Dietary Cholesterol Affects Plasma Lipid Levels, the Intravascular Processing of Lipoproteins and Reverse Cholesterol Transport without Increasing the Risk for Heart Disease


Abstract
:1. Introduction
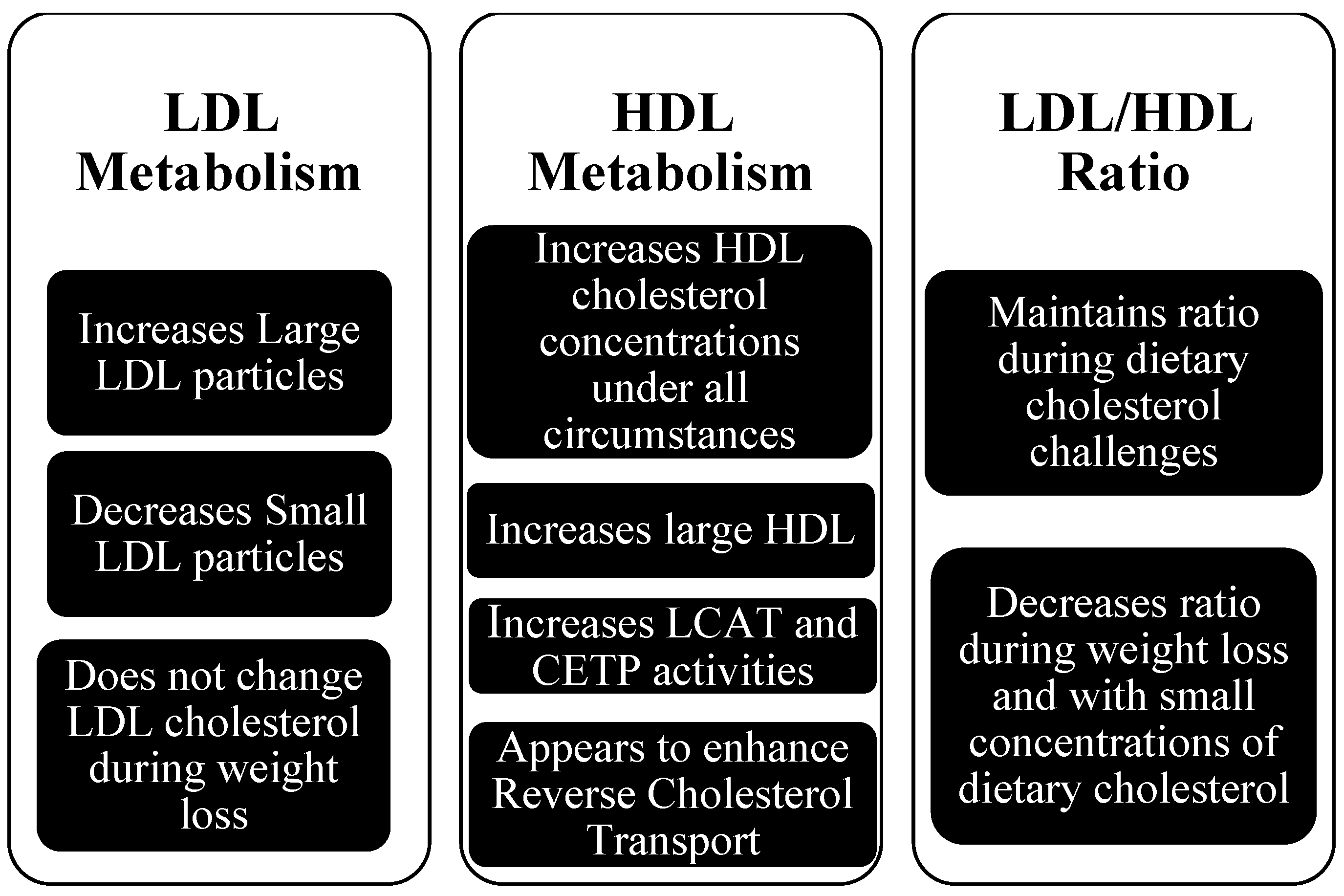
2. Dietary Cholesterol and Plasma LDL and HDL
3. Dietary Cholesterol and LDL
4. Dietary Cholesterol and HDL
5. Dietary Cholesterol and Reverse Cholesterol Transport
{kind=link}
6. Compensatory Mechanisms: Cholesterol Absorption and Cholesterol Synthesis
7. Conclusions
Conflict of Interest
References
- Imes, C.C.; Austin, M.A. Low-density lipoprotein cholesterol, apolipoprotein B, and risk of coronary heart disease: From familial hyperlipidemia to genomics. Biol. Res. Nurs. 2012, in press. [Google Scholar]
- Lim, S.; Park, Y.M.; Sakuma, I.; Koh, K.K. How to control residual cardiovascular risk despite statin treatment: Focusing on HDL cholesterol. Int. J. Cardiol. 2012, in press. [Google Scholar]
- Fernandez, M.L.; Webb, D. The LDL to HDL cholesterol ratio as a valuable tool to evaluate coronary heart disease risk. J. Am. Coll. Nutr. 2008, 27, 1–5. [Google Scholar]
- McNamara, D.J.; Kolb, R.; Parker, T.S.; Batwin, H.; Samuel, P.; Brown, C.D.; Ahrens, E.H., Jr. Heterogeneity of cholesterol homeostasis in man. Response to changes in dietary fat quality and cholesterol quantity. J. Clin. Invest. 1987, 79, 1729–1739. [Google Scholar] [CrossRef]
- Fernandez, M.L. Effects of eggs on plasma lipoproteins in healthy populations. Food Funct. 2010, 1, 156–160. [Google Scholar] [CrossRef]
- Ballesteros, M.N.; Cabrera, R.M.; Saucedo Mdel, S.; Fernandez, M.L. Dietary cholesterol does not increase biomarkers for chronic disease in a pediatric population from northern Mexico. Am. J. Clin. Nutr. 2004, 80, 855–861. [Google Scholar]
