Use of RGB Vegetation Indexes in Assessing Early Effects of Verticillium Wilt of Olive in Asymptomatic Plants in High and Low Fertility Scenarios

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Physiological Measurements
2.3. RGB Vegetation Indexes
2.4. Leaf Pigment Extractions
2.5. Data Analysis
3. Results
3.1. Physiological Measurements
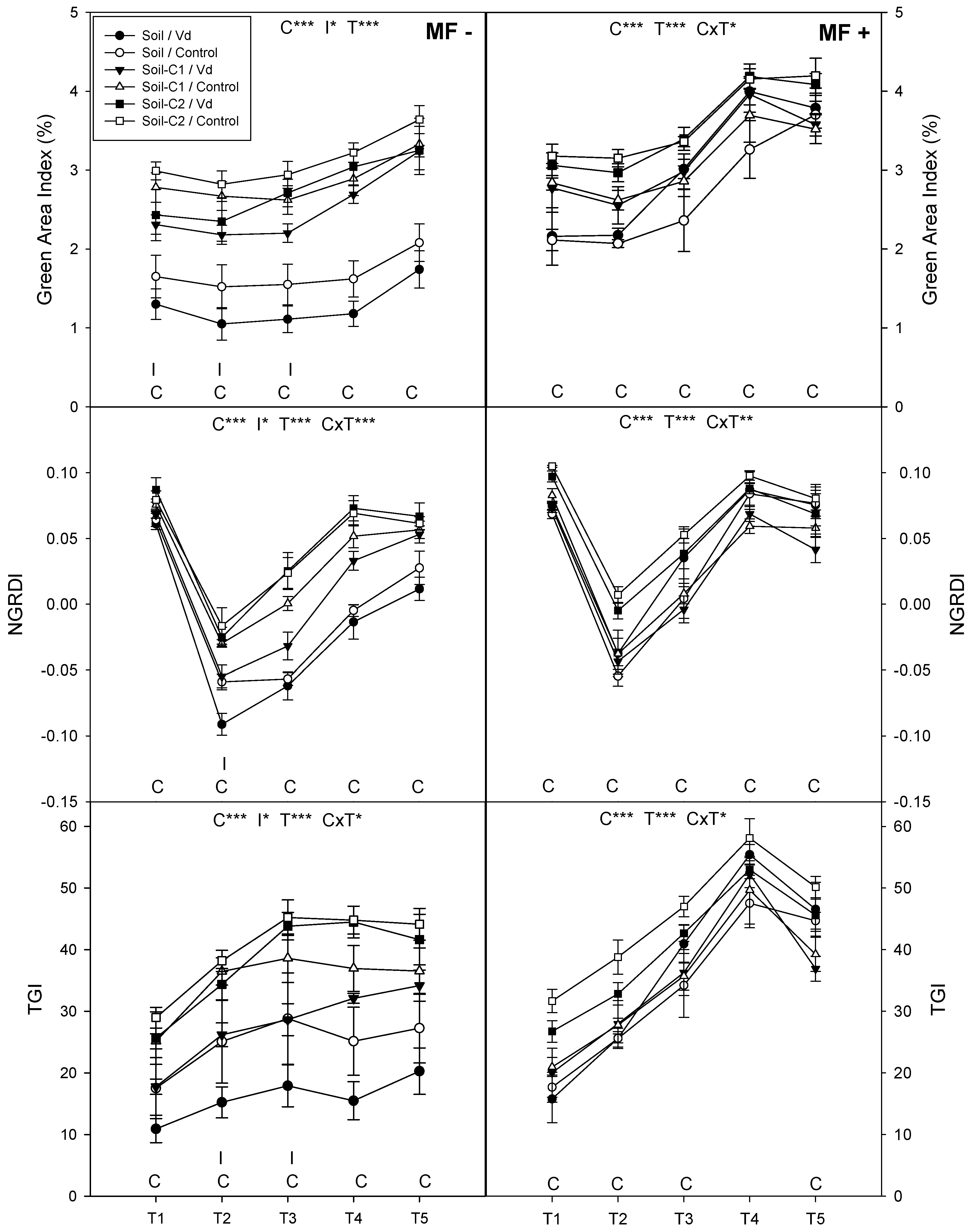
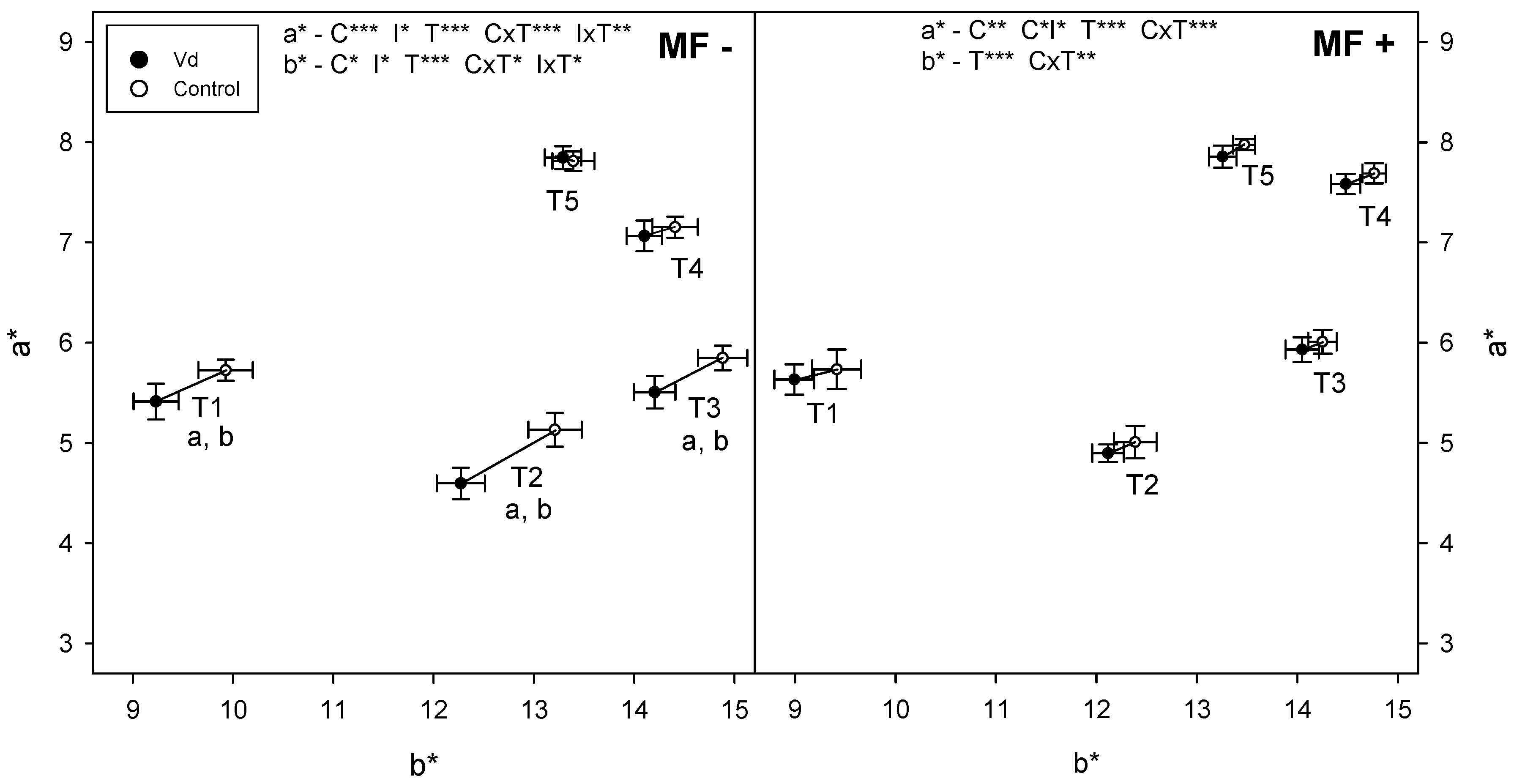
3.2. RGB Vegetation Indexes
3.3. Leaf Pigment Extractions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Keykhasaber, M.; Pham, K.T.K.; Thomma, B.P.H.J.; Hiemstra, J.A. Reliable detection of unevenly distributed Verticillium dahliae in diseased olive trees. Plant Pathol. 2017, 66, 641–650. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Bostock, R.M.; Davis, R.M.; Usami, T.; Platt, H.W.; Subbarao, K.V. Phylogenetics and taxonomy of the fungal vascular wilt pathogen Verticillium, with the descriptions of five new species. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Messner, R.; Schweigkofler, W.; Ibl, M.; Berg, G.; Prillinger, H. Molecular characterization of the plant pathogen Verticillium dahliae Kleb. Using RAPD-PCR and sequencing of the 18SrRNA-Gene. J. Phytopathol. 1996, 144, 347–354. [Google Scholar] [CrossRef]
- Wilhelm, S. Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopathology 1955, 45, 180–181. [Google Scholar]
- Prieto, P.; Navarro-Raya, C.; Valverde-Corredor, A.; Amyotte, S.G.; Dobinson, K.F.; Mercado-Blanco, J. Colonization process of olive tissues by Verticillium dahliae and its in planta interaction with the biocontrol root endophyte Pseudomonas fluorescens PICF7. Microb. Biotechnol. 2009, 2, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Pegg, G.; Brady, B. Verticillium Wilts; CAB International: Wallingford, UK, 2002. [Google Scholar]
- Scholz, S.S.; Schmidt-Heck, W.; Guthke, R.; Furch, A.C.U.; Reichelt, M.; Gershenzon, J.; Oelmüller, R. Verticillium dahliae-Arabidopsis interaction causes changes in gene expression profiles and jasmonate levels on different time scales. Front. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef] [PubMed]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef]
- Fradin, E.F.; Thomma, B.P.H.J. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Roca, L.F.; Moral, J.; Trapero, C.; Blanco-López, M.Á.; López-Escudero, F.J. Effect of inoculum density on Verticillium wilt incidence in commercial olive orchards. J. Phytopathol. 2016, 164, 61–64. [Google Scholar] [CrossRef]
