Metabolaid® Combination of Lemon Verbena and Hibiscus Flower Extract Prevents High-Fat Diet-Induced Obesity through AMP-Activated Protein Kinase Activation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Extraction and Extract Combination Preparation
2.2. HFD Mice and Administration of LV, HS, and MetA
2.3. Tissue Weight and Histological Analysis
2.4. Serum Assay of Biochemical Parameters
2.5. Rectal-Temperature Measurement in Cold Response
2.6. Intraperitoneal Glucose Tolerance Test (IGTT)
2.7. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.8. Western Blotting Analysis
2.9. High-Performance Liquid Chromatography (HPLC) Analysis of MetA
2.10. Statistical Analysis
3. Results
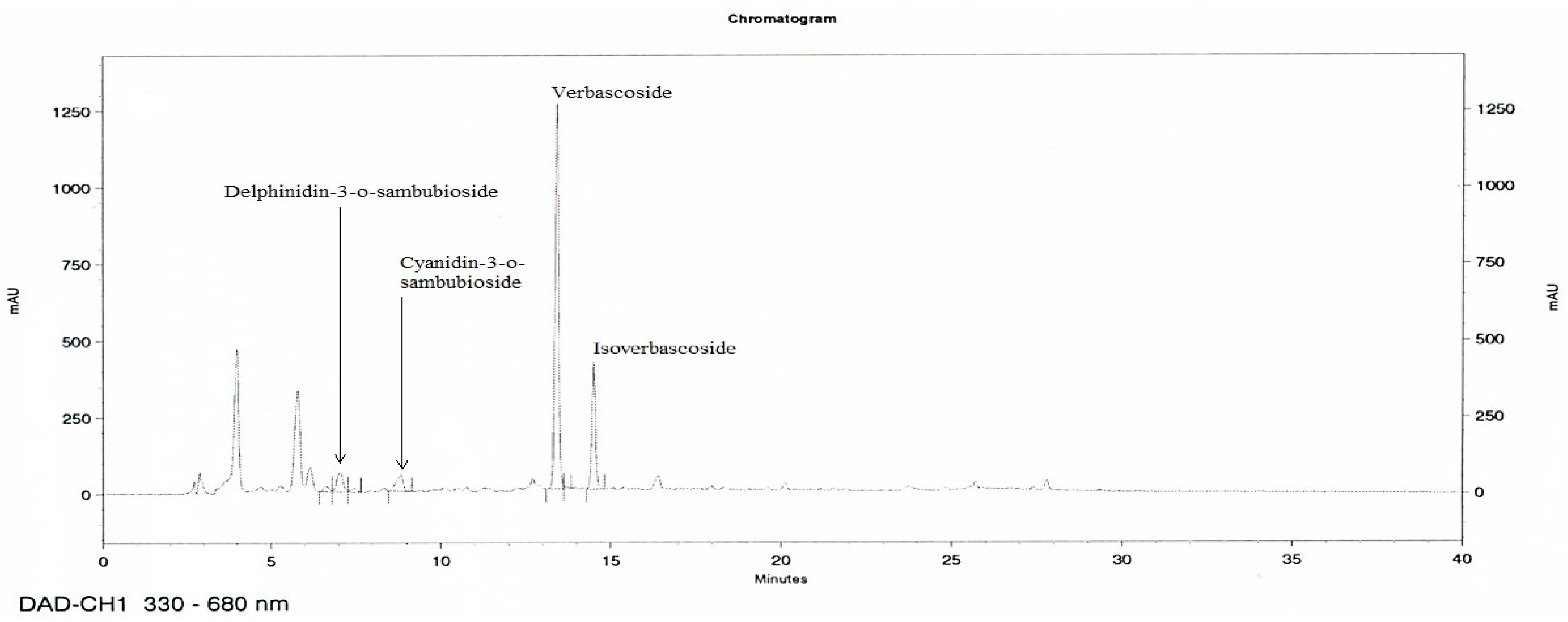
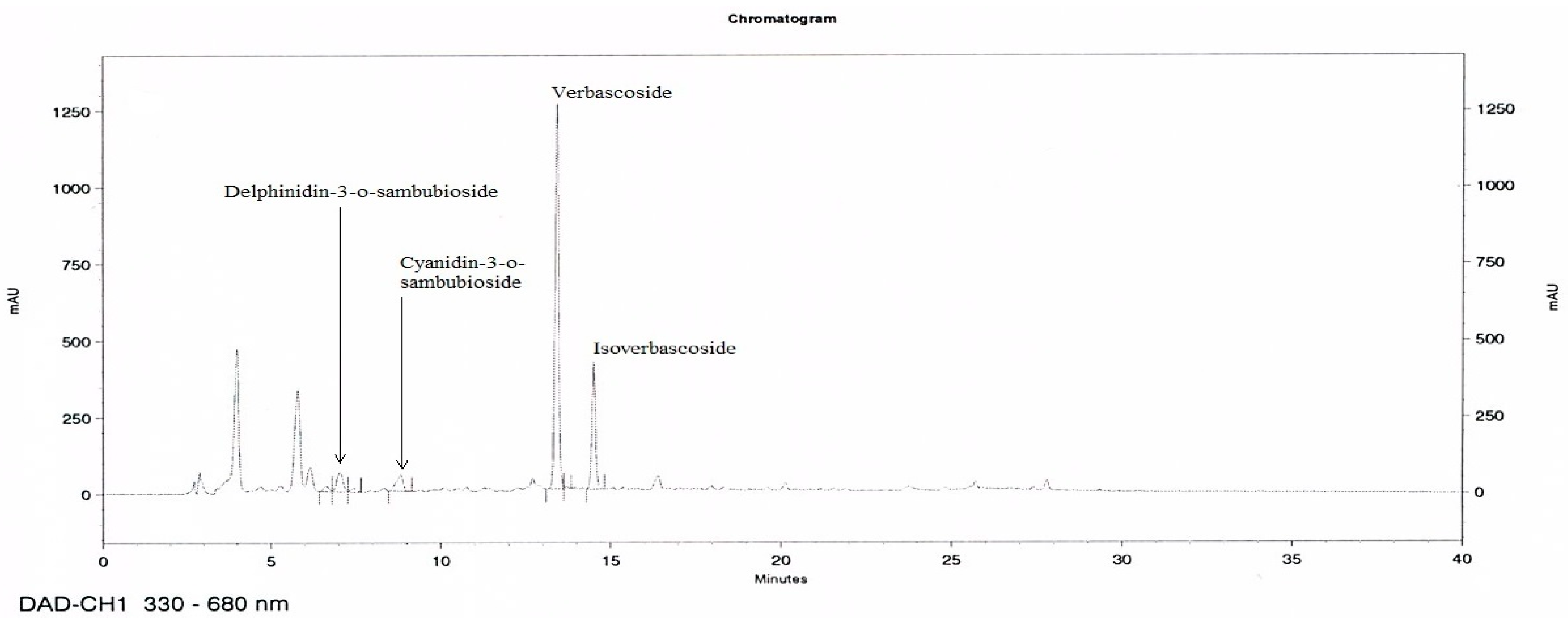
3.1. HPLC Analysis of MetA
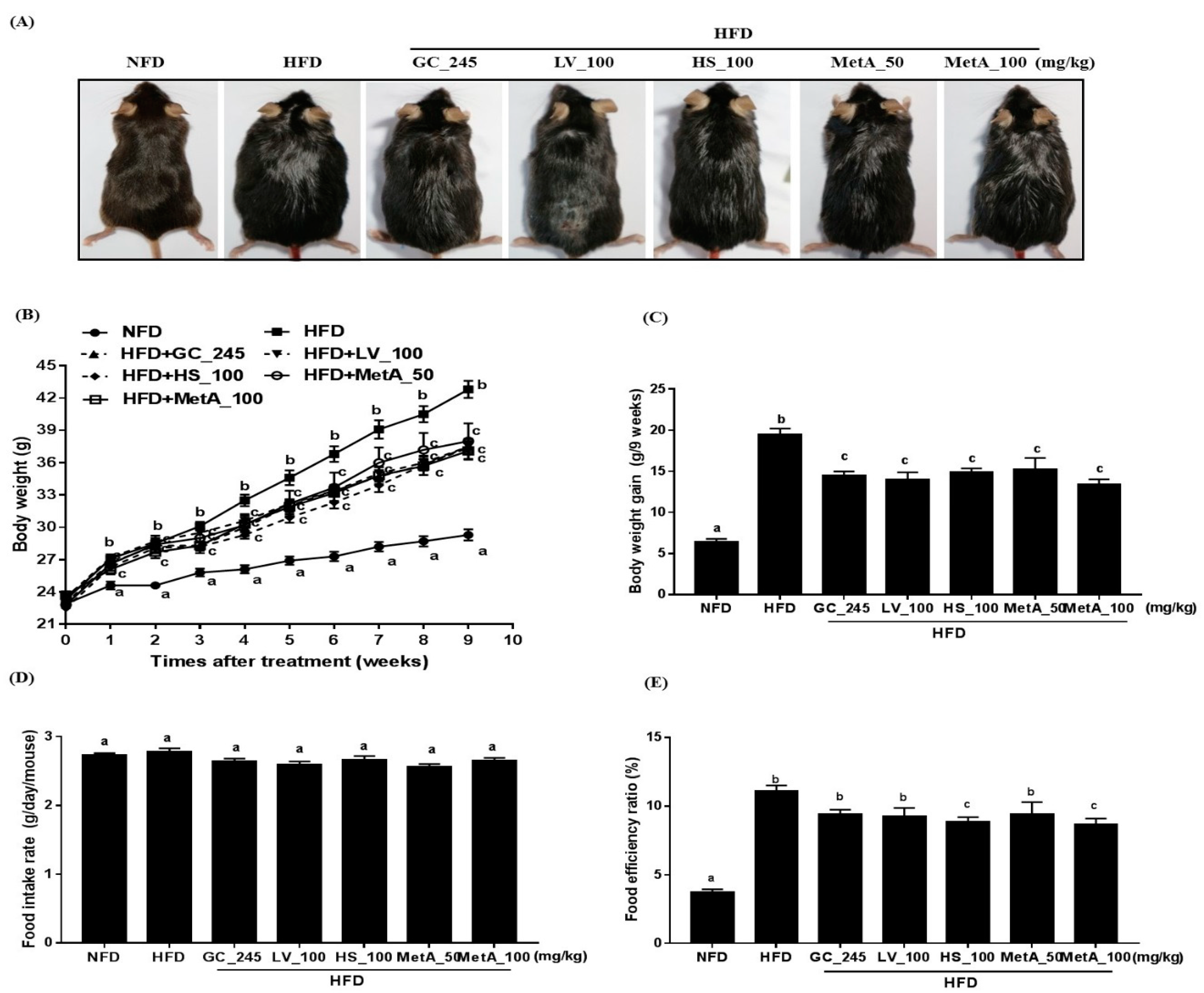
3.2. Effect of LV, HS, and the MetA on Body Weight and Food Intake of HFD-Induced Obese Mice
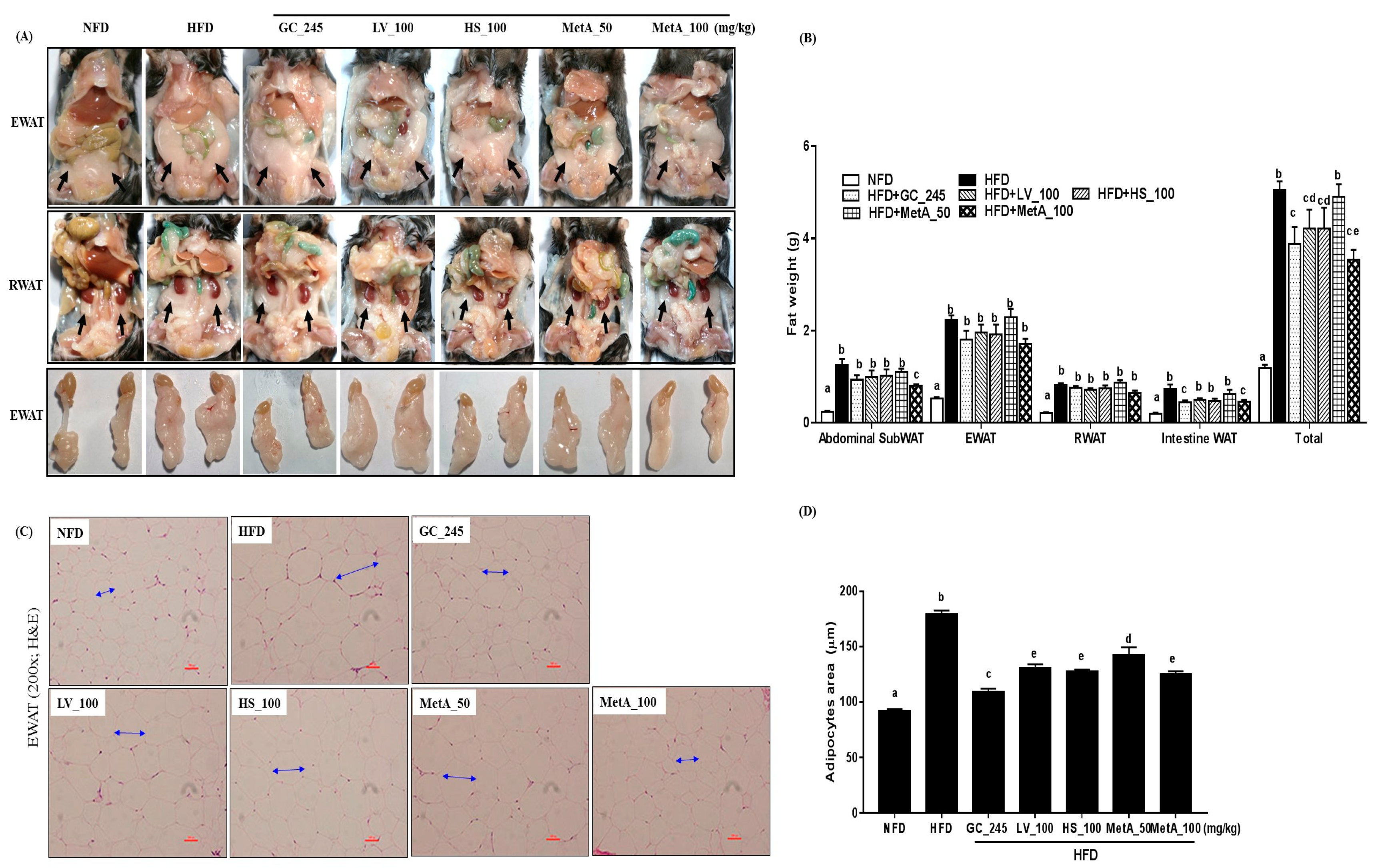
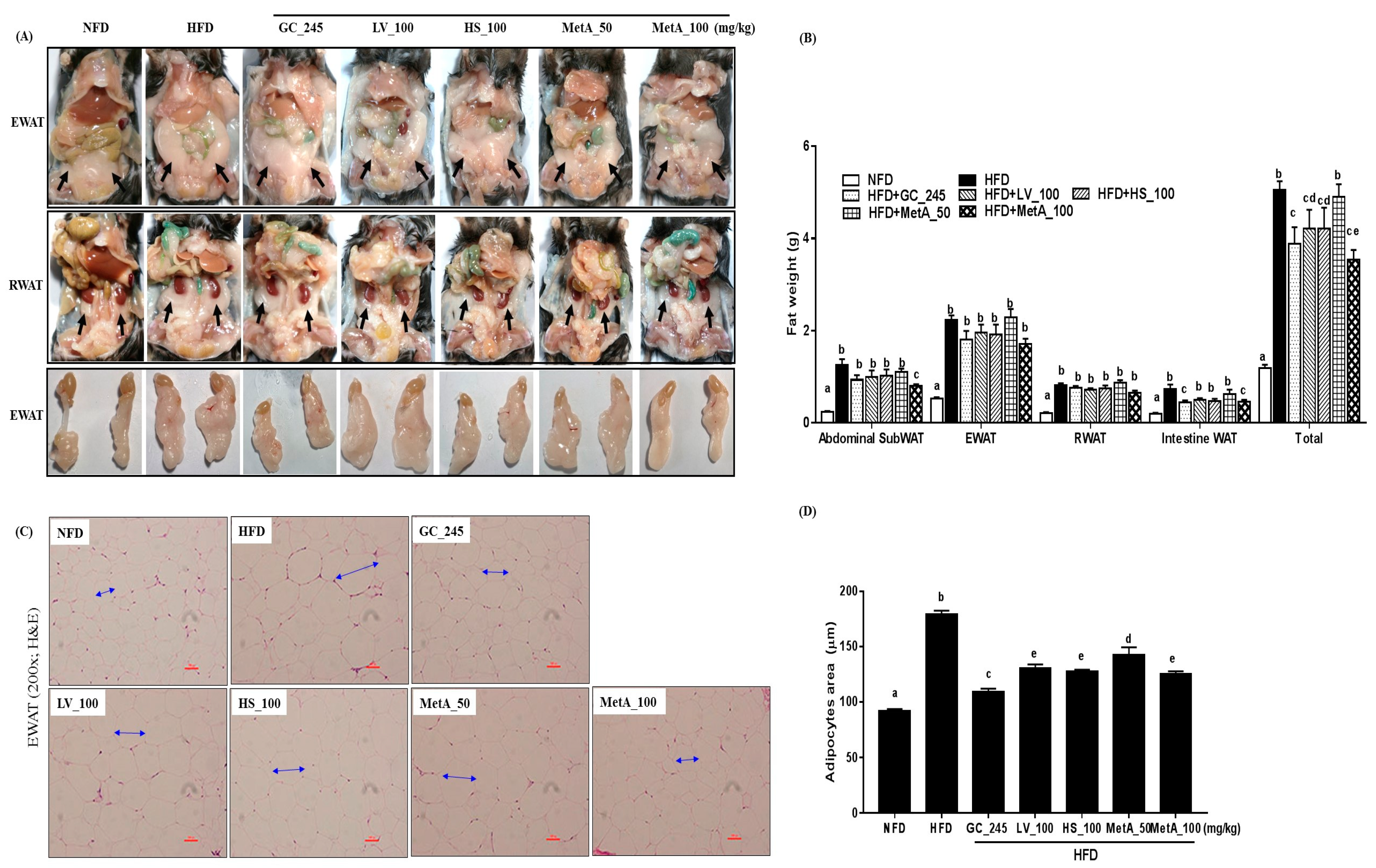
3.3. Effect of LV, HS, and the MetA on Adipose Tissue Weight and Morphology of HFD-Induced Obese Mice
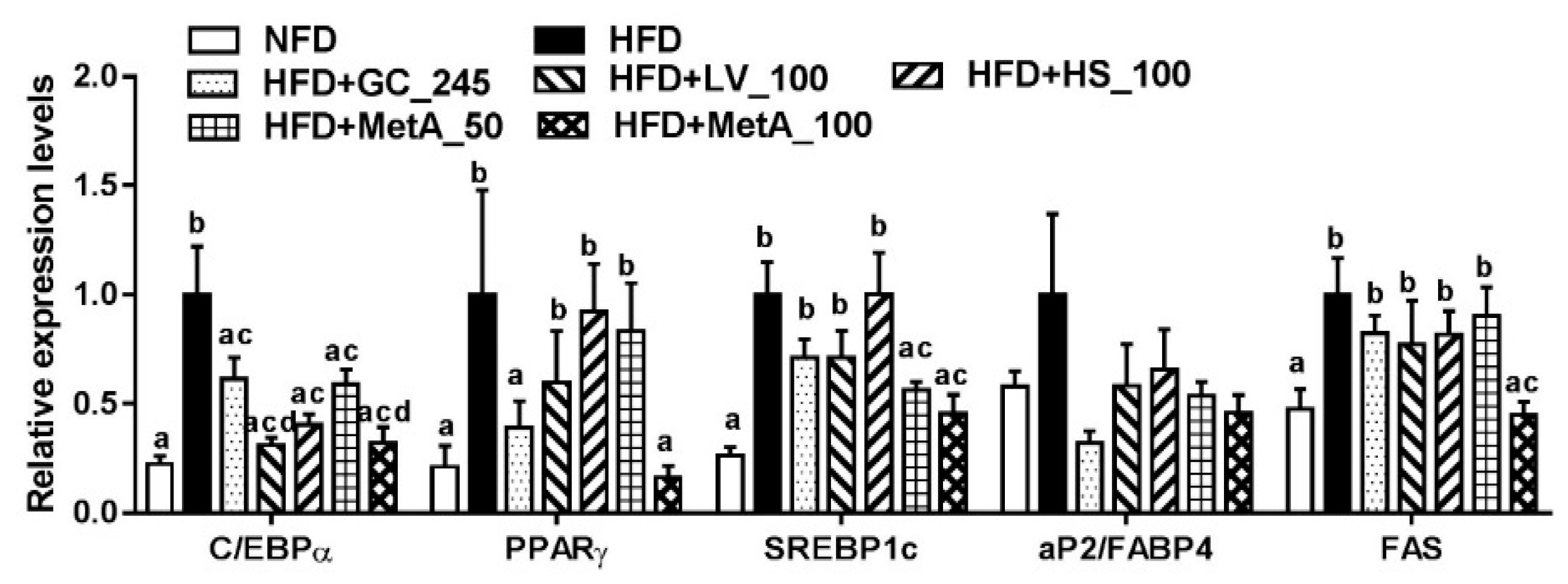
3.4. Effect of LV, HS, and MetA on Expression of Adipogenic Regulator Genes in HFD-Induced Obese Mice
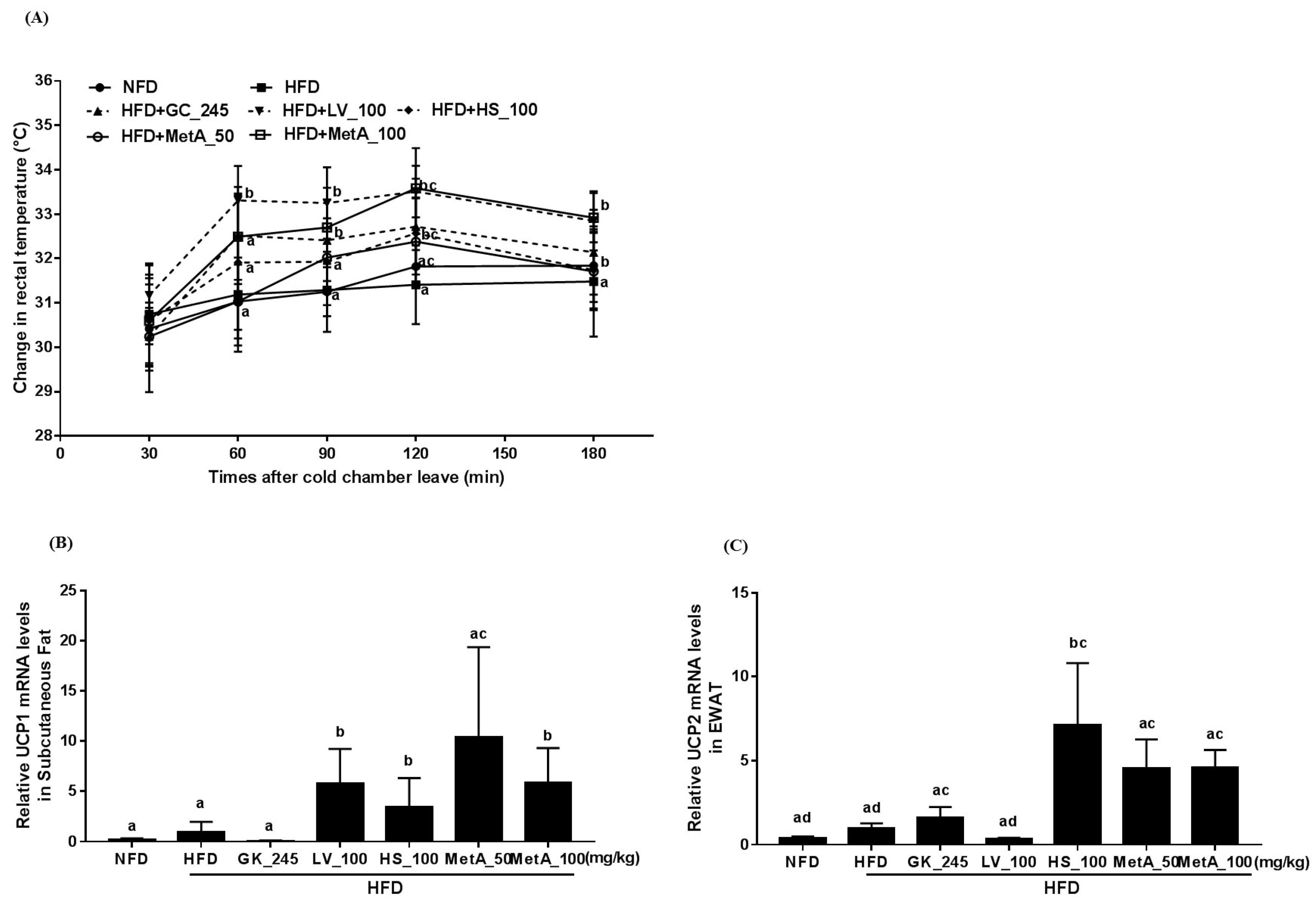
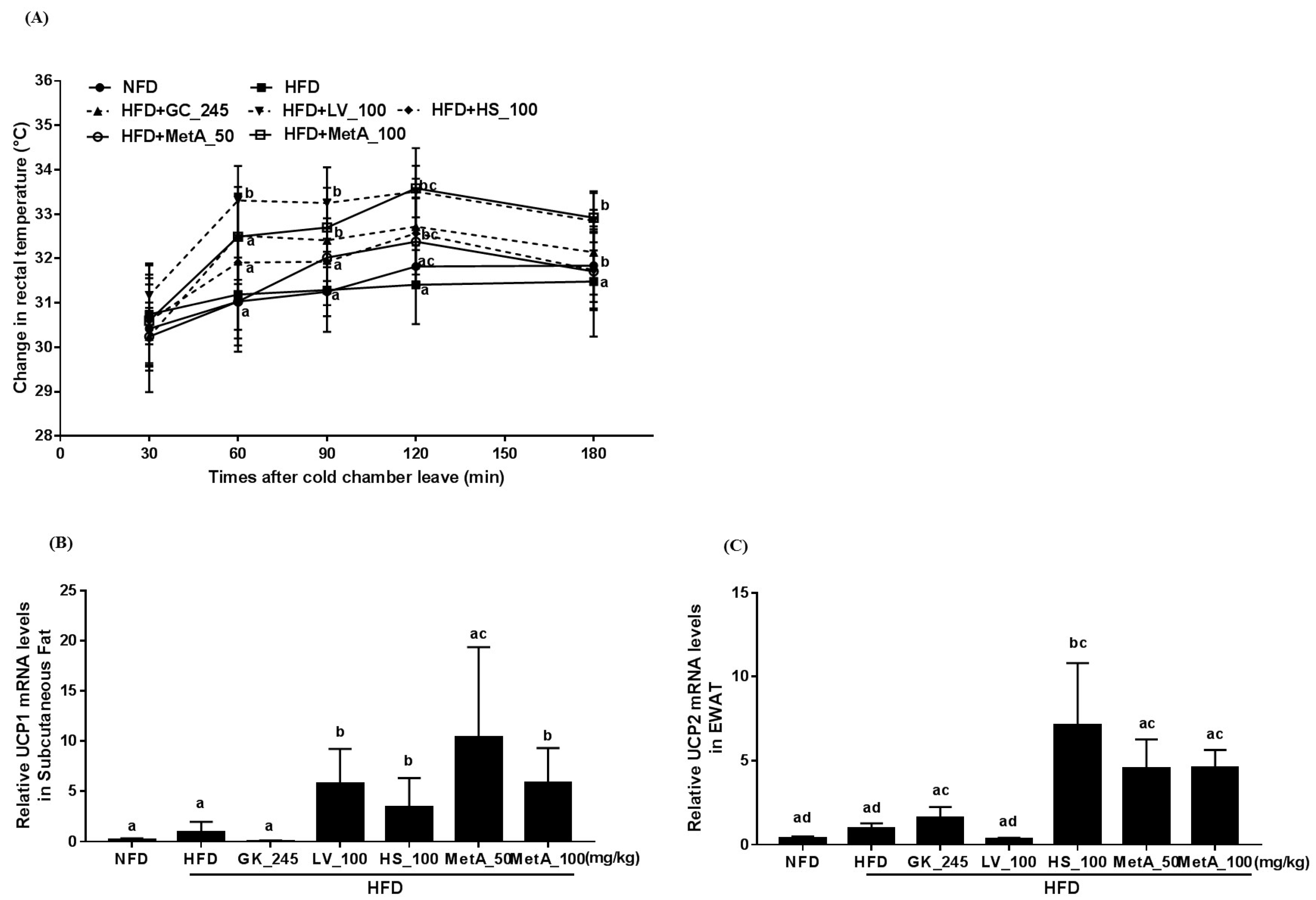
3.5. Effect of LV, HS, and MetA on Energy Expenditure and Thermogenesis-Related Gene Expression in HFD-Induced Obese Mice
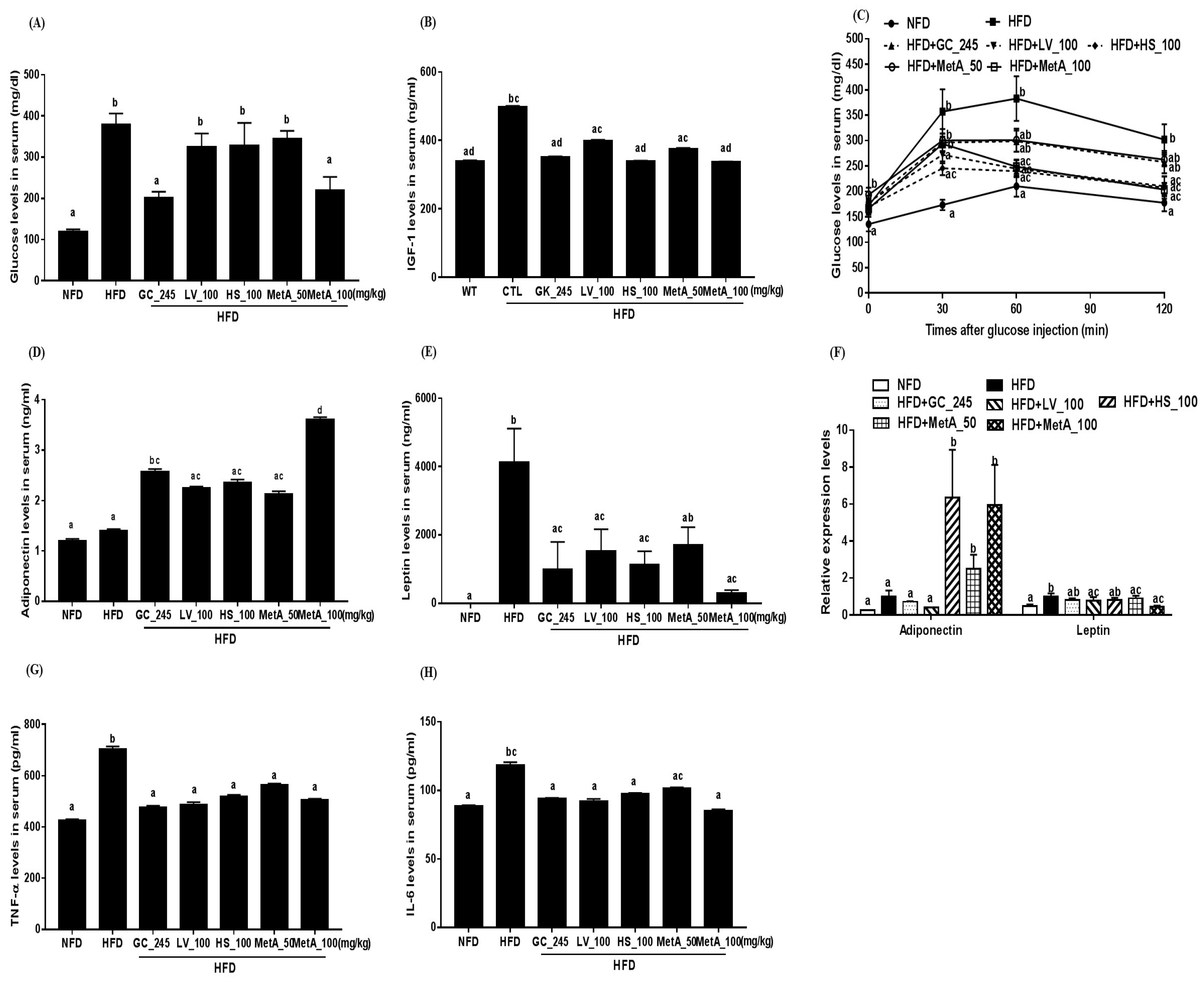
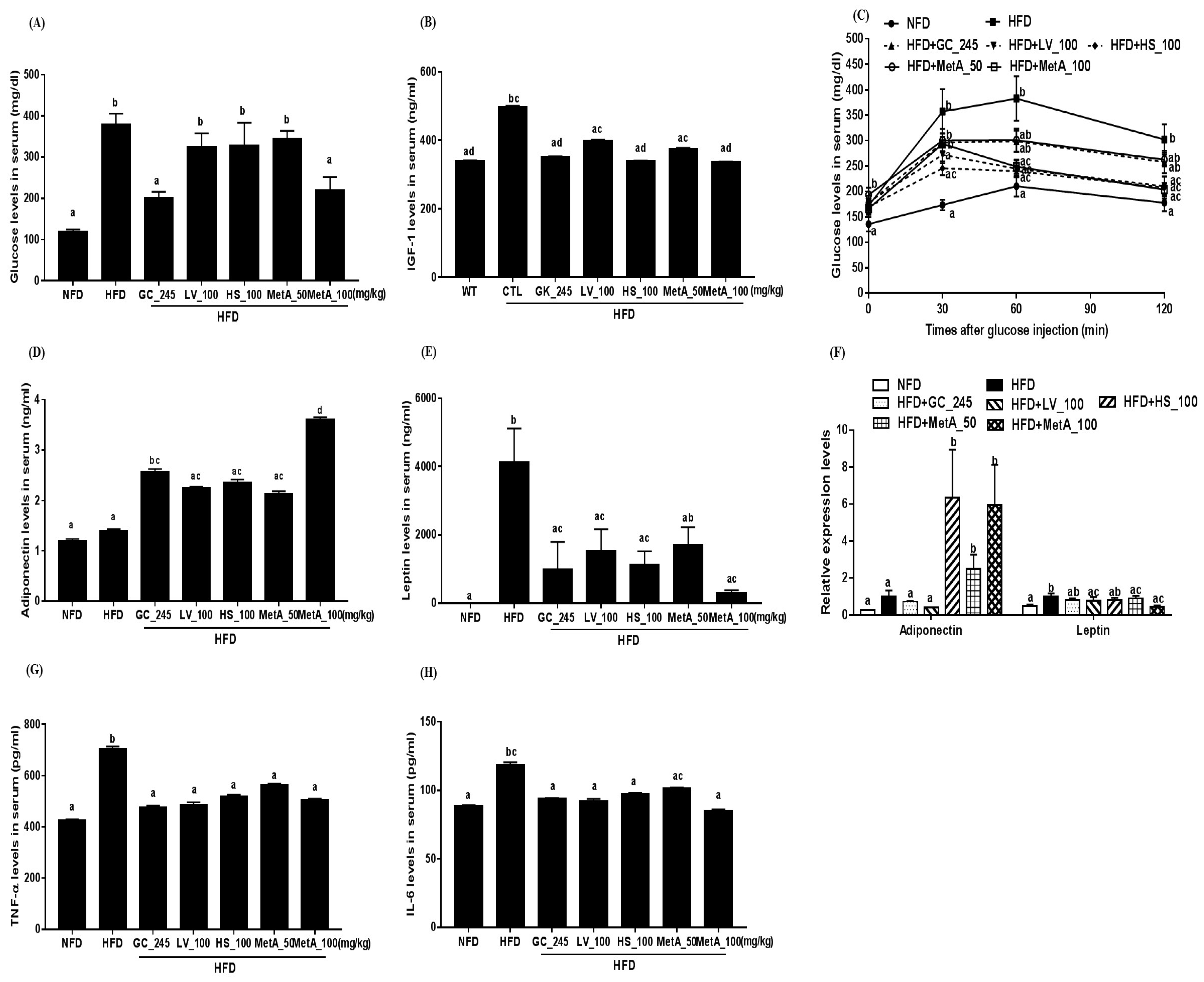
3.6. Effect of LV, HS, and MetA on Biochemical Parameters of HFD-Induced Obese Mice
3.7. Effect of LV, HS, and MetA on Systemic Glucose Tolerance in HFD-Induced Obese Mice
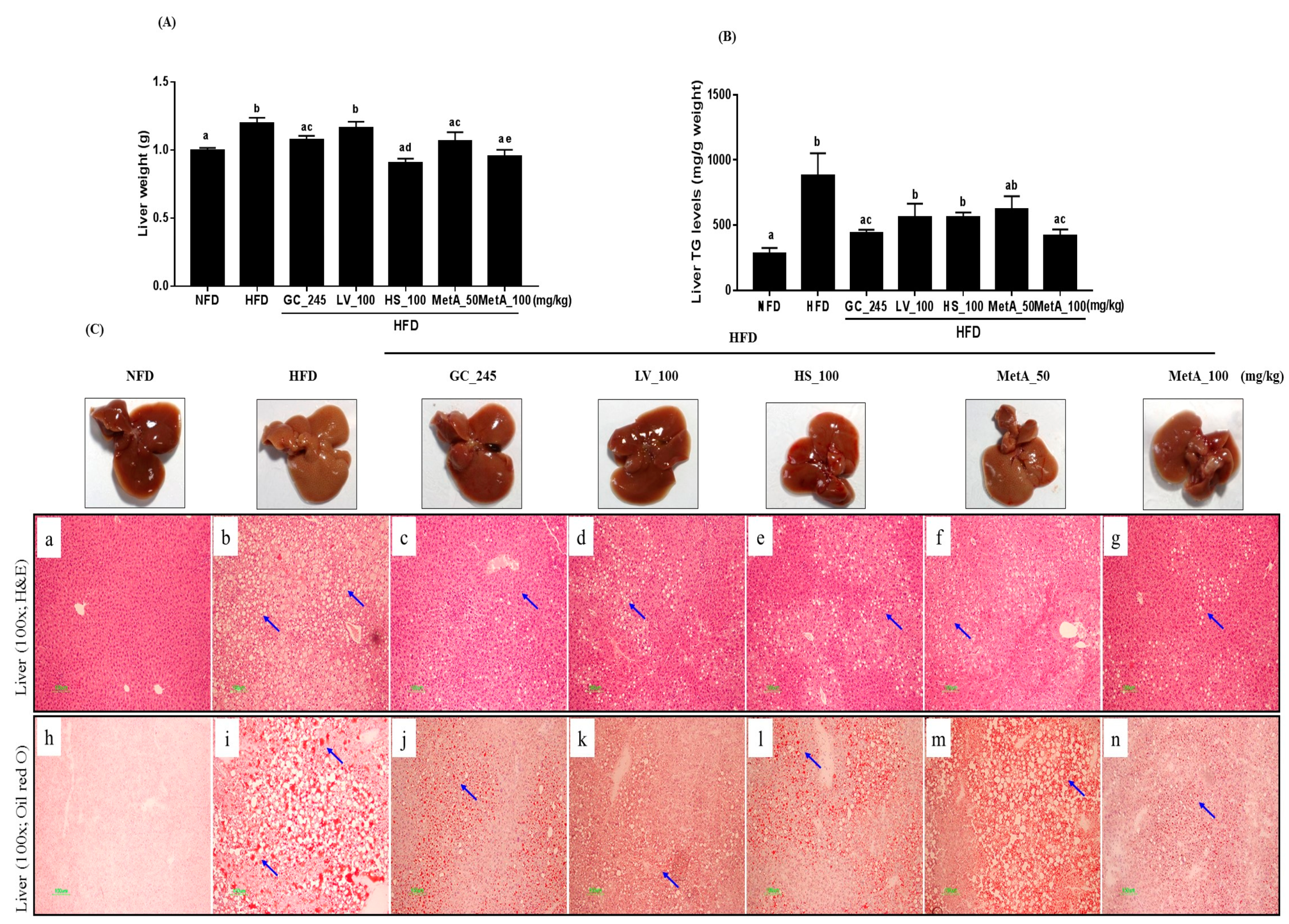
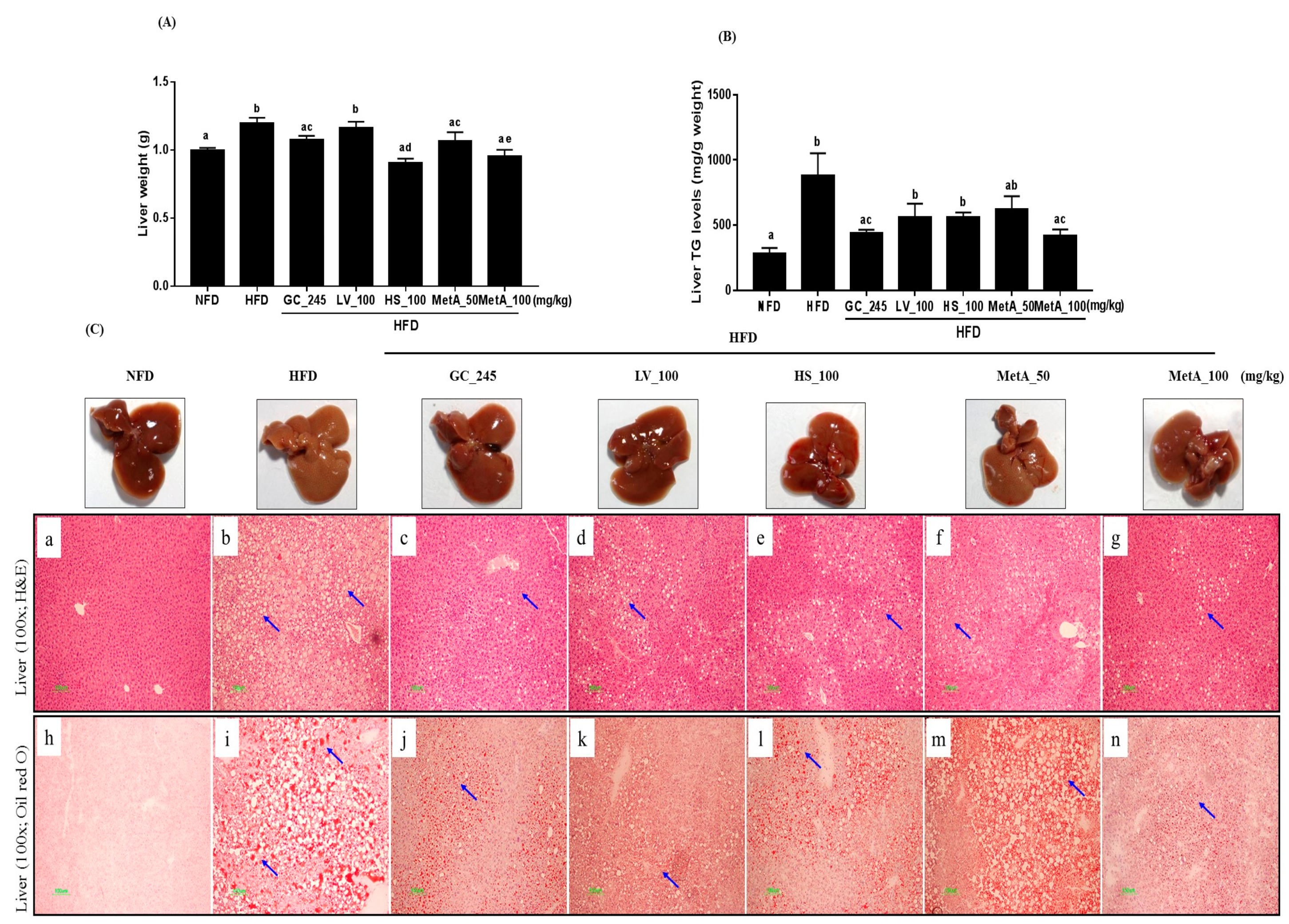
3.8. Effect of LV, HS, and the MetA on Liver Weight and Hepatic Lipid Accumulation
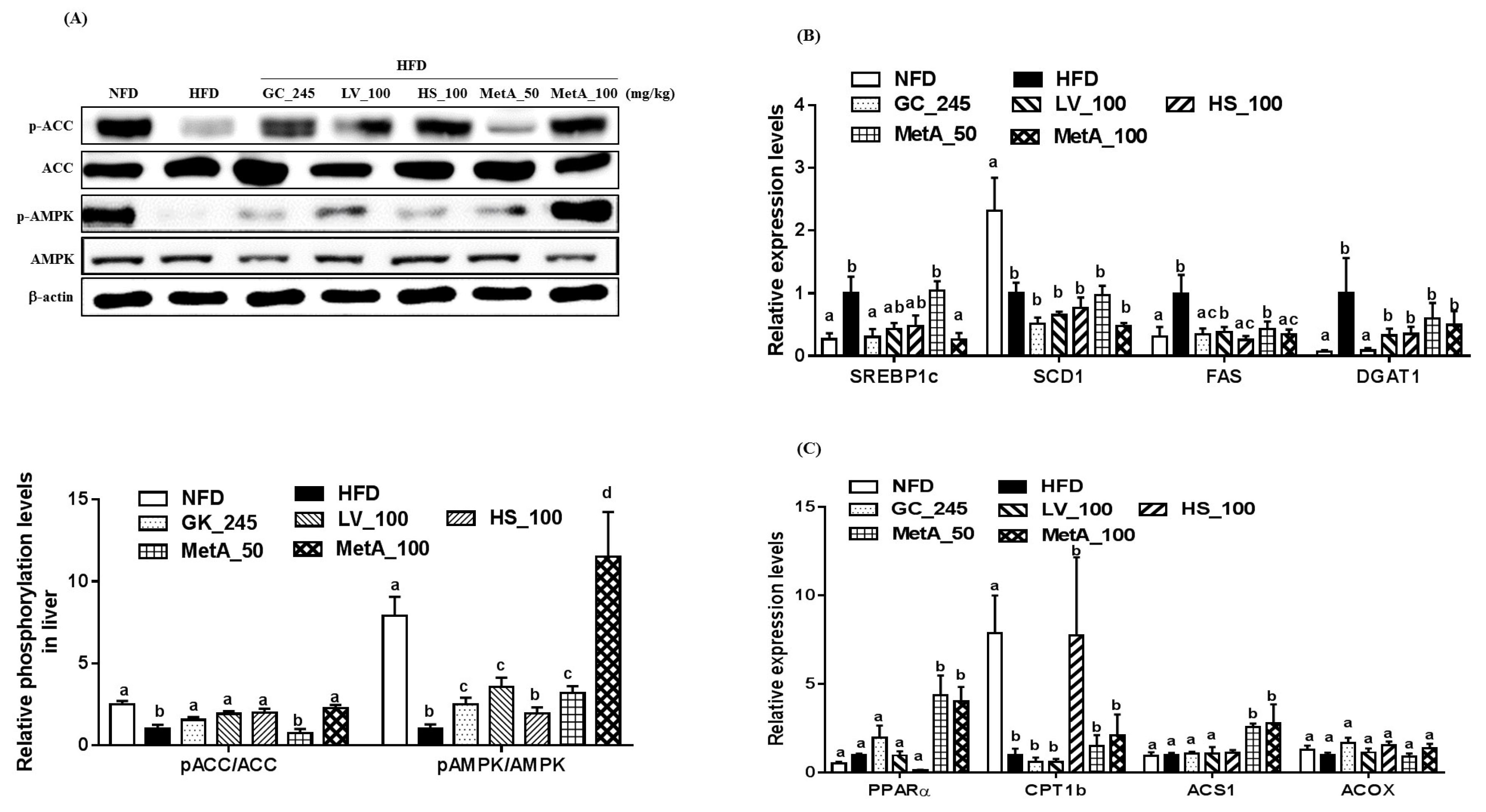
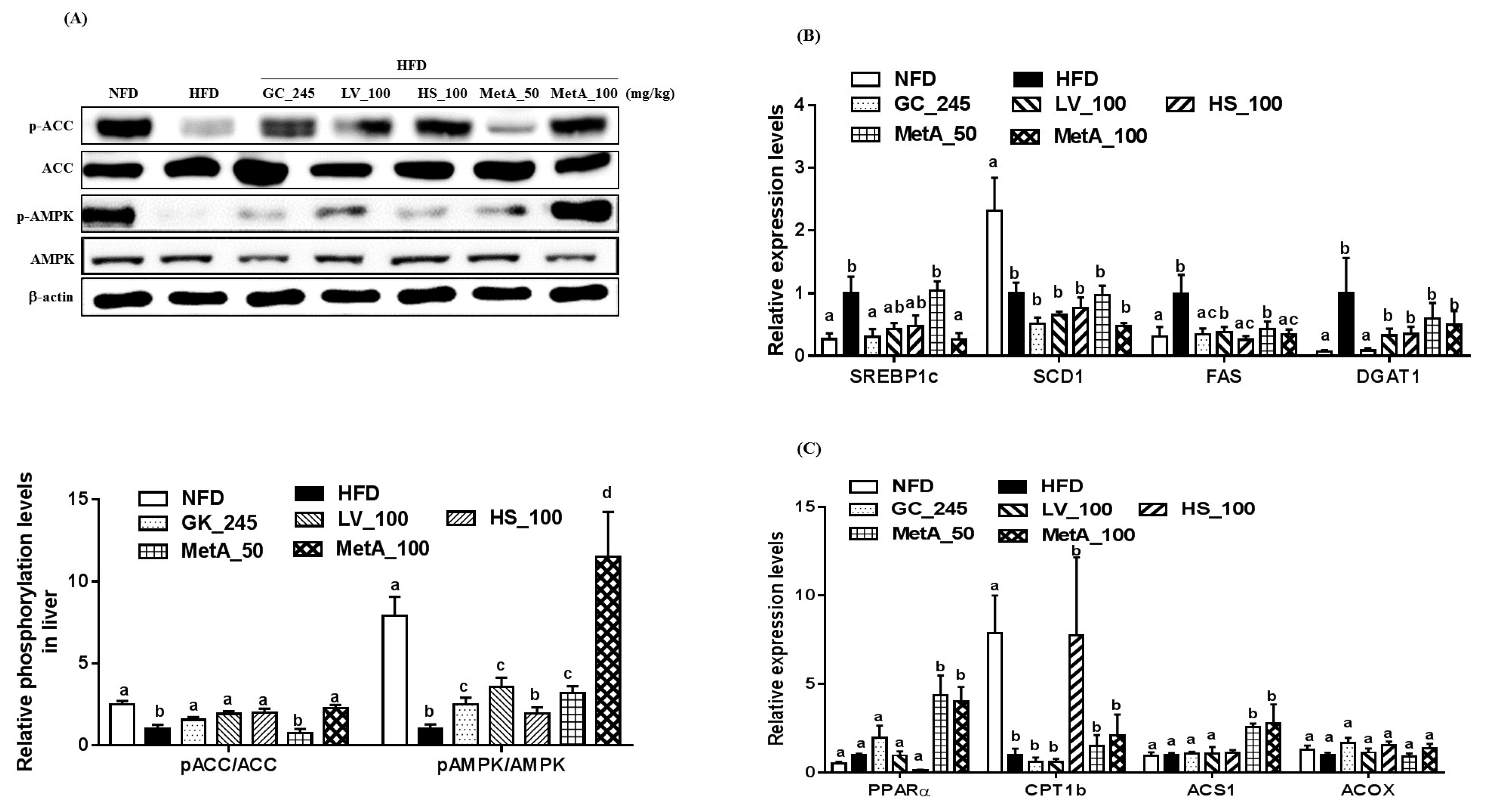