- Herron, K.L.; Vega-Lopez, S.; Conde, K.; Ramjiganesh, T.; Roy, S.; Shachter, N.S.; Fernandez, M.L. Pre-menopausal women, classified as hypo- or hyperresponders, do not alter their LDL/HDL ratio following a high dietary cholesterol challenge. J. Am. Coll. Nutr. 2002, 21, 250–258. [Google Scholar]
- Herron, K.L.; Vega-Lopez, S.; Conde, K.; Ramjiganesh, T.; Shachter, N.S.; Fernandez, M.L. Men classified as hypo- or hyperresponders to dietary cholesterol feeding exhibit differences in lipoprotein metabolism. J. Nutr. 2003, 133, 1036–1042. [Google Scholar]
- Greene, C.M.; Zern, T.L.; Wood, R.J.; Shrestha, S.; Aggarwal, D.; Sharman, M.J.; Volek, J.S.; Fernandez, M.L. Maintenance of the LDL cholesterol: HDL cholesterol ratio in an elderly population given a dietary cholesterol challenge. J. Nutr. 2005, 135, 2793–2798. [Google Scholar]
- Mutungi, G.; Ratliff, J.; Puglisi, M.; Torres-Gonzalez, M.; Vaishnav, U.; Leite, J.O.; Quann, E.; Volek, J.S.; Fernandez, M.L. Dietary cholesterol from eggs increases plasma HDL cholesterol in overweight men consuming a carbohydrate-restricted diet. J. Nutr. 2008, 138, 272–276. [Google Scholar]
- Harman, N.L.; Leeds, A.R.; Griffin, B.A. Increased dietary cholesterol does not increase plasma low density lipoprotein when accompanied by an energy-restricted diet and weight loss. Eur. J. Nutr. 2008, 47, 287–293. [Google Scholar] [CrossRef]
- Mayurasakorn, K.; Srisura, W.; Sitphahul, P.; Hongto, P.O. High-density lipoprotein cholesterol changes after continuous egg consumption in healthy adults. J. Med. Assoc. Thai. 2008, 91, 400–407. [Google Scholar]
- U.S. Department of AgricultureU.S. Department of Health and Human ServicesDietary Guidelines for Americans, 2010, 7th ed; U.S. Government Printing Office: Washington, DC, USA, 2010.
