Diurnal and Seasonal Variations in Chlorophyll Fluorescence Associated with Photosynthesis at Leaf and Canopy Scales

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
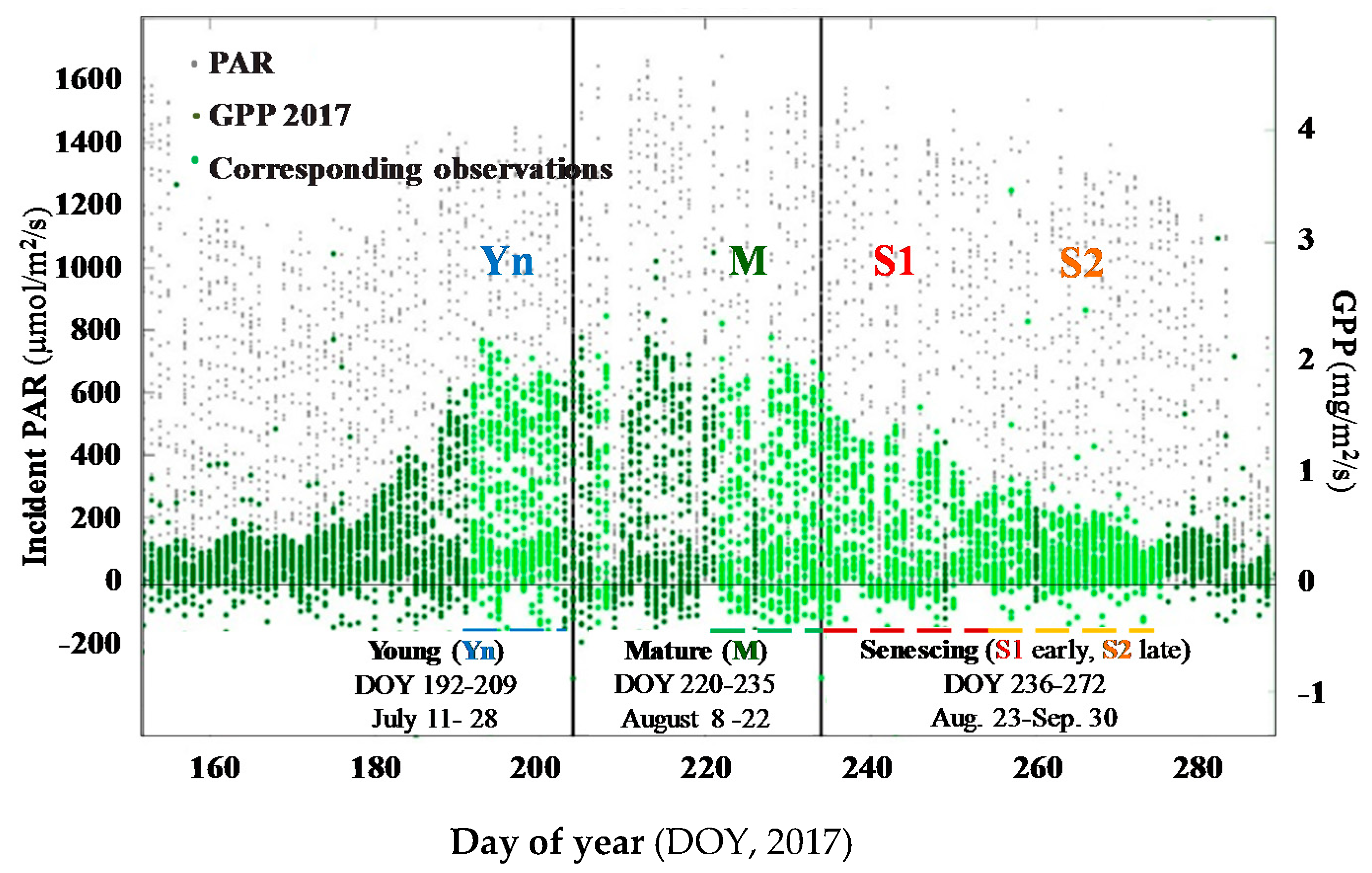
2.1. Study Site and Crop Condition
2.2. Measurements
2.3. Data Processing and Analysis
3. Results
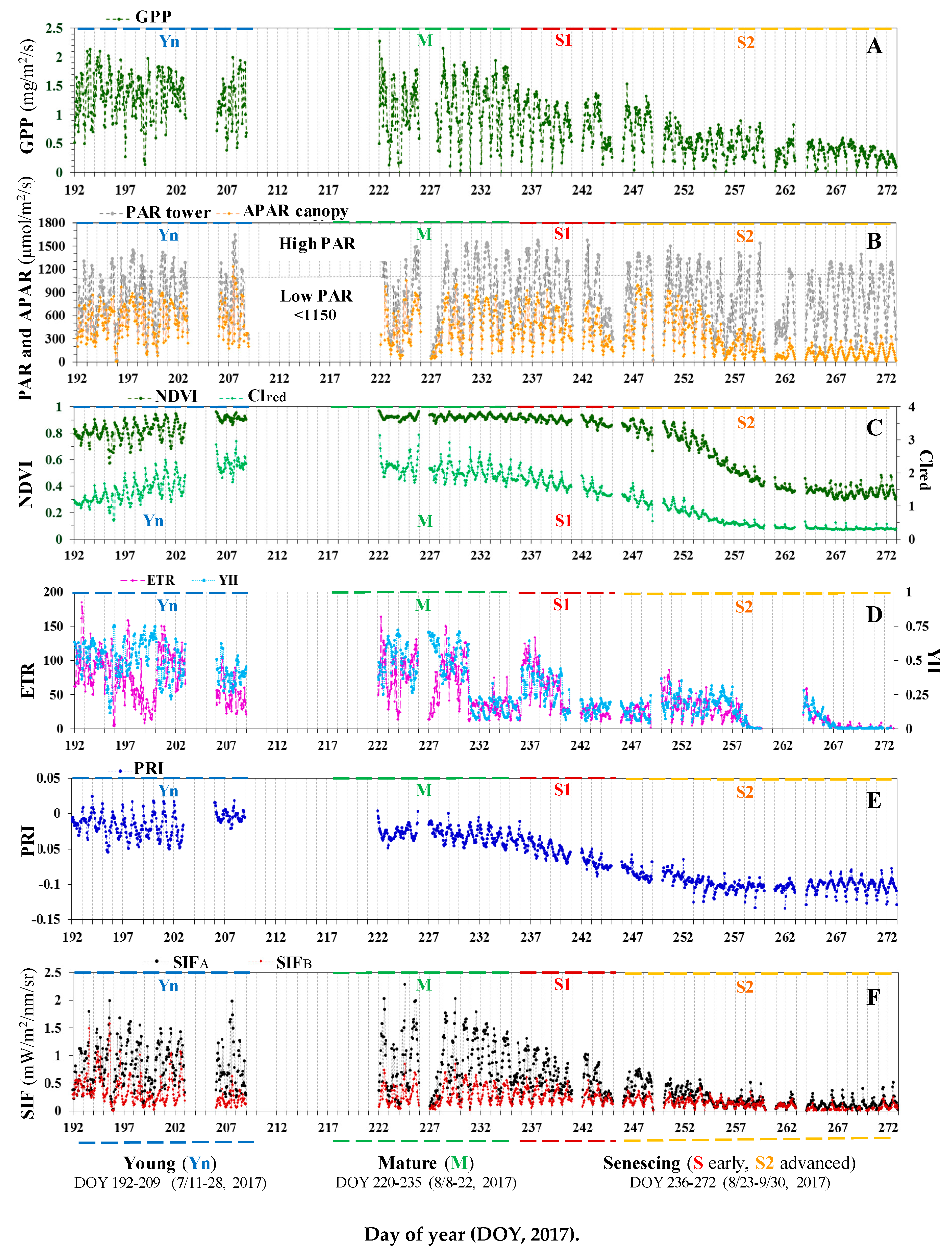
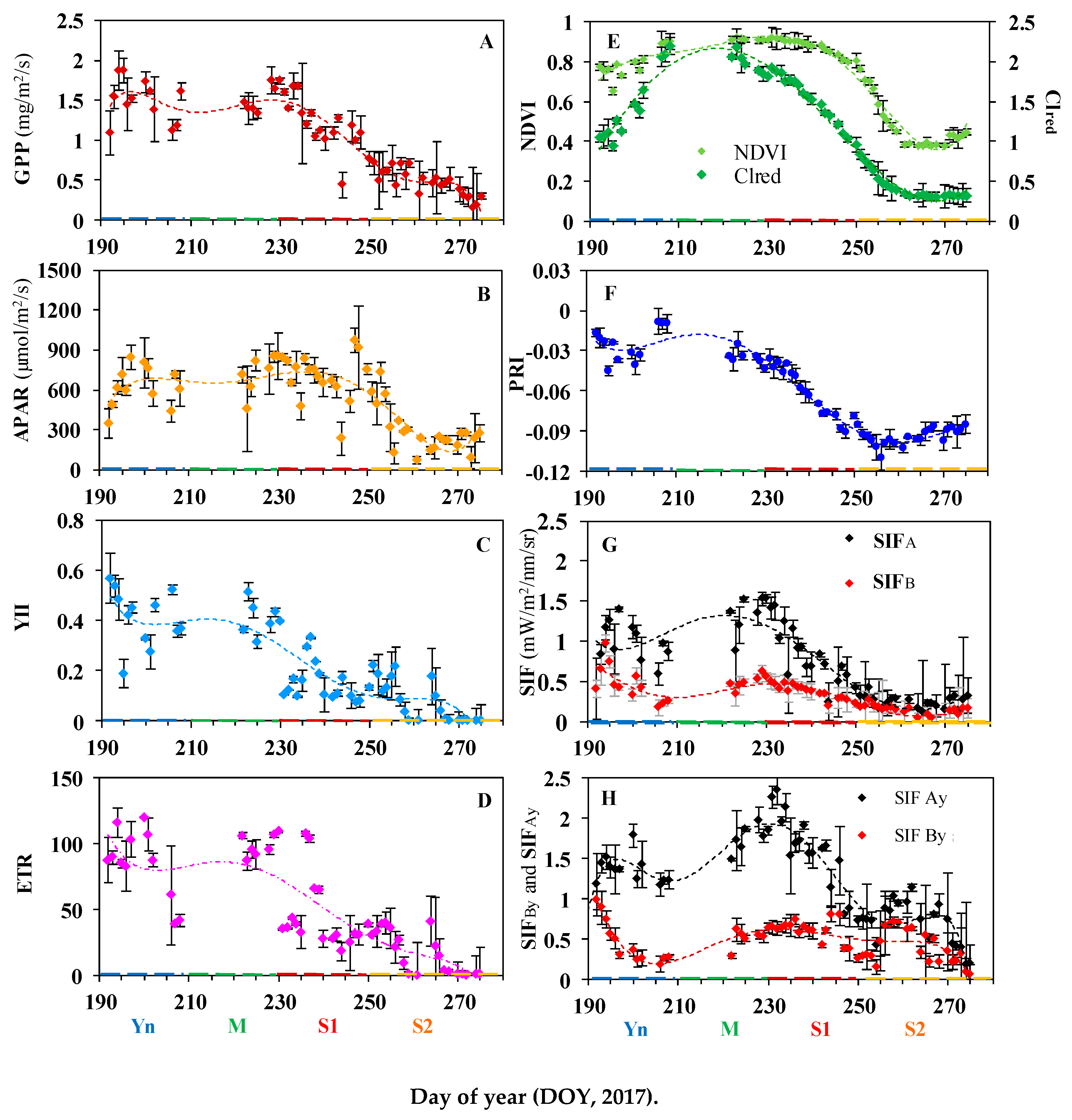
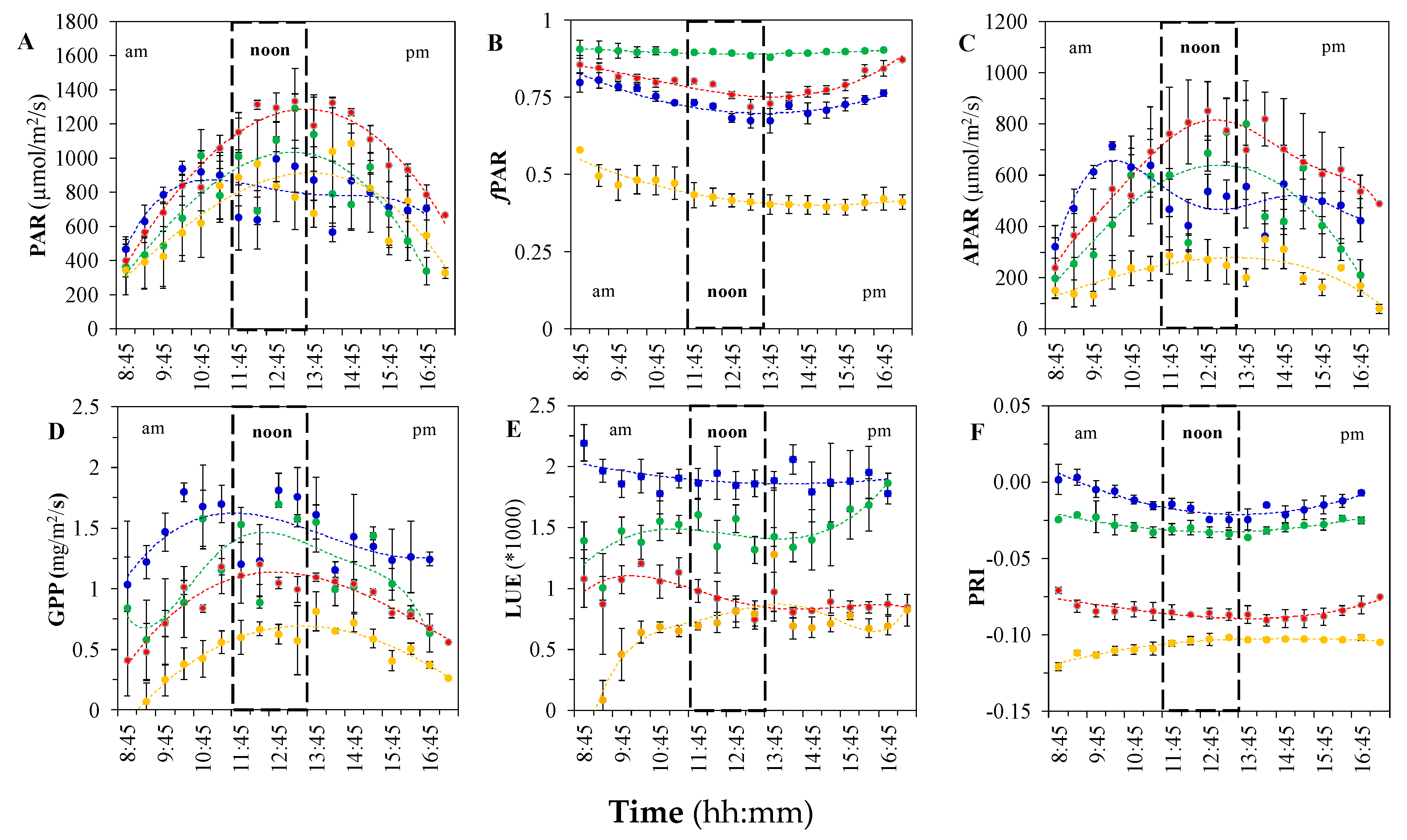
3.1. Diurnal and Seasonal Variations in Photosynthesis and the Associated Reflectance and Fluorescence Metrics
3.1.1. Variation in GPP, PAR, and LUE
3.1.2. Variation in Reflectance VIs
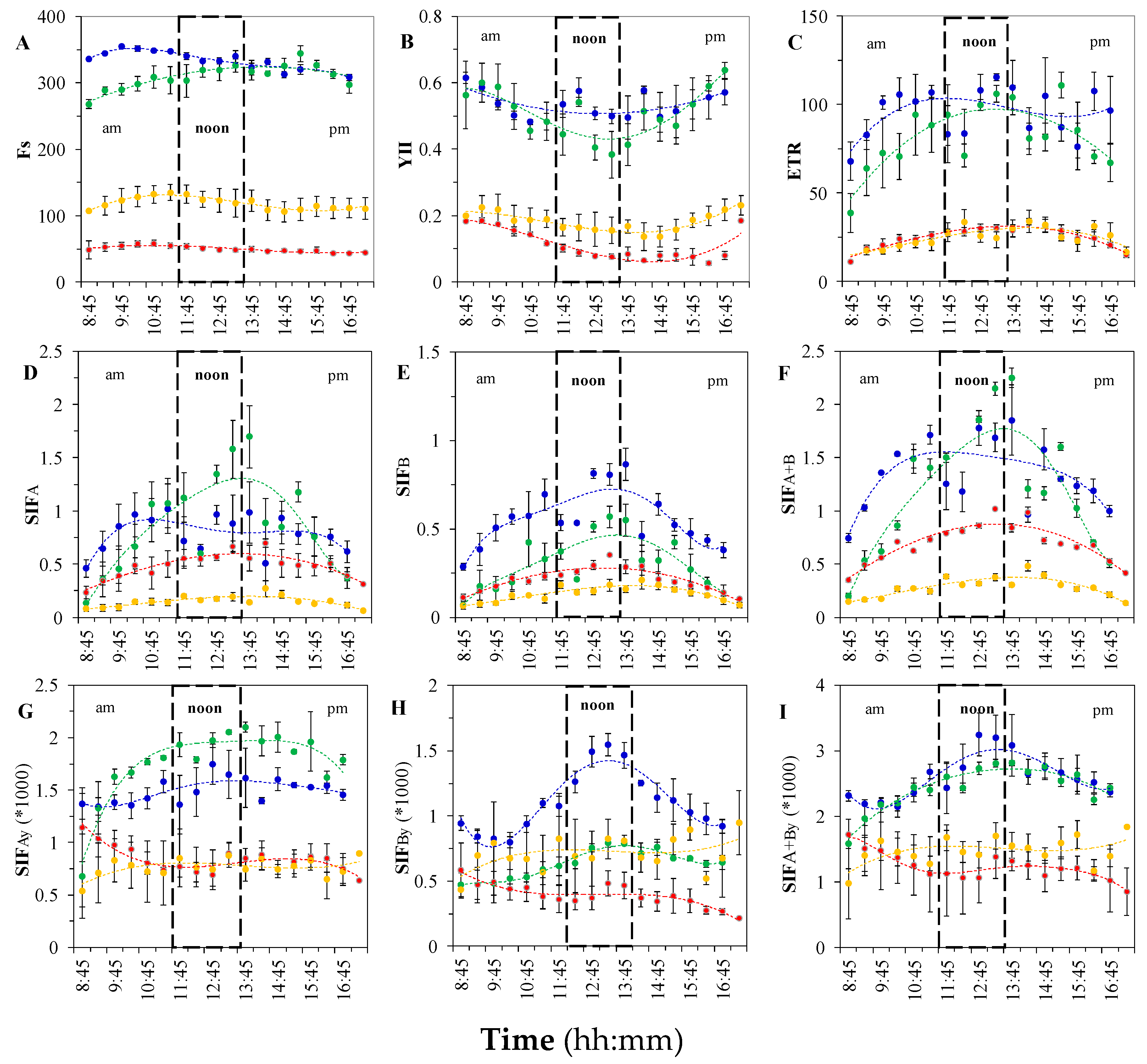
3.1.3. Variation in Leaf-level Chlorophyll Fluorescence
3.1.4. Variation in Canopy SIF Metrics
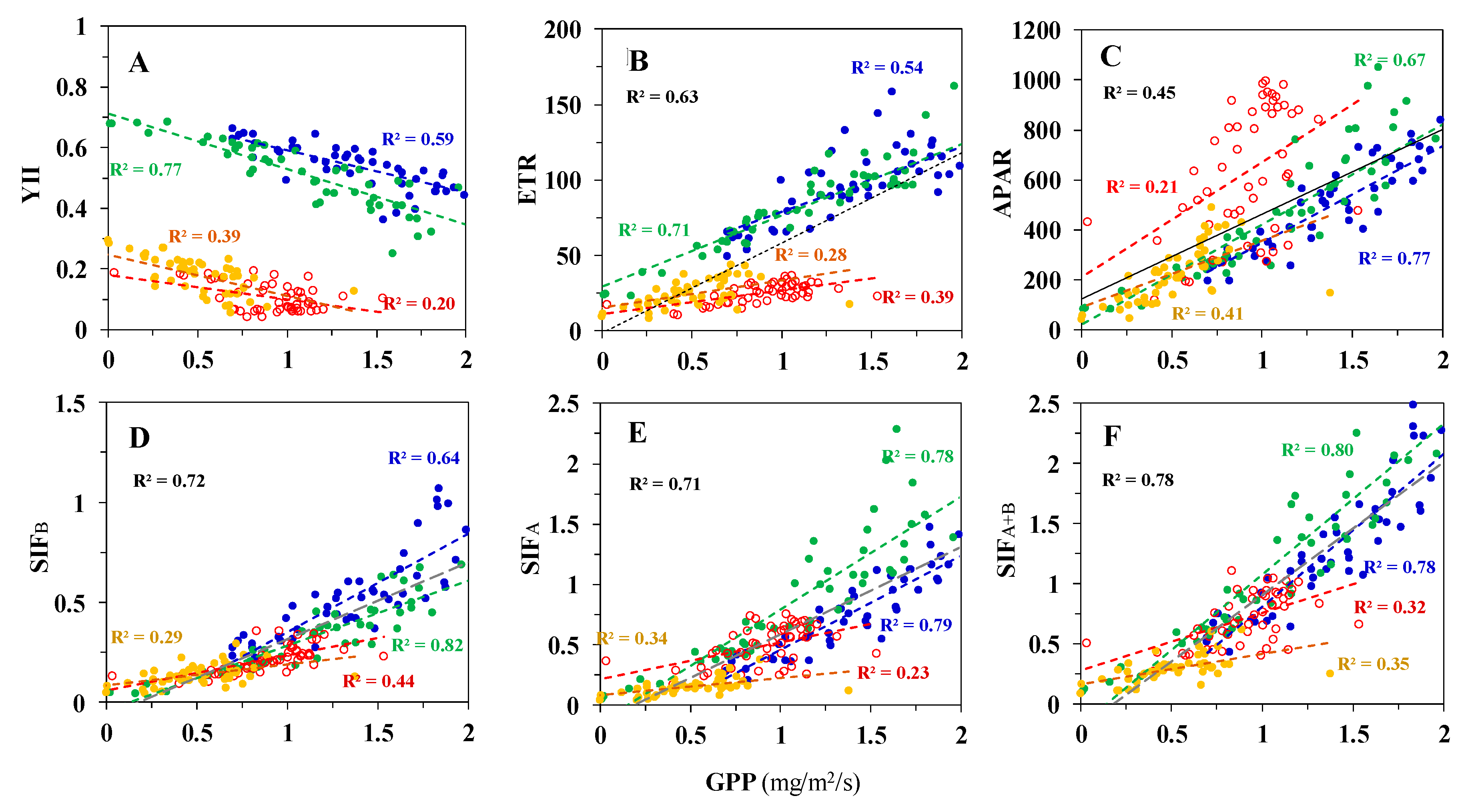
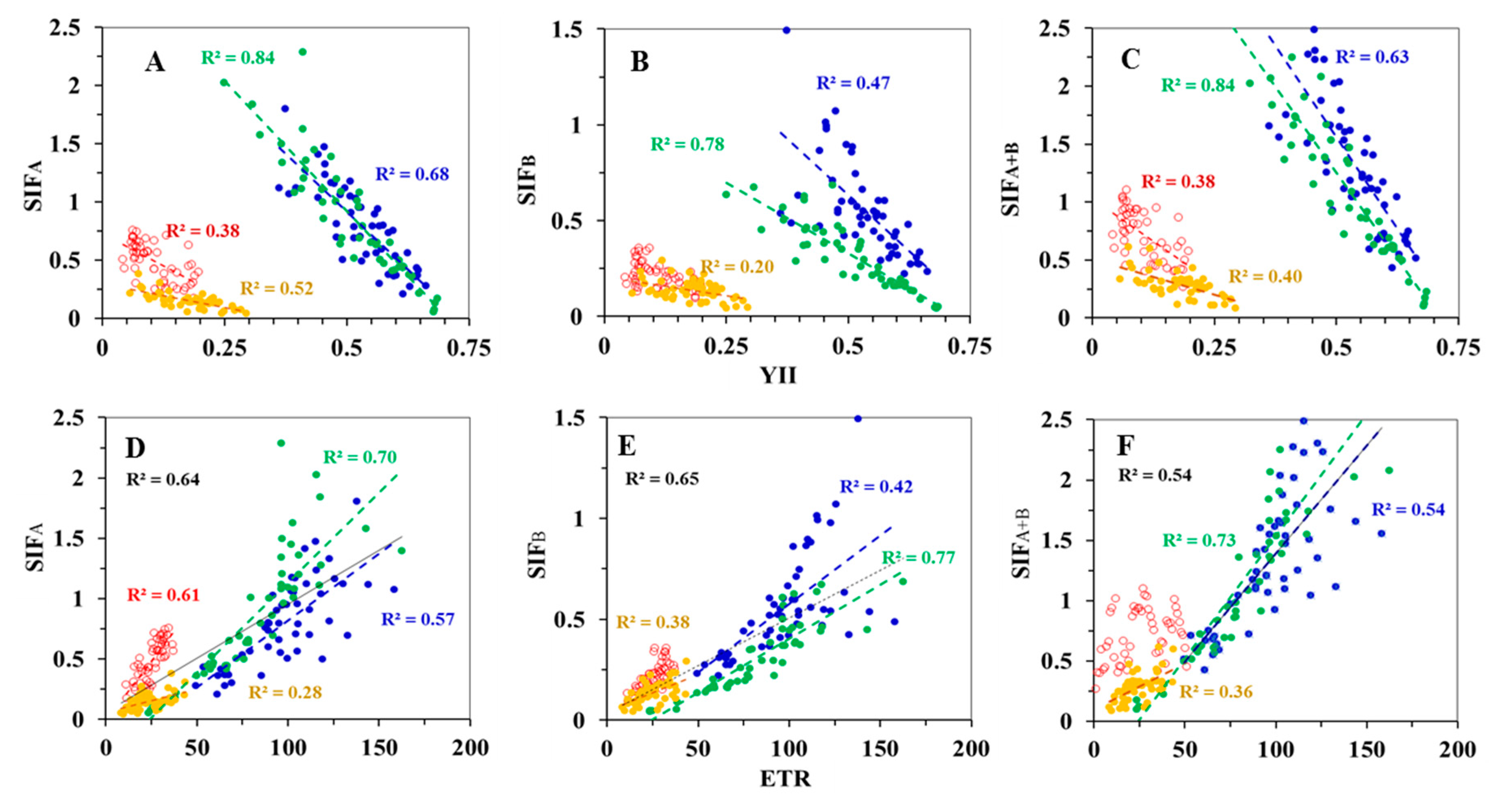
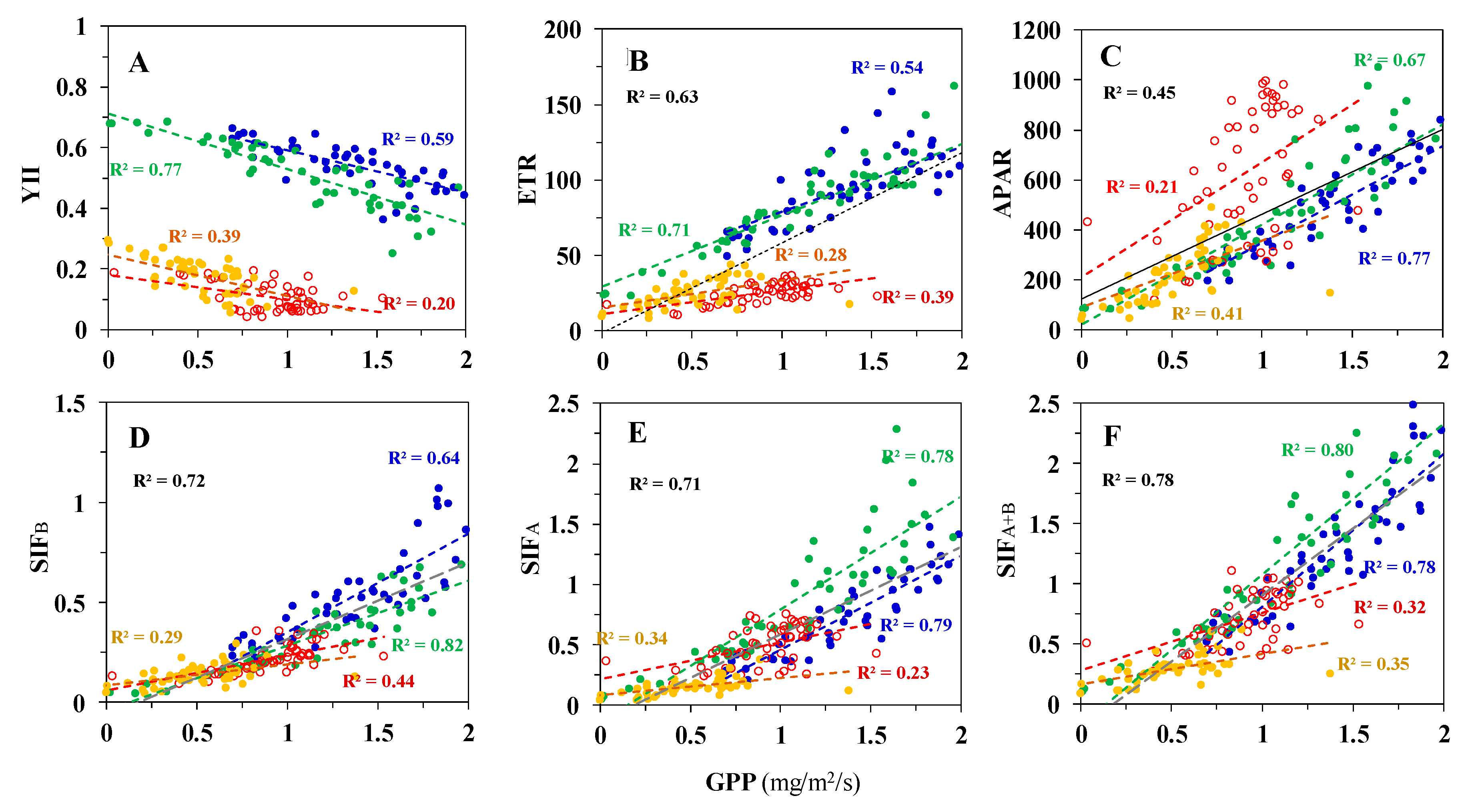
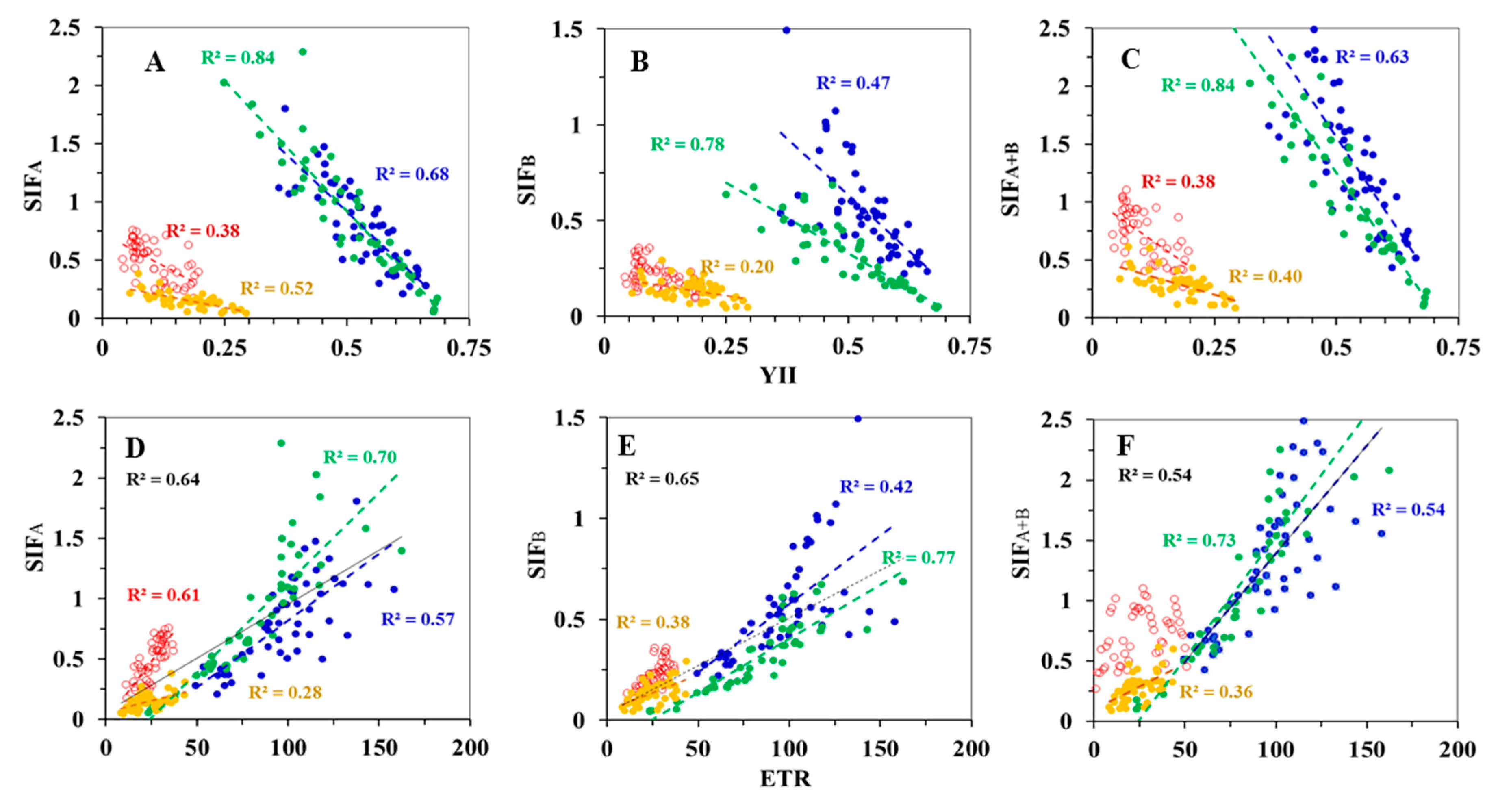
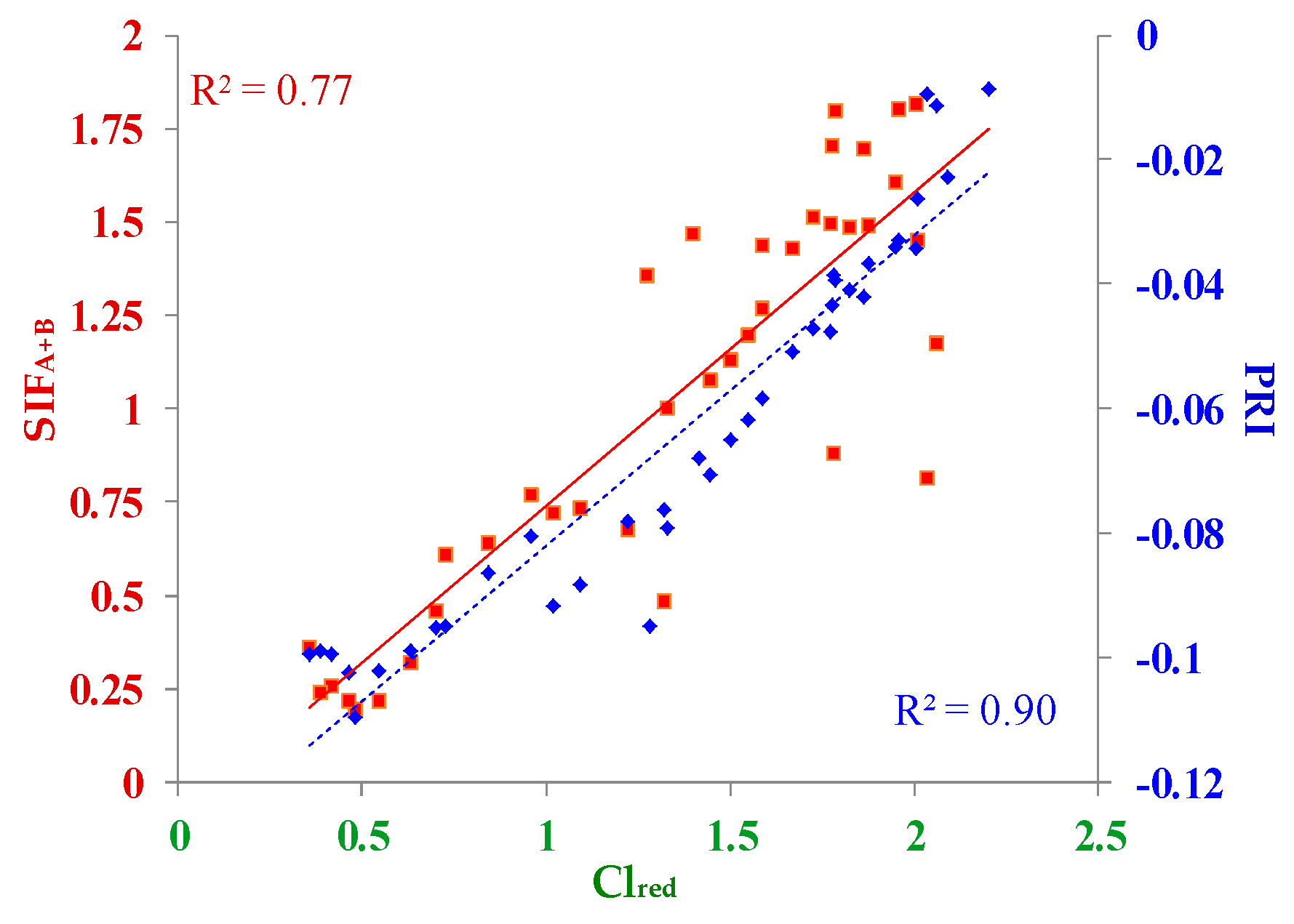
3.2. Correlations between Diurnal Sets of Observations across Corn Growth Stages
3.3. Correlations between Observations Collected under Different Conditions for Photosynthesis
3.3.1. Low PAR Conditions Including the Morning, Midday, and Afternoon Time Periods
Correlations to GPP
Correlations to LUE
3.3.2. High Light Conditions (i.e., High PAR) at Midday
Correlations to GPP
Correlations to LUE
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology, 3rd ed.; Springer: Berlin, Germany, 2003. [Google Scholar]
- Gitelson, A.A.; Buschmann, C.; Lichtenthaler, H.K. Leaf chlorophyll fluorescence corrected for re-absorption by means of absorption and reflectance measurements. J. Plant Physiol. 1998, 152, 283–296. [Google Scholar] [CrossRef]
- Gitelson, A.; Solovchenko, A. Non-invasive Quantification of Foliar Pigments: Principles and Implementation. In Hyperspectral Remote Sensing of Vegetation, 2nd ed.; Thenkabail, P.S., Lyon, J.G., Huete, A., Eds.; CRC Press-Taylor and Francis Group: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2018; Chapter 5; Volume II, pp. 135–162. [Google Scholar]
