Mapping Bush Encroaching Species by Seasonal Differences in Hyperspectral Imagery

Abstract
:
1. Introduction
2. Material and Methods
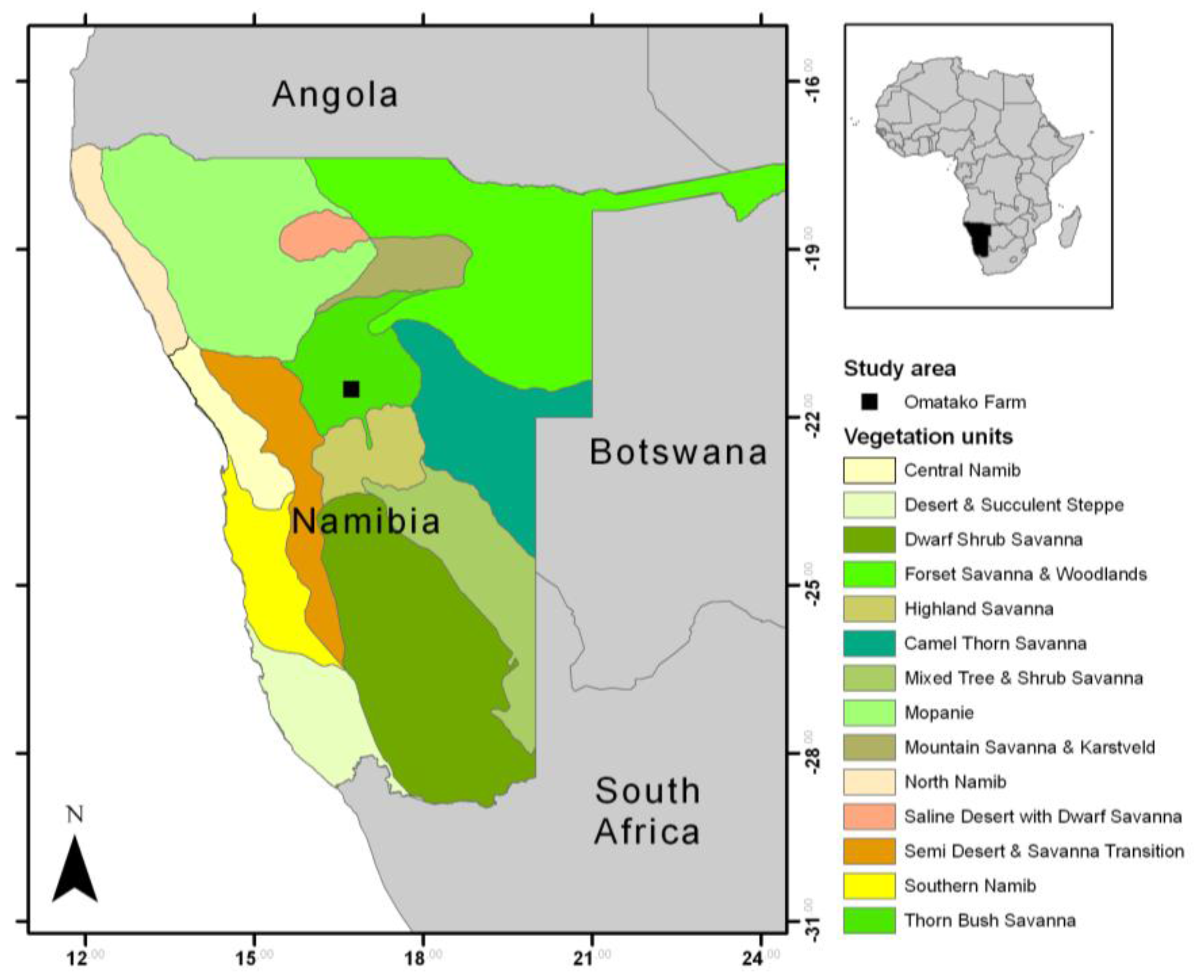
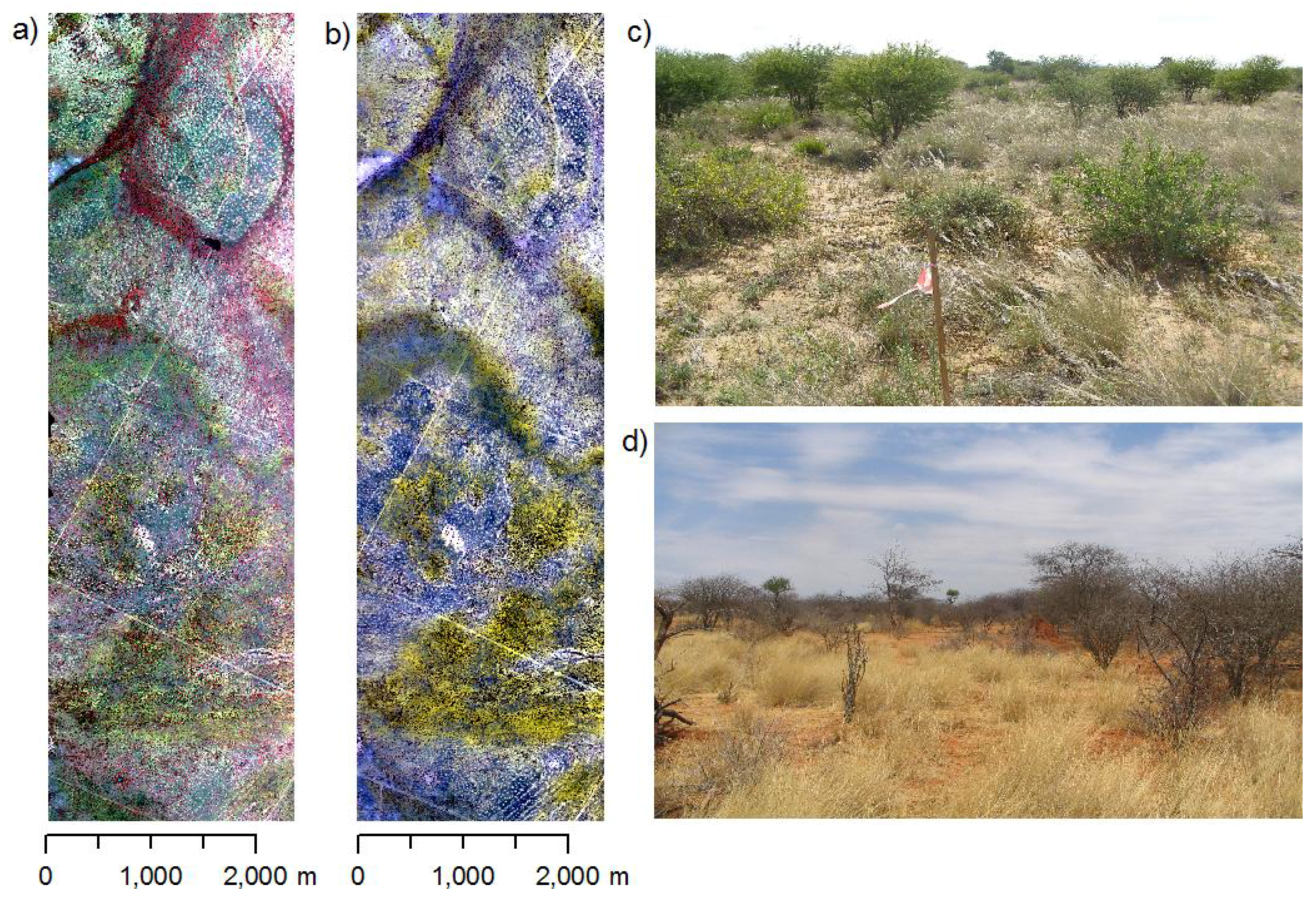
2.1. Study Region

2.2. Vegetation Sampling
2.3. Image Acquisition

2.4. Vegetation Index Differencing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nr. | Index | Full name | Feature | Reference |
|---|---|---|---|---|
| 1 | CARI | Chlorophyll Absorption in Reflectance Index | Chlorophyll | [40] |
| 2 | DGVI | Derivative Green Vegetation Index (1st order) | Greenness | [41] |
| 3 | LWVI | Leaf Water Vegetation Index | Water | [42] |
| 4 | NDLI | Normalized Difference Lignin Index | Lignin | [43] |
| 5 | NDNI | Normalized Difference Nitrogen Index | Nitrogen | [43] |
| 6 | CAI | Cellulose Absorption Index | Cellulose | [44] |
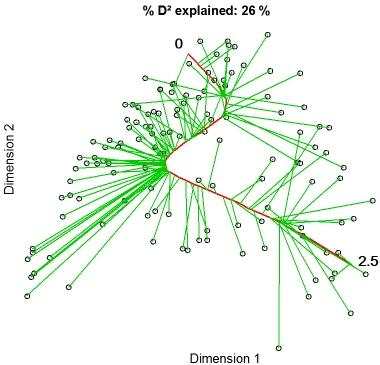
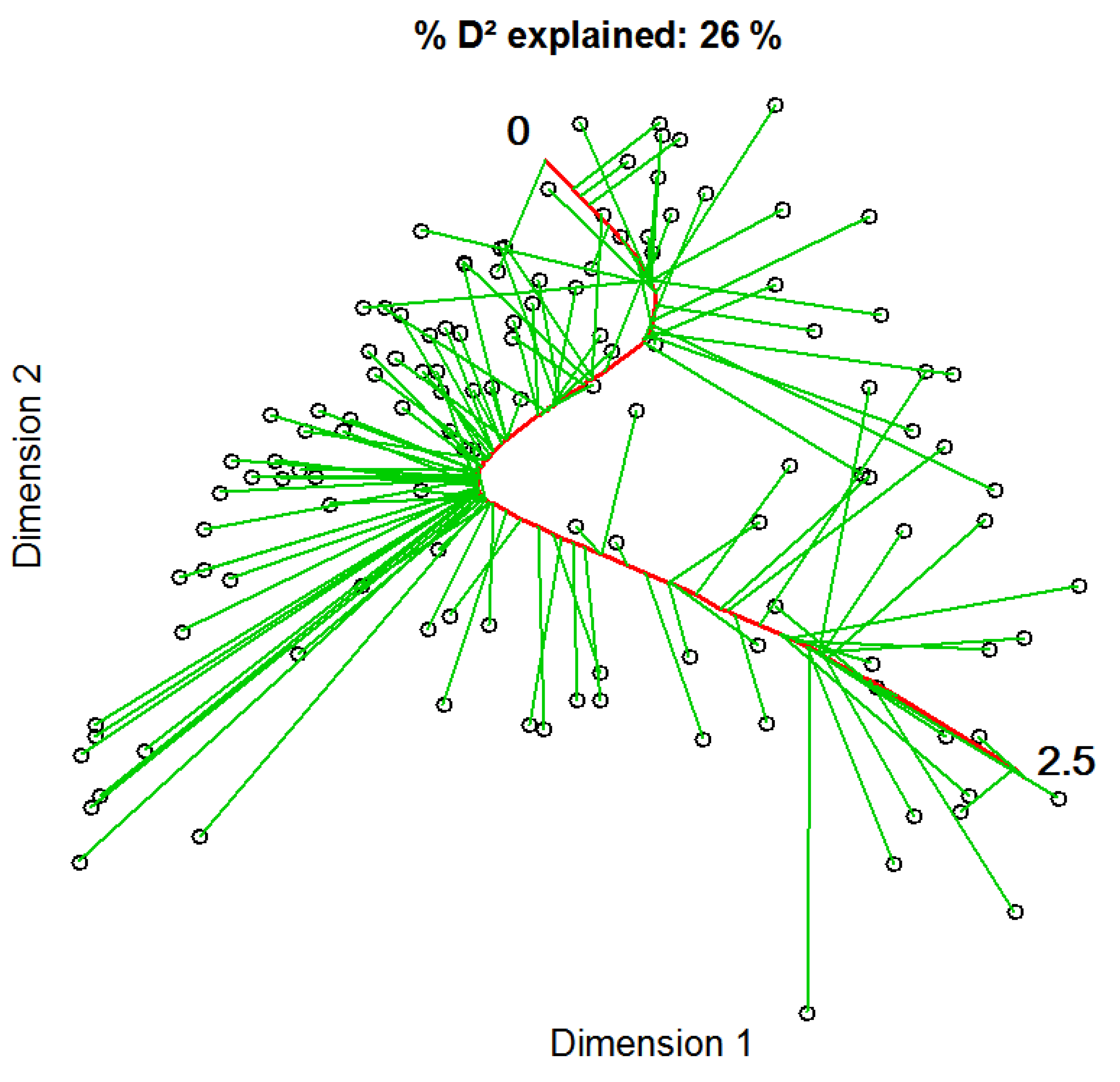
2.5. Constrained Principal Curves
2.6. Mapping and Validation of Plant Species Cover
3. Results
3.1. Constrained Principal Curves

| slope | Std. Error | t value | p | |
|---|---|---|---|---|
| intercept | 2.6646 | 0.3826 | 6.9647 | <0.001 |
| Δ CARI | −3.7014 | 0.6685 | −5.5365 | <0.001 |
| Δ LWVI | 38.5399 | 9.4153 | 4.0933 | <0.001 |
| Δ CAI | 10.3821 | 4.5146 | 2.2997 | <0.05 |
| Δ NDLI | −18.3285 | 13.1827 | −1.3903 | <0.1 |
| Δ NDNI | −32.9389 | 5.589 | −5.8935 | <0.001 |
| Δ DGVI | 14.3914 | 3.4836 | 4.1312 | <0.001 |
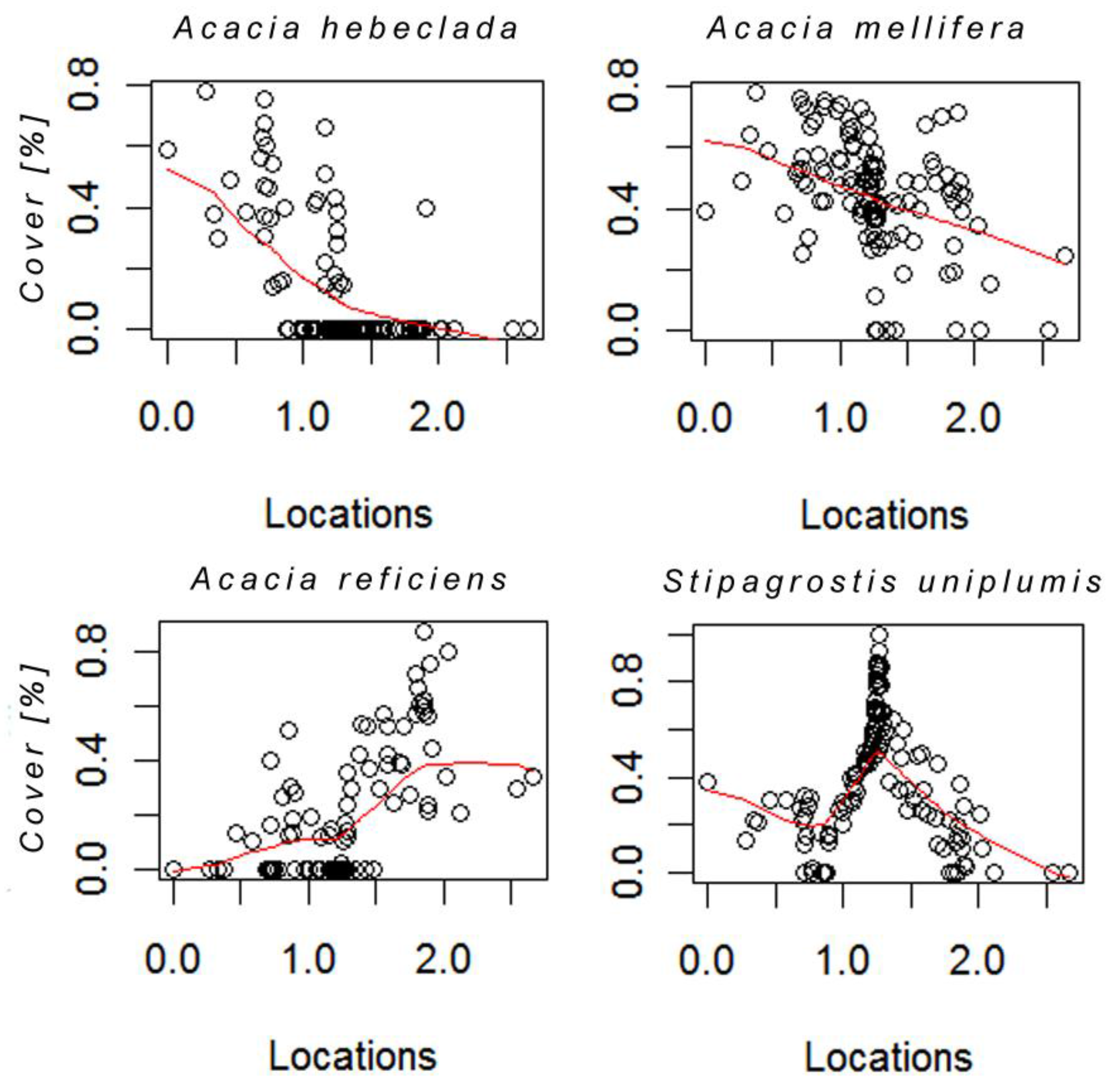
3.2. Species Responses

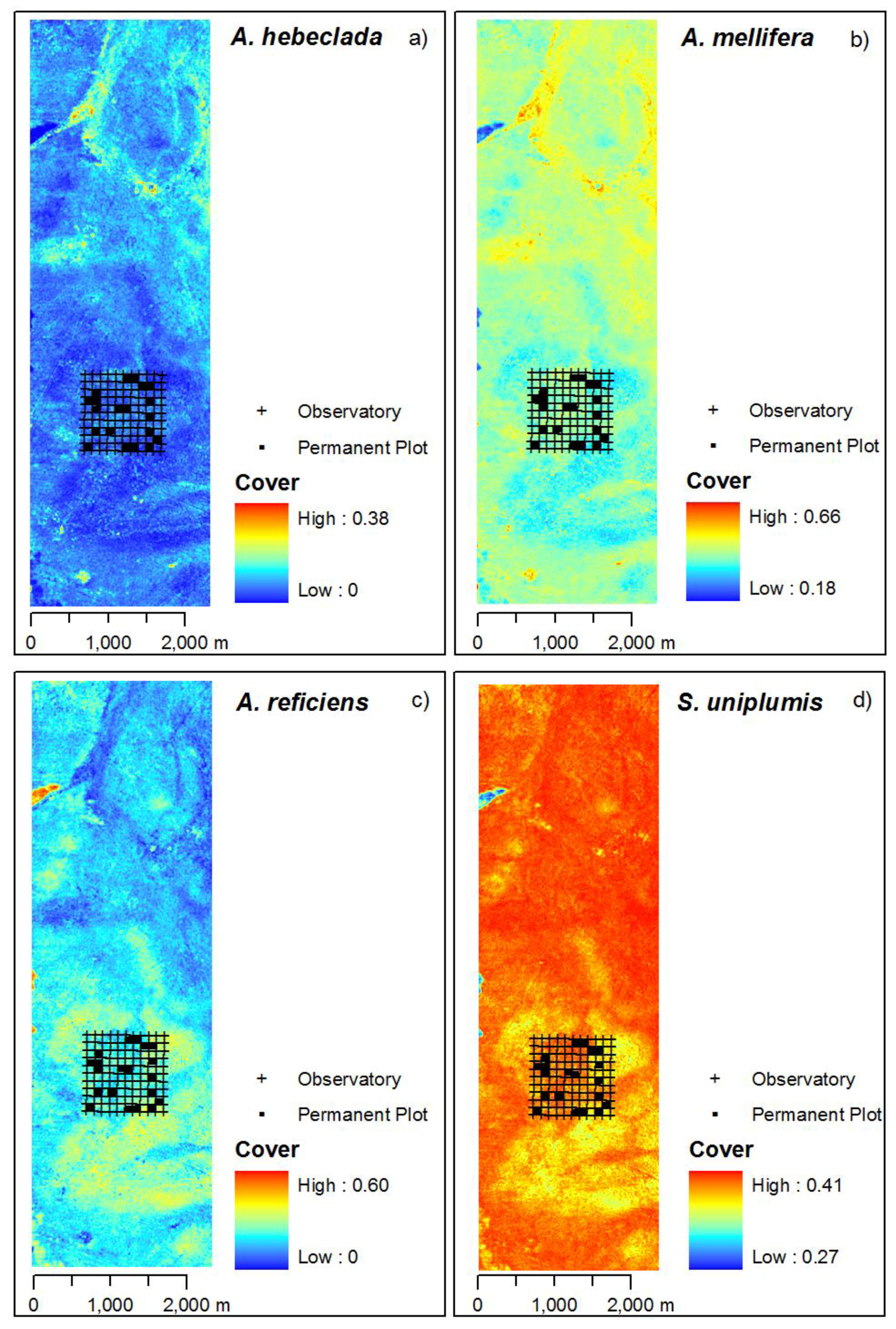
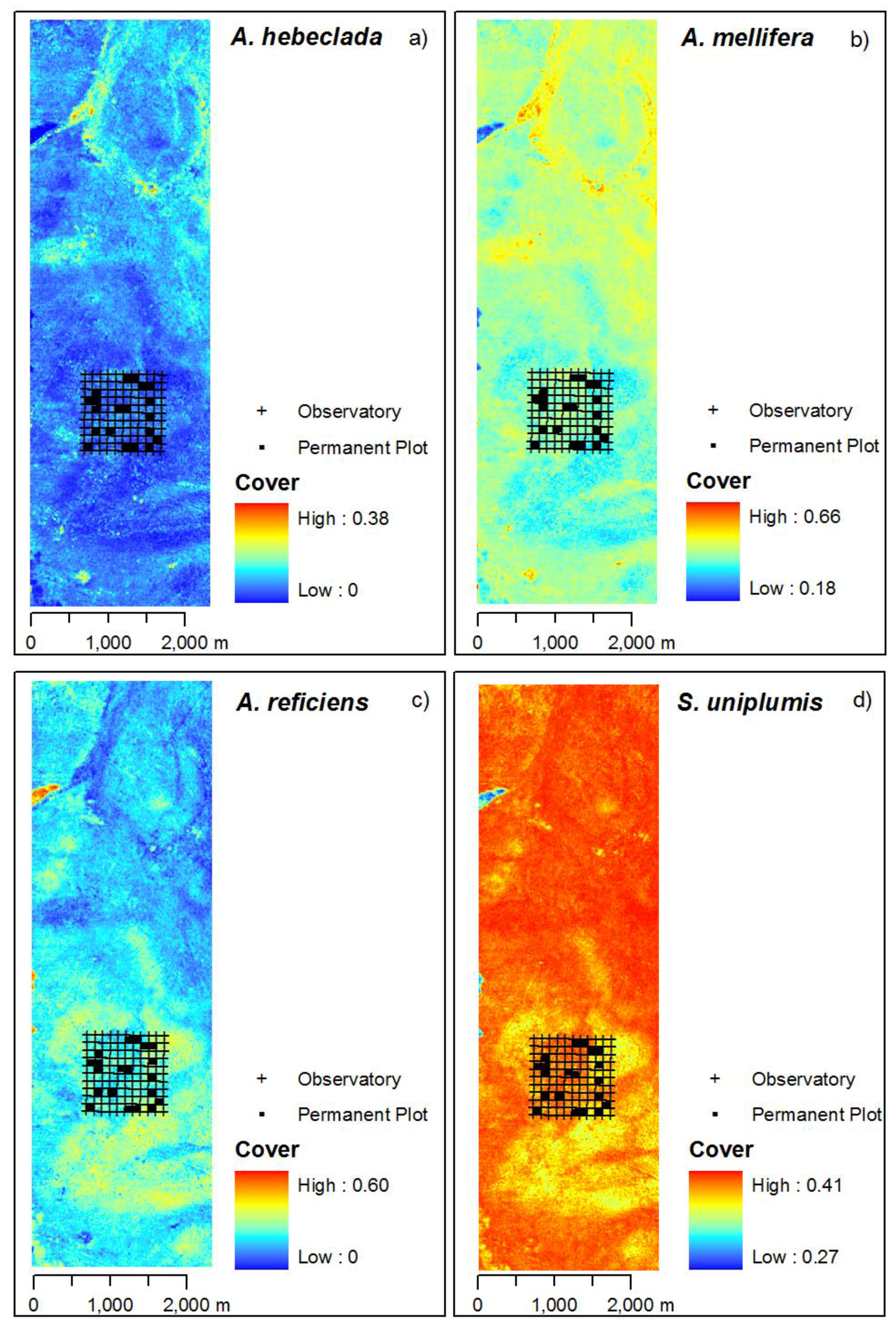
3.3. Mapping of the Principal Curve and Species Cover

3.4 Validation of Species Cover Maps
| Size | Species | n | Pearson’s r | p | year | Spearmann’s r | p | year |