- Calderón, R.; Lucena, C.; Trapero-Casas, J.L.; Zarco-Tejada, P.J.; Navas-Cortés, J.A. Soil temperature determines the reaction of olive cultivars to Verticillium dahliae pathotypes. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Bowden, R.L.; Rouse, D.I.; Sharkey, T.D. Mechanism of photosynthesis decrease by Verticillium dahliae in potato. Plant Physiol. 1990, 94, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Sadras, V.O.; Quiroz, F.; Echarte, L.; Escande, A.; Pereyra, V.R. Effect of Verticillium dahliae on photosynthesis, leaf expansion and senescence of field-grown sunflower. Ann. Bot. 2000, 86, 1007–1015. [Google Scholar] [CrossRef]
- Mercado-Blanco, J.; Rodriguez-Jurado, D.; Pérez-Artés, E.; Jiménez-Díaz, R.M. Detection of the nondefoliating pathotype of Verticillium dahliae in infected olive plants by nested PCR. Plant Pathol. 2001, 50, 609–619. [Google Scholar] [CrossRef]
- Maldonado-González, M.M.; Bakker, P.A.H.M.; Prieto, P.; Mercado-Blanco, J. Arabidopsis thaliana as a tool to identify traits involved in Verticillium dahliae biocontrol by the olive root endophyte Pseudomonas fluorescens PICF7. Front. Microbiol. 2015, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, E.C. Recovery of olive trees with Verticillium wilt after individual application of soil solarization in established olive orchards. Plant Dis. 1991, 75, 557. [Google Scholar] [CrossRef]
- Bejarano-AIcazar, J. Etiology, Importance, and distribution of Verticillium wilt of cotton in Southern Spain. Plant Dis. 1996, 80, 1233. [Google Scholar] [CrossRef]
- Isaac, I. A Comparative study of pathogenic isolates of Verticillium. Trans. Br. Mycol. Soc. 1949, 32, 137-IN5. [Google Scholar] [CrossRef]
- Jiménez-Díaz, R.M.; Cirulli, M.; Bubici, G.; del Mar Jiménez-Gasco, M.; Antoniou, P.P.; Tjamos, E.C. Verticillium wilt, a major threat to olive production: Current status and future prospects for its management. Plant Dis. 2012, 96, 304–329. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.F.; Cook, R.J. Biological control of plant pathogens. Am. Phytopathol. Soc. St. Paul MN 1974. [Google Scholar] [CrossRef]
- Avilés, M.; Borrero, C.; Trillas, M.I. Review on compost as an inducer of disease suppression in plants grown in soilless culture. Dyn. Soil Dyn. Plant 2011, 5, 1–11. [Google Scholar]