3.9. Effects of LV, HS, and the MetA on Lipid Metabolism-Related Genes and Protein Expression in the Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Spieglman, B.M.; Filer, J.S. Obesity and the regulation of energy balance. Cell 2001, 104, 531–543. [Google Scholar] [CrossRef]
- Lei, F.; Zhang, X.N.; Wang, W.; Xing, D.M.; Xie, W.D.; Su, H.; Du, L.J. Evidence of anti-obesity effects of the pomegranate leaf extract in high-fat diet induced obese mice. Int. J. Obes. (Lond.) 2007, 31, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daneschvar, H.L.; Aronson, M.D.; Smetana, G.W. FDA-approved anti-obesity drugs in the United States. Am. J. Med. 2016, 129, 879.e1–879.e6. [Google Scholar] [CrossRef] [PubMed]
- Heber, D. Herbal preparations for obesity: Are they useful? Prim. Care 2003, 30, 441–463. [Google Scholar] [CrossRef]
- Rios-Hoyo, A.; Gutierrez-Salmean, G. New dietary supplements for obesity: What we currently know. Curr. Obes. Rep. 2016, 5, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Newall, C.A.; Anderson, L.A.; Phillipson, J.D. Herbal Medicines: A Guide for Healthcare Professionals; Pharmaceutical Press: London, UK, 1996; p. 296. [Google Scholar]
- Jamila, F.; Mostafa, E. Ethnobotanical survey of medicinal plants used by people in oriental Morocco to manage various ailments. J. Ethnopharmacol. 2014, 154, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, L.; Rossary, A.; Joubert-Zakeyh, J.; Vergnaud-Gauduchon, J.; Farges, M.C.; Fraisse, D.; Texier, O.; Lamaison, J.-L.; Vasson, M.P.; Felgines, C. Lemon verbena infusion consumption attenuates oxidative stress in dextran sulfate sodium-induced colitis in the rat. Dig. Dis. Sci. 2011, 56, 3534–3545. [Google Scholar] [CrossRef] [PubMed]
- Herranz-López, M.; Barrajón-Catalán, E.; Segura-Carretero, A.; Menéndez, J.A.; Joven, J.; Micol, V. Lemon verbena (Lippia citriodora) polyphenols alleviate obesity-related disturbances in hypertrophic adipocytes through AMPK-dependent mechanisms. Phytomedicine 2015, 22, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Haji Faraji, M.; Haji Tarkhani, A. The effect of sour tea (Hibiscus sabdariffa) on essential hypertension. J. Ethnopharmacol. 1999, 65, 231–236. [Google Scholar] [CrossRef]