|---|---|---|---|---|---|---|---|---|
| 10 × 10 | A.hebeclada | 3 | −0.11 | 0.642 | 2004 | −0.27 | 0.248 | 2004 |
| A.mellifera | 12 | 0.54 | 0.014 | 2005 | 0.21 | 0.375 | 2005 | |
| A.reficiens | 9 | 0.23 | 0.333 | 2005 | 0.46 | 0.043 | 2004 | |
| S.uniplumis | 20 | 0.37 | 0.105 | 2005 | 0.32 | 0.174 | 2005 | |
| 20 × 50 | A.hebeclada | 6 | 0.13 | 0.592 | 2004 | 0.18 | 0.443 | 2004 |
| A.mellifera | 19 | 0.45 | 0.047 | 2004 | 0.45 | 0.049 | 2004 | |
| A.reficiens | 17 | 0.32 | 0.165 | 2005 | 0.52 | 0.020 | 2005 | |
| S.uniplumis | 20 | 0.38 | 0.099 | 2004 | 0.26 | 0.262 | 2005 |
4. Discussion
4.1. Phenological Gradient
4.2. Species Responses
4.3. Mapped Vegetation Pattern
4.4. Validation of Species Cover Maps
4.5. Remote Sensing and Bush Encroachment
4.6. Principal Curves
5. Conclusion
Acknowledgements
References
- Archer, S.R. Tree-grass dynamics in a Prosopis-thornscrub savanna parkland: Reconstructing the past and predicting the future. Ecoscience 1995, 2, 83–99. [Google Scholar]
- Archer, S.; Boutton, T.W.; Hibbard, K.A. Trees in grasslands: Biogeochemical consequences of woody plant expansion. In Global Biogeochemical Cycles in the Climate System; Schulze, E.D., Harrison, S.P., Heimann, M., Holland, E.A., Lloyd, J., Prentice, I.C., Schimel, D., Eds.; Academic Press: San Diego, CA, USA, 2000; p. 47. [Google Scholar]
- Ward, D. Do we understand the causes of bush encroachment in African savannas? Afr. J. Range Forage Sci. 2005, 22, 101–105. [Google Scholar] [CrossRef]
- de Klerk, J.N. Bush Encroachment in Namibia; Ministry of Environment and Tourism: Windhoek, Namibia, 2004; p. 255. [Google Scholar]
- Tews, J.; Moloney, K.; Jeltsch, F. Modeling seed dispersal in a variable environment: A case study of the fleshy-fruited savanna shrub Grewia flava. Ecol. Model. 2004, 175, 65–76. [Google Scholar] [CrossRef]