- Segarra, G.; Santpere, E.; Elena, G.; Trillas, M.I. Enhanced Botrytis cinerea resistance of Arabidopsis plants grown in compost may be explained by increased expression of defense related genes, as revealed by microarray analysis. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Segarra, G.; Elena, G.; Trillas, I. Systemic Resistance against Botrytis cinerea in Arabidopsis triggered by an olive marc compost substrate requires functional SA signalling. Physiol. Mol. Plant Pathol. 2013, 82, 46–50. [Google Scholar] [CrossRef]
- Raviv, M. Compost as a tool to suppress plant diseases: Established and putative mechanisms. Acta Hortic. 2016, 1146, 11–24. [Google Scholar] [CrossRef]
- Papasotiriou, F.G.; Varypatakis, K.G.; Christofi, N.; Tjamos, S.E.; Paplomatas, E.J. Olive Mill Wastes: A source of resistance for plants against Verticillium dahliae and a reservoir of biocontrol agents. Biol. Control 2013, 67, 51–60. [Google Scholar] [CrossRef]
- Castaño, R.; Avilés, M. Factors that affect the capacity of growing media to suppress Verticillium wilt. Acta Hortic. 2013, 1013, 465–471. [Google Scholar] [CrossRef]
- Vergara-Díaz, O.; Zaman-Allah, M.A.; Masuka, B.; Hornero, A.; Zarco-Tejada, P.; Prasanna, B.M.; Cairns, J.E.; Araus, J.L. A novel remote sensing approach for prediction of maize yield under different conditions of nitrogen fertilization. Front. Plant Sci. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kefauver, S.C.; El-Haddad, G.; Vergara-Diaz, O.; Araus, J.L. RGB Picture vegetation indexes for high-throughput phenotyping platforms (HTPPs). Remote. Sens. Agr. Ecosyst. Hydrol. Xviii. 2015, 9637, 96370J. [Google Scholar]
- Maloney, P.V.; Petersen, S.R.A.; Navarro, D.; Marshall, A.L.; McKendry, J.M.; Costa, J.P.M. digital image analysis method for estimation of fusarium-damaged kernels in wheat. Crop Sci. 2014, 54, 2077–2083. [Google Scholar] [CrossRef]
- Zhou, B.; Elazab, A.; Bort, J.; Vergara, O.; Serret, M.D.; Araus, J.L. Low-cost assessment of wheat resistance to yellow rust through conventional RGB images. Comput. Electron. Agric. 2015, 116, 20–29. [Google Scholar] [CrossRef]
- Vergara-Diaz, O.; Kefauver, S.C.; Elazab, A.; Nieto-Taladriz, M.T.; Araus, J.L. Grain yield losses in yellow-rusted durum wheat estimated using digital and conventional parameters under field conditions. Crop J. 2015, 3, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Casadesús, J.; Kaya, Y.; Bort, J.; Nachit, M.M.; Araus, J.L.; Amor, S.; Ferrazzano, G.; Maalouf, F.; Maccaferri, M.; Martos, V.; et al. Using vegetation indices derived from conventional digital cameras as selection criteria for wheat breeding in water-limited environments. Ann. Appl. Biol. 2007, 150, 227–236. [Google Scholar] [CrossRef]
