5 Key Challenges and Solutions for Governing Complex Adaptive (Food) Systems
by
Mollie Chapman
1,*, 
Susanna Klassen
1,2,
Maayan Kreitzman
1,
Adrian Semmelink
1,
Kelly Sharp
1,
Gerald Singh
3 and
Kai M. A. Chan
1 1
Institute for Resources, Environment and Sustainability, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
2
Centre for Sustainable Food Systems, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
3
Nippon Foundation NEREUS Program, Institute for Oceans and Fisheries, University of British Columbia, Vancouver, BC V6T 1Z4, Canada
*
Author to whom correspondence should be addressed.
Sustainability 2017, 9(9), 1594; https://doi.org/10.3390/su9091594
Submission received: 26 July 2017
/
Revised: 23 August 2017
/
Accepted: 28 August 2017
/
Published: 7 September 2017
(This article belongs to the Special Issue Environmental Governance for Sustainability)
Abstract
:There is increasing recognition in academic circles of the importance of adaptive governance for the sustainability of social-ecological systems, but little examination of specific implications for the 34% of land-use where human activities are pervasive but potentially commensurate with functioning ecosystems: agricultural production systems. In this paper, we argue for the need to view food systems and agro-ecosystems as multi-scalar complex adaptive systems and identify five key challenging characteristics of such systems: multi-causality; cumulative impacts; regime shifts; teleconnections and mismatch of scales. These characteristics are necessary features of multi-scalar adaptive systems, and apply equally to social and natural subsystems. We discuss the implications of these characteristics for agricultural production systems and consider how governance can rise to these challenges. We present five case studies that highlight these issues: pollinator declines; payments for ecosystem services; pest control and pesticide resistance; downstream aquatic systems in Tasman Bay, New Zealand; and riparian buffers in Puget Sound, USA. From these case studies we derive recommendations for managing agricultural systems, both specific and general. Ultimately, adaptive governance of agro-ecosystems will likely hinge upon three paradigm shifts: viewing farmers and ranchers not only as food producers but also as land and water managers; seeking not yield maximization but rather resilient management of food ecosystems; and critically, as it transcends the production-system literature, engaging broad audiences not only as consumers but also citizens.
1. Introduction
Agriculture represents the single largest driver of environmental impacts globally [1,2,3], and further growth in food production is on a collision course with sustainability goals, including Aichi Targets for biodiversity (e.g., Strategic Goal C: Safeguarding ecosystems) and some Sustainable Development Goals (e.g., Goal 15: Protecting life on land). Modern agricultural production systems are designed to maximize a single goal: production of food, fuel or fiber; yet we depend on agricultural lands to provide a host of other ecosystem services such as water regulation and filtration, pollination, disease suppression and carbon sequestration [1]. In conjunction with protected areas, forest fragments and riparian buffers, agricultural lands can also serve as an essential matrix for supporting biodiversity [4]. A key challenge is to improve agricultural production in response to growing demand for food, while maintaining or enhancing the biodiversity and full suite of ecosystem services provided by agricultural lands [2,5].
Food systems solutions are challenging, in part because though food systems are clearly complex adaptive social-ecological systems, they are generally managed as if they were engineered systems. The trend over human history, but particularly the past two hundred years, has been increased mechanization of agriculture, supplementing and replacing diverse functions to support biomass production that were performed by ecosystems, with more efficient yet environmentally degrading biochemical and mechanical anthropogenic inputs [6]. Thus, nutrient cycling to maintain soil fertility has been largely replaced by chemical fertilizers, pest control has largely been supplanted by synthetic pesticides, and diverse native pollinators have been occluded by domesticated honeybees. These changes have yielded problems in the form of (1) externalities such as greenhouse gas emissions and the eutrophication of downstream water bodies [7], plus the declines of amphibians [8] and various insects; and (2) vulnerabilities and on-farm failures such as colony collapse of honeybees [9] and pollination limitation [10]. Agricultural production systems operate in the face of drastic ecological and social changes (e.g., climate change, demographic changes), which make it all the more essential to approach them using a lens of complex adaptive systems (CAS) that seeks robustness over efficiency.
A crucial question is, how specifically might CAS thinking enhance the management of agricultural systems, broadly conceived to include the terrestrial, sea and freshwater systems they inhabit and impact? CAS literature has defined system characteristics in a variety of ways (such as [11,12]). We focus here on a short list of key characteristics (with an emphasis on mechanisms) that might usefully inform management of socio-ecological systems generally and agricultural production systems specifically. As well, many authors have examined the idea of resilience (related to systems thinking) in agricultural contexts (local cases as well as global concerns around food security and biodiversity) [13,14,15,16,17]. Similarly, Bennett et al. suggest a focus on experimentation, novel solutions and innovation [18]. While resilience offers one important lens, we seek to offer here a more specific set of characteristics as well as approaches to the governance of CAS, addressing the gap between global conceptual work and locally specific case studies. Agriculture can benefit from CAS ”thinking” as it has been applied to other environmental systems, such as natural resource management [19], fisheries [20], marine ecosystems (e.g., [21]), conservation planning (e.g., [22]), tourism [23], and rangelands [24]; and adaptive management applied to agricultural innovation [25]. To consider more broadly the lessons of CAS for agriculture, we have distilled from these other literatures five characteristics that are helpful to understand change in social-ecological systems. (1) Multi-causality: any given pattern may be caused by several different processes, and the action of each is dependent on context [26]. (2) Cumulative impacts: processes do not operate separately but rather accumulate and interact (e.g., [27]). (3) Non-linearity: social and ecological patterns do not gradually change as a linear function of relevant processes but rather display thresholds, time lags, and generally complex behavior (including regime shifts) [28,29,30]. (4) Teleconnections: in this connected world, patterns in one location may result from processes and events in distant locations [31]. (5) Multi-scalarity: relevant processes are simultaneously operating at a diversity of scales, manifesting in patterns at multiple scales [32,33].
In this paper, we discuss the ways in which agricultural systems operate as CAS, considering each of the five factors in turn with a case study for each (selected to effectively highlight each characteristic and cover a range of key issues for managing agricultural production systems, though not a representative or comprehensive set of CAS dynamics in such systems. The five factors are summarized in Table 1 and their relation to each case study is summarized in Table 2). We close with policy options and potential paradigm shifts to enable food systems governance for the Anthropocene. While generally we consider SES where farmers are an integral component, in some sections of the paper the CAS framing is used in keeping with ecological literatures that externalize the role of people and focus on the role of ecological sub-systems. Furthermore, most of the article follows norms in applied ecology literatures (including in agro-ecosystems) which treat consumers and supply chains as external. However, we recognize that these other components are actually crucial and treat those considerations in the final section. If we are to effectively manage agricultural production systems, we will need a clear understanding of how these systems operate and how they might be governed more effectively.
2. Characteristics of Complex Adaptive (Food) Systems
2.1. Multi-Causality: There Is No Smoking Gun and No Silver Bullet
Any given ecological or social pattern is simultaneously the product of many different processes [26]. For example, the distribution and variable abundance of an insectivorous bird is simultaneously a function of the habitat selection of individual birds—which itself results from a bird’s observation of available prey, predation risk (to hawks, etc., as well as nest predation), and nest resources—but also of their relative reproductive success relating to a variety of factors. Seeking to attribute a given pattern (or change in pattern) to a single factor is further complicated by the fact that the effect of all those factors is variable in a context-dependent way.
Context dependence occurs when the same event or process produces different results in different contexts. This can be exacerbated by the interactive nature of ecosystems where changes to one part can ripple through food webs, or ricochet between biotic and abiotic factors, or even between social and ecological subsystems. For example, the effect of a loss of insectivorous birds depends on several interrelated factors, including (1) whether those birds will prey more upon predatory insects than on the pests themselves, (2) the abundance of predatory insects, (3) the prevalence of pesticide applications, (4) the presence of hedgerows, and proximity to natural habitats, etc. In the southern Indian state of Kerala, a suite of interacting factors resulted in a loss of productivity when farmers switched from locally adapted cows to the more productive Holsteins, because of their lack of resistance to the local conditions—heat, humidity and tropical diseases [45].
Interactions between different factors may not only alter the magnitude of impacts but even whether they are positive or negative. For example, forest cover can have different impacts on base water flow in a watershed in wet versus dry years [46]. In wet years the forested watershed infiltrates more water than an urbanized watershed but in dry years the forested watershed transpires more water than the urbanized one leading to a lower base water flow [46].
2.2. Cumulative Impacts: Death by a Thousand Cuts
Not only do the multiple potential causes of a given pattern confound the search for a single or primary concern, they also accumulate to produce outcomes that could not have occurred with any single concern. Thus, many authors have argued for the necessity of considering cumulative impacts on ecosystem. While some literature on cumulative impacts defines them as the drivers of change and some defines them as the effects, we use the definition following Singh et al.: “the combined total effect of multiple effects that limit the ability of people to enjoy ecosystem services” [34]. Cumulative impacts can derive from either the accumulation of many small impacts of the same kind or two or more impacts of different types. Impacts on ecosystems can accumulate (1) additively, through simple addition of several impacts, (2) synergistically, where the actual impact is greater than the sum of individual impacts, and (3) antagonistically, where impacts mitigate or subdue the combined effect so that the total impact is less than would be expected from their sum. Though there is much concern regarding synergistic impacts from researchers and policy makers, antagonistic impacts can also pose management challenges, and reducing a mitigating stressor can cause a primary stressor to degrade an ecosystem further [47]. All three types of impact interaction have been found in experiments and ecosystems. Experimental manipulation of whole ecosystems in boreal lakes showed that acidification, warming and drought generated synergistic impacts on consumers and antagonistic impacts on producers [48]. One meta-analysis of 57 experiments found 20 synergistic effects, 24 antagonistic effects and 13 additive effects, indicating that non-additive impacts are common [27].
The ecological impacts of agricultural production can contribute to cumulative impacts on aquatic food webs. For example, the die-offs of turtle grass beds in several tropical and subtropical regions illustrates the cumulative nature of agricultural impacts (sedimentation, eutrophication) and historical overfishing. Large herbivores (e.g., sea turtles and dugongs/sea cows) kept grass short. Their removal increased decomposition-fueled microbial activity, which led to hypoxia, changing the chemistry of the ecosystem. This altered ecosystem was more vulnerable to subsequent impacts (including those caused by crop or livestock production systems) such as nutrient loading, disease, sedimentation, and turbidity [49]. This system change is also an example of multi-causality as biotic and abiotic factors created interactions leading to ecosystem changes. Here, herbivory of turtle grass by the primary consumers (sea turtles) helps maintain healthy turtle grass beds.
Climate change can lead to cumulative impacts from and in agricultural production systems both directly and indirectly. Indirect climate impacts can (1) amplify or diminish local impacts. For example, climate variability interacts with land use change; in one case land use change amplified climate driven decreases in runoff and soil water while diminishing climate-driven decreases in evapotranspiration [50]. Climate change can also (2) alter conditions in such a way that exposes the system to new impacts. For example, climate change can expose agriculture to new pests and diseases [51].
2.3. Regime Shifts: Systems Change in Fits and Spurts, and Can Shift Unexpectedly
If multiple causes and cumulative impacts resulted only in linear changes to social and ecological systems, many problems would have straightforward management implications. However, systems change non-linearly. At the extreme, system shifts involve a fundamental change in system components or interactions that cannot be easily returned to the original state by removing the source of disturbance. Regime shifts between multiple stable states of an ecosystem were first established from theoretical models that combined predator-prey interactions with other random events (like disturbance) and heterogeneity of temporal and spatial scales. In his seminal paper, Holling [28] showed that predator-prey systems can have multiple “basins of attraction” (the set of initial conditions that eventually lead an ecosystem to a certain state). This contrasted to previous conceptions of ecosystems that suggested that systems had a single global equilibrium state that was dictated by “bottom up” properties like substrate and climate [38]. After the initial theorization of regime shifts, a substantial literature has accumulated documenting them in a diversity of systems including terrestrial, freshwater, and marine (reviewed in [30,52]). These reviews suggest that human disturbances lead to regime shifts, and advance the prominent theoretical interpretation that an erosion of resilience zones around a particular basin of attraction can push formerly stable systems to shift.
