.J.; Castro Cabezas, M. A physician’s guide for the management of hypertriglyceridemia: The etiology of hypertriglyceridemia determines treatment strategy. Panminerva Med. 2012, 54, 91–103. [Google Scholar]
- Innerarity, T.L.; Young, S.G.; Poksay, K.S.; Mahley, R.W.; Smith, R.S.; Milne, R.W.; Marcel, Y.L.; Weisgraber, K.H. Structural relationship of human apolipoprotein B48 to apolipoprotein B100. J. Clin. Invest. 1987, 80, 1794–1798. [Google Scholar] [CrossRef]
- Goldberg, I.J.; Eckel, R.H.; Abumrad, N.A. Regulation of fatty acid uptake into tissues: lipoprotein lipase- and CD36-mediated pathways. J. Lipid Res. 2009, 50, S86–S90. [Google Scholar] [CrossRef]
- Dallinga-Thie, G.M.; Franssen, R.; Mooij, H.L.; Visser, M.E.; Hassing, H.C.; Peelman, F.; Kastelein, J.J.; Peterfy, M.; Nieuwdorp, M. The metabolism of triglyceride-rich lipoproteins revisited: new players, new insight. Atherosclerosis 2010, 211, 1–8. [Google Scholar] [CrossRef]
- Davies, B.S.; Beigneux, A.P.; Barnes, R.H., II; Tu, Y.; Gin, P.; Weinstein, M.M.; Nobumori, C.; Nyren, R.; Goldberg, I.; Olivecrona, G.; et al. GPIHBP1 is responsible for the entry of lipoprotein lipase into capillaries. Cell Metab. 2010, 12, 42–52. [Google Scholar] [CrossRef]
- Davies, B.S.; Beigneux, A.P.; Fong, L.G.; Young, S.G. New wrinkles in lipoprotein lipase biology. Curr. Opin. Lipidol. 2012, 23, 35–42. [Google Scholar] [CrossRef]
- Karpe, F.; Dickmann, J.R.; Frayn, K.N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation. Diabetes 2011, 60, 2441–2449. [Google Scholar] [CrossRef]
- McQuaid, S.E.; Hodson, L.; Neville, M.J.; Dennis, A.L.; Cheeseman, J.; Humphreys, S.M.; Ruge, T.; Gilbert, M.; Fielding, B.A.; Frayn, K.N.; et al. Downregulation of adipose tissue fatty acid trafficking in obesity: A driver for ectopic fat deposition? Diabetes 2011, 60, 47–55. [Google Scholar] [CrossRef]
- Ooi, E.M.; Barrett, P.H.; Chan, D.C.; Watts, G.F. Apolipoprotein C-III: Understanding an emerging cardiovascular risk factor. Clin. Sci. (Lond.) 2008, 114, 611–624. [Google Scholar] [CrossRef]
- Brunzell, J.D.; Hazzard, W.R.; Porte, D., Jr.; Bierman, E.L. Evidence for a common, saturable, triglyceride removal mechanism for chylomicrons and very low density lipoproteins in man. J. Clin. Invest. 1973, 52, 1578–1585. [Google Scholar] [CrossRef]
- Baldo, A.; Sniderman, A.D.; St-Luce, S.; Avramoglu, R.K.; Maslowska, M.; Hoang, B.; Monge, J.C.; Bell, A.; Mulay, S.; Cianflone, K. The adipsin-acylation stimulating protein system and regulation of intracellular triglyceride synthesis. J. Clin. Invest. 1993, 92, 1543–1547. [Google Scholar] [CrossRef]
- Germinario, R.; Sniderman, A.D.; Manuel, S.; Lefebvre, S.P.; Baldo, A.; Cianflone, K. Coordinate regulation of triacylglycerol synthesis and glucose transport by acylation-stimulating protein. Metabolism 1993, 42, 574–580. [Google Scholar] [CrossRef]
- Abumrad, N.A.; Davidson, N.O. Role of the gut in lipid homeostasis. Physiol. Rev. 2012, 92, 1061–1085. [Google Scholar] [CrossRef]
- Evans, K.; Burdge, G.C.; Wootton, S.A.; Clark, M.L.; Frayn, K.N. Regulation of dietary fatty acid entrapment in subcutaneous adipose tissue and skeletal muscle. Diabetes 2002, 51, 2684–2690. [Google Scholar] [CrossRef]
- Mahley, R.W.; Ji, Z.S. Remnant lipoprotein metabolism: key pathways involving cell-surface heparan sulfate proteoglycans and apolipoprotein E. J. Lipid Res. 1999, 40, 1–16. [Google Scholar]
- Mahley, R.W.; Huang, Y.; Rall, S.C., Jr. Pathogenesis of type III hyperlipoproteinemia (dysbetalipoproteinemia). Questions, quandaries, and paradoxes. J. Lipid Res. 1999, 40, 1933–1949. [Google Scholar]
- Sultan, F.; Lagrange, D.; Jansen, H.; Griglio, S. Inhibition of hepatic lipase activity impairs chylomicron remnant-removal in rats. Biochim. Biophys. Acta 1990, 1042, 150–152. [Google Scholar] [CrossRef]
- Kowal, R.C.; Herz, J.; Goldstein, J.L.; Esser, V.; Brown, M.S. Low density lipoprotein receptor-related protein mediates uptake of cholesteryl esters derived from apoprotein E-enriched lipoproteins. Proc. Natl. Acad. Sci. USA 1989, 86, 5810–5814. [Google Scholar] [CrossRef]
- Hussain, M.M.; Maxfield, F.R.; Mas-Oliva, J.; Tabas, I.; Ji, Z.S.; Innerarity, T.L.; Mahley, R.W. Clearance of chylomicron remnants by the low density lipoprotein receptor-related protein/alpha 2-macroglobulin receptor. J. Biol. Chem. 1991, 266, 13936–13940. [Google Scholar]
- Beisiegel, U.; Weber, W.; Bengtsson-Olivecrona, G. Lipoprotein lipase enhances the binding of chylomicrons to low density lipoprotein receptor-related protein. Proc. Natl. Acad. Sci. USA 1991, 88, 8342–8346. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. The LDL receptor. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 431–438. [Google Scholar] [CrossRef]