- Tietjen, B.; Jeltsch, F.; Zehe, E.; Classen, N.; Groengroeft, A.; Schiffers, K.; Oldeland, J. Effects of climate change on the coupled dynamics of water and vegetation in drylands. Ecohydrol. 2010. [Google Scholar] [CrossRef]
- Mugasi, S.K.; Sabiiti, E.N.; Tayebwa, B.M. The economic implications of bush encroachment on livestock farming in rangelands of Uganda. Afr. J. Range Forage Sci. 2000, 17, 64–69. [Google Scholar] [CrossRef]
- Van Eck, J.A.J.; Swanepol, D.J. Chemical bush control on various invader species, using different arboricides and control methods—an analysis. Agricola. 2008, 18, 37–50. [Google Scholar]
- Meik, J.M.; Jeo, R.M.; Mendelson, J.R., III; Jenks, K.E. Effects of bush encroachment on an assemblage of diurnal lizard species in central Namibia. Biol. Conserv. 2002, 106, 29–36. [Google Scholar] [CrossRef]
- Sirami, C.; Seymour, C.; Midgley, G.; Barnard, P. The impact of shrub encroachment on savanna bird diversity from local to regional scale. Divers. Distrib. 2009, 15, 948–957. [Google Scholar] [CrossRef]
- Skarpe, C. Structure of the woody vegetation in disturbed and undisturbed Arid Savanna, Botswana. Vegetatio 1990, 87, 11–18. [Google Scholar] [CrossRef]
- Hudak, A.T.; Wessman, C.A. Textural analysis of historical aerial photography to characterize woody plant encroachment in South African savanna. Remote Sens. Environ. 1998, 66, 317–330. [Google Scholar] [CrossRef]
- Hudak, A.T.; Wessman, C.A. Textural analysis of high resolution imagery to quantify bush encroachment in Madikwe Game Reserve, South Africa, 1955–1996. Int. J. Remote Sens. 2001, 22, 2731–2740. [Google Scholar] [CrossRef]