One challenge of regime shifts is that the threshold that will move the system from one state to another is often unknown a priori. A particular state (e.g., forest) may be robust up to some biotic or abiotic threshold (e.g., reduced moisture, grazing), but once those thresholds are exceeded, it may shift into a different state (e.g., grassland) [53]. Even returning conditions back to former levels can fail to shift the system to its original state [54]. For example, shallow lakes can be characterized by their nutrient concentrations: either nutrient-poor with an abundance of aquatic plants or nutrient-rich dominated by algae. However, even if nutrient levels are somewhat reduced in an algae dominated lake a shift may not occur as it still remains in a stable state [55]. In addition to a dramatic change from one system state to another, systems may also shift to multiple stable attractors (e.g., bifurcations) and in cases oscillate between two different stable attractors (periodic behavior) [39]. Such bifurcations and oscillations might pose extra difficulties in managing a system across scales where the smaller scale dynamics are less predictable and more variable than on the landscape scale (e.g., cycles of pest outbreaks may be more predictable on a landscape scale than on a farm scale).
Non-linearity and regime shifts can apply to agricultural production systems in two ways. On one hand, we can consider agricultural production systems as actively managed (via human inputs) to be far away from ecological equilibria in order to optimize the production of particular system components (e.g., crops). Different agricultural systems may be within the basins of attraction of different stable states (some more desirable than others), but are maintained at non-equilibrium states. Using this concept, true stable states are only reached when active management stops or fails.
An alternative perspective is that different agricultural systems (like the diversified perennial system and the intensely sprayed monoculture) are themselves alternative stable states, with shifts between the two driven by ecological or socioeconomic forces or interactions between these [17]. By this view, human management is the feedback that maintains the system. That a small change, such as no longer applying pesticides to an intensively managed crop, could result in dramatic consequences, could be seen as a factor that keeps that agricultural production system in a stable state (albeit one requiring substantial human energy and input, and vulnerable to environmental, social, and economic changes).
2.4. Teleconnections: ”Transporting” Impacts across Time and Space
Not only are systems buffeted by local forces of disturbance, but important forces of change can originate from distant areas via teleconnections—links between distant areas that are enabled via larger scale processes. Originally a concept from climate science, the teleconnections idea has been used to understand phenomena such as tropical deforestation, impacts of urban land use and vulnerability [41,42,56,57], as well as sustainability more generally [58]. Adger et al. describe three mechanisms of teleconnections: biophysical links and feedbacks; market links; and flows of resources, people and information [42].
For example, large-scale migration is a biophysical teleconnection mechanism that links multiple populations of a species, thereby maintaining a species’ presence in a lower quality habitat, including in agricultural matrices. A species can have a “source” in its preferred habitat, where birth rates exceed death rates, and then migrate to “sink” habitats as the source habitat approaches or exceeds its carrying capacity [59]. Without immigration from the source habitat, sink habitats either cannot support a species at all, or can only support a lower density. This has important implications for conservation. If priority areas for conservation are chosen based on the presence of a species rather than more detailed demographic understanding, the sites may be exclusively sink habitats, resulting in a possible extirpation of the species [59]. The obvious solution is to protect source habitats; however, they can often be difficult to determine without long-term studies [59]. By this mechanism, un-fished refuges can slow population collapse in fished areas (e.g., Jamaican reef fishes), but over time these refuges are often eliminated, leading to a sudden collapse of fisheries elsewhere [49]. Sources/sink dynamics can create desirable states of “stable maladaptation,” whereby the conditions in the source prevent the species from adapting to the sink habitat [59]. In agricultural (or aquacultural) systems, such stable maladaptation can prevent pest resistance [60].
Regional productivity serves as an economic example of teleconnections: here, an increase in global supply of commodity crops leads to price drops that harm producers of the same crop elsewhere [42]. Teleconnections linked through global markets are also integral to the idea of land sparing—the idea that intensive production might “spare” wild and natural areas elsewhere. This idea is underpinned by the assumption that due to globalized commodity markets, increasing production in one area may decrease pressure for production in another (distant or proximate) area [61]. Similarly, the “race to the bottom” narrative indicates that expanded regulations in one location may result in the transference of production to a region with lower environmental standards. This occurs when regions are engaged in international markets and the commodity can be produced elsewhere, with less rigorous standards. In a different example, demand for fair trade and organically certified coffee in rich nations can (in the right circumstances) facilitate better wages and lower pesticide use in poorer coffee-producing nations.
An example of the teleconnected flows of resources, people and information can be seen in Eakin et al.’s study of coffee production in Mexico and Vietnam. They show how coffee producers’ vulnerability “is connected not only through the structure of the global coffee commodity chain, but also through global ideological shifts affecting national policy, the movement of labor, the material flow of coffee stocks, channels of information, and, in reverse, through the broader environmental and institutional implications of local adaptive action”. (p. 399) [43]. A second example is tropical deforestation, which while context dependent, has at times been driven by teleconnected relationships to distant places. In the 1980s and 90s, rural population growth was the primary driver of tropical deforestation, but by the early 2000s it was more often associated with high rates of urban population growth and engagement in agricultural export markets [56]. The mechanisms behind these teleconnections could be demand from distant urban areas via globalized markets or regional urbanization leading to greater demand for agricultural products from rural areas [56]. Teleconnections also include impacts on urban areas such as when a rural crisis leads to rapid urban migration [41]. Rural farming communities can face major changes due to losses of traditional production livelihoods and/or inflows of urbanites seeking a rural lifestyle, often referred to as amenity migration [62]. For another example, extreme weather in the Midwest had a ripple effect of ecological, social, and economic factors. When apple crops failed, migrant farm labourers did not come to work, which led to the de-facto failure of vegetable crops that went unharvested [63].
2.5. Multi-Scalarity: Drivers and Impacts Cross Scales
Whereas teleconnections occur between disparate areas via larger scale processes, systems are also often subject to multi-scalarity, in that drivers exert impacts at multiple temporal or spatial scales. Patterns can therefore emerge at one scale via changes at other scales [26].
The importance of spatial scale can be seen in hydrological modeling for payment for ecosystem services programs. These models, using a parcel to represent an entire watershed, seek to understand the impact of land use change on the watershed and thus determine the appropriate payment for conservation. Modelers face a signal-to-noise problem as high variability across the landscape makes it difficult to detect change in one small parcel [46]. Given that year to year variability is high, it is difficult to determine that change in a small number of parcels was responsible for landscape-level changes.
Temporal scale is important for the choice of baselines and evaluation of the impacts of changes. Time lags in responses of ecological changes present a key management challenge. Even relatively small differences in time can have important consequences. Tilman et al. studied the impact of plant biodiversity on productivity relative to nitrogen addition, watering, elevated CO2, fire and herbivory [64]. During the first five years the non-biodiversity factors had a greater influence on productivity than biodiversity factors. However as the experiment continued, biodiversity became one of the most important factors: the effects of biodiversity on productivity increased over time, while the impact of other factors (especially nitrogen fertilization) decreased. From year 9 onward all three biodiversity treatments (1 versus 16 species, 2 versus 16 species and 4 versus 16 species) were more important than any of the other factors [64]. This experiment suggests that even effectively implemented land use and management policy changes may not show their effects for five or more years, and that governance systems operating over shorter timescales could disregard important dynamics.
Temporal scale is also important in the choice of baselines. When managing, restoring or assessing ecosystems, a longer historical perspective is important. In their work on fisheries from the historical record, Jackson et al. argue for the importance of a deep historical and even archaeological perspective in ecosystem management [49]. Through proxies of past species abundance and ecosystem structure (paleo-archaeological, archaeological, historical, ecological) they show that our current baselines have massively shifted from previous species abundance [49]. They showed that people have been altering ecosystems far before Europeans colonized what they assumed to be “natural” landscapes and that these impacts continue to drive even substantial contemporary consequences [49].
3. Case Studies
3.1. Searching for a Smoking Gun for Pollinator Declines
The crucial role of pollinators in our agricultural systems has been highlighted in recent years, as research increasingly shows that various key species of bees, both wild and managed, are experiencing declines [65]. Public media campaigns, a growing body of scientific evidence, and increased attention from governments have brought the health of pollinator populations into public discourse. As we depend on pollinator species for livelihoods, approximately 1/3 of the global food supply [66], and high-quality nutrition [67], scientists, policy-makers and activists have been actively searching for solutions to stem pollinator losses, which are likely to increase with climate change. How best to do so remains highly debated, stunting policy response, due in part to a lack of agreement on causes, and a problematic search—particularly by the public and media—for a single smoking gun.
Honeybees (predominantly Apis mellifera) are the primary pollinator of monoculture crops globally [66]. They serve as an integral component of a highly managed agricultural system, wherein producers pay apiarists to transport colonies to flowering crops during pollination season [68]. Declines have been documented since the 1940s, including a decrease of over 50 per cent of managed honeybees in the United States from the late 1940s to the early 21st century [69]. Alongside recent media and academic interest surrounding honeybee populations, emerging research points to the importance of native pollinator species in the agricultural landscape. Wild species have been proven to increase crop yields, and are an ecologically important and economically significant pollinator [10,70,71,72,73]. Their role is increasingly critical as honeybee populations decline.
Declines in bee numbers, particularly honeybees, are often attributed to a group of pesticides called neonicotinoids (neonics), considered to be harmful and potentially lethal to bees [74,75]. Growing evidence for their toxic impacts on bee populations range from reduced queen production [73], disorientation [76], reduced immunity [77], and mortality [76,78]. Though much of the research on the effect of neonics has focused on honeybees, evidence has also shown that such pesticides are also harmful to wild bee populations [79]. In response to the influx of scientific evidence and public pressure, governments have been taking action to reduce, and in the case of the European Union, to ban the use of neonicotinoid pesticides. Despite growing research underscoring the dangers of neonics, there remain significant gaps in our knowledge about the impacts of field-realistic doses, and how different neonics treatments affect different bee taxa [80], particularly as much of the research has focused solely on European and North American bees [81].
Debate continues about whether evidence conclusively links large-scale pollinator deaths directly to neonic use; this debate has been interpreted by some individuals, including policy-makers, researchers, journalists and members of the public, as indication that neonic exposure is not a cause. Here we present a more comprehensive examination of the socio-ecological system dynamics and highlight a series of additional drivers to illustrate the multiple causes contributing to the declines in pollinator species.
Often characterized by homogenous landscapes with little to no habitat or refuge for biodiversity, modern intensive agricultural systems themselves currently entail at least one cause of pollinator declines due to a lack of floral and nesting resources [82,83,84,85]. In addition to vegetated sites for refuge, pollinator health is dependent on the availability of a diverse range of plants (and their pollen) with non-overlapping flowering periods typically found in (semi-) natural habitats, as the flowering period of a single plant species found within managed crops systems is often shorter than the activity period of pollinators [86]. The general trend of intensification at smaller spatial scales combined with the extensification of agriculture across the landscape exerts multi-scalar pressures on pollinator populations [87,88].
Pollinators are also exposed to biophysical pressures, as are any other crop or animal species. Honeybees contend with a host of pests and pathogens [89] and colonies that are transported to and networked between multiple farms enhance the risk of rapid spread of diseases and pathogens between managed and wild populations [68,90]. Moreover, climate change introduces a layer of ecological complexity, and is projected to impact the phenology of mutualistic plant-pollinator relationships [91,92].
These additional pressures should not cast doubt on whether neonics are negatively impacting pollinator health, but demonstrate the complex reality that in CAS, “stressors do not act in isolation” [93]. Multiple causes are currently contributing to decreased health and vigour of pollinators, making them more vulnerable to that “last straw” that may push populations over a given threshold [82,94].
While there is little uncertainty that neonics negatively can impact honeybee health in several ways, there is no single smoking gun associated with pollinator declines, and no silver bullet for the larger challenge of maintaining robust pollinator populations in our intensively managed agro-ecosystems. We would argue that the disparity between the growing body of evidence of the various causes of pollinator declines and the lack of adequate policy response is not due to a lack of knowledge about the value of pollinators [65], but a generally inadequate approach to governing solutions to a problem involving multiple causes, facets and scales. Given the critical nature of pollinators to our food systems, definitive precautionary action to mitigate several likely contributors to pollinator declines is needed, even without proof of large impacts from each separately.
3.2. Paying for a Thousand Band-Aids? Rethinking PES to Integrate Complexity in Solving Distant Problems
Throughout Latin America and in many other nations, incentive programs have been envisioned as a solution to improving environmental outcomes associated with agriculture. These programs are referred to variously, most popularly as payments for ecosystem (or environmental) services (PES). Although their design directly addresses several characteristics of CAS, there is further potential for such programs to foster more resilient and sustainable food systems by accounting for more of the five identified characteristics of CAS. We draw upon a broad-ranging consideration of problems and potential solutions for PES [95].