- Lambert, G.; Sjouke, B.; Choque, B.; Kastelein, J.J.; Hovingh, G.K. The PCSK9 decade: Thematic Review Series: New Lipid and Lipoprotein Targets for the Treatment of Cardiometabolic Diseases. J. Lipid Res. 2012, 53, 2515–2524. [Google Scholar] [CrossRef]
- Raal, F.; Scott, R.; Somaratne, R.; Bridges, I.; Li, G.; Wasserman, S.M.; Stein, E.A. Low-density lipoprotein cholesterol-lowering effects of AMG 145, a monoclonal antibody to proprotein convertase subtilisin/kexin type 9 serine protease in patients with heterozygous familial hypercholesterolemia: The reduction of LDL-C with PCSK9 inhibition in heterozygous familial hypercholesterolemia disorder (RUTHERFORD) randomized trial. Circulation 2012, 126, 2408–2417. [Google Scholar] [CrossRef]
- Patsch, J.R.; Miesenbock, G.; Hopferwieser, T.; Muhlberger, V.; Knapp, E.; Dunn, J.K.; Gotto, A.M., Jr.; Patsch, W. Relation of triglyceride metabolism and coronary artery disease. Studies in the postprandial state. Arterioscler. Thromb. 1992, 12, 1336–1345. [Google Scholar] [CrossRef]
- Engelberg, H. Serum lipemia: An overlooked cause of tissue hypoxia. Cardiology 1983, 70, 273–279. [Google Scholar] [CrossRef]
- Simons, L.A.; Dwyer, T.; Simons, J.; Bernstein, L.; Mock, P.; Poonia, N.S.; Balasubramaniam, S.; Baron, D.; Branson, J.; Morgan, J.; et al. Chylomicrons and chylomicron remnants in coronary artery disease: A case-control study. Atherosclerosis 1987, 65, 181–189. [Google Scholar] [CrossRef]
- Ryu, J.E.; Howard, G.; Craven, T.E.; Bond, M.G.; Hagaman, A.P.; Crouse, J.R., III. Postprandial triglyceridemia and carotid atherosclerosis in middle-aged subjects. Stroke 1992, 23, 823–828. [Google Scholar] [CrossRef]
- Karpe, F.; Steiner, G.; Uffelman, K.; Olivecrona, T.; Hamsten, A. Postprandial lipoproteins and progression of coronary atherosclerosis. Atherosclerosis 1994, 106, 83–97. [Google Scholar] [CrossRef]
- Meyer, E.; Westerveld, H.T.; de Ruyter-Meijstek, F.C.; van Greevenbroek, M.M.; Rienks, R.; van Rijn, H.J.; Erkelens, D.W.; de Bruin, T.W. Abnormal postprandial apolipoprotein B-48 and triglyceride responses in normolipidemic women with greater than 70% stenotic coronary artery disease: A case-control study. Atherosclerosis 1996, 124, 221–235. [Google Scholar] [CrossRef]
- Groot, P.H.; van Stiphout, W.A.; Krauss, X.H.; Jansen, H.; van Tol, A.; van Ramshorst, E.; Chin-On, S.; Hofman, A.; Cresswell, S.R.; Havekes, L. Postprandial lipoprotein metabolism in normolipidemic men with and without coronary artery disease. Arterioscler. Thromb. 1991, 11, 653–662. [Google Scholar] [CrossRef]
- Ginsberg, H.N.; Jones, J.; Blaner, W.S.; Thomas, A.; Karmally, W.; Fields, L.; Blood, D.; Begg, M.D. Association of postprandial triglyceride and retinyl palmitate responses with newly diagnosed exercise-induced myocardial ischemia in middle-aged men and women. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 1829–1838. [Google Scholar] [CrossRef]
- Sakata, K.; Miho, N.; Shirotani, M.; Yoshida, H.; Takada, Y.; Takada, A. Remnant-like particle cholesterol is a major risk factor for myocardial infarction in vasospastic angina with nearly normal coronary artery. Atherosclerosis 1998, 136, 225–231. [Google Scholar] [CrossRef]
- Ellsworth, J.L.; Fong, L.G.; Kraemer, F.B.; Cooper, A.D. Differences in the processing of chylomicron remnants and β-VLDL by macrophages. J. Lipid Res. 1990, 31, 1399–1411. [Google Scholar]
- Genest, J.; Sniderman, A.; Cianflone, K.; Teng, B.; Wacholder, S.; Marcel, Y.; Kwiterovich, P., Jr. Hyperapobetalipoproteinemia. Plasma lipoprotein responses to oral fat load. Arteriosclerosis 1986, 6, 297–304. [Google Scholar] [CrossRef]
- Castro Cabezas, M.; de Bruin, T.W.; Jansen, H.; Kock, L.A.; Kortlandt, W.; Erkelens, D.W. Impaired chylomicron remnant clearance in familial combined hyperlipidemia. Arterioscler. Thromb. 1993, 13, 804–814. [Google Scholar] [CrossRef]
- Castro Cabezas, M.; de Bruin, T.W.; de Valk, H.W.; Shoulders, C.C.; Jansen, H.; Willem Erkelens, D. Impaired fatty acid metabolism in familial combined hyperlipidemia. A mechanism associating hepatic apolipoprotein B overproduction and insulin resistance. J. Clin. Invest. 1993, 92, 160–168. [Google Scholar] [CrossRef]
- Castro Cabezas, M.; de Bruin, T.W.; Kock, L.A.; Kortlandt, W.; Van Linde-Sibenius Trip, M.; Jansen, H.; Erkelens, D.W. Simvastatin improves chylomicron remnant removal in familial combined hyperlipidemia without changing chylomicron conversion. Metabolism 1993, 42, 497–503. [Google Scholar] [CrossRef]
- Capell, W.H.; Zambon, A.; Austin, M.A.; Brunzell, J.D.; Hokanson, J.E. Compositional differences of LDL particles in normal subjects with LDL subclass phenotype A and LDL subclass phenotype B. Arterioscler Thromb. Vasc. Biol. 1996, 16, 1040–1046. [Google Scholar] [CrossRef]