- Peng, Y.; Nguy-Robertson, A.; Arkebauer, T.; Gitelson, A.A. Assessment of Canopy Chlorophyll Content Retrieval in Maize and Soybean: Implications of Hysteresis on the Development of Generic Algorithms. Remote Sens. 2017, 9, 226. [Google Scholar] [CrossRef]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. In Third Earth Resources Technology Satellite-1 Symposium; Freden, S.C., Mercanti, E.P., Becker, M., Eds.; Technical Presentations, NASA SP-351; NASA: Washington, DC, USA, 1974; Volume 1, pp. 309–317. [Google Scholar]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A Narrow-Waveband Spectral Index That Tracks Diurnal Changes in Photosynthetic Efficiency. Remote Sens. Environ. 1992, 44, 35–44. [Google Scholar] [CrossRef]
- Hunt, E.R.; Doraiswamy, P.C., Jr.; McMurtrey, J.E.; Daughtry, C.S.T.; Perry, E.M.; Akhmedov, B. A visible band index for remote sensing leaf chlorophyll content at the canopy scale. Int. J. Appl. Earth Obs. Geoinf. 2013, 21, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Filella, I.; Gamon, J.A. Assessment of photosynthetic radiation-use efficiency with spectral reflectance. New Phytol. 1995, 131, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The photochemical reflectance index: An optical indicator of photosynthetic radiation-use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.M.; Huemmrich, K.F.; Landis, D.; Black, T.A.; Barr, A.; McCaughey, J.H. Remote sensing of ecosystem light use efficiency using MODIS. Remote Sens. Environ. 2016, 187, 345–366. [Google Scholar] [CrossRef]
- Ranson, K.J.; Daughtry, C.S.T.; Biehl, L.L. Sun angle, view angle, and background effects on spectral response of simulated balsam fir canopies. Photogrammic Eng. Remote Sens. 1986, 52, 649–658. [Google Scholar]
- Zarco-Tejada, P.J.; González-Dugo, V.; Williams, L.E.; Suárez, L.; Berni, J.; Goldhamer, D.; Fereres, E. A PRI-based water stress index combining structural and chlorophyll effects: Assessment using diurnal narrow-band airborne imagery and the CWSI thermal index. Remote Sens. Environ. 2013, 138, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Lichtenhaler, H.K.; Buschmann, C.; Rinderle, U.; Shmuck, G. Application of chlorophyll fluorescence in ecophysiology. Radiat. Environ. Biophys. 1986, 25, 297–308. [Google Scholar] [CrossRef]
- Jones, H.G.; Vaughan, R.A. Remote Sensing of Vegetation: Principles, Techniques and Applications; Oxford University Press Inc.: New York, NY, USA, 2011; ISBN 978-0-19-920779-4. [Google Scholar]