PES programs are often looked to as solutions to problems in downstream systems, with the notion that external beneficiaries may be willing to pay for improved environmental outcomes [96]. Indeed, PES programs exemplify the recognition that on-farm actions can have considerable consequences at other scales, e.g., in downstream aquatic ecosystems where sedimentation and eutrophication may serve as important stressors [97,98]. While this recognition of teleconnections and multi-scalar dynamics is potentially helpful for food-system sustainability, it is also the case that those downstream systems should not be expected to change linearly as a result of altered inputs via the PES (see Sourcing Stifling Sediment, below). Instead, it may often be the case that the downstream systems will exhibit nonlinear dynamics complete with multiple stable states [30,33], time lags [49], and path-dependency or “hysteresis” [99,100]. Accordingly, it may take dedicated efforts at managing multiple stressors, and patience, before downstream systems are restored.
Given their voluntary nature, PES programs enjoy an important advantage in addressing multi-causality. Whereas demonstrated proof of harm may be expected before legislation is likely to prohibit or mandate management change (expected but not necessarily required) [101], policymakers or downstream beneficiaries may be satisfied with much less perfect science before addressing a stressor via PES [102].
Cumulative Impacts
Agriculture is the source of several stressors among many, but within agriculture, PES programs target the actions of individual producers as separate impacts to be addressed by “purchasing” behavior change (via an incremental addition of an extrinsic motivation). The changes in management are then expected to accumulate with commensurate positive effect. This approach misses the point that there are larger system dynamics at play, and addressing individual producers as separate rational agents may backfire. Rather interventions might seek to change norms, not just individual behaviours. Addressing each producer as a separate contributor to impacts could backfire in two ways. First, the pursuit of individual-level additionality—as sought in much of the economic-oriented literature on PES—risks distracting policymakers from a larger program objective that is perceived to be fair and achieves program-wide goals in the near and long terms [95]. Inequities—perceived or “real”—undermine the achievement of such goals [103]. Moreover, a focus on individual-level additions also seems to require considerable monitoring and enforcement costs, impeding solutions that might achieve the same collective objectives via trust and peer monitoring [95].
Second, the notion that motivation is a simple linear function of aggregate incentives risks crowding out intrinsic motivations via the addition of new monetary extrinsic ones, such as via PES [104]. The cumulative effect of the diverse motivations may be that the new instrumental one undermines the existing moral one [105,106].
Perhaps a more effective way forward is to ensure that institutions articulate appropriate values [104], placing rights and responsibilities where they are conducive to long-term sustainability [95]. To do so is to address the larger dynamics of the system [107], seeking to change the system’s norms and values rather than simply individual behavior via simplistic notions of producers as rational agents [108].
3.3. Regime Shifts and Pests: Pesticide Resistance and Pest Control
Agricultural pest management provides instructive examples of regime shifts. Such shifts can be ecological, as in long-term changes in the arthropod community, or evolutionary, as in large-scale genetic changes that are not easily reversed, such as the evolution of pesticide resistance. The use of pesticides and other management practices can drive these system shifts, changing a landscape with complex relationships between crops, pests and predators to a landscape dominated by pesticide use and simplified food webs.
In Figure 1, we present a regime-shift framework for thinking about pest management approaches following the logic of basins of attraction from resilience theory [38]. Under this framework, some agricultural systems maintain functional interactions within the food web, providing the ecosystem services of pest control and resistance mitigation (type 1 basins in Figure 1, green circles). These systems might generally revert to a high-complexity food web in the absence or failure of agricultural management. Other systems do not sustain the ecosystem service of pest control, and revert to widespread or acute pest outbreaks and associated crop collapses in the absence or failure of management (termed type 2 basins in Figure 1).
Regime shifts between these two types of basins are often not from one stable point to another, but rather from an artificially maintained non-equilibrium steady state within one basin of attraction to a different non-equilibrium state in another basin (Figure 1, arrow A). Pest control regime shift dynamics can function through either ecological or evolutionary mechanisms (purple and red circles, respectively, Figure 1).
Ecological Shift
Predators of agricultural pests are subject to a range of cumulative impacts that reduce their populations, including habitat degradation, climate change and pesticides. Pesticides can eliminate the predators of pests thereby decreasing pest predation [110], in cases leading to an ecological shift whereby predator species can no longer functionally control the pest population (arrow A, Figure 1) [111]. An ecological flip can also occur when pesticides lead to secondary pest outbreaks; pesticides, which are used to eliminate one type of pest, result in an increase in population of a new agricultural pest (often due to the elimination of a predator [112]. In this situation, the key components (i.e., pest species and pesticide type) change, but the dynamics of the pesticide dominated system remain similar (arrows between hatched purple circles, Figure 1).
Unless they play out at a landscape scale, ecological shifts are sometimes reversible as natural enemy populations can migrate into pesticide dominated areas or adjacent refuge areas [113]. In other cases, natural enemies may have to be re-introduced into the landscape and have their natural habitat protected or restored. The degree of active intervention required to move from a pest control system that is pesticide-dominated to one that is ecologically-mediated (beyond limiting the use of pesticides) depends on the existence of native natural enemies, connectivity to other arthropod communities, availability of predator habitat, and economic viability of imperfect crops.
Evolutionary Shift
The long-term and intensive use of pesticides is commonly referred to as the “pesticide treadmill”, where producers have to keep increasing the dose or changing the type of pesticide they use, as pests develop resistance to pesticides [114]. The evolution of resistance is well understood theoretically [115,116] and empirically [117]. Over 500 animal pests have developed resistance to one or more types of pesticide [118]. When pests evolve resistance to pesticides, the results can be widespread, long-term, and cause drastic changes in crop production and the agro-ecosystem. Pesticide resistance results in crop collapses bringing the system to a local point of stability (Figure 1 steady state type 3). If the producers adapt by switching pesticides or crops, resistance causes a shift between one pesticide dominated system to a different pesticide dominated system (red open circle, Figure 1). However, as resistance accumulates (perhaps to multiple pesticides), the stable state attractor of crop collapse becomes harder to escape, both because the genetics become efficient and less reversible [119], and because alternative pest management systems to flip to become more scarce.
Predators also play a secondary role in pest control: resistance mitigation. Predators can delay the evolution of pesticide resistance by preferentially predating insecticide resistant pests over susceptible pests. Initially modeled by Gould, Kennedy & Johnson [120], this role was subsequently supported through empirical work [121,122,123]. In these cases, the evolutionary pressure exerted by pest predators operates in the opposite direction to that of the pesticide, thereby attenuating the fast evolutionary response to a single strong selection agent.
Management Implications
A resilience theory perspective situates pest control approaches within different basins of attraction that surround distinct stable states, thereby clarifying the risks and potential benefits of transitions between these basins. Despite the risks of being within a type 2 basin of attraction, producers on the pesticide treadmill may not be able to easily transition to a type 1 system. This could be because (1) the environment cannot support a functioning arthropod food web that could provide sufficient pest control services, and/or (2) because socio-political factors do not incentivize such a transition, instead encouraging switching crops or pesticides, but remaining in the same (or similar) precarious position within the basin of a type 2 stable state.
Despite the abundance of theoretical and empirical studies about agricultural pests, the most effective components of alternative systems to conventional agriculture are not well understood quantitatively [124]. For example, organic agriculture has been shown to sustain higher levels of insect trophic complexity, and species evenness than conventional agriculture [125,126], and organic perennial systems can maintain stable pest levels over the long term [127]. However, landscape and management characteristics such as fallow management and planting date appear to be more important for the function of arthropod food webs in some cases than the use of pesticides themselves [128]. The characteristics that enable partially self-sustaining pest control that poses fewer risks in the event of management failure (i.e. type 1 basin systems) are clearly more complex than the presence or absence of pesticides alone. This may account for the conflicting results of different studies comparing pest damage between conventional and organic agriculture [124]. A quantitative understanding of these characteristics (such as fallow practices, crop rotations, landscape scale complexity) on both arthropod communities and resulting yields are needed to enact resilience-conscious pest management.
3.4. Sourcing Stifling Sediment: Teleconnections between Oyster Beds and Farms via Nitrogen Run-Off in Tasman Bay, New Zealand
Historically, Tasman Bay, New Zealand, is a site of dense mussel, scallop, and oyster reefs, formerly providing food and fisheries jobs for many commercial and recreational fishers, as well as local iwi (Maori tribes or nations) [129]. Shellfish populations have declined steeply and Tasman Bay commercial scallop harvest has been suspended since 2005 [130]. Today, prominent economic activities around the bay include aquaculture for green-lipped mussels and livestock ranching (on land) [130,131,132]. Both aquaculture and ranching are private operations that harvest private resources, potentially changing employment opportunities in the bay. Both activities are sources of tension, as aquaculture can compete with, rather than substitute, wild fisheries and ranching can contribute to contaminant runoff and sedimentation in the bays [34]. Despite attempts to rebuild wild shellfish populations through larval rearing and enhancement, achieving self-sustaining historic shellfish stocks (without enhancement) remain elusive [129,133]. What happened to a once-abundant resource in Tasman Bay? The current state of research on the topic provides some clues, discussed below.
Tasman Bay has undergone a radical biophysical transformation because of historical fishing practices. Commercial dredge fishing from the late 1800s to the 1960s reduced the complex three dimensional benthic habitat formed by layered mussel beds into a relatively flat, silty bottom [129]. At the same time, the coastline of Tasman Bay was filled in and dredged in an effort to cater to lifestyles and industry of the settler community, rather than that of the indigenous Maori [129,134]. These historical changes are legacy effects because they have long-term repercussions that shape how current social-ecological interactions occur [49,135]. The particular legacy effect is the transition to the flat, silty state of the bay, which is a regime shift from one system attractor to another [136]. Bivalves have been unable to sufficiently resettle to recreate historic benthic habitats conducive to mussels, scallops, and oysters (hard reef substrate, and multi-layered beds) largely because of continuous commercial dredge fishing as well as the continuous press disturbance of sediment to the bay from landuse [129,137]. Sediment can smother shellfish and prevent larval settlement of mussels, oysters, and scallops [137,138]. That Tasman Bay has not returned to historical conditions despite recent efforts to enhance shellfish biomass by artificially raising populations suggests that hysteresis effects are operating—that is, reversing ecosystem change will require a rebuilding of feedback processes that maintained the old system [139].
The new system is maintained by a variety of processes that add and maintain sediment in the bay—many of which are regulated through human activity. First, sediment is added to the bay mainly through input from terrestrial ecosystems, illustrating the importance of cross-system impacts [140]. Most land use around Tasman Bay is associated with dairy, beef, and sheep ranching, as well as forestry [34,132]. Runoff carrying sediment is directed through prominent currents towards Tasman Bay [140]. New Zealand is unique as an industrial country in that most of its export economy is driven by agriculture and its agricultural sector is fully exposed to international markets (New Zealand has no protection oriented subsidies, [141]). The influence of foreign markets on land use around New Zealand, including Tasman Bay, therefore teleconnects international commerce to important impacts on Tasman Bay ecology (such as [42] describe for other areas of the world). Second, commercial fishing and shipping vessels can re-suspend sediment that already exists in the bay, ensuring that sediment remains an outer layer in the benthos [34]. Finally, suspended mussel-lines in aquaculture operations can contribute debris and detritus (faeces and shell fragments) that further smother formerly productive wild shellfish habitat [131]. Multiple prominent contributions of sediment as an impact, interacting with other impacts such as shellfish harvest and pollution in the bay (some of which binds to sediment), contribute to cumulative impacts that prevent a return to a formerly productive shellfish ecosystem [34].
Temporal Scale
Central to Tasman Bay’s current state are historical changes that maintain a new system, meanwhile, memory of the bay’s historic state lingers, influencing current resource conflicts. Many fishers and conservationists wish for a return of productive wild fisheries with more diverse marine biodiversity, while some younger generations may be content with the bay’s current state, knowing only that state and increasingly relying on aquaculture for employment [130,142]. Shifting baselines that influence what people think the bay ought to be may reduce conflict in the future as memories fade [143]. Unless management addresses the wide-ranging feedbacks that maintain the current system, past ecosystems are unlikely to be reclaimed in any case [34]. Future climate change may also reinforce current feedbacks as more intense storms may increase sediment runoff to the bay [34].
Spatial Scale
The historical and current feedbacks that maintain their respective systems are drastically different in spatial scale. Whereas the past state of Tasman Bay was maintained by shellfish ecology maintaining adequate settlement substrate for new recruits, the current bay is maintained by human activities across ecosystems and international trade. Reclaiming Tasman Bay will likely require addressing the current cross-scale impacts that maintain the system’s current state [34].