- Hokanson, J.E.; Krauss, R.M.; Albers, J.J.; Austin, M.A.; Brunzell, J.D. LDL physical and chemical properties in familial combined hyperlipidemia. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 452–459. [Google Scholar] [CrossRef]
- Clemente-Postigo, M.; Queipo-Ortuno, M.I.; Fernandez-Garcia, D.; Gomez-Huelgas, R.; Tinahones, F.J.; Cardona, F. Adipose tissue gene expression of factors related to lipid processing in obesity. PLoS One 2011, 6, e24783. [Google Scholar] [CrossRef]
- Peterson, J.; Bihain, B.E.; Bengtsson-Olivecrona, G.; Deckelbaum, R.J.; Carpentier, Y.A.; Olivecrona, T. Fatty acid control of lipoprotein lipase: a link between energy metabolism and lipid transport. Proc. Natl. Acad. Sci. USA 1990, 87, 909–913. [Google Scholar] [CrossRef]
- Karpe, F.; Olivecrona, T.; Walldius, G.; Hamsten, A. Lipoprotein lipase in plasma after an oral fat load: Relation to free fatty acids. J. Lipid Res. 1992, 33, 975–984. [Google Scholar]
- Tchernof, A.; Lamarche, B.; Prud’Homme, D.; Nadeau, A.; Moorjani, S.; Labrie, F.; Lupien, P.J.; Despres, J.P. The dense LDL phenotype. Association with plasma lipoprotein levels, visceral obesity, and hyperinsulinemia in men. Diabetes Care 1996, 19, 629–637. [Google Scholar]
- Packard, C.J. Triacylglycerol-rich lipoproteins and the generation of small, dense low-density lipoprotein. Biochem. Soc. Trans. 2003, 31, 1066–1069. [Google Scholar] [CrossRef]
- Klop, B.; Proctor, S.D.; Mamo, J.C.; Botham, K.M.; Castro Cabezas, M. Understanding postprandial inflammation and its relationship to lifestyle behaviour and metabolic diseases. Int. J. Vasc. Med. 2012, 2012, 947417. [Google Scholar] [CrossRef]
- Proctor, S.D.; Mamo, J.C. Intimal retention of cholesterol derived from apolipoprotein B100- and apolipoprotein B48-containing lipoproteins in carotid arteries of Watanabe heritable hyperlipidemic rabbits. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1595–1600. [Google Scholar] [CrossRef]
- Proctor, S.D.; Vine, D.F.; Mamo, J.C. Arterial retention of apolipoprotein B(48)- and B(100)-containing lipoproteins in atherogenesis. Curr. Opin. Lipidol. 2002, 13, 461–470. [Google Scholar] [CrossRef]
- Tabas, I.; Williams, K.J.; Boren, J. Subendothelial lipoprotein retention as the initiating process in atherosclerosis: Update and therapeutic implications. Circulation 2007, 116, 1832–1844. [Google Scholar] [CrossRef]
- Klop, B.; Castro Cabezas, M. Chylomicrons: A key biomarker and risk factor for cardiovascular disease and for the understanding of obesity. Curr. Cardiovasc. Risk. Rep. 2012, 6, 27–34. [Google Scholar] [CrossRef]
- Subramanian, S.; Chait, A. Hypertriglyceridemia secondary to obesity and diabetes. Biochim. Biophys. Acta 2012, 1821, 819–825. [Google Scholar] [CrossRef]
- Pacifico, L.; Anania, C.; Osborn, J.F.; Ferraro, F.; Bonci, E.; Olivero, E.; Chiesa, C. Low 25(OH)D3 levels are associated with total adiposity, metabolic syndrome, and hypertension in Caucasian children and adolescents. Eur. J. Endocrinol. 2011, 165, 603–611. [Google Scholar] [CrossRef]
- Proctor, S.D.; Vine, D.F.; Mamo, J.C. Arterial permeability and efflux of apolipoprotein B-containing lipoproteins assessed by in situ perfusion and three-dimensional quantitative confocal microscopy. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2162–2167. [Google Scholar] [CrossRef]
- Watts, G.F.; Chan, D.C.; Barrett, P.H.; Martins, I.J.; Redgrave, T.G. Preliminary experience with a new stable isotope breath test for chylomicron remnant metabolism: A study in central obesity. Clin. Sci. (Lond.) 2001, 101, 683–690. [Google Scholar] [CrossRef]
- Taskinen, M.R.; Adiels, M.; Westerbacka, J.; Soderlund, S.; Kahri, J.; Lundbom, N.; Lundbom, J.; Hakkarainen, A.; Olofsson, S.O.; Orho-Melander, M.; et al. Dual metabolic defects are required to produce hypertriglyceridemia in obese subjects. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2144–2150. [Google Scholar] [CrossRef]
- Caron, S.; Verrijken, A.; Mertens, I.; Samanez, C.H.; Mautino, G.; Haas, J.T.; Duran-Sandoval, D.; Prawitt, J.; Francque, S.; Vallez, E.; et al. Transcriptional activation of apolipoprotein CIII expression by glucose may contribute to diabetic dyslipidemia. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 513–519. [Google Scholar] [CrossRef]
- Mamo, J.C.; Watts, G.F.; Barrett, P.H.; Smith, D.; James, A.P.; Pal, S. Postprandial dyslipidemia in men with visceral obesity: An effect of reduced LDL receptor expression? Am. J. Physiol. Endocrinol. Metab. 2001, 281, E626–E632. [Google Scholar]
- Castro Cabezas, M.; Erkelens, D.W. The direct way from gut to vessel wall: Atheroinitiation. Eur. J. Clin. Invest. 1998, 28, 504–505. [Google Scholar] [CrossRef]