- Moleele, N.; Ringrose, S.; Arnberg, W.; Lunden, B.; Vanderpost, C. Assessment of vegetation indexes useful for browse (forage) prediction in semi-arid rangelands. Int. J. Remote Sens. 2001, 22, 741–756. [Google Scholar] [CrossRef]
- Wagenseil, H. Woody Vegetation Cover and Bush Encroachment in Namibia: A Modelling Approach Based on Remote Sensing. Ph.D. Thesis, University of Erlangen-Nürnberg, Erlangen, Germany, 2008; p. 216. [Google Scholar]
- Zimmermann, I.; Joubert, D.; Smit, G.N. A problem tree to diagnose a problem bush. Agricola 2008, 18, 27–36. [Google Scholar]
- Kotiluoto, R.; Ruokolainen, K.; Kettunen, M. Invasive Acacia auriculiformis Benth. in different habitats in Unguja, Zanzibar. Afr. J. Ecol. 2009, 47, 77–86. [Google Scholar] [CrossRef]
- Moleele, N.M.; Ringrose, S.; Matheson, W.; Vanderpost, C. More woody plants? The status of bush encroachment in Botswana’s grazing areas. J. Environ. Manage. 2002, 64, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Joubert, D.F.; Rothauge, A.; Smit, G.N. A conceptual model of vegetation dynamics in the semiarid Highland savanna of Namibia, with particular reference to bush thickening by Acacia mellifera. J. Arid Environ. 2008, 72, 2201–2210. [Google Scholar] [CrossRef]
- Curtis, B.A.; Mannheimer, C.A. Tree Atlas of Namibia; National Botanical Research Institute (NBRI): Windhoek, Namibia, 2005; p. 688. [Google Scholar]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trend. Ecol. Evolut. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Hüttich, C.G.U.; Herold, M.; Strohbach, B.J.; Schmidt, M.; Keil, M.; Dech, S. On the suitability of MODIS time series metrics to map vegetation types in dry savanna ecosystems: A case study in the Kalahari of NE Namibia. Remote Sens. 2009, 1, 620–643. [Google Scholar] [CrossRef]