- Akindahunsi, A.A.; Olaleye, M.T. Toxicological investigation of aqueous methanolic extract of calyces of Hibiscus sabdariffa L. J. Ethnopharmacol. 2003, 89, 161–164. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, J.K.; Kim, H.J.; Moon, S.R.; Shin, B.C.; Park, K.W.; Yang, H.O.; Kim, S.M.; Park, R. Hibiscus extract inhibits the lipid droplet accumulation and adipogenic transcription factors expression of 3T3-L1 preadipocytes. J. Altern. Complement. Med. 2003, 9, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Hansawasdi, C.; Kawabata, J.; Kasai, T. Hibiscus acid as an inhibitor of starch digestion in the Caco-2 cell model system. Biosci. Biotechnol. Biochem. 2001, 65, 2087–2089. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, L.A.; Adebayo, J.O.; Oguntoye, O.B.; Olatunde, N.O.; Olatunji, V.A.; Soladote, A.O. Effects of aqueous extracts of petals of red and green Hibiscus sabdariffa. on plasma lipid and hematological variables in rats. Pharm. Biol. 2005, 43, 471–474. [Google Scholar] [CrossRef]
- Hirunpanich, V.; Utaipat, A.; Morales, N.P.; Bunyapraphatsara, N.; Sato, H.; Herunsale, A.; Suthisisang, C.H. Hypocholesterolemic and antioxidant effects of aqueous extracts from the dried calyx of Hibiscus sabdariffa L. in hypercholesterolemic rats. J. Ethnopharmacol. 2006, 103, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Joven, J.; Espinel, E.; Rull, A.; Aragonès, G.; Rodríguez-Gallego, E.; Camps, J.; Micol, V.; Herranz-López, M.; Menéndez, J.A.; Borrás, I.; et al. Plant-derived polyphenols regulate expression of miRNA paralogs miR-103/107 and miR-122 and prevent diet-induced fatty liver disease in hyperlipidemic mice. Biochim. Biophys. Acta 2012, 1820, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Herranz-López, M.; Olivares-Vicente, M.; Encinar, J.A.; Barrajón-Catalán, E.; Segura-Carretero, A.; Joven, J.; Micol, V. Multi-Targeted molecular effects of Hibiscus sabdariffa polyphenols: An opportunity for a global approach to obesity. Nutrients 2017, 9, 907. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Lee, W.K.; Lee, C.W.; Yoon, W.K.; Kim, N.; Park, S.K.; Lee, H.S.; Park, H.K.; Han, S.B.; Yun, J.; et al. Improvement of high-fat diet-induced obesity by a mixture of red grape extract, soy isoflavone and l-carnitine: Implications in cardiovascular and non-alcoholic fatty liver diseases. Food Chem. Toxicol. 2011, 49, 2453–2458. [Google Scholar] [CrossRef] [PubMed]
- Cristancho, A.G.; Lazar, M.A. Forming functional fat: A growing understanding of adipocyte differentiation. Nat. Rev. Mol. Cell Biol. 2011, 28, 722–734. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, J.; Cannon, B. The changed metabolic world with human brown adipose tissue: Therapeutic visions. Cell Metab. 2010, 11, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Townsend, K.L.; Tseng, Y.H. Brown fat fuel utilization and thermogenesis. Trends Endocrinol. Metab. 2014, 25, 168–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Bostrom, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Wang, Y.; Lu, S.F.; Hong, H.; Fu, S.; He, S.; Li, Q.; Yue, J.; Xu, B.; Zhu, B.M. Acupuncture promotes white adipose tissue browning by inducing UCP1 expression on DIO mice. BMC Complement. Altern. Med. 2014, 14, 501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Esteves, T.C. Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3. Cell Metab. 2005, 2, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Bracale, R.; Petroni, M.L.; Davinelli, S.; Bracale, U.; Scapagnini, G.; Carruba, M.O.; Nisoli, E. Muscle uncoupling protein 3 expression is unchanged by chronic ephedrine/caffeine treatment: Results of a double blind, randomised clinical trial in morbidly obese females. PLoS ONE 2014, 9, e98244. [Google Scholar] [CrossRef] [PubMed]
- Rashid, S.; Watanabe, T.; Sakaue, T.; Lewis, G.F. Mechanisms of HDL lowering in insulin resistant, hypertriglyceridemic states: The combined effect of HDL triglyceride enrichment and elevated hepatic lipase activity. Clin. Biochem. 2003, 36, 421–429. [Google Scholar] [CrossRef]