- Van Wittenberghe, S.; Alonso, L.; Verrelst, J.J. Bidirectional sun-induced chlorophyll fluorescence emission is influenced by leaf structure and light scattering properties—A bottom-up approach. Remote Sens. Environ. 2015, 158, 169–179. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence as a tool in plant physiology. Photosynth. Res. 1984, 5, 139–157. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Jee, G. Advances in Photosynthesis and Respiration, Volume 19: Chlorophyll a fluorescence: A signature of photosynthesis. Photosynth. Res. 2005, 83, 101–105. [Google Scholar]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Franck, F.; Juneau, P.; Popovic, R. Resolution of the Photosystem I and Photosystem II contributions to chlorophyll fluorescence of intact leaves at room temperature. Biochim. Biophys. Acta 2002, 1556, 246. [Google Scholar] [CrossRef]
- Vilfan, N.; van der Tol, C.; Muller, O.; Rascher, U.; Verhoef, W. Fluspect-B: A model for leaf fluorescence, reflectance and transmittance spectra. Remote Sens. Environ. 2016, 186, 596–615. [Google Scholar] [CrossRef]
- Porcar-Castel, A.; Pfundel, E.; Korhonen, J.F.J.; Juurola, E. A new monitoring PAM fluorimeter (MONI-PAM) to study the short- and long-term acclimation of photosystem II in field conditions. Photosynth. Res. 2008, 96, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Baldacci, D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: Past, present and future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Cheng, Y.-B.; Middleton, E.M.; Zhang, Q.; Huemmrich, K.F.; Campbell, P.K.E.; Corp, L.A.; Cook, B.D.; Kustas, W.P.; Daughtry, C.S.T. Integrating Solar Induced Fluorescence and the Photochemical Reflectance Index for Estimating Gross Primary Production in a Cornfield. Remote Sens. 2013, 5, 6857–6879. [Google Scholar] [CrossRef] [Green Version]
- Damm, A.; Elbers, J.; Gioli, B.; Hamidi, K.; Hutjes, R.; Kosvancova, M.; Meroni, M.; Migleta, F.; Moersch, A.; Moreno, J.; et al. Remote sensing of sun-induced fluorescence to improve modeling of diurnal courses of gross primary production (GPP). Glob. Chang. Biol. 2010, 16, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Rossini, M.; Nedbal, L.; Guanter, L.; A’c, A.; Alonso, L.; Burkart, A.; Cogliati, S.; Colombo, R.; Damm, A.; Drusch, M.; et al. Red and far red Sun-induced chlorophyll fluorescence as a measure of plant photosynthesis. Geophys. Res. Lett. 2015, 42, 1632–1639. [Google Scholar] [CrossRef] [Green Version]