- Spence, J.D.; Jenkins, D.J.; Davignon, J. Dietary cholesterol and egg yolks: Not for patients at risk of vascular disease. Can. J. Cardiol. 2010, 26, e336–e339. [Google Scholar] [CrossRef]
- Blesso, C.N.; Andersen, C.J.; Barona, J.; Volk, B.; Volek, J.S.; Fernandez, M.L. A moderate carbohydrate-restricted diet results in weight loss and improves clinical parameters of metabolic syndrome in adult men and women. Addition of egg yolk further improves inflammation. FASEB J 2012, 26. (Suppl.), 819.30. [Google Scholar]
- Gardner, C.D.; Fortmann, S.P.; Krauss, R.M. Association of small low-density lipoprotein particles with the incidence of coronary artery disease in men and women. JAMA 1996, 276, 875–881. [Google Scholar] [CrossRef]
- Mensink, R.P.; Katan, M.B. Effect of dietary fatty acids on serum lipids and lipoproteins. A meta-analysis of 27 trials. Arterioscler. Thromb. 1992, 12, 911–919. [Google Scholar] [CrossRef]
- Krauss, R.M. Atherogenic lipoprotein phenotype and diet-gene interactions. J. Nutr. 2001, 131, 340–343. [Google Scholar]
- Yuan, Y.; Li, P.; Ye, J. Lipid homeostasis and the formation of macrophage-derived foam cells in atherosclerosis. Protein Cell 2010, 3, 173–181. [Google Scholar] [CrossRef]
- Herron, K.L.; Lofgren, I.E.; Sharman, M.; Volek, J.S.; Fernandez, M.L. High intake of cholesterol results in less atherogenic low-density lipoprotein particles in men and women independent of response classification. Metabolism 2004, 53, 823–830. [Google Scholar] [CrossRef]
- Zanni, E.E.; Zannis, V.I.; Blum, C.B.; Herbert, P.N.; Breslow, J.L. Effect of egg cholesterol and dietary fats on plasma lipids, lipoproteins, and apoproteins of normal women consuming natural diets. J. Lipid Res. 1987, 28, 518–527. [Google Scholar]
- Greene, C.M.; Waters, D.; Clark, R.M.; Contois, J.H.; Fernandez, M.L. Plasma LDL and HDL characteristics and carotenoid content are positively influenced by egg consumption in an elderly population. Nutr. Metab. (Lond.) 2006, 3, 6. [Google Scholar] [CrossRef]
- Mutungi, G.; Waters, D.; Ratliff, J.; Puglisi, M.; Clark, R.M.; Volek, J.S.; Fernandez, M.L. Eggs distinctly modulate plasma carotenoid and lipoprotein subclasses in adult men following a carbohydrate-restricted diet. J. Nutr. Biochem. 2010, 21, 261–267. [Google Scholar] [CrossRef]
- Ginsberg, H.; Le, N.A.; Mays, C.; Gibson, J.; Brown, W.V. Lipoprotein metabolism in nonresponders to increased dietary cholesterol. Arteriosclerosis 1981, 1, 463–470. [Google Scholar] [CrossRef]
- Fernandez, M.L.; Calle, M. Revisiting dietary cholesterol recommendations: Does the evidence support a limit of 300 mg/d? Curr. Atheroscler. Rep. 2010, 12, 377–383. [Google Scholar] [CrossRef]
- Beynen, A.C.; Katan, M.B.; Van Zutphen, L.F. Hypo- and hyperresponders: Individual differences in the response of serum cholesterol concentration to changes in diet. Adv. Lipid Res. 1987, 22, 115–171. [Google Scholar]
- Jeyarajah, E.J.; Cromwell, W.C.; Otvos, J.D. Lipoprotein particle analysis by nuclear magnetic resonance spectroscopy. Clin. Lab. Med. 2006, 26, 847–870. [Google Scholar] [CrossRef]
- Krauss, R.M. Lipoprotein subfractions and cardiovascular disease risk. Curr. Opin. Lipidol. 2010, 21, 305–311. [Google Scholar] [CrossRef]