- Trussell, H.J.; Saber, E.; Vrhel, M. Color image processing. IEEE Signal Process. Mag. 2005, 22, 14–22. [Google Scholar] [CrossRef]
- Hunt, E.R.; Cavigelli, M.; Daughtry, C.S.T.; McMurtrey, J.E.; Walthall, C.L. Evaluation of digital photography from model aircraft for remote sensing of crop biomass and nitrogen status. Precis. Agric. 2005, 6, 359–378. [Google Scholar] [CrossRef]
- Raymond Hunt, E.R.; Daughtry, C.S.T.; Eitel, J.U.H.; Long, D.S. Remote sensing leaf chlorophyll content using a visible band index. Agron. J. 2011, 103, 1090–1099. [Google Scholar] [CrossRef]
- Hunt, E.R.; Doraiswamy, P.; McMurtrey, J.; Daughtry, C.; Perry, E.; Akhmedov, B. A visible band index for remote sensing leaf chlorophyll content at the canopy scale. Intl. J. Appl. Earth. Obs. Geoinf. 2013. [Google Scholar] [CrossRef]
- Garcia-Ruiz, G.M.; Trapero, C.; Del Rio, C.; Lopez-Escudero, F.J. Evaluation of resistance of Spanish olive cultivars to Verticillium dahliae in inoculations conducted in greenhouse. Phytoparasitica 2014, 42, 205–212. [Google Scholar] [CrossRef]
- Aviles, M.; Borrero, C. Identifying characteristics of Verticillium wilt suppressiveness in olive mill composts. Plant Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, J.D.; Israelstam, G.F. A Method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric analysis of chlorophylls and carotenoids from commonly grown fern species by using various extracting solvents. Synlett 2014, 25, 97–101. [Google Scholar]
- Romanyà, J.; Sancho-Adamson, M.; Ortega, D.; Trillas, M.I. Early stage effects of Verticillium wilt of olive (VWO) on nutrient use in young olive trees grown in soils amended with compost and mineral fertilisation. Plant Soil 2018. [Google Scholar] [CrossRef]
- Bibi, N.; Li, F.; Ahmed, I.M.; Fan, K.; Yuan, S.N.; Wang, X.D. Impact of Verticillium dahliae toxin on morphogenetic, physiological and biochemical characteristics of upland cotton. Pak. J. Bot. 2017, 49, 191–199. [Google Scholar]
- Buhtz, A.; Hohe, A.; Schwarz, D.; Grosch, R. Effects of Verticillium dahliae on tomato root morphology considering plant growth response and defence. Plant Pathol. 2017, 66, 667–676. [Google Scholar] [CrossRef]