3.5. Conflicting Scales in Governance of Puget Sound Riparian Restoration
In Washington State’s Puget Sound region, controversy has erupted over demands by the Treaty Tribes of Western Washington for stricter regulation and enforcement to protect salmon habitat. Agricultural producers are opposed to the proposed solutions and feel unfairly targeted. Passionate debate surrounded one small rule change for voluntary conservation programs on agricultural land. Why was such a small rule change such an intractable problem? Part of the answer can be understood by applying a CAS lens.
We studied this case using semi-structured interviews with 22 producers and 13 experts in addition to document analysis from policy and scientific meetings, public media campaigns, white papers and reports. We sought to understand the key values and perspectives of different groups, including: land managers/producers, Conservation Districts, government environmental agencies, and Treaty Tribes.
Good habitat for salmon is lacking or threatened throughout the Puget Sound. While salmon face multiple and cumulative threats (no smoking gun, death by a thousand cuts), lack of habitat is considered to be the limiting factor for healthy salmon populations. Strips of forest and vegetation along stream sides, called riparian buffers, provide many of the needed habitat functions. As a migrating species, salmon not only connect upstream producers and downstream fishers, but also respond to ocean conditions such as temperature and food supply. Thus use of fossil fuels in distant and proximate areas teleconnects to Puget Sound salmon management via climate change.
A variety of programs support producers to plant riparian buffers. The rule change in question would increase the minimum width of supported riparian buffer projects from 10 to 30 m. The Treaty Rights at Risk (TRAR) initiative has argued that 30 m is the minimum riparian buffer width necessary to create functional salmon habitat. TRAR is opposed to using federal and state funds for riparian buffer projects under this width, arguing that while narrow buffers improve water quality, they will not provide the needed habitat functions for salmon recovery. Conservation Districts (CDs) administer the funds and write contracts with producers and rural landowners to implement riparian restoration. The CDs do not support the change, arguing that increasing the minimum width will ultimately result in poorer salmon habitat and water quality as fewer landowners will be willing to sign up for the voluntary riparian restoration projects.
Temporal Scale
For the tribes, this is a long-standing problem to be solved via a long-run strategy. When tribes signed the Treaty of Point Elliott in 1855 they were assured to their right to fish salmon, recognizing the centrality of this resource. Yet even assuring this right to fish at traditional grounds took over 100 years. The struggle is captured by the quote from Chief Red Cloud: “They made us many promises, more than I can remember, but they never kept but one; they promised to take our land, and they took it.” Today fishing rights are established, yet now the struggle is to ensure that there are sufficient salmon to be fished. Legal battles have granted treaty holding tribes the status of co-managers of the salmon resource and established US government responsibility to assure adequate numbers of fish returns. For the TRAR group then, a short term reduction in riparian restoration programs is a small price to pay for the longer term goal of establishing treaty holding tribes as co-managers of salmon resources.
Agricultural producers on the other hand, talk of year-to-year profitability and generations, not centuries. Many in the agricultural community are worried about the loss of farmland in the region over the last few decades and the increasingly difficult economic situation many family producers face. They have watched farmland disappear from their communities and see strict rules for riparian buffers as one more threat. Based on data from the last few years, CDs worry that the proposed changes would impact their projects in the new few years.
Spatial Scale
These groups tend to view the problem at different spatial scales as well. Reports from the TRAR group present maps and statistics of the entire Puget Sound region, via data on acres of quality salmon habitat, total fish passages blocked and salmon returns. CDs and producers much more often examine these problems at a parcel scale. Considering one property at a time, they think about what projects can be accomplished to meet the landowner’s goals and to improve a suite of conservation-related goals, including salmon habitat. From a parcel-scale view, stricter rules mean fewer trees in the ground as producers will enroll less area or opt out of the program altogether.
4. Discussion: Rethinking Agriculture
Agriculture will have to contend with complex dynamics that affect the health of ecosystems in which it occurs, and thus threatens to undermine production in the long-term. Here we propose that such efforts will be greatly aided by paradigm shifts on three scales: governance, production and people (encompassing individuals and communities).
4.1. Governance Scale Shift: Farming Is Not Only Agricultural Production, but Also Land Management
Given the critical role that agricultural production systems play in mediating a wide range of environmental impacts, it will be more effective to address food production and environmental management goals simultaneously in farming, rather than farming only for food and addressing the ecological impacts separately. The case studies on Puget Sound and Tasman Bay show the challenges of (and potential conflict from) managing ecosystems without managing impacts from agriculture. Yet, managing agricultural land for both production and ecological goals faces a number of barriers: farmers/ranchers may identify as producers, not as land managers; because environmental impacts often manifest downstream, they may not be significant or obvious at the farm-scale; and managing agricultural lands for multiple goals can lead to real or perceived reductions in agricultural output. This shift does not require a change in farmer/rancher identity from “productivist” to “conservationist”. Rather, it facilitates a more explicit focus on land management goals, and a broadening of what producers view as a “good” farmer/rancher that includes more conservation related goals [144]. Underlying this shift are two strategies for change.
Strategy 1. Coordinate Governance of Agricultural Landscapes across Sectors
Many impacts to and from agricultural production require coordinated management. When there is no silver bullet, approaches will need to consider changes across landscapes and across management sectors (agriculture, fishing, forestry, urban land use planning, etc.), as shown in the Puget Sound and Tasman Bay case studies. In contrast to the marine context where it has become de rigeur to call for simultaneous management for fisheries, shipping, and habitat, in land based systems there is less integration of the agricultural production/environmental impact nexus with ecosystem and landscape scale spatial planning [61]. In the case of marine policy in North America, the management of primary industries is housed in the same government agencies as environmental protection (e.g., Fisheries and Oceans Canada, National Oceanic and Atmospheric Administration in the US). Yet, in a terrestrial context it is still common for agriculture to be managed by separate agencies than the species at risk that inhabit those landscapes and ecological impacts regarding air and water quality (e.g., Agriculture and Agri-Food Canada and Environment and Climate Change Canada; or in the US, the Environmental Protection Agencies and Fish and Wildlife Service manage water and air quality and species at risk listings whereas agriculture is managed by the US Department of Agriculture). For example, the Puget Sound Partnership coordinates dialogue across sectors to develop goals and strategies to address multiple interlinked challenges around issues such as salmon recovery, flood management and farmland protection. Such integrated approaches mean that producers have something to gain in the process (such as protection of farmland or better flood management) and may be more open to dialogue than when the focus is only on increasing regulations for environmental impacts from agriculture. Such integrated approaches can also consider impacts to yields and output, e.g., by creating protections for highly productive farmland that is often at risk from urban encroachment or by strategically targeting areas of the landscape for conservation or production priorities.
A variety of tools and policy mechanisms exist to support cross-sector governance (including the design of regulatory and voluntary programs discussed below). Approaches such as ecosystem-based management [145] and spatial/land-use planning [146,147,148] focus on science-based integrated management with active local participation. Ecosystem-based management (EBM) considers the interactive and cumulative effects that humans have on linked social and ecological systems [145]. Spatial planning is a way to implement EBM via a participatory process [146]. It involves selecting areas and times for activities in order to achieve economic, ecological and social goals [146]. Such integrative approaches seek to account for the various spatial, temporal, ecological and social systems as well as their interactions [149]. Various tools have been developed to help in participatory decision-making processes in the context of implementing ecosystem-based management or conducting land or marine-based spatial planning such as Structured Decision Making [150], Integrated Valuation of Ecosystem Services and Trade-offs (InVEST) [151,152], MARXAN with Zones, Formative Scenario Analysis [153], QUEST [154], ALCES [155], mediated modeling [156], and alternative futures research [157]. Such approaches could help to inform collaborative governance for challenges including loss of quality farmland, downstream impacts from chemical and nutrient use, and maintenance of pollinator habitat across the landscape.
Strategy 2. Drive Landscape Scale Change by Enabling (Where Possible) and Regulating (When Necessary)
There has recently been an emphasis on voluntary approaches to environmental management, but it’s important to note that many voluntary approaches work in tandem with underlying prescriptive regulations [104,158]. For governance processes to yield changes across the landscape (and seascape), some combination of regulatory and voluntary programs are likely needed. Voluntary (including incentive-based) programs can support motivated producers to steward their land, while regulations provide broader protection, but potentially less efficiently; combining these can both “crowd in” producers that are already interested in environmentally conscious land management and create hard limits within which all producers must operate. For example, China has taken the approach to strictly regulate pollution from large agri-business while offering incentives and support to small producers to accomplish environmental goals while maintaining rural lifestyles and cultures [159,160]. Voluntary programs like conservation easement purchase programs can be effective when landowner and conservation priorities naturally align. For example, they can help to keep out subdivisions and fences from rangelands, thereby preserving cultural landscapes that are important to ranchers, and also potentially helping conserve biodiversity on these lands [161]. While ranching is by no means always ecologically benign, in some contexts it may be the best option for preserving large-scale ecological processes [162], such as mimicking natural prairie systems [163].
Producers are often drawn to programs that reduce the threat of regulatory requirements, provide technical expertise and/or financial incentives [164]. Incentive-based programs compensate for, or share marginal costs with, producers for practices that have large-scale social/environmental benefits. Payments for ecosystem services (PES) programs enable subsidies to conditional on management practices that support or enhance ecosystem services [165]. Another option that can be used to directly target environmental risks and impacts is cost-sharing programs such as Canada’s Environmental Farm Plan (EFP) program. This program assists producers with addressing environmental risks by incentivising the adoption of environmentally beneficial management practices. The incentives typically take the form of one-off partial payments for specific practices (e.g., the purchasing of vegetation to plant in degraded riparian areas). Similar programs are facilitated via Conservation Districts in many US states, which provide technical and administrative support as well as cost-sharing. Cost-sharing (rather than paying the full opportunity cost) can “crowd in intrinsic motivations” for adopting environmentally beneficial management practices [95]. As agriculture is subsidized in many parts of the world, cost-sharing programs targeting ecosystem services offer an opportunity to use subsidies as economic incentives for sustainable land management. These kinds of conservation-focused subsidies can also account for local impacts driven by distant drivers via teleconnected relationships.
4.2. Production Scale Shift: From Prescriptive to Place-Based Farming Practices
Given that agricultural challenges are multi-causal, networked and teleconnected at multiple scales, solutions for sustainable agricultural systems are context-specific and cannot be reliably attained via specific practices applied at coarse scales [166]. This paradigm shift thus suggests that no one mode of farming (conventional, organic, small-scale, polyculture, etc.) can be the solution for both food production and environmental degradation, in line with other calls to move beyond panaceas [167]. Instead of globally-prescriptive solutions, place-based approaches to production would account for the socio-cultural, ecological, and economic context of a particular place, and select appropriate practices for that context. The concept of “place” goes beyond simple geography, and encompasses relational significance derived from everyday experience and practice, and emphasizes the nested and networked nature of food systems [168,169,170]. Place-based approaches to agricultural systems management that reflect the complex, multi-scalar, relational, and values-based nature of food systems have the potential to enhance socio-ecological resilience [171].
While established prescriptive regulatory frameworks such as organic certification offer some important policy interventions and guidelines for agriculture, they do not guarantee improved environmental or production outcomes. For example, while the organic movement has focused on a holistic farming system (including the cultivation of polycultures, enhancing soil quality, and fostering human, animal and societal health), organic certification as a label has largely focused on substituting inputs with “chemical-free” products [166,172]. A strict adherence to such a principle-based approach could impede the search for sustainable agricultural systems since the pathways to sustainability cannot be one-size-fits-all in a CAS. As such, effective and innovative solutions may need to cut across or even transcend different conventions or established approaches: conventional farms that cultivate varietals that are vigorous and well-adapted to climate extremes may be more “sustainable” than input-intensive organic certified farms, particularly those growing disease-sensitive or low-yielding products. Production systems that are reflective of place may need to integrate approaches from seemingly disparate sides of the management spectrum to achieve optimal outcomes, and the strategies employed may differ across farms and regions. This will require careful consideration of the needs of a particular place, while also considering the ways that place is linked to larger scale markets, policies and networks.
Strategy 3. Cultivate Production Systems Robust to Multiple Stressors
In contrast to the prevailing approach that focuses research dollars to maximize yield of single genetic variants under ideal circumstances or in response to single threats (e.g., drought, specific pests) [173], here we call for a strategy that eschews efficiency maximization in pursuit of farming that can better attend to the diverse challenges that necessarily arise from operating within a CAS. Agricultural production systems can expect to experience cumulative impacts as climate change will lead to combined stressors on production systems (such as drought and heat waves). As conventional agriculture has focused on increasing yields, it has at the same time decreased the diversity of production approaches, and therefore agricultural system adaptability and robustness. This means that unexpected events could impact a much greater area, making the consequences more substantial than in a more diverse system. For example, consolidations of slaughterhouses can increase the total risk in the food system as contamination would spread more quickly and impact many more animals, and subsequently, people. While some regime shifts are unexpected, some are well understood and predictable. As discussed in case study 3, agriculture can be seen as managed at far-from-equilibrium states, but the economic and environmental risks of a management failure depends on the relevant basin of attraction. Management that explicitly accounts for these risks might place more emphasis on robustness than short-term profitability. Some agricultural systems might minimize this tradeoff, but systems that simultaneously provide high yields, stability, and positive environmental outcomes remain elusive and require more research.