- Jorgensen, A.B.; Frikke-Schmidt, R.; West, A.S.; Grande, P.; Nordestgaard, B.G.; Tybjaerg-Hansen, A. Genetically elevated non-fasting triglycerides and calculated remnant cholesterol as causal risk factors for myocardial infarction. Eur. Heart J. 2012. [Google Scholar] [CrossRef]
- Senti, M.; Nogues, X.; Pedro-Botet, J.; Rubies-Prat, J.; Vidal-Barraquer, F. Lipoprotein profile in men with peripheral vascular disease. Role of intermediate density lipoproteins and apoprotein E phenotypes. Circulation 1992, 85, 30–36. [Google Scholar] [CrossRef]
- Vogel, R.A.; Corretti, M.C.; Plotnick, G.D. Effect of a single high-fat meal on endothelial function in healthy subjects. Am. J. Cardiol. 1997, 79, 350–354. [Google Scholar] [CrossRef]
- Lundman, P.; Eriksson, M.; Schenck-Gustafsson, K.; Karpe, F.; Tornvall, P. Transient triglyceridemia decreases vascular reactivity in young, healthy men without risk factors for coronary heart disease. Circulation 1997, 96, 3266–3268. [Google Scholar] [CrossRef]
- Steinberg, H.O.; Tarshoby, M.; Monestel, R.; Hook, G.; Cronin, J.; Johnson, A.; Bayazeed, B.; Baron, A.D. Elevated circulating free fatty acid levels impair endothelium-dependent vasodilation. J. Clin. Invest. 1997, 100, 1230–1239. [Google Scholar] [CrossRef]
- Van Oostrom, A.J.; van Wijk, J.; Castro Cabezas, M. Lipaemia, inflammation and atherosclerosis: Novel opportunities in the understanding and treatment of atherosclerosis. Drugs 2004, 64, 19–41. [Google Scholar] [CrossRef]
- Alipour, A.; Elte, J.W.; van Zaanen, H.C.; Rietveld, A.P.; Castro Cabezas, M. Novel aspects of postprandial lipemia in relation to atherosclerosis. Atheroscler. Suppl. 2008, 9, 39–44. [Google Scholar]
- Couillard, C.; Bergeron, N.; Prud’homme, D.; Bergeron, J.; Tremblay, A.; Bouchard, C.; Mauriege, P.; Despres, J.P. Postprandial triglyceride response in visceral obesity in men. Diabetes 1998, 47, 953–960. [Google Scholar] [CrossRef]
- Taira, K.; Hikita, M.; Kobayashi, J.; Bujo, H.; Takahashi, K.; Murano, S.; Morisaki, N.; Saito, Y. Delayed post-prandial lipid metabolism in subjects with intra-abdominal visceral fat accumulation. Eur. J. Clin. Invest. 1999, 29, 301–308. [Google Scholar] [CrossRef]
- Su, J.W.; Nzekwu, M.M.; Cabezas, M.C.; Redgrave, T.; Proctor, S.D. Methods to assess impaired post-prandial metabolism and the impact for early detection of cardiovascular disease risk. Eur. J. Clin. Invest. 2009, 39, 741–754. [Google Scholar] [CrossRef]
- Castro Cabezas, M.; Halkes, C.J.; Meijssen, S.; van Oostrom, A.J.; Erkelens, D.W. Diurnal triglyceride profiles: A novel approach to study triglyceride changes. Atherosclerosis 2001, 155, 219–228. [Google Scholar] [CrossRef]
- Van Oostrom, A.J.; Castro Cabezas, M.; Ribalta, J.; Masana, L.; Twickler, T.B.; Remijnse, T.A.; Erkelens, D.W. Diurnal triglyceride profiles in healthy normolipidemic male subjects are associated to insulin sensitivity, body composition and diet. Eur. J. Clin. Invest. 2000, 30, 964–971. [Google Scholar] [CrossRef]
- Halkes, C.J.; Castro Cabezas, M.; van Wijk, J.P.; Erkelens, D.W. Gender differences in diurnal triglyceridemia in lean and overweight subjects. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 1767–1774. [Google Scholar] [CrossRef]
- Lean, M.E.; Han, T.S.; Morrison, C.E. Waist circumference as a measure for indicating need for weight management. Br. Med. J. 1995, 311, 158–161. [Google Scholar] [CrossRef]
- Van Gaal, L.F.; Mertens, I.L.; de Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar]
- Deeb, S.S.; Zambon, A.; Carr, M.C.; Ayyobi, A.F.; Brunzell, J.D. Hepatic lipase and dyslipidemia: Interactions among genetic variants, obesity, gender, and diet. J. Lipid Res. 2003, 44, 1279–1286. [Google Scholar] [CrossRef]
- Hill, M.J.; Metcalfe, D.; McTernan, P.G. Obesity and diabetes: Lipids, “nowhere to run to”. Clin. Sci. (Lond.) 2009, 116, 113–123. [Google Scholar] [CrossRef]
- Bjorntorp, P.; Bergman, H.; Varnauskas, E. Plasma free fatty acid turnover rate in obesity. Acta Med. Scand. 1969, 185, 351–356. [Google Scholar] [CrossRef]
- Mook, S.; Halkes, C.C.; Bilecen, S.; Castro Cabezas, M. In vivo regulation of plasma free fatty acids in insulin resistance. Metabolism 2004, 53, 1197–1201. [Google Scholar] [CrossRef]
- Van Oostrom, A.J.; van Dijk, H.; Verseyden, C.; Sniderman, A.D.; Cianflone, K.; Rabelink, T.J.; Castro Cabezas, M. Addition of glucose to an oral fat load reduces postprandial free fatty acids and prevents the postprandial increase in complement component 3. Am. J. Clin. Nutr. 2004, 79, 510–515. [Google Scholar]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vascul. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef]
- Lottenberg, A.M.; Afonso Mda, S.; Lavrador, M.S.; Machado, R.M.; Nakandakare, E.R. The role of dietary fatty acids in the pathology of metabolic syndrome. J. Nutr. Biochem. 2012, 23, 1027–1040. [Google Scholar] [CrossRef]