- López-Escudero, F.J.; Del Río, C.; Caballero, J.M.; Blanco-López, M.A. Evaluation of olive cultivars for resistance to verticillium dahliae. Eur. J. Plant Pathol. 2004, 110, 79–85. [Google Scholar] [CrossRef]
- Olesen, J.E.; Petersen, B.M.; Berntsen, J.; Hansen, S.; Jamieson, P.D.; Thomsen, A.G. Comparison of methods for simulating effects of nitrogen on green area index and dry matter growth in winter wheat. F. Crop. Res. 2002, 74, 131–149. [Google Scholar] [CrossRef]
- Tzeng, D.D.; Devay, J.E. Physiological responses of Gossypium hirsutum L. to infection by defoliating and non-defoliating pathotypes Verticillium dahliae Kleb. Physiol. Plant. Path. 1985, 26, 57–72. [Google Scholar] [CrossRef]
- Hampton, R.E.; Wullschleger, S.D.; Oosterhuis, D.M. Impact of Verticillium wilt on net photosynthesis, respiration and photorespiration in field-grown cotton (Gossypium hirsutum L.). Physiol. Mol. Plant Pathol. 1990, 37, 271–280. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | EC (mS cm−1) | C (%) | C/N | N (%) | P Olsen (mg/kg) | P (%) | K (%) | S (%) | Ca (%) | Mg (%) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil | 8.58 | 111.8 | 0.86 | 10.75 | 0.08 | 12.4 | ND | ND | ND | ND | ND |
| C1 | 9.51 | 3345 | 33.91 | 22.31 | 1.52 | 59.3 | 0.15 | 2.26 | 0.19 | 7.14 | 1.29 |
| Soil + C1 | 8.81 | 403 | 2.56 | 15.06 | 0.17 | 21 | ND | ND | ND | ND | ND |
| C2 | 8.7 | 3655 | 28.71 | 14.58 | 1.97 | 340 | 0.42 | 1.9 | 0.5 | 9.05 | 0.97 |
| Soil + C2 | 8.9 | 650.5 | 2.39 | 11.38 | 0.21 | 63.1 | ND | ND | ND | ND | ND |
| MF- | MF+ | |||||||
|---|---|---|---|---|---|---|---|---|
| Physiological Parameters | Overall | I | C | T | Overall | I | C | T |
| ΦPSII Y | 0.0000 | NS | NS | 0.0000 | 0.0074 | NS | NS | 0.0000 |
| Fv/Fm Y | 0.0000 | NS | NS | 0.0000 | 0.0001 | NS | NS | NS |
| Gs Y | 0.0000 | NS | NS | 0.0000 | 0.0014 | NS | NS | 0.0000 |
| ΦPSII M | 0.0027 | NS | 0.0294 | 0.0000 | 0.0018 | NS | 0.0000 | 0.0000 |
| Fv/Fm M | 0.0000 | NS | 0.0245 | 0.0000 | 0.0005 | NS | 0.0001 | 0.0015 |
| Gs M | 0.0001 | NS | NS | 0.0000 | 0.0095 | NS | NS | 0.0000 |
| RGB Indexes | ||||||||
| Intensity | 0.0000 | NS | 0.0126 | 0.0000 | 0.0000 | NS | 0.0005 | 0.0000 |
| Hue | 0.0000 | NS | 0.0019 | 0.0000 | 0.0000 | NS | 0.0480 | 0.0000 |
| Saturation | 0.0000 | 0.0260 | 0.0026 | 0.0000 | 0.0000 | NS | 0.0007 | 0.0000 |
| Lightness | 0.0000 | NS | NS | 0.0000 | 0.0000 | NS | 0.0001 | 0.0000 |