- Resasco, J.; Hale, A.N.; Henry, M.C.; Gorchov, D.L. Detecting an invasive shrub in a deciduous forest understory using late-fall Landsat sensor imagery. Int. J. Remote Sens. 2007, 28, 3739–3745. [Google Scholar] [CrossRef]
- Castro-Esau, K.L.; Sanchez-Azofeifa, G.A.; Rivard, B.; Wright, S.J.; Quesada, M. Variability in leaf optical properties of Mesoamerican trees and the potential for species classification. Amer. J. Bot. 2006, 93, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Castro-Esau, K.L.; Kalacska, M. Tropical dry forest phenology and discrimination of tropical tree species using hyperspectral data. In Hyperspectral Remote Sensing of Tropical and Sub-Tropical Forests; Kalacska, M., Sanchez-Azofeifa, G.A., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–26. [Google Scholar]
- Giess, W. A preliminary vegetation map of South West Africa. Dinteria 1971, 4, 5–114. [Google Scholar]
- Petersen, A. Pedodiversity of Southern African Drylands. Ph.D. Thesis, University of Hamburg, Hamburg, Germany, 2008; p. 408. [Google Scholar]
- Rolecek, J.; Chytry, M.; Hajek, M.; Lvoncik, S.; Tichy, L. Sampling design in large-scale vegetation studies: Do not sacrifice ecological thinking to statistical purism! Folia Geobot. 2007, 42, 199–208. [Google Scholar] [CrossRef]
- Kent, M.; Coker, P. Vegetation Description and Analysis: A Practical Approach; John Wiley & Sons: Chichester, UK, 1995; p. 363. [Google Scholar]
- Rao, C.R. A review of canonical coordinates and an alternative to correspondence analysis using Hellinger Distance. Questiio 1995, 19, 23–63. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Cocks, T.; Jenssen, R.; Stewart, A.; Wilson, I.; Shields, T. The hymap hyperspectral sensor: The system, calibration and performance. In Proceedings of 1st EARSEL Workshop on Imaging Spectroscopy, Zurich, Switzerland; 1998; p. 7. [Google Scholar]