- Sears, B.; Ricordi, C. Role of fatty acids and polyphenols in inflammatory gene transcription and their impact on obesity, metabolic syndrome and diabetes. Eur. Rev. Med. Pharmacol. Sci. 2012, 16, 1137–1154. [Google Scholar]
- Kopp, A.; Gross, P.; Falk, W.; Bala, M.; Weigert, J.; Buechler, C.; Neumeier, M.; Scholmerich, J.; Schaffler, A. Fatty acids as metabolic mediators in innate immunity. Eur. J. Clin. Invest. 2009, 39, 924–933. [Google Scholar] [CrossRef]
- Sniderman, A.D.; Maslowska, M.; Cianflone, K. Of mice and men (and women) and the acylation-stimulating protein pathway. Curr. Opin. Lipidol. 2000, 11, 291–296. [Google Scholar] [CrossRef]
- Cianflone, K.; Vu, H.; Walsh, M.; Baldo, A.; Sniderman, A. Metabolic response of Acylation Stimulating Protein to an oral fat load. J. Lipid Res. 1989, 30, 1727–1733. [Google Scholar]
- Cianflone, K.; Maslowska, M. Differentiation-induced production of ASP in human adipocytes. Eur. J. Clin. Invest. 1995, 25, 817–825. [Google Scholar] [CrossRef]
- Maslowska, M.; Scantlebury, T.; Germinario, R.; Cianflone, K. Acute in vitro production of acylation stimulating protein in differentiated human adipocytes. J. Lipid Res. 1997, 38, 1–11. [Google Scholar]
- Saleh, J.; Summers, L.K.; Cianflone, K.; Fielding, B.A.; Sniderman, A.D.; Frayn, K.N. Coordinated release of acylation stimulating protein (ASP) and triacylglycerol clearance by human adipose tissue in vivo in the postprandial period. J. Lipid Res. 1998, 39, 884–891. [Google Scholar]
- Skidgel, R.A. Basic carboxypeptidases: Regulators of peptide hormone activity. Trends Pharmacol. Sci. 1988, 9, 299–304. [Google Scholar] [CrossRef]
- Maslowska, M.; Sniderman, A.D.; Germinario, R.; Cianflone, K. ASP stimulates glucose transport in cultured human adipocytes. Int. J. Obes. Relat. Metab. Disord. 1997, 21, 261–266. [Google Scholar]
- Van Oostrom, A.J.; Alipour, A.; Plokker, T.W.; Sniderman, A.D.; Castro Cabezas, M. The metabolic syndrome in relation to complement component 3 and postprandial lipemia in patients from an outpatient lipid clinic and healthy volunteers. Atherosclerosis 2007, 190, 167–173. [Google Scholar] [CrossRef]
- Volp, A.C.; Barbosa, K.B.; Bressan, J. Triacylglycerols and body fat mass are possible independent predictors of C3 in apparently healthy young Brazilian adults. Nutrition 2012, 28, 544–550. [Google Scholar] [CrossRef]
- Hernandez-Mijares, A.; Banuls, C.; Bellod, L.; Jover, A.; Sola, E.; Morillas, C.; Victor, V.M.; Rocha, M. Effect of weight loss on C3 and C4 components of complement in obese patients. Eur. J. Clin. Invest. 2012, 42, 503–509. [Google Scholar] [CrossRef]
- Meijssen, S.; van Dijk, H.; Verseyden, C.; Erkelens, D.W.; Castro Cabezas, M. Delayed and exaggerated postprandial complement component 3 response in familial combined hyperlipidemia. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 811–816. [Google Scholar] [CrossRef]
- Halkes, C.J.; van Dijk, H.; de Jaegere, P.P.T.; Plokker, H.W.M.; van der Helm, Y.; Erkelens, D.W.; Castro Cabezas, M. Postprandial increase of complement component 3 in normolipidemic patients with coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1526–1530. [Google Scholar] [CrossRef]
- Kristiansson, K.; Perola, M.; Tikkanen, E.; Kettunen, J.; Surakka, I.; Havulinna, A.S.; Stancakova, A.; Barnes, C.; Widen, E.; Kajantie, E.; et al. Genome-wide screen for metabolic syndrome susceptibility Loci reveals strong lipid gene contribution but no evidence for common genetic basis for clustering of metabolic syndrome traits. Circ. Cardiovasc. Genet. 2012, 5, 242–249. [Google Scholar] [CrossRef]
- Alipour, A.; van Oostrom, A.J.; Van Wijk, J.P.; Verseyden, C.; Plokker, H.W.; Jukema, J.W.; Rabelink, A.J.; Castro Cabezas, M. Mannose binding lectin deficiency and triglyceride-rich lipoprotein metabolism in normolipidemic subjects. Atherosclerosis 2009, 206, 444–450. [Google Scholar] [CrossRef]
- Meijssen, S.; Castro Cabezas, M.; Twickler, T.B.; Jansen, H.; Erkelens, D.W. In vivo evidence of defective postprandial and postabsorptive free fatty acid metabolism in familial combined hyperlipidemia. J. Lipid Res. 2000, 41, 1096–1102. [Google Scholar]
- Halkes, C.J.; van Dijk, H.; Verseyden, C.; de Jaegere, P.P.; Plokker, H.W.; Meijssen, S.; Erkelens, D.W.; Castro Cabezas, M. Gender differences in postprandial ketone bodies in normolipidemic subjects and in untreated patients with familial combined hyperlipidemia. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1875–1880. [Google Scholar] [CrossRef]
- Castro Cabezas, M.; van Wijk, J.P.; Elte, J.W.; Klop, B. Effects of metformin on the regulation of free Fatty acids in insulin resistance: A double-blind, placebo-controlled study. J. Nutr. Metab. 2012, 2012. [Google Scholar] [CrossRef]