- Mora, S.; Otvos, J.D.; Rifai, N.; Rosenson, R.S.; Buring, J.E.; Ridker, P.M. Lipoprotein particle profiles by nuclear magnetic resonance compared with standard lipids and apolipoproteins in predicting incident cardiovascular disease in women. Circulation 2009, 119, 931–939. [Google Scholar] [CrossRef]
- Soedamah-Muthu, S.S.; Chang, Y.F.; Otvos, J.; Evans, R.W.; Orchard, T.J. Lipoprotein subclass measurements by nuclear magnetic resonance spectroscopy improve the prediction of coronary artery disease in Type 1 diabetes. A prospective report from the Pittsburgh Epidemiology of Diabetes Complications Study. Diabetologia 2003, 46, 674–682. [Google Scholar]
- Clifton, P.M.; Kestin, M.; Abbey, M.; Drysdale, M.; Nestel, P.J. Relationship between sensitivity to dietary fat and dietary cholesterol. Arteriosclerosis 1990, 10, 394–401. [Google Scholar] [CrossRef]
- Nestel, P.J. Dietary cholesterol and plasma lipoproteins. Ann. N. Y. Acad. Sci. 1993, 676, 1–10. [Google Scholar] [CrossRef]
- Romano, G.; Tilly-Kiesi, M.K.; Patti, L.; Taskinen, M.R.; Pacioni, D.; Cassader, M.; Riccardi, G.; Rivellese, A.A. Effects of dietary cholesterol on plasma lipoproteins and their subclasses in iddm patients. Diabetologia 1998, 41, 193–200. [Google Scholar] [CrossRef]
- Kesaniemi, Y.A.; Ehnholm, C.; Miettinen, T.A. Intestinal cholesterol absorption efficiency in man is related to apoprotein E phenotype. J. Clin. Invest. 1987, 80, 578–581. [Google Scholar] [CrossRef]
- Clifton, P.M.; Nestel, P.J. Influence of gender, body mass index, and age on response of plasma lipids to dietary fat plus cholesterol. Arterioscler. Thromb. 1992, 12, 955–962. [Google Scholar] [CrossRef]
- Clifton, P.M.; Abbey, M.; Noakes, M.; Beltrame, S.; Rumbelow, N.; Nestel, P.J. Body fat distribution is a determinant of the high-density lipoprotein response to dietary fat and cholesterol in women. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 1070–1078. [Google Scholar] [CrossRef]
- Rader, D.J.; Alexander, E.T.; Weibel, G.L.; Billheimer, J.; Rothblat, G.H. The role of reverse cholesterol transport in animals and humans and relationship to atherosclerosis. J. Lipid Res. 2009, 50, S189–S194. [Google Scholar]
- Bruce, C.; Chouinard, R.A., Jr.; Tall, A.R. Plasma lipid transfer proteins, high-density lipoproteins, and reverse cholesterol transport. Ann. Rev. Nutr. 1998, 18, 297–330. [Google Scholar] [CrossRef]
- Ohashi, R.; Mu, H.; Wang, X.; Yao, Q.; Chen, C. Reverse cholesterol transport and cholesterol efflux in atherosclerosis. QJM 2005, 98, 845–856. [Google Scholar] [CrossRef]
- Sutherland, W.H.; Ball, M.J.; Walker, H. The effect of increased egg consumption on plasma cholesteryl ester transfer activity in healthy subjects. Eur. J. Clin. Nutr. 1997, 51, 172–176. [Google Scholar]
- Huesca-Gomez, C.; Carreon-Torres, E.; Nepomuceno-Mejia, T.; Sanchez-Solorio, M.; Galicia-Hidalgo, M.; Mejia, A.M.; Montano, L.F.; Franco, M.; Posadas-Romero, C.; Perez-Mendez, O. Contribution of cholesteryl ester transfer protein and lecithin: Cholesterol acyltransferase to HDL size distribution. Endocr. Res. 2004, 30, 403–415. [Google Scholar] [CrossRef]
- Leite, J.O.; Fernandez, M.L. Should we take high-density lipoprotein cholesterol levels at face value? Am. J. Cardiovasc. Drugs 2010, 10, 1–3. [Google Scholar]
- Tall, A.R. Plasma lipid transfer proteins. J. Lipid Res. 1986, 27, 361–367. [Google Scholar]