- Saadeh, S. Nonalcoholic fatty liver disease and obesity. Nutr. Clin. Pract. 2007, 22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.B.; Zhou, G.; Li, C. AMPK: An emerging drug target for diabetes and the metabolic syndrome. Cell Metab. 2009, 9, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, N.; Prentki, M. AMP kinase and malnoy-CoA: Target for therapy of the metabolic syndrome. Nat. Rev. Drug Discov. 2004, 3, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Paschke, R. Regulation of adipocytokines and insulin resistance. Diabetologia 2003, 46, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-α: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Lagathu, C.; Bastard, J.P.; Auclair, M.; Maachi, M.; Capeau, J.; Caron, M. Chronic interleukin-6 (IL-6) treatment increased IL-6 secretion and induced insulin resistance in adipocyte: Prevention by rosiglitazone. Biochem. Biophys. Res. Commun. 2003, 311, 372–379. [Google Scholar] [CrossRef]
- Fried, S.K.; Ricci, M.R.; Russell, C.D.; Laferrère, B. Regulation of leptin production in humans. J. Nutr. 2000, 130, 3127S–3131S. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S. Adipose tissue as an endocrine organ. Obesity 2006, 14, 242S–249S. [Google Scholar] [CrossRef] [PubMed]
- Funes, L.; Fernández-Arroyo, S.; Laporta, O.; Pons, A.; Roche, E.; Segura-Carretero, A.; Fernández-Gutiérrez, A.; Micol, V. Correlation between plasma antioxidant capacity and verbascoside levels in rats after oral administration of lemon verbena extract. Food Chem. 2009, 117, 589–598. [Google Scholar] [CrossRef]
- Herranz-Lopez, M.; Fernandez-Arroyo, S.; Perez-Sanchez, A.; Barrajon-Catalan, E.; Beltran-Debon, R.; Menendez, J.A.; Alonso-Villaverde, C.; Segura-Carretero, A.; Joven, J.; Micol, V. Synergism of plant-derived polyphenols in adipogenesis: Perspectives and implications. Phytomedicine 2012, 19, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Di Giancamillo, A.; Rossi, R.; Pastorelli, G.; Deponti, D.; Carollo, V.; Casamassima, D. The effects of dietary verbascoside on blood and liver oxidative stress status induced by a high-6 polyunsaturated fatty acids diet in piglets. J. Anim. Sci. 2015, 93, 2849–2859. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, P.; Villarreal, M.; Herrera-Ruiz, M.; Zamilpa, A.; Jiménez-Ferrer, E.; Trejo-Tapia, G. In vivo anti-inflammatory and anti-ulcerogenic activities of extracts from wild growing and in vitro plants of Castilleja tenuiflora Benth. (Orobanchaceae). J. Ethnopharmacol. 2013, 150, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Corbi, G.; Conti, V.; Komici, K.; Manzo, V.; Filippelli, A.; Palazzo, M.; Vizzari, F.; Davinelli, S.; Di Costanzo, A.; Scapagnini, G.; et al. Phenolic plant extracts induce sirt1 activity and increase antioxidant levels in the rabbit’s heart and liver. Oxid. Med. Cell Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Scapagnini, G.; Davinelli, S.; Kaneko, T.; Koverech, G.; Koverech, A.; Calabrese, E.J.; Calabrese, V. Dose response biology of resveratrol in obesity. J. Cell Commun. Signal. 2014, 8, 385–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felgines, C.; Fraisse, D.; Besson, C.; Vasson, M.P.; Texier, O. Bioavailability of lemon verbena (Aloysia triphylla) polyphenols in rats: Impact of colonic inflammation. Br. J. Nutr. 2014, 111, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Frank, T.; Janssen, M.; Netzel, M.; Strass, G.; Kler, A.; Kriesl, E.; Bitsch, I. Pharmacokinetics of anthocyanidin-3-glycosides following consumption of Hibiscus sabdariffa L. extract. J. Clin. Pharmacol. 2005, 45, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Boix-Castejón, M.; Herranz-López, M.; Pérez Gago, A.; Olivares-Vicente, M.; Caturla, N.; Roche, E.; Micol, V. Hibiscus and lemon verbena polyphenols modulate appetite-related biomarkers in overweight subjects: A randomized controlled trial. Food Funct. 2018, 9, 3173–3184. [Google Scholar] [CrossRef] [PubMed]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.—A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef] [PubMed]