- Lopez-Miranda, J.; Williams, C.; Lairon, D. Dietary, physiological, genetic and pathological influences on postprandial lipid metabolism. Br. J. Nutr. 2007, 98, 458–473. [Google Scholar] [CrossRef]
- De Ruyter, J.C.; Olthof, M.R.; Seidell, J.C.; Katan, M.B. A trial of sugar-free or sugar-sweetened beverages and body weight in children. N. Engl. J. Med. 2012, 367, 1397–1406. [Google Scholar] [CrossRef]
- Patalay, M.; Lofgren, I.E.; Freake, H.C.; Koo, S.I.; Fernandez, M.L. The lowering of plasma lipids following a weight reduction program is related to increased expression of the LDL receptor and lipoprotein lipase. J. Nutr. 2005, 135, 735–739. [Google Scholar]
- Laimer, M.W.; Engl, J.; Tschoner, A.; Kaser, S.; Ritsch, A.; Tatarczyk, T.; Rauchenzauner, M.; Weiss, H.; Aigner, F.; Patsch, J.R.; et al. Effects of weight loss on lipid transfer proteins in morbidly obese women. Lipids 2009, 44, 1125–1130. [Google Scholar] [CrossRef]
- Wang, Y.; Snel, M.; Jonker, J.T.; Hammer, S.; Lamb, H.J.; de Roos, A.; Meinders, A.E.; Pijl, H.; Romijn, J.A.; Smit, J.W.; et al. Prolonged caloric restriction in obese patients with type 2 diabetes mellitus decreases plasma CETP and increases apolipoprotein AI levels without improving the cholesterol efflux properties of HDL. Diabetes Care 2011, 34, 2576–2580. [Google Scholar] [CrossRef]
- Chan, D.C.; Watts, G.F.; Barrett, P.H.; Mamo, J.C.; Redgrave, T.G. Markers of triglyceride-rich lipoprotein remnant metabolism in visceral obesity. Clin. Chem. 2002, 48, 278–283. [Google Scholar]
- James, A.P.; Watts, G.F.; Barrett, P.H.; Smith, D.; Pal, S.; Chan, D.C.; Mamo, J.C. Effect of weight loss on postprandial lipemia and low-density lipoprotein receptor binding in overweight men. Metabolism 2003, 52, 136–141. [Google Scholar] [CrossRef]
- Roberts, C.K.; Barnard, R.J.; Liang, K.H.; Vaziri, N.D. Effect of diet on adipose tissue and skeletal muscle VLDL receptor and LPL: Implications for obesity and hyperlipidemia. Atherosclerosis 2002, 161, 133–141. [Google Scholar] [CrossRef]
- Maraki, M.I.; Aggelopoulou, N.; Christodoulou, N.; Anastasiou, C.A.; Toutouza, M.; Panagiotakos, D.B.; Kavouras, S.A.; Magkos, F.; Sidossis, L.S. Lifestyle intervention leading to moderate weight loss normalizes postprandial triacylglycerolemia despite persisting obesity. Obesity (Silver Spring) 2011, 19, 968–976. [Google Scholar] [CrossRef]
- Cruz-Teno, C.; Perez-Martinez, P.; Delgado-Lista, J.; Yubero-Serrano, E.M.; Garcia-Rios, A.; Marin, C.; Gomez, P.; Jimenez-Gomez, Y.; Camargo, A.; Rodriguez-Cantalejo, F.; et al. Detary fat modifies the postprandial inflammatory state in subjects with metabolic syndrome: The LIPGENE study. Mol. Nutr. Food Res. 2012, 56, 854–865. [Google Scholar] [CrossRef]
- Yin, R.X.; Wu, D.F.; Miao, L.; Aung, L.H.; Cao, X.L.; Yan, T.T.; Long, X.J.; Liu, W.Y.; Zhang, L.; Li, M. Several genetic polymorphisms interact with overweight/obesity to influence serum lipid levels. Cardiovasc. Diabetol. 2012, 11. [Google Scholar] [CrossRef]
- Corella, D.; Peloso, G.; Arnett, D.K.; Demissie, S.; Cupples, L.A.; Tucker, K.; Lai, C.Q.; Parnell, L.D.; Coltell, O.; Lee, Y.C.; et al. APOA2, dietary fat, and body mass index: replication of a gene-diet interaction in 3 independent populations. Arch. Intern. Med. 2009, 169, 1897–1906. [Google Scholar] [CrossRef]
- Lai, C.Q.; Corella, D.; Demissie, S.; Cupples, L.A.; Adiconis, X.; Zhu, Y.; Parnell, L.D.; Tucker, K.L.; Ordovas, J.M. Dietary intake of n-6 fatty acids modulates effect of apolipoprotein A5 gene on plasma fasting triglycerides, remnant lipoprotein concentrations, and lipoprotein particle size: The Framingham Heart Study. Circulation 2006, 113, 2062–2070. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Ordovas, J.M.; Smith, C.E.; Baraza, J.C.; Lee, Y.C.; Garaulet, M. APOA5 gene variation interacts with dietary fat intake to modulate obesity and circulating triglycerides in a Mediterranean population. J. Nutr. 2012, 141, 380–385. [Google Scholar]
- Thomas, T.R.; Horner, K.E.; Langdon, M.M.; Zhang, J.Q.; Krul, E.S.; Sun, G.Y.; Cox, R.H. Effect of exercise and medium-chain fatty acids on postprandial lipemia. J. Appl. Physiol. 2001, 90, 1239–1246. [Google Scholar]
- Ferguson, M.A.; Alderson, N.L.; Trost, S.G.; Essig, D.A.; Burke, J.R.; Durstine, J.L. Effects of four different single exercise sessions on lipids, lipoproteins, and lipoprotein lipase. J. Appl. Physiol. 1998, 85, 1169–1174. [Google Scholar]
- Harrison, M.; Moyna, N.M.; Zderic, T.W.; O’Gorman, D.J.; McCaffrey, N.; Carson, B.P.; Hamilton, M.T. Lipoprotein particle distribution and skeletal muscle lipoprotein lipase activity after acute exercise. Lipids. Health Dis. 2012, 11. [Google Scholar] [CrossRef]