- Escola-Gil, J.C.; Llaverias, G.; Julve, J.; Jauhiainen, M.; Mendez-Gonzalez, J.; Blanco-Vaca, F. The cholesterol content of western diets plays a major role in the paradoxical increase in high-density lipoprotein cholesterol and upregulates the macrophage reverse cholesterol transport pathway. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2493–2499. [Google Scholar] [CrossRef]
- Andersen, C.J.; Blesso, C.N.; Park, Y.K.; Barona, J.; Pham, T.; Lee, J.Y.; Fernandez, M.L. Carbohydrate restriction favorably affects HDL metabolism in men and women with metabolic syndrome. Addition of egg yolk further increases HDL particles. FASEB J. 2012, 26. (Suppl.), 254.5.. [Google Scholar]
- Karadeniz, M.; Erdogan, M.; Ayhan, Z.; Yalcin, M.; Olukman, M.; Cetinkalp, S.; Alper, G.E.; Eroglu, Z.; Tetik, A.; Cetintas, V.; et al. Effect of G2706A and G1051A polymorphisms of the ABCA1 gene on the lipid, oxidative stress and homocystein levels in Turkish patients with polycystic ovary syndrome. Lipids Health Dis. 2011, 10, 193. [Google Scholar] [CrossRef]
- Katan, M.B.; Grundy, S.M.; Jones, P.; Law, M.; Miettinen, T.; Paoletti, R. Efficacy and safety of plant stanols and sterols in the management of blood cholesterol levels. Mayo. Clin. Proc. 2003, 78, 965–978. [Google Scholar]
- Fernandez, M.L. Soluble fiber and nondigestible carbohydrate effects on plasma lipids and cardiovascular risk. Cur. Opin. Lipidol. 2001, 12, 35–40. [Google Scholar] [CrossRef]
- Wang, L.J.; Song, B.L. Niemann-Pick C1-Like 1 and cholesterol uptake. Biochim. Biophys. Acta 1821, 964–972. [Google Scholar]
- Connor, W.E.; Lin, D.S. The intestinal absorption of dietary cholesterol by hypercholesterolemic (Type II) and normocholesterolemic humans. J. Clin. Invest. 1974, 53, 1062–1070. [Google Scholar] [CrossRef]
- Nestel, P.J.; Poyser, A. Changes in cholesterol synthesis and excretion when cholesterol intake is increased. Metabolism 1976, 25, 1591–1599. [Google Scholar] [CrossRef]
- Kern, F., Jr. Normal plasma cholesterol in an 88-year-old man who eats 25 eggs a day. Mechanisms of adaptation. N. Engl. J. Med. 1991, 324, 896–899. [Google Scholar] [CrossRef]
- Schmitz, G.; Langmann, T.; Heimerl, S. Role of ABCG1 and other ABCG family members in lipid metabolism. J. Lipid Res. 2001, 42, 1513–1520. [Google Scholar]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar]
- Herron, K.L.; McGrane, M.M.; Waters, D.; Lofgren, I.E.; Clark, R.M.; Ordovas, J.M.; Fernandez, M.L. The ABCG5 polymorphism contributes to individual responses to dietary cholesterol and carotenoids in eggs. J. Nutr. 2006, 136, 1161–1165. [Google Scholar]
- Tobert, J.A. New developments in lipid-lowering therapy: The role of inhibitors of hydroxymethylglutaryl-coenzyme a reductase. Circulation 1987, 76, 534–538. [Google Scholar] [CrossRef]
- Illingworth, D.R.; Tobert, J.A. A review of clinical trials comparing HMG-CoA reductase inhibitors. Clin. Ther. 1994, 16, 366–385. [Google Scholar]
- Vega, G.L.; Grundy, S.M. Effect of statins on metabolism of apo-B-containing lipoproteins in hypertriglyceridemic men. Am. J. Cardiol. 1998, 81, 36–42. [Google Scholar] [CrossRef]