- Schickling, A.; Matveeva, M.; Damm, A.; Schween, J.; Wahner, A.; Graf, A.; Crewell, S.; Rascher, U. Combining Sun-induced chlorophyll fluorescence and photochemical reflectance index improves diurnal modeling of gross primary productivity. Remote Sens. 2016, 8, 574. [Google Scholar] [CrossRef]
- Middleton, E.M.; Rascher, U.; Corp, L.A.; Huemmrich, K.F.; Cook, B.D.; Noormets, A.; Schickling, A.; Pinto, F.; Alonso, L.; Damm, A.; et al. The 2013 FLEX - US Airborne Campaign at the Parker Tract Loblolly Pine Plantation in North Carolina, USA. Remote Sens. 2017, 9, 612. [Google Scholar] [CrossRef]
- Colombo, R.; Celesti, M.; Bianchi, R.; Campbell, P.; Cogliati, S.; Cook, B.; Corp, L.; Damm, A.; Domec, J.; Guanter, L.; et al. Variability of sun-induced chlorophyll fluorescence according to stand age-related processes in a managed loblolly pine forest. Glob. Chang. Biol. 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, C.; Fisher, J.B.; Worden, J.; Badgley, G.; Saatchi, S.S.; Lee, J.E.; Toon, G.C.; Butz, A.; Jung, M.; Kuze, A. New global observations of the terrestrial carbon cycle from GOSAT: Patterns of plant fluorescence with gross primary productivity. Geophys. Res. Lett. 2011, 38, 1–6. [Google Scholar] [CrossRef]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Middleton, E.M. First observations of global and seasonal terrestrial chlorophyll fluorescence from space. Biogeosciences 2011, 8, 637–651. [Google Scholar] [CrossRef] [Green Version]
- Joiner, J.; Yoshida, Y.; Vasilkov, A.P.; Schaefer, K.; Jung, M.; Guanter, L.; Zhang, Y.; Garrity, S.; Middleton, E.M.; Huemmrich, K.F. The seasonal cycle of satellite chlorophyll fluorescence observations and its relationship to vegetation phenology and ecosystem atmosphere carbon exchange. Remote Sens. Environ. 2014, 152, 375–391. [Google Scholar] [CrossRef] [Green Version]
- Joiner, J.; Yoshida, Y.; Guanter, L.; Middleton, E.M. New methods for the retrieval of chlorophyll red fluorescence from hyperspectral satellite instruments: Simulations and application to GOME-2 and SCIAMACHY. Atmos. Meas. Tech. 2016, 9. [Google Scholar] [CrossRef]
- Sanders, A.F.; Verstraeten, W.W.; Kooreman, M.L.; van Leth, T.C.; Beringer, J.; Joiner, J. Spaceborne sun-induced vegetation fluorescence time series from 2007 to 2015 evaluated with Australian flux tower measurements. Remote Sens. 2016, 8, 895. [Google Scholar] [CrossRef]
- Wood, J.D.; Griffis, T.J.; Baker, J.M.; Frankenberg, C.; Verma, M.; Yuen, K. Multiscale analyses of solar-induced florescence and gross primary production. Geophys. Res. Lett. 2017, 44, 533–541. [Google Scholar] [CrossRef]