- Bahramsoltani, R.; Rostamiasrabadi, P.; Shahpiri, Z.; Marques, A.M.; Rahimi, R.; Farzaei, M.H. Aloysia citrodora Paláu (Lemon verbena): A review of phytochemistry and pharmacology. J. Ethnopharmacol. 2018, 222, 34–51. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | GeneBank Accession NO | Sequences |
|---|---|---|
| C/EBPα | BC011118.1 | Forward:5′-TGGACAAGAACAGCAACGAGTAC-3′ Reverse: 5′-CGGTCATTGTCACTGGTCAACT-3′ |
| PPARγ | NM_011146 | FAM: 5′-TCGGAATCAGCTCTGTGGACCTCTCC-3′ |
| SREBP1c | BC056922.1 | Forward: 5′-AGCCTGGCCATCTGTGAGAA-3′ Reverse: 5′-CAGACTGGTACGGGCCACAA-3′; |
| aP2/FABP4 | NM_024406 | Forward: 5′-TGGGAACCTGGAAGCTTGTCTC-3′ Reverse: 5′-GAATTCCACGCCCAGTTTGA-3′ |
| FAS | NM_007988 | Forward: 5′-CTGAGATCCCAGCACTTCTTGA-3′ Reverse: 5′-GCCTCCGAAGCCAAATGAG-3′; |
| SCD1 | AH002082.2 | Forward: 5′-CATCGC CTGCTCTACCCTTT-3′ Reverse: 5′-GAACTG CGCTTGGAAACCTG-3′ |
| DGAT1 | NM_10046 | Forward: 5′-TGCTACGACGAGTTCTTGAG-3′ Reverse: 5′-CTCTGCCACAGCATTGAGAC-3′ |
| PPARα | BC016892 | Forward: 5′-GCCTGTCTGTCGGGATGT-3′ Reverse: 5′-GGCTTCGTGGATTCTCTTG-3′ |
| CTP1b | AF017174.2 | Forward: 5′-GTCGCTTCTTCAAGGTCTGG-3′ Reverse: 5′-AAGAAAGCAGCACGTTCGAT-3′ |
| ACS1 | U15977.1 | Forward: 5′-TCCTACAAAGAGGTGGCAGAACT-3′ Reverse: 5′-GGCTTGAACCCCTTCTGGAT-3′ |
| ACOX | BC056448 | Forward: 5′-CAGGAAGAGCAAGGAAGTGG-3′ Reverse: 5′-CCTTTCTGGCTGATCCCATA-3′ |
| UCP1 | NM_009463.3 | Forward: 5′-GACTCAGTCCAAGAGTACTTCTCTTC -3′ Reverse: 5′-GCCGGCTGAGATCTTGTTTC-3′ |
| UCP2 | NM_011671 | Forward: 5′-CCGCATTGGCCTCTACGACTCT-3′ Reverse: 5′-CCCCGAAGGCAGAAGTGAAGTG-3′ |
| Adiponectin | NM_009605 | Forward: 5′-GTCTCAGCTGTCGGTCTTCCCCT-3′ Reverse: 5′-CCCTGGCTTTATGCTCTTTGC-3′ |
| Leptin | NM_008493 | Forward: 5′-CCAAAACCCTCATCAAGACC-3′ Reverse: 5′-GTCCAACTGTTGAAGAATGTCCC-3′ |
| GAPDH | NM_008084 | VIC: 5′-TGCATCCTGCACCACCAACTGCTTAG-3′ |
| NFD | HFD | HFD | |||||
|---|---|---|---|---|---|---|---|
| GC_245 | LV_100 | HS_100 | MetA_50 | MetA_100 | |||