Conventional agriculture has benefited from decades of intensive funding. Research on alternative methods are in their infancy, but can produce practical results, e.g., microbial herbicides [174,175]. For example, organic agriculture suffers a yield gap compared to conventional agriculture, but use of Best Management Practices minimizes this gap [176]. Further research into organic practices could help to optimize yields in organic production systems. As well, organic agriculture is often more resistant to drought and excessive rainfall [176]. Perhaps the aspects of organic systems that account for yield gaps could be decoupled from those that generate resilient ecosystem services: e.g., a farm using mostly organic methods for long-term soil fertility, but supplementing with synthetic N might close a yield gap while preserving important water regulation ecosystem services. While in practice such novel solutions might run into conflict with farming philosophies, research can play a key role in exploring possibilities. Research on systems that transcend the organic/conventional divide is much needed to provide the empirical foundations for place based agriculture as this paper envisions.
Strategy 4. Account for Regime Shift Risks via Adaptation Plans: Make A Plan B and C
Rather than relying on mechanisms such as crop insurance to avoid catastrophic losses, agricultural communities might consider alternative crops or markets or even alternative economic activities, given potentially major shifts in climate and economic conditions. Instead of attempting to maintain present conditions, agricultural systems should be managed and governed to be prepared for many possible futures. Whereas in strategy 3 we discuss the first component of robustness—resistance to irreversible and dramatic change—here we focus on the second: recovery from change in the form of alternative pathways [177].
Agricultural production systems may experience pressure to adapt stemming from the impacts they cause systems (such as in Tasman Bay or Puget Sound). As well, agricultural production systems themselves may be vulnerable to system shifts (such a crashes in pollinator populations, pesticide resistance, or shifting climates). Place-based approaches and coordinated spatial planning (as described in strategy 1) can serve as mechanisms for collaborative dialogue and the consideration of alternate pathways. A key component of CAS thinking is acknowledging that there will be surprises, and that efforts to maintain a particular state, despite changing context, will accumulate risk as systems become more and more rigid, because disturbances are inevitable. If governance systems at various scales facilitate the realization of a “Plan B” (and C, D, etc.), the diversity in responses enables modularity and robustness, especially at coarser scales [178,179]. If a particular industry or crop is a backbone of a place, what other options exist in the face of crop failure or market price crashes?
Such alternative plans should also consider how to provide agricultural producers assistance to transition to new crops or production practices. In catastrophic cases where agriculture itself may not be a viable future for producers, supplementary or alternative livelihood options are key. Many factors impact the adoption of new practices by producers, including the quality and accessibility of information, integration in local agricultural networks (e.g., producer or watershed management groups), and financial capacity [180]. Programs such as those discussed in strategy 2 can facilitate such transitions.
Strategy 5. Employ Teleconnections to Drive and Spread Innovations and Political Movements towards More Sustainable Production
The search for solutions should not be a choice between global panaceas and locally imagined grassroots solutions; perhaps the best of both worlds can stem from a global network of food practitioners identifying place-appropriate solutions through teleconnected and place-specific knowledge. Agricultural intensification and associated environmental impacts are already influenced (arguably negatively) by teleconnections from consumer demand and—in some cases—pressure from integrated value-chain retailers (e.g., Walmart). The physical and social distancing that has resulted from the complexity of the global agri-food system has obscured connections and inserted a “metabolic rift” between sites of production and consumption. Where biogeochemical (i.e. nutrient cycling) and social processes (i.e. producer and eater relationships) were once tightly bound, the growing distancing is contributing to environmental crises [181,182,183].
Achieving robust agricultural systems may require addressing the negative effects of such market teleconnections, in part by exploiting other teleconnections via knowledge networks. Shifting cultural norms, laws, and regulations are large-scale processes that could positively teleconnect different landscapes vis-à-vis management regimes. Several food movements seek to change the governance of food systems, and empower individuals, groups and agencies through democratic processes. By creating opportunities for inclusive participation of producers and citizens alike, food and farmer movements that seek inclusive policies and structural changes in the food system can serve as drivers of more sustainable and place-based approaches to production, distribution and consumption [183,184]. Given the linkages between supply chains, consumption patterns, and production, these elements may need to be addressed in tandem (as elaborated in the following section). In the Global South, movements such as the MST (or Landless Rural Workers Movement) seek not only land reform to improve land access, but are also working to promote agroecological approaches to land management that improve both ecological and social outcomes of farming systems [185,186]. More broadly, the global mobilization of civil society groups around the concept of Food Sovereignty (championed by the international coalition La Via Campesina) seeks to regain citizen and community control in the food system [187]. These movements integrate concerns of power relations in the food system with attention to ecological dynamics and social equity—necessary considerations to address contemporary ecological and social crises related to agricultural production [183,188].
4.3. People Scale Shift: Seeking Systems that Support Participation of All People as Both Citizens and Consumers
While thus far we have focused on applying CAS thinking to agricultural production, a complete understanding of food systems must also include the consumption that ultimately drives production. There is too often a focus on production systems without actively engaging with demand-side systems, which are absolutely crucial to understanding and intervening in the dynamics of production. Some research has already addressed elements of the coupled challenge. For example, food security literature has examined potential consequences of changes in demand for high-impact animal based foods [189,190,191,192]. The movement for and concept of Food Sovereignty has highlighted the importance of changing not only production and consumption, but also power relations and governance in food systems than encompass both supply and demand aspects, as well as structural barriers to such changes [187]. Supply chain management and certification literature has described a multitude of labels signifying changes in production practices that might (under some circumstances) enable consumers to fund sustainability innovations in agriculture [193,194]. Related but largely distinct, payments for ecosystem services literature has examined multiple mechanisms by which beneficiaries of ecosystem services might fund sustainable management of agricultural systems to achieve their private (and sometimes collective) goals [96].
Yet challenges remain. One challenge is that ideas about citizen empowerment and social equity remain largely elusive in mainstream discussions and solutions around food systems sustainability and food security [188,195]. The second challenge is practical: how do we implement large-scale and systemic changes to enable collective citizen empowerment whilst our current system espouses individualistic action? Dominant and mainstream efforts have generally sought to enable the expression of consumer values—what an individual might want for themselves from their private consumption (either of agricultural products or ecosystem services). It will be crucial to develop mechanisms that enable such values effectively and efficiently, for example, without being stymied by analysis paralysis (associated with too many options, each too complex) [196,197].
Yet, a focus on consumer driven change not only assumes that consumer preferences should be guided by environmental sustainability [198], but that consumers are in a socio-economic position to act on these preferences. Moreover, in the role of “consumer”, our ability to vote is limited by the size of our pocket book and the composition of our plate. Therefore, equally crucial and arguably receiving less attention, are mechanisms that enable all people (regardless of socio-economic status and context) to express citizen values: preferences, principles and virtues about production processes and resulting distributions of harms and benefits. Citizen values, pertaining to things we want for society as a whole, are not well elicited via consumer purchases [199], which are limited by free-rider dynamics and the available options (or even more so, socio-economic access to resources and social context). Such points resonate with the food-sovereignty literature’s focus on moral dimensions of food systems and also power relations. Of course, such citizen values cannot be expected to generate uniformly supported visions of food systems, as values vary widely (e.g., some groups see environmental directions as pathways to unwanted government regulation and economic stagnation) [200]. Nevertheless, enabling the expression of such values is crucial from a CAS perspective, in that it presents a crucial feedback in the system (sensu Meadows): the goals of the system cannot be expected to serve people unless those people have a real opportunity to shape the offerings via new information flows [107].
Ultimately, a CAS-informed governance of agricultural production systems and their ecological impacts will require changes and action throughout the food system, including a more equitable distribution of responsibilities among producers, individuals (in their roles both as citizens and consumers), other players in supply chains, and policymakers, as well as rights and power between and within these groups. Such broad changes will require collective action and cannot be accomplished via individual consumption choices alone; they go beyond voting with one’s fork or wallet.
5. Conclusions
Given the scope of this paper, we focused on guiding principles for policy objectives, rather than specific policies, which we suggest be developed to account for the particularities of politics and context. While implementation of changes in the directions we suggest certainly presents a challenge, a number of papers in this special issue offer useful insights towards this end, as do other popular and academic literatures (e.g., [197]). Systems will fare better when we recognize that massive disturbances are here and coming, via governance systems that reflect the complex adaptive nature of agricultural production systems and landscapes. We hope that highlighting these 5 characteristics can inform such collective action towards governance of food systems as CAS. To this end, we have sought to bridge ideas from otherwise disparate literatures and to provide way-posts (in the form of strategies, paradigm shifts and system characteristics) to guide the governance of food and agricultural systems as CAS.
Acknowledgments
We would like to thank the Social Sciences and Humanities Research Council of Canada (Grant 22R75814) and the Natural Sciences and Engineering Research Council of Canada (Grant 17R82930) for support. We would also like to thank Navin Ramankutty, whose Global Food Security class inspired and informed several of the authors.