- Amoros-Lopez, J.; Gomez-Chova, L.; Vila-Frances, J.; Alonso, L.; Calpe, J.; Moreno, J.; del Valle-Tascon, S. Evaluation of remote sensing of vegetation fluorescence by the analysis of diurnal cycles. Int. J. Remote Sens. 2008, 29, 5423–5436. [Google Scholar] [CrossRef]
- Campbell, P.K.E.; Middleton, E.M.; Corp, L.A.; Kim, M.S. Contribution of chlorophyll fluorescence to the apparent vegetation reflectance. Sci. Total Environ. 2008, 404, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.K.E.; Middleton, E.M.; Corp, L.A.; van der Tol, C.; Huemmrich, K.F.; Cendrero-Mateo, M.P.; Leifeld, J. Diurnal and phenological changes in vegetation fluorescence and reflectance, indicative of vegetation photosynthetic properties and function. In Proceedings of the 5th International Workshop on Remote Sensing of Vegetation Fluorescence, Paris, France, 22–24 April 2014; p. 9. [Google Scholar]
- Meroni, M.; Rossini, M.; Guanter, L.; Alonso, L.; Rascher, U.; Colombo, R.; Moreno, J. Remote sensing of solar-induced chlorophyll fluorescence: Review of methods and applications. Remote Sens. Environ. 2009, 113, 2037–2051. [Google Scholar] [CrossRef]
- Wyber, R.; Malenovský, Z.; Ashcroft, M.B.; Osmond, B.; Robinson, S.A. Do daily and seasonal trends in leaf solar induced fluorescence reflect changes in photosynthesis, growth or light exposure? Remote Sens. 2017, 9, 604. [Google Scholar] [CrossRef]
- Alonso, L.; Van Wittenberghe, S.; Amorós-López, J.; Vila-Francés, J.; Gómez-Chova, L.; Moreno, J. Diurnal cycle relationships between passive fluorescence, PRI and NPQ of vegetation in a controlled stress experiment. Remote Sens. 2017, 9, 770. [Google Scholar] [CrossRef]
- Yang, H.; Yang, X.; Zhang, Y.; Heskel, M.A.; Lu, X.; Munger, J.W. Chlorophyll fluorescence tracks seasonal variations of photosynthesis from leaf to canopy in a temperate forest. Glob. Chang. Biol. 2017, 23, 2874–2886. [Google Scholar] [CrossRef] [PubMed]
- Goulas, Y.; Fournier, A.; Daumard, F.; Champagne, S.; Ounis, A.; Marloie, O. Gross Primary Production of a Wheat Canopy Relates Stronger to Far Red Than to Red Solar-Induced Chlorophyll Fluorescence. Remote Sens. 2017, 9, 97. [Google Scholar] [CrossRef]
- Middleton, E.M.; Corp, L.A.; Cook, B. FUSION Canopy Tower System for Remote Sensing Observations of Terrestrial Ecosystems. 2013. Available online: ftp://fusionftp.gsfc.nasa.gov/FUSION/ (accessed on 25 February 2019).