| TG (mg/dL) | 91.5 ± 4.1 a | 121.9 ± 10.5 | 106.6 ± 6.0 ab | 75.7 ± 4.5 ac | 100.4 ± 3.0 ab | 109.4 ± 8.0 ab | 105.4 ± 2.5 ab |
| NEFA (mEq/L) | 3.0 ± 0.1 a | 3.3 ± 0.2 a | 2.8 ± 0.1 a | 2.5 ± 0.1 b | 2.6 ± 0.1 b | 3.3 ± 0.17 a | 2.6 ± 0.1 b |
| T-CHO (mg/dL) | 118.4 ± 2.4 a | 317.1 ± 28.3 b | 186.8 ± 10.4 | 262.8 ± 36.2 b | 321.4 ± 34.4 b | 282.2 ± 15.7 b | 149.7 ± 6.3 |
| LDL-CHO (mg/dL) | 10.1 ± 0.5 a | 16.7 ± 1.5 b | 8.9 ± 0.8 ac | 13.2 ± 1.7 ab | 13.3 ± 1.4 ab | 11.3 ± 0.6 | 10.5 ± 0.6 ac |
| HDL-CHO (mg/dL) | 48.4 ± 1.1 a | 52.9 ± 2.2 a | 54.6 ± 2.6 a | 59.3 ± 3.4 a | 60.1 ± 2.6 b | 72.0 ± 3.6 c | 59.3 ± 2.5 a |
| ALT (mg/dL) | 20.7 ± 0.7 a | 61.6 ± 12.4 b | 24.8 ± 2.2 a | 21.4 ± 1.3 | 35.8 ± 5.7 a | 37.5 ± 8.3 a | 29.2 ± 4.1 a |
| AST (mg/dL) | 81.1 ± 8.6 | 118.0 ± 35.1 | 77.9 ± 9.6 | 91.4 ± 16.3 | 80.0 ± 7.1 | 82.1 ± 5.0 | 81.1 ± 5.8 |
| Creatinine (mg/dL) | 0.29 ± 0.01 | 0.34 ± 0.01 | 0.35 ± 0.01 | 0.33 ± 0.02 | 0.29 ± 0.01 | 0.34 ± 0.02 | 0.35 ± 0.01 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-S.; Yang, W.-K.; Kim, H.Y.; Min, B.; Caturla, N.; Jones, J.; Park, Y.-C.; Lee, Y.-C.; Kim, S.-H. Metabolaid® Combination of Lemon Verbena and Hibiscus Flower Extract Prevents High-Fat Diet-Induced Obesity through AMP-Activated Protein Kinase Activation. Nutrients 2018, 10, 1204. https://doi.org/10.3390/nu10091204
Lee Y-S, Yang W-K, Kim HY, Min B, Caturla N, Jones J, Park Y-C, Lee Y-C, Kim S-H. Metabolaid® Combination of Lemon Verbena and Hibiscus Flower Extract Prevents High-Fat Diet-Induced Obesity through AMP-Activated Protein Kinase Activation. Nutrients. 2018; 10(9):1204. https://doi.org/10.3390/nu10091204
Chicago/Turabian StyleLee, Young-Sil, Won-Kyung Yang, Hwa Yeon Kim, Bokkee Min, Nuria Caturla, Jonathan Jones, Yang-Chun Park, Young-Cheol Lee, and Seung-Hyung Kim. 2018. "Metabolaid® Combination of Lemon Verbena and Hibiscus Flower Extract Prevents High-Fat Diet-Induced Obesity through AMP-Activated Protein Kinase Activation" Nutrients 10, no. 9: 1204. https://doi.org/10.3390/nu10091204