- Schläpfer, D.; Richter, R. Geo-atmospheric processing of airborne imaging spectrometry data. Part 1: Parametric orthorectification. Int. J. Remote Sens. 2002, 23, 2609–2630. [Google Scholar] [CrossRef]
- Richter, R.; Schläpfer, D. Geo-atmospheric processing of airborne imaging spectrometry data, Part 2: Atmospheric/topographic correction. Int. J. Remote Sens. 2002, 23, 2631–2649. [Google Scholar] [CrossRef]
- Singh, A. Digital change detection techniques using remotely-sensed data. Int. J. Remote Sens. 1989, 10, 989–1003. [Google Scholar] [CrossRef]
- Coppin, P.; Jonckheere, I.; Nackaerts, K.; Muys, B.; Lambin, E. Digital change detection methods in ecosystem monitoring: A review. Int. J. Remote Sens. 2004, 25, 1565–1596. [Google Scholar] [CrossRef]
- Lu, D.; Mausel, P.; Brondizio, E.; Moran, E. Change detection techniques. Int. J. Remote Sens. 2004, 25, 2365–2407. [Google Scholar] [CrossRef]
- Oldeland, J.; Dorigo, W.; Lieckfeld, L.; Jürgens, N. Connecting spectral indices, constrained ordination and fuzzy classification as an innovative approach for mapping vegetation types. Remote Sens. Environ. 2010, 114, 1155–1166. [Google Scholar] [CrossRef]
- Dorigo, W.; Richter, R.; Baret, F.; Bamler, R.; Wagner, W. Enhanced automated canopy characterization from hyperspectral data by a novel two step radiative transfer model inversion approach. Remote Sens. 2009, 1, 1139–1170. [Google Scholar] [CrossRef]
- Kim, M.S.; Daughtry, C.S.T.; Chappelle, E.W.; McMurtrey, J.E., III; Walthall, C.L. The use of high spectral resolution bands for estimating absorbed photosynthetically active radiation (APAR). In Proceedings of the 6th Symposium on Physical Measurements and Signatures in Remote Sensing, Val D’Isere, France, 1994; pp. 299–306.