- Slivkoff-Clark, K.M.; James, A.P.; Mamo, J.C. The chronic effects of fish oil with exercise on postprandial lipaemia and chylomicron homeostasis in insulin resistant viscerally obese men. Nutr. Metab. (Lond.) 2012, 9, 9. [Google Scholar] [CrossRef]
- Sullivan, S.; Kirk, E.P.; Mittendorfer, B.; Patterson, B.W.; Klein, S. Randomized trial of exercise effect on intrahepatic triglyceride content and lipid kinetics in nonalcoholic fatty liver disease. Hepatology 2012, 55, 1738–1745. [Google Scholar] [CrossRef]
- Magkos, F. Exercise and fat accumulation in the human liver. Curr. Opin. Lipidol. 2010, 21, 507–517. [Google Scholar] [CrossRef]
- van Herpen, N.A.; Schrauwen-Hinderling, V.B.; Schaart, G.; Mensink, R.P.; Schrauwen, P. Three weeks on a high-fat diet increases intrahepatic lipid accumulation and decreases metabolic flexibility in healthy overweight men. J. Clin. Endocrinol. Metab. 2012, 96, E691–E695. [Google Scholar]
- Mestek, M.L. Physical activity, blood lipids, and lipoproteins. Am. J. Lifestyle Med. 2009, 3, 279–283. [Google Scholar] [CrossRef]
- Thompson, P.D.; Rader, D.J. Does exercise increase HDL cholesterol in those who need it the most? Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1097–1098. [Google Scholar] [CrossRef]
- Maki, K.C.; Pelkman, C.L.; Finocchiaro, E.T.; Kelley, K.M.; Lawless, A.L.; Schild, A.L.; Rains, T.M. Resistant starch from high-amylose maize increases insulin sensitivity in overweight and obese men. J. Nutr. 2012, 142, 717–723. [Google Scholar] [CrossRef]
- Robertson, M.D.; Wright, J.W.; Loizon, E.; Debard, C.; Vidal, H.; Shojaee-Moradie, F.; Russell-Jones, D.; Umpleby, A.M. Insulin-sensitizing effects on muscle and adipose tissue after dietary fiber intake in men and women with metabolic syndrome. J. Clin. Endocrinol. Metab. 2012, 97, 3326–3332. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Ma, X.Q.; Wu, C.; Lu, J.; Zhang, S.S.; Guo, J.; Wu, S.Q.; Ye, X.F.; Xu, J.F.; He, J. Effect of anti-obesity drug on cardiovascular risk factors: A systematic review and meta-analysis of randomized controlled trials. PLoS One 2012, 7, e39062. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Julia, Z.; Poitou, C.; Bouillot, J.L.; Basdevant, A.; Chapman, M.J.; Clement, K.; Guerin, M. Effect of bariatric surgery-induced weight loss on SR-BI-, ABCG1-, and ABCA1-mediated cellular cholesterol efflux in obese women. J. Clin. Endocrinol. Metab. 2011, 96, 1151–1159. [Google Scholar] [CrossRef]
- Catapano, A.L.; Reiner, Z.; de Backer, G.; Graham, I.; Taskinen, M.R.; Wiklund, O.; Agewall, S.; Alegria, E.; Chapman, M.J.; Durrington, P.; et al. ESC/EAS Guidelines for the management of dyslipidaemias: The Task Force for the management of dyslipidaemias of the European Society of Cardiology (ESC) and the European Atherosclerosis Society (EAS). Atherosclerosis 2011, 217, 1–44. [Google Scholar]
- Kushner, R.F. Clinincal assessment and management of adult obesity. Circulation 2012, 126, 2870–2877. [Google Scholar] [CrossRef]
- Berglund, L.; Brunzell, J.D.; Goldberg, A.C.; Goldberg, I.J.; Sacks, F.; Murad, M.H.; Stalenhoef, A.F. Evaluation and treatment of hypertriglyceridemia: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2012, 97, 2969–2989. [Google Scholar] [CrossRef]
- Sniderman, A.D.; Williams, K.; Contois, J.H.; Monroe, H.M.; McQueen, M.J.; de Graaf, J.; Furberg, C.D. A meta-analysis of low-density lipoprotein cholesterol, non-high-density lipoprotein cholesterol, and apolipoprotein B as markers of cardiovascular risk. Circ. Cardiovasc. Qual. Outcomes 2011, 4, 337–345. [Google Scholar] [CrossRef]
- Mora, S.; Glynn, R.J.; Boekholdt, S.M.; Nordestgaard, B.G.; Kastelein, J.J.; Ridker, P.M. On-treatment non-high-density lipoprotein cholesterol, apolipoprotein B, triglycerides, and lipid ratios in relation to residual vascular risk after treatment with potent statin therapy: JUPITER (justification for the use of statins in prevention: An intervention trial evaluating rosuvastatin). J. Am. Coll. Cardiol. 2012, 59, 1521–1528. [Google Scholar]
- Boekholdt, S.M.; Arsenault, B.J.; Mora, S.; Pedersen, T.R.; LaRosa, J.C.; Nestel, P.J.; Simes, R.J.; Durrington, P.; Hitman, G.A.; Welch, K.M.; et al. Association of LDL cholesterol, non-HDL cholesterol, and apolipoprotein B levels with risk of cardiovascular events among patients treated with statins: A meta-analysis. JAMA 2012, 307, 1302–1309. [Google Scholar] [CrossRef]
- Robinson, J.G.; Wang, S.; Jacobson, T.A. Meta-analysis of comparison of effectiveness of lowering apolipoprotein B versus low-density lipoprotein cholesterol and nonhigh-density lipoprotein cholesterol for cardiovascular risk reduction in randomized trials. Am. J. Cardiol 2012, 110, 1468–1476. [Google Scholar] [CrossRef]