Author Contributions
Case studies were led as follows: (1) Klassen and Sharp; (2) Chan; (3) Kreitzman and Semmelink; (4) Singh; (5) Chapman. All authors contributed to writing and revising the remaining sections of the paper. Kreitzman conceptualized and created the regime shift figure and framework.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Robertson, G.P.; Swinton, S.M. Reconciling agricultural productivity and environmental integrity: A grand challenge for agriculture. Front. Ecol. Environ. 2005, 3, 38–46. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, C.D.; Karp, D.S.; Meyer, C.F.J.; Hadly, E.A.; Daily, G.C. Predicting biodiversity change and averting collapse in agricultural landscapes. Nature 2014, 509, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Balzar, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Donner, S.D.; Kucharik, C.J. Corn-based ethanol production compromises goal of reducing nitrogen export by the Mississippi River. Proc. Natl. Acad. Sci. USA 2008, 105, 4513–4518. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; McCoy, K.A. A qualitative meta-analysis reveals consistent effects of atrazine on freshwater fish and amphibians. Environ. Health Persp. 2010, 118, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Van Engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar]
- Garibaldi, L.A.; Aizen, M.A.; Klein, A.M.; Cunningham, S.A.; Harder, L.D. Global growth and stability of agricultural yield decrease with pollinator dependence. Proc. Natl. Acad. Sci. USA 2011, 108, 5909–5914. [Google Scholar] [CrossRef] [PubMed]
- Arthur, B.W.; Durlauf, S.N.; Lane, D.A. The Economy as an Evolving Complex System II; Addison-Wesley: Reading, MA, USA, 1997; Volume 27, pp. 1–6. [Google Scholar]
- Holland, J.H. Hidden Order; Addison-Wesley: Reading, MA, USA, 1995. [Google Scholar]
- Bullock, J.M.; Dhanjal Adams, K.L.; Milne, A.; Oliver, T.H.; Todman, L.C.; Whitmore, A.P.; Pywell, R.F. Resilience and food security: Rethinking an ecological concept. J. Ecol. 2017, 105, 880–884. [Google Scholar] [CrossRef]
- Wittman, H.; Chappell, M.J.; Abson, D.J.; Kerr, R.B.; Blesh, J.; Hanspach, J.; Perfecto, I.; Fischer, J. A social-ecological perspective on harmonizing food security and biodiversity conservation. Reg. Environ. Chang. 2016, 17, 1291–1301. [Google Scholar] [CrossRef]
- Wittman, H. Reframing agrarian citizenship: Land, life and power in Brazil. J. Rural Stud. 2009, 25, 120–130. [Google Scholar] [CrossRef]
- Darnhofer, I. Resilience and why it matters for farm management. Eur. Rev. Agric. Econ. 2014, 41, 461–484. [Google Scholar] [CrossRef]
- Vandermeer, J.H.; Perfecto, I. Syndromes of production in agriculture: Prospects for social-ecological regime change. Ecol. Soc. 2012, 17, 388–395. [Google Scholar] [CrossRef]
- Bennett, E.M.; Carpenter, S.; Gordon, L.; Ramankutty, N. Toward a more resilient agriculture. Solutions 2014, 5, 65–75. [Google Scholar]
- Rammel, C.; Stagl, S.; Wilfing, H. Managing complex adaptive systems—A co-evolutionary perspective on natural resource management. Ecol. Econ. 2007, 63, 9–21. [Google Scholar] [CrossRef]
- Mahon, R.; McConney, P.; Roy, R.N. Governing fisheries as complex adaptive systems. Mar. Policy 2008, 32, 104–112. [Google Scholar] [CrossRef]
- Levin, P.S.; Mollmann, C. Marine ecosystem regime shifts: Challenges and opportunities for ecosystem-based management. Philos. Trans. R. Soc. B 2015, 370, 1659. [Google Scholar] [CrossRef]
- Ban, N.C.; Mills, M.; Tam, J.; Hicks, C.C.; Klain, S.C.; Stoeckl, N.; Bottrill, M.C.; Levine, J.; Pressey, R.L.; Satterfield, T.; et al. A social-ecological approach to conservation planning: Embedding social considerations. Front. Ecol. Environ. 2013, 11, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Schianetz, K.; Kavanagh, L. Sustainability indicators for tourism destinations: A complex adaptive systems approach using systemic indicator systems. J. Sustain. Tour. 2008, 16, 601–628. [Google Scholar] [CrossRef]
- Gross, J.E.; McAllister, R.R.J.; Abel, N.; Smith, D.M.S.; Maru, Y. Australian rangelands as complex adaptive systems: A conceptual model and preliminary results. Environ. Modell. Softw. 2006, 21, 1264–1272. [Google Scholar] [CrossRef]
- Klerkx, L.; Aarts, N.; Leeuwis, C. Adaptive management in agricultural innovation systems: The interactions between innovation networks and their environment. Agric. Syst. 2010, 103, 390–400. [Google Scholar] [CrossRef]
- Levin, S.A. The problem of pattern and scale in ecology: The Robert H. MacArthur Award lecture. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Darling, E.S.; Côté, I.M. Quantifying the evidence for ecological synergies. Ecol. Lett. 2008, 11, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative stable states in ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- Liu, J.; Dietz, T.; Carpenter, S.R.; Alberti, M.; Folke, C.; Moran, E.; Pell, A.N.; Deadman, P.; Kratz, T.; Lubchenco, J. Complexity of coupled human and natural systems. Science 2007, 317, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Cash, D.; Adger, W.N.; Berkes, F.; Garden, P. Scale and cross-scale dynamics: Governance and information in a multilevel world. Ecol. Soc. 2006, 11, 3213–3217. [Google Scholar] [CrossRef]
- Levin, S.A. Multiple scales and the maintenance of biodiversity. Ecosystems 2000, 3, 498–506. [Google Scholar] [CrossRef]
- Singh, G.G.; Sinner, J.; Ellis, J.; Kandlikar, M.; Halpern, B.S.; Satterfield, T.; Chan, K.M.A. Mechanisms and risk of cumulative impacts to coastal ecosystem services: An expert elicitation approach. J. Environ. Manag. 2017, 199, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and cumulative effects of multiple human stressors in marine systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Mcleod, K.L.; Rosenberg, A.A.; Crowder, L.B. Managing for cumulative impacts in ecosystem-based management through ocean zoning. Ocean Coast. Manag. 2008, 51, 203–211. [Google Scholar] [CrossRef]
- O’Brien, K.; Leichenko, R.; Kelkar, U.; Venema, H.; Aandahl, G.; Tompkins, H.; Javed, A.; Bhadwal, S.; Barg, S.; Nygaard, L.; et al. Mapping vulnerability to multiple stressors: climate change and globalization in India. Glob. Environ. Chang. 2004, 14, 303–313. [Google Scholar] [CrossRef]
- Folke, C. Resilience: The emergence of a perspective for social–ecological systems analyses. Glob. Environ. Chang. 2006, 16, 253–267. [Google Scholar] [CrossRef]
- Otto, S.P.; Day, T. A Biologist’s Guide to Mathematical Modeling in Ecology and Evolution; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Lewontin, R.C. The meaning of stability. Brookhaven Symp. Biol. 1969, 22, 13–24. [Google Scholar] [PubMed]
- Seto, K.C.; Reenberg, A.; Boone, C.G.; Fragkias, M.; Haase, D.; Langanke, T.; Marcotullio, P.; Munroe, D.K.; Olah, B.; Simon, D. Urban land teleconnections and sustainability. Proc. Natl. Acad. Sci. USA 2012, 109, 7687–7692. [Google Scholar] [CrossRef] [PubMed]
- Adger, W.N.; Eakin, H.; Winkels, A. Nested and teleconnected vulnerabilities to environmental change. Front. Ecol. Environ. 2008, 7, 150–157. [Google Scholar] [CrossRef]
- Eakin, H.; Winkels, A.; Sendzimir, J. Nested vulnerability: Exploring cross-scale linkages and vulnerability teleconnections in Mexican and Vietnamese coffee systems. Environ. Sci. Policy 2009, 12, 398–412. [Google Scholar] [CrossRef]
- Collins, S.L.; Carpenter, S.R.; Swinton, S.M.; Orenstein, D.E.; Childers, D.L.; Gragson, T.L.; Grimm, N.B.; Grove, J.M.; Harlan, S.L.; Kaye, J.P.; et al. An integrated conceptual framework for long-term social–ecological research. Front. Ecol. Environ. 2011, 9, 351–357. [Google Scholar] [CrossRef]
- Eisler, M.C.; Lee, M.R.; Tarlton, J.F.; Martin, G.B.; Beddington, J.; Dungait, J.A.; Greathead, H.; Liu, J.; Mathew, S.; Miller, H. Agriculture: Steps to sustainable livestock. Nature 2014, 507, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Guswa, A.J.; Brauman, K.A.; Brown, C.; Hamel, P.; Keeler, B.L.; Sayre, S.S. Ecosystem services: Challenges and opportunities for hydrologic modeling to support decision making. Water Resour. Res. 2014, 50, 4535–4544. [Google Scholar] [CrossRef]
- Brown, C.J.; Saunders, M.I.; Possingham, H.P.; Richardson, A.J. Managing for interactions between local and global stressors of ecosystems. PLoS ONE 2013, 8, e65765. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.R.; Graham, M.D.; Vinebrooke, R.D.; Findlay, D.L.; Paterson, M.J.; Turner, M.A. Multiple anthropogenic stressors cause ecological surprises in boreal lakes. Glob. Chang. Biol. 2006, 12, 2316–2322. [Google Scholar] [CrossRef]
- Jackson, J.B.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, W.Z.; Zhang, X.C.; Zheng, F.L. Impacts of land use change and climate variability on hydrology in an agricultural catchment on the Loess Plateau of China. J. Hydrol. 2009, 377, 35–42. [Google Scholar] [CrossRef]
- Piao, S.; Ciais, P.; Huang, Y.; Shen, Z.; Peng, S.; Li, J.; Zhou, L.; Liu, H.; Ma, Y.; Ding, Y.; et al. The impacts of climate change on water resources and agriculture in China. Nature 2010, 467, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.; Meyers, J. Thresholds in ecological and social–ecological systems: A developing database. Ecol. Soc. 2004, 9, 3438–3447. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Meadows, D.H. Thinking in Systems; Chelsea Green Publishing: Windsor County, VT, USA, 2008. [Google Scholar]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Cardille, J.A.; Bennett, E.M. Ecology: Tropical teleconnections. Nat. Geosci. 2010, 3, 154–155. [Google Scholar] [CrossRef]
- Güneralp, B.; Seto, K.C.; Ramachandran, M. Evidence of urban land teleconnections and impacts on hinterlands. Curr. Opin. Environ. Sustain. 2013, 5, 445–451. [Google Scholar] [CrossRef]
- Liu, J.; Hull, V.; Batistella, M.; DeFries, R.; Dietz, T.; Fu, F.; Hertel, T.W.; Izaurralde, R.C.; Lambin, E.F.; Li, S.; et al. Framing sustainability in a telecoupled world. Ecol. Soc. 2013, 18, 344–365. [Google Scholar] [CrossRef]
- Dias, P.C. Sources and sinks in population biology. Trends Ecol. Evol. 1996, 11, 326–330. [Google Scholar] [CrossRef]
- Kreitzman, M.; Ashander, J.; Driscoll, J.; Bateman, A.W.; Chan, K.M.A.; Lewis, M.A.; Krkosek, M. Wild salmon sustain the effectiveness of parasite control on salmon farms: Conservation implications from an evolutionary ecosystem service. Conserv. Lett. 2017. [Google Scholar] [CrossRef]
- Fischer, J.; Abson, D.J.; Butsic, V.; Chappell, M.J.; Ekroos, J.; Hanspach, J.; Kuemmerle, T.; Smith, H.G.; Wehrden, H. Land sparing versus land sharing: Moving forward. Conserv. Lett. 2014. [Google Scholar] [CrossRef]
- Gosnell, H.; Abrams, J. Amenity migration: Diverse conceptualizations of drivers, socioeconomic dimensions, and emerging challenges. GeoJournal 2011, 76, 303–322. [Google Scholar] [CrossRef]
- Durand, F. What’s the Deal with Interstellar’s Idea that Corn and Okra Are the Only Crops Left in the Future? Let’s Ask a Scientist. Available online: http://www.thekitchn.com/could-a-crop-blight-really-wipe-out-everything-but-corn-and-okra-food-on-film-212647 (accessed on 9 June 2015).