- Porcar-Castell, A.; Tyystjärvi, E.; Atherton, J.; van der Tol, C.; Flexas, J.; Pfündel, E.E.; Moreno, J.; Frankenberg, C.; Berry, J.A. Linking chlorophyll a fluorescence to photosynthesis for remote sensing applications: Mechanisms and challenges. J. Exp. Bot. 2014, 65, 4065–4095. [Google Scholar] [CrossRef] [PubMed]
- Cogliati, S.; Rossini, M.; Julitta, T.; Meroni, M.; Schickling, A.; Burkart, A.; Pinto, F.; Rascher, U.; Colombo, R. Continuous and long-term measurements of reflectance and sun-induced chlorophyll fluorescence by using novel automated field spectroscopy systems. Remote Sens. Environ. 2015, 164, 270–281. [Google Scholar] [CrossRef]
- Damm, A.; Guanter, L.; Verhoef, W.; Schläpfer, D.; Garbari, S.; Schaepman, M.E. Impact of varying irradiance on vegetation indices and chlorophyll fluorescence derived from spectroscopy data. Remote Sens. Environ. 2015, 156, 202–215. [Google Scholar] [CrossRef]
- Liu, L.; Guan, L.; Liu, X. Directly estimating diurnal changes in GPP for C3 and C4 crops using far-red sun-induced chlorophyll fluorescence. Agric. For. Meteorol. 2017, 232, 1–9. [Google Scholar] [CrossRef]
- Springer, K.R.; Wang, R.; Gamon, J.A. Parallel Seasonal Patterns of Photosynthesis, Fluorescence, and Reflectance Indices in Boreal Trees. Remote Sens. 2017, 9, 691. [Google Scholar] [CrossRef]
- Drusch, M.; Moreno, J.; Del Bello, U.; Franco, R.; Goulas, Y.; Huth, A.; Kraft, S.; Middleton, E.; Miglietta, F.; Mohammed, G.; et al. The FLuorescence EXplorer Mission Concept-ESA’s Earth Explorer 8. IEEE Trans. Geosci. Remote Sens. 2017, 55, 1273–1284. [Google Scholar] [CrossRef]
- PhenoCam—An Ecosystem Phenology Web Camera Network. Available online: https://phenocam.sr.unh.edu/webcam/ (accessed on 20 December 2018).
- FLUXNET 2015, FLUXNET-ORNL. Hosted at the Oak Ridge National Laboratory, USA. Available online: URL https://fluxnet.ornl.gov/) (accessed on 10 December 2018).
- Julitta, T.; Burkart, A.; Rossini, M.; Schickling, A.; Colombo, R.; Rascher, U.; Cogliati, S.M. FLoX: A System for Automatic Long Term Measurements of Top of Canopy Sun Induced Chlorophyll Fluorescence. In FLEX 2017 Workshop, ESA-ESRIN. ESA: FLEX 2017, Frascati, Italy. Available online: https://www.dropbox.com/s/w8umv5j9nvk5p5w/Abstract-book.pdf?dl=0 (accessed on 25 February 2019).