- Elvidge, C.D.; Chen, Z. Comparison of broad-band and narrow-band red and near-infrared vegetation indices. Remote Sens. Environ. 1995, 54, 38–48. [Google Scholar] [CrossRef]
- Galváo, L.S.; Formaggio, A.R.; Tisot, D.A. Discrimination of sugarcane varieties in southeastern Brazil with EO-1 Hyperion data. Remote Sens. Environ. 2005, 94, 523–534. [Google Scholar] [CrossRef]
- Serrano, L.; Penuelas, J.; Ustin, S.L. Remote sensing of nitrogen and lignin in Mediterranean vegetation from AVIRIS data: Decomposing biochemical from structural signals. Remote Sens. Environ. 2002, 81, 355–364. [Google Scholar] [CrossRef]
- Nagler, P.L.; Inoue, Y.; Glenn, E.P.; Russ, A.L.; Daughtry, C.S.T. Cellulose absorption index (CAI) to quantify mixed soil-plant litter scenes. Remote Sens. Environ. 2003, 87, 310–325. [Google Scholar] [CrossRef]
- Hastie, T.; Stuetzle, W. Principal curves. J. Amer. Statist. Assn. 1989, 84, 502–516. [Google Scholar] [CrossRef]
- Ter Braak, C.J.F.; Prentice, C. A theory of gradient analysis. Adv. Ecol. Res. 2004, 34, 235–282. [Google Scholar]
- De’ath, G. Principal curves: A new technique for indirect and direct gradient analysis. Ecology 1999, 80, 2237–2253. [Google Scholar] [CrossRef]
- Izenman, A.J. Modern Multivariate Statistical Techniques: Regression, Classification, and Manifold Learning; Springer: Berlin, Germany, 2008; p. 734. [Google Scholar]
- De’ath, G. Extended dissimilarity: A method of robust estimation of ecological distances from high beta diversity data. Plant Ecol. 1999, 144, 191–199. [Google Scholar] [CrossRef]
- Hastie, T.; De’ath, G.; Walsh, C. pcurve: Principal Curve Analysis; R Package Version 0.6-2; 2005. [Google Scholar]
- R-Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009; Available online: http://www.R-project.org (accessed on May 15, 2009).