- Tilman, D.; Reich, P.B.; Isbell, F. Biodiversity impacts ecosystem productivity as much as resources, disturbance, or herbivory. Proc. Natl. Acad. Sci. USA 2012, 109, 10394–10397. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Singh, G.M.; Mozaffarian, D.; Myers, S.S. Effects of decreases of animal pollinators on human nutrition and global health: A modelling analysis. Lancet 2015, 386, 1964–1972. [Google Scholar] [CrossRef]
- Gordon, R.; Bresolin-Schott, N.; East, I.J. Nomadic beekeeper movements create the potential for widespread disease in the honeybee industry. Aust. Vet. J. 2014, 92, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Committee on the Status of Pollinators in North America; Board on Life Sciences; Resources, Board on Agriculture and Natural Resources; Division on Earth and Life Studies; National Research Council of the National Academies. Status of Pollinators in North America; National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native bees provide insurance against ongoing honey bee losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.A.; Garratt, M.P.D.; Wickens, J.B.; Wickens, V.J.; Potts, S.G.; Raine, N.E. Neonicotinoid pesticide exposure impairs crop pollination services provided by bumblebees. Nature 2015, 528, 548–550. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Cutler, G.C.; Scott-Dupree, C.D.; Drexler, D.M. Honey bees, neonicotinoids and bee incident reports: The Canadian situation. Pest Manag. Sci. 2013, 70, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Blacquière, T.; Field, L.M.; Hails, R.S.; Potts, S.G.; Raine, N.E.; Vanbergen, A.J.; McLean, A.R. A restatement of recent advances in the natural science evidence base concerning neonicotinoid insecticides and insect pollinators. Proc. Roy. Soc. B Biol. Sci. 2015, 282, 1818. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed]
- Mogren, C.L.; Lundgren, J.G. Neonicotinoid-contaminated pollinator strips adjacent to cropland reduce honey bee nutritional status. Sci. Rep. 2016, 6, 29608. [Google Scholar] [CrossRef] [PubMed]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Lundin, O.; Rundlöf, M.; Smith, H.G.; Fries, I.; Bommarco, R. Neonicotinoid insecticides and their impacts on bees: A systematic review of research approaches and identification of knowledge gaps. PLoS ONE 2015, 10, e0136928. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.J.; Baldock, K.C.R.; Brown, M.J.F.; Cresswell, J.E.; Dicks, L.V.; Fountain, M.T.; Garratt, M.P.D.; Gough, L.A.; Heard, M.S.; Holland, J.M.; et al. Protecting an Ecosystem Service: Approaches to Understanding and Mitigating Threats to Wild Insect Pollinators, 1st ed.; Elsevier: Toronto, ON, Canada, 2016; Volume 54, pp. 135–206. [Google Scholar]
- Vanbergen, A.J.; Initiative, T.I.P. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Williams, N.M.; Thorp, R.W. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA 2002, 99, 16812–16816. [Google Scholar] [CrossRef] [PubMed]
- Kovács-Hostyánszki, A.; Espíndola, A.; Vanbergen, A.J.; Settele, J.; Kremen, C.; Dicks, L.V. Ecological intensification to mitigate impacts of conventional intensive land use on pollinators and pollination. Ecol. Lett. 2017, 20, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Senapathi, D.; Biesmeijer, J.C.; Breeze, T.D.; Kleijn, D.; Potts, S.G.; Carvalheiro, L.G. Pollinator conservation—The difference between managing for pollination services and preserving pollinator diversity. Curr. Opin. Insect Sci. 2015, 12, 93–101. [Google Scholar] [CrossRef]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed]
- Runckel, C.; Flenniken, M.L.; Engel, J.C.; Ruby, J.G.; Ganem, D.; Andino, R.; DeRisi, J.L. Temporal analysis of the honey bee microbiome reveals four novel viruses and seasonal prevalence of known viruses, nosema, and crithidia. PLoS ONE 2011, 6, e20656. [Google Scholar] [CrossRef] [PubMed]
- Fürst, M.A.; McMahon, D.P.; Osborne, J.L.; Paxton, R.J.; Brown, M.J.F. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant-pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Hegland, S.J.; Nielsen, A.; Lázaro, A.; Bjerknes, A.-L.; Totland, Ø. How does climate warming affect plant-pollinator interactions? Ecol. Lett. 2009, 12, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E. The canary in the coalmine; bee declines as an indicator of environmental health. Sci. Prog. 2016, 99, 312–326. [Google Scholar] [CrossRef] [PubMed]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan Dewenter, I.; Szentgyörgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.M.A.; Anderson, E.; Chapman, M.; Jespersen, K.; Olmsted, P. Payments for ecosystem services: Rife with problems and potential—for transformation towards sustainability. Ecol. Econ. 2017, 140, 110–122. [Google Scholar] [CrossRef]
- Wunder, S. When payments for environmental services will work for conservation. Conserv. Lett. 2013, 6, 230–237. [Google Scholar] [CrossRef]
- Owens, P.N.; Batalla, R.J.; Collins, A.J.; Gomez, B.; Hicks, D.M.; Horowitz, A.J.; Kondolf, G.M.; Marden, M.; Page, M.J.; Peacock, D.H.; et al. Fine-grained sediment in river systems: Environmental significance and management issues. River Res. Appl. 2005, 21, 693–717. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Mumby, P.J.; Hastings, A.; Edwards, H.J. Thresholds and the resilience of Caribbean coral reefs. Nature 2007, 450, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Groffman, P.M.; Baron, J.S.; Blett, T.; Gold, A.J.; Goodman, I.; Gunderson, L.H.; Levinson, B.M.; Palmer, M.A.; Paerl, H.W.; Peterson, G.D.; et al. Ecological thresholds: The key to successful environmental management or an important concept with no practical application? Ecosystems 2006, 9, 1–13. [Google Scholar] [CrossRef]
- Oreskes, N. Science and public policy: What’s proof got to do with it? Environ. Sci. Policy 2004, 7, 369–383. [Google Scholar] [CrossRef]
- Naeem, S.; Ingram, J.C.; Varga, A.; Agardy, T.; Barten, P.; Bennett, G.; Bloomgarden, E.; Bremer, L.L.; Burkill, P.; Cattau, M.; Ching, C.; et al. Get the science right when paying for nature’s services. Science 2015, 347, 1206–1207. [Google Scholar] [CrossRef] [PubMed]
- Pascual, U.; Phelps, J.; Garmendia, E.; Brown, K.; Corbera, E.; Martin, A.; Gómez-Baggethun, E.; Muradian, R. Social equity matters in payments for ecosystem services. BioScience 2014, 64, 1027–1036. [Google Scholar] [CrossRef]
- Vatn, A. An institutional analysis of payments for environmental services. Ecol. Econ. 2010, 69, 1245–1252. [Google Scholar] [CrossRef]
- Frey, B.S.; Jegen, R. Motivation crowding theory. J. Econ. Surv. 2001, 15, 589–611. [Google Scholar] [CrossRef]
- Fehr, E.; Falk, A. Psychological foundations of incentives. Eur. Econ. Rev. 2002, 46, 687–724. [Google Scholar] [CrossRef]
- Meadows, D.H. Leverage points. Solu. J. 2010, 1, 41–49. [Google Scholar]
- Levine, J.; Chan, K.M.A.; Satterfield, T. Ecological economics. Ecol. Econ. 2015, 114, 22–32. [Google Scholar] [CrossRef]
- Roopnarine, P.D.; Angielczyk, K.D. Community stability and selective extinction during the Permian-Triassic mass extinction. Science 2015, 350, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Bommarco, R.; Miranda, F.; Bylund, H.; Björkman, C. Insecticides suppress natural enemies and increase pest damage in cabbage. J. Econ. Entomol. 2011, 104, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Angelo, M.J. The Law and Ecology of Pesticides and Pest Management; Taylor & Francis Group: Abingdon, UK, 2013; pp. 1–330. [Google Scholar]
- Dutcher, J.D. A review of resurgence and replacement causing pest outbreaks in IPM. In General Concepts in Integrated Pest and Disease Management; Springer: Dordrecht, The Netherlands, 2007; pp. 27–43. [Google Scholar]
- Rand, T.A.; Tylianakis, J.M.; Tscharntke, T. Spillover edge effects: The dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitats. Ecol. Lett. 2006, 9, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Arbuckle, J.G. Farmer perspectives on pesticide resistance. Exten. Community Econ. Dev. Publ. 2014, 25, 1–8. [Google Scholar]
- Comins, H.N. The development of insecticide resistance in the presence of migration. J. Theor. Biol. 1977, 64, 177–197. [Google Scholar] [CrossRef]
- May, R.M.; Dobson, A.P. Population Dynamics and the Rate of Evolution of Pesticide Resistance; National Academy Press: Washington, DC, USA, 1986; pp. 170–193. [Google Scholar]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Whalon, M.E.; Mota-Sanchez, D.; Hollingworth, R.M. Analysis of Global Pesticide Resistance in Arthropods; CABI: Wallingford, UK, 2008; pp. 5–31. [Google Scholar]
- Uyenoyama, M.K. Pleiotropy and the evolution of genetic systems conferring resistance to pesticides. In Pesticide Resistance: Strategies and Tactics for Management; National Academies Press: Washington, DC, USA, 1986; pp. 207–221. [Google Scholar]
- Gould, F.; Kennedy, G.G.; Johnson, M.T. Effects of natural enemies on the rate of herbivore adaptation to resistant host plants. Entomol. Exp. Appl. 2011, 58, 1–14. [Google Scholar] [CrossRef]
- Liu, X.; Chen, M.; Collins, H.L.; Onstad, D.W.; Roush, R.T.; Zhang, Q.; Earle, E.D.; Shelton, A.M. Natural enemies delay insect resistance to Bt crops. PLoS ONE 2014, 9, e90366. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, A.J.; Hannon, E.R.; Sisterson, M.S.; Stock, S.P.; Carrière, Y.; Tabashnik, B.E. Effects of entomopathogenic nematodes on evolution of pink bollworm resistance to Bacillus thuringiensis toxin Cry1Ac. J. Econ. Entomol. 2012, 105, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- Berticat, C.; Duron, O.; Heyse, D.; Raymond, M. Insecticide resistance genes confer a predation cost on mosquitoes, Culex pipiens. Genet. Res. 2004, 83, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Letourneau, D.K.; Bothwell, S.G. Comparison of organic and conventional farms: Challenging ecologists to make biodiversity functional. Front. Ecol. Environ. 2008, 6, 430–438. [Google Scholar] [CrossRef]
- Macfadyen, S.; Gibson, R.; Polaszek, A.; Morris, R.J.; Craze, P.G.; Planqué, R.; Symondson, W.O.C.; Memmott, J. Do differences in food web structure between organic and conventional farms affect the ecosystem service of pest control? Ecol. Lett. 2009, 12, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Crowder, D.W.; Northfield, T.D.; Strand, M.R.; Snyder, W.E. Organic agriculture promotes evenness and natural pest control. Nature 2010, 466, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Vandermeer, J.; Perfecto, I.; Philpott, S. Ecological complexity and pest control in organic coffee production: Uncovering an autonomous ecosystem service. BioScience 2010, 60, 527–537. [Google Scholar] [CrossRef]
- Letourneau, D.K.; Goldstein, B. Pest damage and arthropod community structure in organic vs. conventional tomato production in California. J. Appl. Ecol. 2001, 38, 557–570. [Google Scholar] [CrossRef]
- Handley, S. An Analysis of Historical Impacts and Composition of the Benthic Environment of Tasman and Golden Bays; National Institute of Water and Atmospheric Research: Nelson, New Zealand, 2006. [Google Scholar]
- Gillespie, P.A.; Forrest, R.W.; Peake, B.M.; Basher, L.R.; Clement, D.M.; Dunmore, R.A.; Hicks, D.M. Spatial delineation of the depositional footprint of the Motueka River outwelling plume in Tasman Bay, New Zealand. N. Z. J. Mar. Freshw. Res. 2011, 45, 455–475. [Google Scholar] [CrossRef]
- Gibbs, M.M. Sedimentation, suspension, and resuspension in Tasman Bay and Beatrix Bay, New Zealand, two contrasting coastal environments which thermally stratify in summer. N. Z. J. Mar. Freshw. Res. 2001, 35, 951–970. [Google Scholar] [CrossRef]
- Cornelisen, C.D.; Gillespie, P.A.; Kirs, M.; Young, R.G.; Forrest, R.W.; Barter, P.J.; Knight, B.R.; Harwood, V.J. Motueka River plume facilitates transport of ruminant faecal contaminants into shellfish growing waters, Tasman Bay. N. Z. J. Mar. Freshw. Res. 2011, 45, 477–495. [Google Scholar] [CrossRef]
- Brown, S.N. Ecology and Enhancement of the Flat Oyster Ostrea chilensis (Philippi, 1845) in Central New Zealand. Ph.D. Thesis, University of Canterbury, Christchurch, New Zealand, 2011. [Google Scholar]
- Thrush, S.F.; Hewitt, J.E.; Cummings, V.J.; Ellis, J.I.; Hatton, C.; Lohrer, A.; Norkko, A. Muddy waters: Elevating sediment input to coastal and estuarine habitats. Front. Ecol. Environ. 2004, 2, 299–306. [Google Scholar] [CrossRef]
- Knick, S.T.; Rotenberry, J.T. Ghosts of habitats past: Contribution of landscape change to current habitats used by shrubland birds. Ecology 2000, 81, 220–227. [Google Scholar] [CrossRef]
- Biggs, R.; Carpenter, S.R.; Brock, W.A. Turning back from the brink: Detecting an impending regime shift in time to avert it. Proc. Natl. Acad. Sci. USA 2009, 106, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Cummings, V.; Hewitt, J.; Thrush, S.; Norkko, A. Determining effects of suspended sediment on condition of a suspension feeding bivalve (Atrina zelandica): Results of a survey, a laboratory experiment and a field transplant experiment. J. Exp. Mar. Biol. Ecol. 2002, 267, 147–174. [Google Scholar] [CrossRef]
- MacKenzie, L.; Adamson, J. Water column stratification and the spatial and temporal distribution of phytoplankton biomass in Tasman Bay, New Zealand: Implications for aquaculture. N. Z. J. Mar. Freshw. Res. 2004, 38, 705–728. [Google Scholar] [CrossRef]
- Schreiber, S.; Rudolf, V.H.W. Crossing habitat boundaries: Coupling dynamics of ecosystems through complex life cycles. Ecol. Lett. 2008, 11, 576–587. [Google Scholar] [CrossRef] [PubMed]
- Tuckey, B.J.; Gibbs, M.T.; Knight, B.R.; Gillespie, P.A. Tidal circulation in Tasman and Golden Bays: Implications for river plume behaviour. N. Z. J. Mar. Freshw. Res. 2006, 40, 305–324. [Google Scholar] [CrossRef]
- Kelsey, J. The New Zealand Experiment: A World Model for Structural Adjustment? Bridget Williams Books: Wellington, New Zealand, 2015. [Google Scholar]
- Gibbs, M.T. Interactions between bivalve shellfish farms and fishery resources. Aquaculture 2004, 240, 267–296. [Google Scholar] [CrossRef]
- Brown, C.J.; Trebilco, R. Unintended cultivation, shifting baselines, and conflict between objectives for fisheries and conservation. Conserv. Biol. 2014, 28, 677–688. [Google Scholar] [CrossRef] [PubMed]
- McGuire, J.; Morton, L.W.; Cast, A.D. Reconstructing the good farmer identity: Shifts in farmer identities and farm management practices to improve water quality. Agric. Hum. Values 2012, 30, 57–69. [Google Scholar] [CrossRef]
- McLeod, K.; Leslie, H. Ecosystem-Based Management for the Oceans; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Ehler, C.; Douvere, F. Navigating the Future of Marine World Heritage: Results from the First World Heritage Marine Site Managers Meeting Honolulu, Hawaii, 1–3 December 2010. Available online: http://www.unesco.org/library/PDF/193111e.pdf (accessed on 6 September 2017).