- Houborg, R.; Anderson, M.C.; Daughtry, C.S.T.; Kustas, W.P.; Rodell, M. Using leaf chlorophyll to parameterize light-use-efficiency within a thermal-based carbon, water and energy exchange model. Remote Sens. Environ. 2011, 115, 1694–1705. [Google Scholar] [CrossRef]
- Cook, B.D.; Davis, K.J.; Wang, W.; Desai, A.; Berger, B.W.; Teclaw, R.M.; Martinm, J.G.; Bolstad, P.; Bawkin, P.S.; Yi, C.; et al. Carbon exchange and venting anomalies in an upland deciduous forest in northern Wisconsin, USA. Agric. For. Meteorol. 2004, 126, 271–295. [Google Scholar] [CrossRef]
- Nichol, C.J.; Huemmrich, K.F.; Black, T.A.; Jarvis, P.G.; Walthall, C.L.; Grace, J. Remote sensing of photosynthetic-light-use efficiency of boreal forest. Agric. For. Meteorol. 2000, 101, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A. Recent Developments in Remote Estimation of Crop Biophysical and Biochemical Properties at Various Scales. In Hyperspectral Remote Sensing of Vegetation, 2nd ed.; Thenkabail, P.S., Lyon, J.G., Huete, A., Eds.; CRC Press—Taylor and Francis Group: Boca Raton, FL, USA; London, UK; New York, NY, USA, 2018; Volume 3, pp. 3–24. [Google Scholar]
- Julitta, T.; Wutzler, T.; Rossini, M.; Colombo, R.; Cogliati, S.; Meroni, M.; Burkart, A.; Migliavacca, M. An R Package for Field Spectroscopy: From System Characterization to Sun-Induced Chlorophyll Fluorescence Retrieval; ESA ESRIN: Frascati, Rome, Italy, 2017. [Google Scholar]
- Julitta, T. FieldSpectroscopy CC and FieldSpectroscopy DP Packages. Online on GitHub Platform. 2017. Available online: https://github.com/tommasojulitta (accessed on 24 January 2019).
- Cogliati, S.; Verhoef, W.; Kraft, S.; Sabater, N.; Alonso, L.; Vicent, J.; Colombo, R. Retrieval of sun-induced fluorescence using advanced spectral fitting methods. Remote Sens. Environ. 2015, 169, 344–357. [Google Scholar] [CrossRef]
- Goward, S.N.; Huemmrich, K.F. Vegetation canopy PAR absorptance and the normalized difference vegetation index: An assessment using the SAIL model. Remote Sens. Environ. 1992, 39, 119–140. [Google Scholar] [CrossRef]
- Hipps, L.E.; Asrar, G.; Kanemasu, E. Assessing the interception of photosynthetically active radiation in winter wheat. Agric. Meteorol. 1983, 28, 253–259. [Google Scholar] [CrossRef]
- Orfanidis, S.J. Introduction to Signal Processing; Prentice Hall: Englewood Cliffs, NJ, USA, 1996. [Google Scholar]
- Miao, G.; Guan, K.; Yang, X.; Bernacchi, C.J.; Berry, J.A.; DeLucia, E.H.; Wu, J.; Moore, C.E.; Meacham, K.; Cai, Y.; et al. Sun-Induced Chlorophyll Fluorescence, Photosynthesis, and Light Use Efficiency of a Soybean Field from Seasonally Continuous Measurements. J. Geophys. Res. Biogeosci. 2018, 123, 610–623. [Google Scholar] [CrossRef]
- Van der Tol, C.; Verhoef, W.; Immermans, J.; Verhoef, A.; Su, Z. An integrated model of soil-canopy spectral radiance observations, photosynthesis, fluorescence, temperature and energy balance. Biogeosci. Discuss. 2009, 6, 6025–6075. [Google Scholar] [CrossRef]
- Van der Tol, C.; Berry, J.A.; Campbell, P.K.E.; Rascher, U. Models of fluorescence and photosynthesis for interpreting measurements of solar-induced chlorophyll fluorescence. J. Geophys. Res. Biogeosci. 2014, 119, 2312–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verrelst, J.; Rivera, J.P.; Alonso, L.; Moreno, J. An Automated Radiative Transfer Models Operator (ARTMO) Toolbox for Automated Retrieval of Biophysical Parameters through Model Inversion; European Geosciences Union (EGU) General Assembly: Vienna, Austria, 2011. [Google Scholar]
- ESAS; National Academies of Sciences, Engineering and Medicine. Thriving on Our Planet: A Decadal Strategy for Earth Observation from Space; The National Academies Press: Washington, DC, USA, 2018. [Google Scholar] [CrossRef]
- Guanter, L.; Kaufmann, H.; Segl, K.; Foerster, S.; Rogass, C.; Chabrillat, S.; Kuester, T.; Hollstein, A.; Rossner, G.; Chlebek, C.; et al. The EnMAP spaceborne imaging spectroscopy mission for earth observation. Remote Sens. 2015, 7, 8830–8857. [Google Scholar] [CrossRef]
- European Space Agency (ESA). Report for Mission Selection, FLEX. In ESA SP-1330/2 Volume 2; European Space Agency: Noordwijk, The Netherlands, 2015. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement Approach | Location Frequency | Parameters (units) |
|---|---|---|
| Eddy covariance | 10 m tower 30 min | Gross primary production (GPP, mg/m2/s), photosynthetically active radiation (PAR, µmol/m2/s) and absorbed PAR (APAR, µmol/m2/s), light use efficiency (LUE, mg CO2/µmol PAR) |
| MoniPAM | leaf-level 10 min | PAM light-adapted fluorescence metrics (i.e., F maximum—Fm’, and F steady state—Fs), used to estimate the operating efficiency of PSII (YII, arbitrary units or au) and electron transport rate (ETR, µmol electrons/m2/s) |
| FLoX | 1.5 m above Canopy < 2 min | Solar induced fluorescence (SIF, mW/m2/nm/sr) in the O2A (SIFA) and O2B (SIFB) bands and their sum (SIFA+B), SIF yields (SIFAy, SIFBy, SIFB+Ay), normalized difference vegetation index (NDVI, au), chlorophyll index (CIred, au), photochemical reflectance index (PRI, au) |
| Spectral Index [Band Resolution] | Reference |
|---|---|
| NDVI = (R800 − R670)/(R800 + R670) [10 nm] | Rouse et al., 1974 [7] |
| CIred = (R785/R705) − 1 [10 nm and 5 nm ] | Gitelson et al., 2018 [4] |
| PRI = (R531 − R570)/(R531 + R570) [2 nm] | Gamon et al., 1992 [11] |
| Growth stage | Young (Yn) | Mature (M) | Early Senescence (S1) | Advanced Senescence (S2) | ANOVA GLM | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time period | am | noon | pm | n3 | am | noon | pm | n | am | noon | pm | n | am | noon | pm | n | n3 | R2 | ||||||||||||
| Canopy CO2 exchange and radiation metrics(Eddy covariance flux tower) | ||||||||||||||||||||||||||||||
| GPP1 | 1.10 | b2 | 1.50 | a | 1.28 | ab | 36 | 0.74 | c | 1.40 | a | 0.84 | c | 36 | 0.49 | d | 1.09 | b | 0.66 | cd | 36 | 0.12 | e | 0.62 | cd | 0.39 | d | 36 | 144 | 0.79 †, + |
| PAR 4 | 572 | d | 812 | c | 690 | cd | 36 | 476 | d | 1,096 | b | 607 | cd | 36 | 490 | d | 1,275 | a | 794 | c | 36 | 307 | de | 865 | c | 536 | d | 36 | 144 | 0.57 †, + |
| APAR | 431 | c | 481 | c | 459 | c | 36 | 272 | d | 653 | b | 370 | d | 35 | 296 | d | 799 | a | 511 | c | 36 | 101 | e | 270 | d | 162 | de | 35 | 142 | 0.54 †, + |
| LUE (x1000) | 1.90 | a | 1.88 | a | 1.93 | a | 36 | 1.31 | b | 1.35 | b | 1.51 | b | 35 | 1.11 | c | 0.87 | d | 0.84 | d | 36 | 0.22 | e | 0.75 | d | 0.74 | d | 36 | 143 | 0.77 ‡ |