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Jürgens, N. Transkontinentale Makrotransekte als Instrument zur Erforschung der Biodiversität. Ein konkreter Vorschlag für einen Projektverbund: BIOTA—Biodiversity Monitoring Transect Analysis. In Biodiversitätsforschung in Deutschland. Potentiale und Perspektiven; Barthlott, W., Gutmann, M., Eds.; Graue Reihe-Europäische Akademie: Bad Neuenahr-Ahrweiler, Germany, 1998; pp. 61–65. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electronica 2001, 4, 1–9. [Google Scholar]
- Dennison, P.E.; Roberts, D.A. The effects of vegetation phenology on endmember selection and species mapping in southern California chaparral. Remote Sens. Environ. 2003, 87, 295–309. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; Vander Zee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113 (Suppl. 1), S78. [Google Scholar] [CrossRef]
- Patton, A.R.; Gieseker, L. Seasonal changes in the lignin and cellulose content of some montana grasses. J. Anim Sci. 1942, 1, 22–26. [Google Scholar]
- Robbins, C.T.; Moen, A.N. Composition and digestibility of several deciduous browses in the northeast. J. Wildlife Manage. 1975, 39, 337–341. [Google Scholar] [CrossRef]
- Skarpe, C. Plant community structure in relation to grazing and environmental-changes along a north-south transect in the western Kalahari. Vegetatio 1986, 68, 3–18. [Google Scholar]
- Scholes, R.J.; Archer, S.R. Tree-grass interactions in savannas. Ann. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Chidumayo, E.N. Climate and phenology of savanna vegetation in southern Africa. J. Veg. Sci. 2001, 12, 347–354. [Google Scholar] [CrossRef]
- Archibald, S.; Scholes, R.J. Leaf green-up in a semi-arid African savanna -separating tree and grass responses to environmental cues. J. Veg. Sci. 2007, 18, 583–594. [Google Scholar] [CrossRef]
- Coates-Palgrave, M. Trees of Southern Africa, 3rd ed.; Coates-Palgrave, K., Ed.; Struik: Cape Town, South Africa, 2002; p. 1212. [Google Scholar]
- Jeltsch, F.; Moloney, K.A.; Schurr, F.M.; Kochy, M.; Schwager, M. The state of plant population modelling in light of environmental change. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 171–189. [Google Scholar] [CrossRef]
- Wiegand, K.; Saitz, D.; Ward, D. A patch-dynamics approach to savanna dynamics and woody plant encroachment—Insights from an arid Savanna. Perspect. Plant Ecol. Evol. Syst. 2006, 7, 229–242. [Google Scholar] [CrossRef]
- Wiegand, K.; Ward, D.; Saltz, D. Multi-scale patterns and bush encroachment in an arid savanna with a shallow soil layer. J. Veg. Sci. 2005, 16, 311–320. [Google Scholar] [CrossRef]
- Schultka, W.; Cornelius, R. Vegetation structure of a heavily grazed range in northern Kenya: Tree and shrub canopy. J. Arid Environ. 1997, 36, 291–306. [Google Scholar] [CrossRef]
- van Vegten, J.A. Thornbush invasion in a Savanna ecosystem in Eastern Botswana. Vegetatio 1984, 56, 3–7. [Google Scholar] [CrossRef]
- Nagendra, H.; Rocchini, D. High resolution satellite imagery for tropical biodiversity studies: The devil is in the detail. Biodivers. Conserv. 2008, 17, 3431–3442. [Google Scholar] [CrossRef]
- Bastiaanssen, W.G.M.; Molden, D.J.; Makin, I.W. Remote sensing for irrigated agriculture: Examples from research and possible applications. Agr. Water Manage. 2000, 46, 137–155. [Google Scholar] [CrossRef]
- Kennedy, R.E.; Townsend, P.A.; Gross, J.E.; Cohen, W.B.; Bolstad, P.; Wang, Y.Q.; Adams, P. Remote sensing change detection tools for natural resource managers: Understanding concepts and tradeoffs in the design of landscape monitoring projects. Remote Sens. Environ. 2009, 113, 1382–1396. [Google Scholar] [CrossRef]
- Banfield, J.D.; Raftery, A.E. Ice floe identification in satellite images using mathematical morphology and clustering about principal curves. J. Amer. Statist. Assn. 1992, 87, 7–16. [Google Scholar] [CrossRef]
- Stanford, D.C.; Raftery, A.E. Finding curvilinear features in spatial point patterns: Principal curve clustering with noise. IEEE Trans. Patt. Anal. Mach. Int. 2000, 22, 601–609. [Google Scholar] [CrossRef]
- Chang, K.-y.; Ghosh, J. Principal curves for nonlinear feature extraction and classification. In SPIE Applications of Artificial Neural Networks in Image Processing III; Nasrabadi, N.M., Katsaggelos, A.K., Eds.; SPIE: Bellingham, WA, USA, 1998; Volume 3307, pp. 120–129. [Google Scholar]
- Armitage, R.P.; Weaver, R.E.; Kent, M. Remote sensing of semi-natural upland vegetation: the relationship between species composition and spectral response. In Vegetation Mapping: From Patch to Planet; Biogeography Research Group Symposia Series; Alexander, R., Millington, A.C., Eds.; John Wiley and Sons Ltd: Chichester, UK, 2000; pp. 83–102. [Google Scholar]
- Schmidtlein, S.; Zimmermann, P.; Schupferling, R.; Weiss, C. Mapping the floristic continuum: Ordination space position estimated from imaging spectroscopy. J. Veg. Sci. 2007, 18, 131–140. [Google Scholar] [CrossRef]
- van Leeuwen, W.J.D.; Davison, J.E.; Casady, G.M.; Marsh, S.E. Phenological characterization of Desert Sky Island vegetation communities with remotely sensed and climate time series data. Remote Sens. 2010, 2, 388–415. [Google Scholar] [CrossRef]
- Palmer, A. R.; Fortescue, A. Remote sensing and change detection in rangelands. Afr. J. Range Forage Sci. 2004, 21, 123–128. [Google Scholar] [CrossRef]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Oldeland, J.; Dorigo, W.; Wesuls, D.; Jürgens, N. Mapping Bush Encroaching Species by Seasonal Differences in Hyperspectral Imagery. Remote Sens. 2010, 2, 1416-1438. https://doi.org/10.3390/rs2061416
Oldeland J, Dorigo W, Wesuls D, Jürgens N. Mapping Bush Encroaching Species by Seasonal Differences in Hyperspectral Imagery. Remote Sensing. 2010; 2(6):1416-1438. https://doi.org/10.3390/rs2061416
Chicago/Turabian StyleOldeland, Jens, Wouter Dorigo, Dirk Wesuls, and Norbert Jürgens. 2010. "Mapping Bush Encroaching Species by Seasonal Differences in Hyperspectral Imagery" Remote Sensing 2, no. 6: 1416-1438. https://doi.org/10.3390/rs2061416