| a* | 0.0000 | 0.0129 | 0.0000 | 0.0000 | 0.0000 | NS | 0.0014 | 0.0000 |
| b* | 0.0000 | 0.0399 | 0.0256 | 0.0000 | 0.0000 | NS | NS | 0.0000 |
| u* | 0.0000 | NS | 0.0000 | 0.0000 | 0.0000 | NS | 0.0007 | 0.0000 |
| v* | 0.0000 | NS | NS | 0.0000 | 0.0000 | NS | NS | 0.0000 |
| GA | 0.0000 | 0.0213 | 0.0000 | 0.0000 | 0.0000 | NS | 0.0004 | 0.0000 |
| GGA | 0.0000 | NS | 0.0000 | 0.0000 | 0.0000 | NS | 0.0000 | 0.0000 |
| CSI | 0.0000 | NS | NS | 0.0000 | 0.0000 | NS | 0.0014 | 0.0000 |
| NGRDI | 0.0000 | 0.0452 | 0.0000 | 0.0000 | 0.0000 | NS | 0.0006 | 0.0000 |
| TGI | 0.0000 | 0.0301 | 0.0000 | 0.0000 | 0.0000 | NS | 0.0004 | 0.0000 |
| Pigment Extractions | ||||||||
| Chl a | 0.0000 | 0.0375 | 0.0012 | 0.0000 | 0.0395 | NS | NS | 0.0101 |
| Chl b | 0.0000 | 0.0049 | 0.0000 | 0.0000 | 0.0339 | NS | NS | 0.0036 |
| Chl a+b | 0.0000 | 0.0194 | 0.0000 | 0.0000 | 0.0050 | NS | NS | 0.0009 |
| Car | 0.0005 | 0.0041 | 0.0000 | NS | 0.0050 | NS | 0.0380 | 0.0000 |
| Car/Chl ratio | 0.0000 | NS | NS | 0.0000 | 0.0017 | NS | NS | 0.0000 |
| MF- | MF+ | |||||
|---|---|---|---|---|---|---|
| Physiological Parameters | Soil | Soil-C1 | Soil-C2 | Soil | Soil-C1 | Soil-C2 |
| PSII Y | 0.633 a | 0.650 a | 0.621 a | 0.704 a | 0.693 a | 0.690 a |
| Fv/Fm Y | 0.780 a | 0.778 a | 0.787 a | 0.809 a | 0.786 a | 0.803 a |
| PSII M | 0.650 a | 0.548 b | 0.650 a | 0.726 a | 0.525 b | 0.709 a |
| Fv/Fm M | 0.760 ab | 0.658 b | 0.779 a | 0.792 a | 0.635 b | 0.784 a |
| RGB Indexes | ||||||
| Saturation | 0.119b | 0.115 b | 0.132 a | 0.120 b | 0.116 b | 0.129 a |
| Hue | 73.854 b | 76.472 a | 77.043 a | 77.791 b | 77.986 b | 79.475 a |
| a* | 5.898 b | 6.087 b | 6.644 a | 6.390 b | 6.210 b | 6.695 a |
| b* | 12.731 b | 12.541 a | 13.402 a | 12.7432 a | 12.528 a | 12.884 a |
| u* | 1.564 c | 1.881 b | 2.226 a | 2.174 b | 2.040 b | 2.471 a |
| v* | 14.129 a | 13.948 a | 14.727 a | 14.118 a | 13.912 a | 14.011 a |
| GA | 1.200 b | 1.635 a | 1.707 a | 2.863 b | 3.137 b | 3.573 a |
| GGA | 0.675 b | 1.333 a | 1.494 a | 1.531 b | 1.545 b | 1.945 a |
| CSI | 55.314 a | 51.563 a | 50.106 a | 47.779 b | 51.839 a | 46.219 b |
| NGRDI | -0.012 c | 0.022 b | 0.044 a | 0.041 b | 0.031 b | 0.063 a |
| TGI | 20.370 c | 31.255 b | 39.117a | 35.401 b | 34.676 b | 42.650 a |
| Pigment Extractions | ||||||
| Chl a | 0.594 b | 0.731 a | 0.771 a | 0.888 a | 0.915 a | 0.919 a |
| Chl b | 0.309 b | 0.372 a | 0.386 a | 0.414 a | 0.436 a | 0.430 a |