- Mistry, P.; Miller, N.E.; Laker, M.; Hazzard, W.R.; Lewis, B. Individual variation in the effects of dietary cholesterol on plasma lipoproteins and cellular cholesterol homeostasis in man. Studies of low density lipoprotein receptor activity and 3-hydroxy-3-methylglutaryl coenzyme a reductase activity in blood mononuclear cells. J. Clin. Invest. 1981, 67, 493–502. [Google Scholar] [CrossRef]
- Torres-Gonzalez, M.; Shrestha, S.; Sharman, M.; Freake, H.C.; Volek, J.S.; Fernandez, M.L. Carbohydrate restriction alters hepatic cholesterol metabolism in guinea pigs fed a hypercholesterolemic diet. J. Nutr. 2007, 137, 2219–2223. [Google Scholar]
- Chambers, C.M.; Ness, G.C. Dietary cholesterol regulates hepatic 3-hydroxy-3-methylglutaryl coenzyme a reductase gene expression in rats primarily at the level of translation. Arch. Biol. Biophys. 1998, 354, 317–322. [Google Scholar] [CrossRef]
- Mutungi, G.; Torres-Gonzalez, M.; McGrane, M.M.; Volek, J.S.; Fernandez, M.L. Carbohydrate restriction and dietary cholesterol modulate the expression of HMG-CoA reductase and the LDL receptor in mononuclear cells from adult men. Lipids Health Dis. 2007, 6, 34. [Google Scholar] [CrossRef]
- Boucher, P.; de Lorgeril, M.; Salen, P.; Crozier, P.; Delaye, J.; Vallon, J.J.; Geyssant, A.; Dante, R. Effect of dietary cholesterol on low density lipoprotein-receptor, 3-hydroxy-3-methylglutaryl-CoAreductase, and low density lipoprotein receptor-related protein mrna expression in healthy humans. Lipids 1998, 33, 1177–1186. [Google Scholar] [CrossRef]
- Hu, F.B.; Stampfer, M.J.; Rimm, E.B.; Manson, J.E.; Ascherio, A.; Colditz, G.A.; Rosner, B.A.; Spiegelman, D.; Speizer, F.E.; Sacks, F.M.; et al. A prospective study of egg consumption and risk of cardiovascular disease in men and women. JAMA 1999, 281, 1387–1394. [Google Scholar]
- Song, W.O.; Kerver, J.M. Nutritional contribution of eggs to American diets. J. Am. Coll. Nutr. 2000, 19, 556–562. [Google Scholar]
- Esrey, K.L.; Joseph, L.; Grover, S.A. Relationship between dietary intake and coronary heart disease mortality: Lipid research clinics prevalence follow-up study. J. Clin. Epidemiol. 1996, 49, 211–216. [Google Scholar] [CrossRef]
- Krumholz, H.M.; Seeman, T.E.; Merrill, S.S.; Mendes de Leon, C.F.; Vaccarino, V.; Silverman, D.I.; Tsukahara, R.; Ostfeld, A.M.; Berkman, L.F. Lack of association between cholesterol and coronary heart disease mortality and morbidity and all-cause mortality in persons older than 70 years. JAMA 1994, 272, 1335–1340. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Barona, J.; Fernandez, M.L. Dietary Cholesterol Affects Plasma Lipid Levels, the Intravascular Processing of Lipoproteins and Reverse Cholesterol Transport without Increasing the Risk for Heart Disease. Nutrients 2012, 4, 1015-1025. https://doi.org/10.3390/nu4081015
Barona J, Fernandez ML. Dietary Cholesterol Affects Plasma Lipid Levels, the Intravascular Processing of Lipoproteins and Reverse Cholesterol Transport without Increasing the Risk for Heart Disease. Nutrients. 2012; 4(8):1015-1025. https://doi.org/10.3390/nu4081015
Chicago/Turabian StyleBarona, Jacqueline, and Maria Luz Fernandez. 2012. "Dietary Cholesterol Affects Plasma Lipid Levels, the Intravascular Processing of Lipoproteins and Reverse Cholesterol Transport without Increasing the Risk for Heart Disease" Nutrients 4, no. 8: 1015-1025. https://doi.org/10.3390/nu4081015