| Leaf chlorophyll fluorescence metrics(Pulse amplitude modulated F, MoniPAM) | ||||||||||||||||||||||||||||||
| YII | 0.59 | a | 0.53 | a | 0.56 | a | 36 | 0.55 | a | 0.41 | b | 0.57 | a | 35 | 0.19 | c | 0.09 | e | 0.11 | e | 35 | 0.24 | c | 0.16 | d | 0.21 | c | 35 | 141 | 0.89 †, + |
| ETR | 80.2 | b | 97.7 | a | 86.8 | ab | 36 | 52.8 | c | 95.0 | a | 76.7 | b | 35 | 15.2 | e | 28.9 | d | 20.7 | de | 35 | 13.2 | e | 28.0 | d | 23.6 | de | 34 | 140 | 0.79 †, + |
| Canopy reflectance and fluorescence metrics(FLuorescence bOX, FLoX) | ||||||||||||||||||||||||||||||
| Reflectance vegetation indices (VIs) | ||||||||||||||||||||||||||||||
| NDVI | 0.84 | b | 0.76 | d | 0.80 | c | 35 | 0.92 | a | 0.92 | a | 0.92 | a | 36 | 0.88 | b | 0.82 | c | 0.86 | b | 36 | 0.59 | e | 0.54 | f | 0.53 | f | 36 | 143 | 0.95 ‡ |
| Clred | 1.28 | c | 1.07 | d | 1.19 | cd | 35 | 2.33 | a | 2.06 | b | 2.29 | a | 36 | 1.38 | c | 1.11 | d | 1.27 | c | 36 | 0.51 | d | 0.48 | d | 0.49 | d | 36 | 143 | 0.94 †, + |
| PRI | 0.002 | a | −0.020 | c | −0.009 | b | 35 | −0.022 | c | −0.032 | d | −0.024 | c | 35 | −0.080 | e | −0.085 | e | −0.083 | e | 35 | −0.106 | f | −0.104 | f | −0.103 | f | 34 | 139 | 0.95 ‡ |
| Solar induced fluorescence metrics (SIF, mW/m2/nm/sr) | ||||||||||||||||||||||||||||||
| SIFA | 0.60 | c | 0.80 | b | 0.70 | bc | 36 | 0.45 | cd | 1.24 | a | 0.59 | c | 36 | 0.28 | e | 0.58 | c | 0.40 | d | 36 | 0.07 | f | 0.18 | f | 0.12 | f | 36 | 144 | 0.65 †, + |
| SIFAy (x1000) | 1.38 | c | 1.56 | b | 1.48 | bc | 36 | 1.68 | b | 1.86 | a | 1.64 | b | 35 | 1.05 | d | 0.77 | e | 0.81 | e | 35 | 0.87 | e | 0.81 | e | 0.77 | e | 35 | 141 | 0.75 + |
| SIFB | 0.36 | b | 0.75 | a | 0.42 | b | 36 | 0.12 | de | 0.43 | b | 0.19 | d | 36 | 0.12 | de | 0.28 | c | 0.14 | de | 36 | 0.06 | e | 0.16 | d | 0.11 | de | 36 | 144 | 0.79 †, + |
| SIFBy (x1000) | 0.88 | c | 1.35 | a | 0.98 | bc | 36 | 0.50 | c | 0.68 | b | 0.58 | c | 36 | 0.53 | d | 0.39 | e | 0.34 | e | 36 | 0.81 | d | 0.75 | d | 0.76 | d | 36 | 144 | 0.64 ‡ |
| SIFA+B | 0.97 | c | 1.48 | b | 1.12 | c | 36 | 0.57 | e | 1.67 | a | 0.79 | d | 35 | 0.41 | ef | 0.87 | d | 0.54 | c | 34 | 0.13 | f | 0.35 | ef | 0.22 | ef | 35 | 140 | 0.66 ‡ |
| SIFA+By (x1000) | 3.64 | c | 4.47 | a | 3.95 | b | 36 | 3.87 | b | 4.45 | a | 3.87 | b | 35 | 2.63 | d | 1.93 | e | 1.96 | e | 34 | 2.55 | d | 2.37 | de | 2.31 | de | 34 | 139 | 0.65+ |
| Variables | Morning Low PAR | Midday | Afternoon Low PAR | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| High PAR 1 | Low PAR 2 | ||||||||||||
| Independent | Dependent | R2 | n3 | p | R2 | n | p | R2 | n | p | R2 | n | p |
| GPP | Photosynthetically active radiation (PAR) and absorbed PAR (APAR) | ||||||||||||
| PAR | 0.60 | 48 | ** | 0.11 | 28 | ns | 0.16 | 19 | ns | 0.43 | 48 | * | |
| APAR | 0.79 | 48 | ** | 0.08 | 28 | ns | 0.75 | 18 | ** | 0.37 | 48 | * | |
| Leaf chlorophyll fluorescence metrics (Pulse amplitude modulated F, MoniPAM) | |||||||||||||
| YII | 0.15 | 48 | ns | 0.81 | 27 | ** | 0.63 | 18 | * | 0.17 | 48 | ns | |
| ETR | 0.75 | 48 | ** | 0.67 | 27 | * | 0.70 | 18 | ** | 0.66 | 48 | ** | |
| Canopy reflectance VIs | |||||||||||||
| NDVI | 0.22 | 48 | ns | 0.23 | 28 | ns | 0.65 | 19 | * | 0.22 | 48 | ns | |
| Clred | 0.09 | 48 | ns | 0.36 | 28 | ns | 0.60 | 19 | ** | 0.09 | 48 | ns | |
| PRI | 0.48 | 48 | ns | 0.78 | 28 | ** | 0.73 | 19 | ** | 0.45 | 48 | * | |
| Canopy solar induced fluorescence (SIF) | |||||||||||||
| SIFA | 0.86 | 48 | ** | 0.65 | 28 | ** | 0.80 | 19 | ** | 0.65 | 48 | ** | |
| SIFB | 0.68 | 47 | ** | 0.68 | 28 | ** | 0.68 | 19 | ** | 0.73 | 48 | * | |
| SIFA+B | 0.88 | 48 | ** | 0.71 | 28 | ** | 0.83 | 19 | ** | 0.75 | 48 | ** | |
| Solar induced fluorescence yield (SIFy = SIF/APAR) | |||||||||||||
| SIFAy | 0.16 | 48 | ns | 0.75 | 28 | ** | 0.74 | 18 | ** | 0.26 | 48 | ns | |
| SIFBy | 0.01 | 48 | ns | 0.62 | 28 | * | 0.05 | 18 | ns | 0.03 | 48 | ns | |
| SIFA+By | 0.12 | 48 | ns | 0.79 | 28 | ** | 0.30 | 18 | ns | 0.21 | 48 | ns | |
| LUE | Photosynthetically active radiation (PAR) and absorbed PAR (APAR) | ||||||||||||
| PAR | 0.16 | 47 | ns | 0.14 | 28 | ns | 0.14 | 18 | ns | 0.26 | 48 | ns | |
| APAR | 0.23 | 47 | ns | 0.02 | 28 | ns | 0.02 | 18 | ns | 0.19 | 48 | ns | |
| Leaf chlorophyll fluorescence metrics (Pulse amplitude modulated F, MoniPAM) | |||||||||||||
| YII | 0.40 | 48 | * | 0.79 | 27 | ** | 0.61 | 18 | * | 0.69 | 48 | ** | |
| ETR | 0.55 | 48 | * | 0.49 | 27 | * | 0.47 | 18 | ns | 0.28 | 48 | ns | |
| Canopy reflectance VIs | |||||||||||||
| NDVI | 0.29 | 48 | ns | 0.19 | 28 | ns | 0.71 | 18 | * | 0.16 | 48 | ns | |
| Clred | 0.11 | 48 | ns | 0.22 | 28 | ns | 0.66 | 18 | * | 0.14 | 48 | ns | |
| PRI | 0.66 | 48 | ** | 0.67 | 28 | * | 0.64 | 18 | ** | 0.71 | 48 | ** | |
| Canopy solar induced fluorescence (SIF, mW/m2/nm/sr) | |||||||||||||
| SIFA | 0.38 | 48 | ns | 0.35 | 28 | ns | 0.43 | 18 | ns | 0.11 | 48 | ns | |
| SIFB | 0.46 | 47 | * | 0.63 | 28 | ** | 0.41 | 18 | ns | 0.32 | 48 | ns | |
| SIFA+B | 0.45 | 47 | * | 0.51 | 28 | * | 0.46 | 18 | * | 0.18 | 48 | ns | |
| Solar induced fluorescence yield (SIFy = SIF/APAR) | |||||||||||||
| SIFAy | 0.30 | 46 | ns | 0.62 | 28 | ** | 0.71 | 18 | ** | 0.34 | 47 | ns | |
| SIFBy | 0.15 | 46 | ns | 0.70 | 28 | * | 0.17 | 18 | ns | 0.29 | 47 | ns | |
| SIFA+By | 0.29 | 46 | ns | 0.71 | 28 | ** | 0.35 | 18 | ns | 0.62 | 47 | * | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campbell, P.K.E.; Huemmrich, K.F.; Middleton, E.M.; Ward, L.A.; Julitta, T.; Daughtry, C.S.T.; Burkart, A.; Russ, A.L.; Kustas, W.P. Diurnal and Seasonal Variations in Chlorophyll Fluorescence Associated with Photosynthesis at Leaf and Canopy Scales. Remote Sens. 2019, 11, 488. https://doi.org/10.3390/rs11050488
Campbell PKE, Huemmrich KF, Middleton EM, Ward LA, Julitta T, Daughtry CST, Burkart A, Russ AL, Kustas WP. Diurnal and Seasonal Variations in Chlorophyll Fluorescence Associated with Photosynthesis at Leaf and Canopy Scales. Remote Sensing. 2019; 11(5):488. https://doi.org/10.3390/rs11050488
Chicago/Turabian StyleCampbell, Petya K. E., Karl F. Huemmrich, Elizabeth M. Middleton, Lauren A. Ward, Tommaso Julitta, Craig S. T. Daughtry, Andreas Burkart, Andrew L. Russ, and William P. Kustas. 2019. "Diurnal and Seasonal Variations in Chlorophyll Fluorescence Associated with Photosynthesis at Leaf and Canopy Scales" Remote Sensing 11, no. 5: 488. https://doi.org/10.3390/rs11050488