- Klop, B.; Cohn, J.S.; van Oostrom, A.J.; van Wijk, J.P.; Birnie, E.; Castro Cabezas, M. Daytime triglyceride variability in men and women with different levels of triglyceridemia. Clin. Chim. Acta 2011, 412, 2183–2189. [Google Scholar] [CrossRef]
- Brunzell, J.D. Clinical practice. Hypertriglyceridemia. N. Engl. J. Med. 2007, 357, 1009–1017. [Google Scholar] [CrossRef]
- Watts, G.F.; Karpe, F. Triglycerides and atherogenic dyslipidaemia: Extending treatment beyond statins in the high-risk cardiovascular patient. Heart 2011, 97, 350–356. [Google Scholar] [CrossRef]
- Chan, D.C.; Watts, G.F. Dyslipidaemia in the metabolic syndrome and type 2 diabetes: Pathogenesis, priorities, pharmacotherapies. Expert Opin. Pharmacother. 2011, 12, 13–30. [Google Scholar] [CrossRef]
- Watts, G.F.; Karpe, F. Why, when and how should hypertriglyceridemia be treated in the high-risk cardiovascular patient? Expert. Rev. Cardiovasc. Ther. 2011, 9, 987–997. [Google Scholar] [CrossRef]
- Dujovne, C.A.; Williams, C.D.; Ito, M.K. What combination therapy with a statin, if any, would you recommend? Cur. Atheroscler. Rep. 2011, 13, 12–22. [Google Scholar]
- Rubenfire, M.; Brook, R.D.; Rosenson, R.S. Treating mixed hyperlipidemia and the atherogenic lipid phenotype for prevention of cardiovascular events. Am. J. Med. 2010, 123, 892–898. [Google Scholar] [CrossRef]
- Toth, P.P. Drug treatment of hyperlipidaemia: A guide to the rational use of lipid-lowering drugs. Drugs 2010, 70, 1363–1379. [Google Scholar] [CrossRef]
- Tenenbaum, A.; Motro, M.; Fisman, E.Z.; Tanne, D.; Boyko, V.; Behar, S. Bezafibrate for the secondary prevention of myocardial infarction in patients with metabolic syndrome. Arch. Intern. Me.d 2005, 165, 1154–1160. [Google Scholar]
- Tenkanen, L.; Manttari, M.; Manninen, V. Some coronary risk factors related to the insulin resistance syndrome and treatment with gemfibrozil. Experience from the Helsinki Heart Study. Circulation 1995, 92, 1779–1785. [Google Scholar] [CrossRef]
- Tenkanen, L.; Manttari, M.; Kovanen, P.T.; Virkkunen, H.; Manninen, V. Gemfibrozil in the treatment of dyslipidemia: An 18-year mortality follow-up of the Helsinki Heart Study. Arch. Intern. Med. 2006, 166, 743–748. [Google Scholar] [CrossRef]
- Rubins, H.B.; Robins, S.J.; Collins, D.; Nelson, D.B.; Elam, M.B.; Schaefer, E.J.; Faas, F.H.; Anderson, J.W. Diabetes, plasma insulin, and cardiovascular disease: subgroup analysis from the Department of Veterans Affairs high-density lipoprotein intervention trial (VA-HIT). Arch. Intern. Med. 2002, 162, 2597–2604. [Google Scholar] [CrossRef]
- Scott, R.; O’Brien, R.; Fulcher, G.; Pardy, C.; D’Emden, M.; Tse, D.; Taskinen, M.R.; Ehnholm, C.; Keech, A. Effects of fenofibrate treatment on cardiovascular disease risk in 9795 individuals with type 2 diabetes and various components of the metabolic syndrome: The Fenofibrate Intervention and Event Lowering in Diabetes (FIELD) study. Diabetes Care 2009, 32, 493–498. [Google Scholar] [CrossRef]
- Ginsberg, H.N.; Elam, M.B.; Lovato, L.C.; Crouse, J.R., III; Leiter, L.A.; Linz, P.; Friedewald, W.T.; Buse, J.B.; Gerstein, H.C.; Probstfield, J.; et al. Effects of combination lipid therapy in type 2 diabetes mellitus. N. Engl. J. Med. 2010, 362, 1563–1574. [Google Scholar] [CrossRef]
- Boden, W.E.; Probstfield, J.L.; Anderson, T.; Chaitman, B.R.; Desvignes-Nickens, P.; Koprowicz, K.; McBride, R.; Teo, K.; Weintraub, W. Niacin in patients with low HDL cholesterol levels receiving intensive statin therapy. N. Engl. J. Med. 2011, 365, 2255–2267. [Google Scholar] [CrossRef]
- Watts, G.F.; Chan, D.C.; Ooi, E.M.; Nestel, P.J.; Beilin, L.J.; Barrett, P.H. Fish oils, phytosterols and weight loss in the regulation of lipoprotein transport in the metabolic syndrome: Lessons from stable isotope tracer studies. Clin. Exp. Pharmacol. Physiol. 2006, 33, 877–882. [Google Scholar] [CrossRef]
- Chan, D.C.; Watts, G.F.; Barrett, P.H.; Beilin, L.J.; Redgrave, T.G.; Mori, T.A. Regulatory effects of HMG CoA reductase inhibitor and fish oils on apolipoprotein B-100 kinetics in insulin-resistant obese male subjects with dyslipidemia. Diabetes 2002, 51, 2377–2386. [Google Scholar] [CrossRef]
- Van Wijk, J.P.; de Koning, E.J.; Martens, E.P.; Rabelink, T.J. Thiazolidinediones and blood lipids in type 2 diabetes. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1744–1749. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218-1240. https://doi.org/10.3390/nu5041218
AMA Style
Klop B, Elte JWF, Cabezas MC. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients. 2013; 5(4):1218-1240. https://doi.org/10.3390/nu5041218
Chicago/Turabian StyleKlop, Boudewijn, Jan Willem F. Elte, and Manuel Castro Cabezas. 2013. "Dyslipidemia in Obesity: Mechanisms and Potential Targets" Nutrients 5, no. 4: 1218-1240. https://doi.org/10.3390/nu5041218