| Chl a + b | 0.902 b | 1.103 a | 1.157 a | 1.302 a | 1.351 a | 1.349 a |
| Car | 0.227 b | 0.259 a | 0.266 a | 0.267 b | 0.293 a | 0.278 ab |
| Car/Chl | 0.264 a | 0.242 a | 0.234 a | 0.207 a | 0.219 a | 0.208 a |
| MF | Measure | hue | sat | a* | b* | a*/b* | GA | GGA | CSI | NGRDI | TGI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MF- | Chl a | 0.5394 *** | −0.2239 * | 0.3855 *** | −0.2792 *** | 0.5304 *** | 0.3762 *** | 0.5236 *** | −0.5235 *** | 0.4654 *** | NS |
| Chl b | 0.6435 *** | NS | 0.5349 *** | NS | 0.6324 *** | 0.4862 *** | 0.6437 *** | −0.6273 *** | 0.5634 *** | 0.2469 * | |
| Chl a + b | 0.5937 *** | −0.2037 * | 0.4477 *** | −0.2391 * | 0.5837 *** | 0.4258 *** | 0.5823 *** | −0.5771 *** | 0.5148 *** | NS | |
| Car | NS | NS | NS | NS | NS | 0.3625 *** | 0.2756 *** | NS | 0.2758 ** | 0.2850 ** | |
| Car/Chl | −0.6044 *** | 0.2381 * | −0.4592 *** | 0.2832 ** | −0.6027 *** | −0.2928 ** | −0.5375 *** | 0.6247 *** | −0.4567 *** | NS | |
| MF+ | Chl a | NS | NS | NS | −0.3043 ** | 0.2624 ** | NS | NS | NS | NS | NS |
| Chl b | 0.4188 *** | NS | 0.2912 ** | NS | NS | 0.2568 * | 0.3153 ** | −0.3497 *** | 0.2410 * | NS | |
| Chl a + b | 0.2457 * | NS | NS | −0.261 * | 0.2147 * | NS | NS | NS | NS | NS | |
| Car | −0.2672 ** | NS | −0.3617 *** | −0.2624 ** | 0.2219 * | −0.2434 * | −0.3596 *** | 0.3570 *** | −0.3240 ** | −0.3096 ** | |
| Car/Chl | −0.5936 *** | NS | −0.4778 *** | NS | NS | −0.3984 *** | −0.5437 *** | 0.5826 *** | −0.4294 *** | −0.2582 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho-Adamson, M.; Trillas, M.I.; Bort, J.; Fernandez-Gallego, J.A.; Romanyà, J. Use of RGB Vegetation Indexes in Assessing Early Effects of Verticillium Wilt of Olive in Asymptomatic Plants in High and Low Fertility Scenarios. Remote Sens. 2019, 11, 607. https://doi.org/10.3390/rs11060607
Sancho-Adamson M, Trillas MI, Bort J, Fernandez-Gallego JA, Romanyà J. Use of RGB Vegetation Indexes in Assessing Early Effects of Verticillium Wilt of Olive in Asymptomatic Plants in High and Low Fertility Scenarios. Remote Sensing. 2019; 11(6):607. https://doi.org/10.3390/rs11060607
Chicago/Turabian StyleSancho-Adamson, Marc, Maria Isabel Trillas, Jordi Bort, Jose Armando Fernandez-Gallego, and Joan Romanyà. 2019. "Use of RGB Vegetation Indexes in Assessing Early Effects of Verticillium Wilt of Olive in Asymptomatic Plants in High and Low Fertility Scenarios" Remote Sensing 11, no. 6: 607. https://doi.org/10.3390/rs11060607