- Lindenmayer, D.; Hobbs, R.J.; Montague-Drake, R.; Alexandra, J.; Bennett, A.; Burgman, M.; Cale, P.; Calhoun, A.; Cramer, V.; Cullen, P.; et al. A checklist for ecological management of landscapes for conservation. Ecol. Lett. 2008, 11, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Opdam, P.; Wascher, D. Climate change meets habitat fragmentation: Linking landscape and biogeographical scale levels in research and conservation. Biol. Conserv. 2004, 117, 285–297. [Google Scholar] [CrossRef]
- Rosenberg, A.A.; McLeod, K. Implementing ecosystem-based approaches to management for the conservation of ecosystem services: Politics and socio-economics of ecosystem-based management of marine resources. Mar. Ecol. Prog. Ser. 2005, 300, 271–274. [Google Scholar] [CrossRef]
- Gregory, R.; Failing, L.; Harstone, M.; Long, G.; McDaniels, T. Structured Decision Making: A Practical Guide to Environmental Management Choices; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Guerry, A.D.; Ruckelshaus, M.H.; Arkema, K.K.; Bernhardt, J.R.; Guannel, G.; Kim, C.K.; Marsik, M.; Papenfus, M.; Toft, J.E.; Verutes, G.; et al. Modeling benefits from nature: Using ecosystem services to inform coastal and marine spatial planning. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2012, 8, 107–121. [Google Scholar] [CrossRef]
- Nelson, E.; Mendoza, G.; Regetz, J.; Polasky, S.; Tallis, H.; Cameron, D.R.; Chan, K.M.A.; Daily, G.C.; Goldstein, J.; Kareiva, P.M.; et al. Modeling multiple ecosystem services, biodiversity conservation, commodity production, and tradeoffs at landscape scales. Front. Ecol. Environ. 2009, 7, 4–11. [Google Scholar] [CrossRef]
- Tietje, O. Identification of a small reliable and efficient set of consistent scenarios. Eur. J. Oper. Res. 2005, 162, 418–432. [Google Scholar] [CrossRef]
- Robinson, J.; Carmichael, J.; Van Wynsberghe, R.; Journeay, M.; Rogers, L. Sustainability as a problem of design: Interactive science in the Georgia Basin. Integr. Asses. 2006, 6, 165–192. [Google Scholar]
- Schneider, R.R.; Stelfox, J.B.; Boutin, S.; Wasel, S. Managing the cumulative impacts of land uses in the western canadian sedimentary basin: A modeling approach. Ecol. Soc. 2003, 7, 928–930. [Google Scholar] [CrossRef]
- Metcalf, S.S.; Wheeler, E.; Bendor, T.K.; Lubinski, K.S.; Hannon, B.M. Sharing the floodplain: Mediated modeling for environmental management. Environ. Model. Softw. 2010, 25, 1282–1290. [Google Scholar] [CrossRef]
- Steinitz, C.; Arias, H.; Bassett, S.; Flaxman, M.; Goode, T.; Maddock, T., III; Mouat, D.; Peiser, R.; Shearer, A. Alternative Futures for Changing Landscapes; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- Porras, I.; Barton, D.N.; Miranda, M. Learning from 20 Years of Payments for Ecosystem Services in Costa Rica; Shaping Sustainable Markets Papers: London, UK, 2013. [Google Scholar]
- Ruhl, J.B. Farms, Their Environmental Harms, and Environmental Law: Taking the Great Leap from Anti-Law to Positive Law in Farm Policy. Available online: http://scholarship.law.berkeley.edu/cgi/viewcontent.cgi?article=1623&context=elq (accessed on 7 September 2017).
- Tanentzap, A.J.; Lamb, A.; Walker, S.; Farmer, A. Resolving conflicts between agriculture and the natural environment. PLoS Biol. 2015, 13, e1002242. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.N. Can cows and conservation mix? BioScience 2001, 51, 85–90. [Google Scholar] [CrossRef]
- Curtin, C.G.; Sayre, N.F.; Lane, B.D. Transformations of the Chihuahuan Borderlands: Grazing, fragmentation, and biodiversity conservation in desert grasslands. Environ. Sci. Policy 2002, 5, 55–68. [Google Scholar] [CrossRef]
- Austin, M.A.; Buffett, D.A.; Nicolson, D.J.; Scudder, G.G.E.; Stevens, V. (Eds.) Taking Nature’s Pulse: The Status of Biodiversity in British Columbia; Biodiversity BC: Victoria, BC, Canada, 2008; p. 268. [Google Scholar]
- Wilcove, D.S.; Lee, J. Using economic and regulatory incentives to restore endangered species: Lessons learned from three new programs. Conserv. Biol. 2004, 18, 639–645. [Google Scholar] [CrossRef]
- Wunder, S. Are direct payments for environmental services spelling doom for sustainable forest management in the tropics? Ecol. Soc. 2006, 11, 473–482. [Google Scholar] [CrossRef]
- Rigby, D.; Cáceres, D. Organic farming and the sustainability of agricultural systems. Agric. Syst. 2001, 68, 21–40. [Google Scholar] [CrossRef]
- Ostrom, E.; Cox, M. Moving beyond panaceas: A multi-tiered diagnostic approach for social-ecological analysis. Environ. Conserv. 2010, 37, 451–463. [Google Scholar] [CrossRef]
- Escobar, A. Culture sits in places: Reflections on globalism and subaltern strategies of localization. Political Geogr. 2001, 20, 139–174. [Google Scholar] [CrossRef]
- Marsden, T. Sustainable place-making for sustainability science: The contested case of agri-food and urban–rural relations. Sustain. Sci. 2013, 8, 213–226. [Google Scholar] [CrossRef]
- Horlings, L.G.; Horlings, L.G. Values in place; a value-oriented approach toward sustainable place-shaping. Reg. Stud. Reg. Sci. 2015, 2, 257–274. [Google Scholar] [CrossRef]
- Klassen, S.E.; Wittman, H. Place-based food systems: “Re-valuing local” and fostering socio-ecological sustainability. In Sustainable Food Futures: Multidisciplinary Solutions; Duncan, J., Bailey, M., Eds.; Routledge: Abingdon, UK, 2017. [Google Scholar]
- Seufert, V.; Ramankutty, N.; Mayerhofer, T. What is this thing called organic?—How organic farming is codified in regulations. Food Policy 2017, 68, 10–20. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Sørensen, M.; Pedersen, S.M.; Weiner, J. Feeding the world: Genetically modified crops versus agricultural biodiversity. Agron. Sustain. Dev. 2013, 33, 651–662. [Google Scholar] [CrossRef]
- Stubbs, T.L.; Kennedy, A.C. Microbial weed control and microbial herbicides. In Herbicides—Environmental Impact Studies and Management Approaches; Alvarez-Fernandez, R., Ed.; InTech: Rijeka, Croatia, 2012; pp. 135–166. [Google Scholar]
- Kennedy, A.C.; Johnson, B.N.; Stubbs, T.L. Host range of a deleterious rhizobacterium for biological control of downy brome. Weed Sci. 2001, 49, 792–797. [Google Scholar] [CrossRef]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Levin, S.A.; Lubchenco, J. Resilience, robustness, and marine ecosystem-based management. BioScience 2008, 58, 27–32. [Google Scholar] [CrossRef]
- Levin, S.; Xepapadeas, T.; Crépin, A.S.; Norberg, J.; de Zeeuw, A.; Folke, C.; Hughes, T.; Arrow, K.; Barrett, S.; Daily, G.; et al. Social-ecological systems as complex adaptive systems: Modeling and policy implications. Environ. Dev. Econ. 2012, 18, 111–132. [Google Scholar] [CrossRef]
- Leslie, P.; McCabe, J.T. Response diversity and resilience in social-ecological systems. Curr. Anthropol. 2013, 54, 114–143. [Google Scholar] [CrossRef] [PubMed]
- Baumgart-Getz, A.; Prokopy, L.S.; Floress, K. Why farmers adopt best management practice in the USA: A meta-analysis of the adoption literature. J. Environ. Manag. 2012, 96, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.W. Environmental crises and the metabolic rift in world-historical perspective. Org. Environ. 2016, 13, 123–157. [Google Scholar] [CrossRef]
- Clapp, J. Distant agricultural landscapes. Sustain. Sci. 2014, 10, 305–316. [Google Scholar] [CrossRef]
- Wittman, H. Reworking the metabolic rift: La Vía Campesina, agrarian citizenship, and food sovereignty. J. Peasant Stud. 2009, 36, 805–826. [Google Scholar] [CrossRef]
- Levkoe, C.Z. Towards a transformative food politics. Local Environ. 2011, 16, 687–705. [Google Scholar] [CrossRef]
- Altieri, M.A. Agroecology, Small Farms, and Food Sovereignty. Available online: http://monthlyreview.org/2009/07/01/agroecology-small-farms-and-food-sovereignty/ (accessed on 1 March 2015).
- Blesh, J.; Wittman, H. “Brasilience:” assessing resilience in land reform settlements in the Brazilian Cerrado. Hum. Ecol. 2015, 43, 531–546. [Google Scholar] [CrossRef]
- Wittman, H. Food sovereignty: A new rights framework for food and nature? Environ. Soc. Adv. Res. 2011, 2, 87–105. [Google Scholar]
- Loos, J.; Abson, D.J.; Chappell, M.J.; Hanspach, J.; Mikulcak, F.; Tichit, M.; Fischer, J. Putting meaning back into “sustainable intensification”. Front. Ecol. Environ. 2014, 12, 356–361. [Google Scholar] [CrossRef]
- Weber, C.L.; Matthews, H.S. Food-Miles and the Relative Climate Impacts of Food Choices in the United States. Environ. Sci. Technol. 2008, 42, 3508–3513. [Google Scholar] [CrossRef] [PubMed]
- Stokstad, E. Could less meat mean more food? Science 2010, 327, 810–811. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Clark, M. Global diets link environmental sustainability and human health. Nature 2014, 515, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Erb, K.H.; Lauk, C.; Kastner, T.; Mayer, A.; Theurl, M.C.; Haberl, H. Exploring the biophysical option space for feeding the world without deforestation. Nat. Commun. 2016, 7, 11382. [Google Scholar] [CrossRef] [PubMed]
- Kremen, C.; Iles, A.; Bacon, C. Diversified farming systems: An agroecological, systems-based alternative to modern industrial agriculture. Ecol. Soc. 2012, 17, 44. [Google Scholar] [CrossRef]
- Raynolds, L.T.; Murray, D.; Heller, A. Regulating sustainability in the coffee sector: A comparative analysis of third-party environmental and social certification initiatives. Agric. Hum. Values 2007, 24, 147–163. [Google Scholar] [CrossRef]
- Schipanski, M.E.; MacDonald, G.K.; Rosenzweig, S.; Chappell, M.J.; Bennett, E.M.; Kerr, R.B.; Blesh, J.; Crews, T.; Drinkwater, L.; Lundgren, J.G.; et al. Realizing resilient food systems. BioScience 2016, 66, 600–610. [Google Scholar] [CrossRef]
- Kahneman, D. Thinking, Fast and Slow; Farrar, Straus and Giroux: New York, NY, USA, 2011. [Google Scholar]
- Heath, C.; Heath, D. Switch: How to Change Things When Change Is Hard, 1st ed.; Crown Business: Danvers, MA, USA, 2010. [Google Scholar]
- Eckhardt, G.M.; Belk, R.; Devinney, T.M. Why don’t consumers consume ethically? J. Consum. Behav. 2010, 9, 426–436. [Google Scholar] [CrossRef]
- Sagoff, M. Aggregation and deliberation in valuing environmental public goods: A look beyond contingent pricing. Ecol. Econ. 1998, 24, 213–230. [Google Scholar] [CrossRef]
- Kahan, D.M.; Braman, D. Cultural cognition and public policy. Yale Law Policy Rev. 2006, 24, 149–172. [Google Scholar]
Figure 1.
A conceptual stability landscape for pest-control: Agricultural systems revert to different stability points (open circles) depending on their properties: complex pest-predator food webs (type 1 basin), or crop collapse due to pest outbreaks (type 2 basin). Agricultural systems are often maintained away from points of stability in non-equilibrium states (hatched circles) due to management (orange arrows). Systems can follow diverse pathways between stability points and non-equilibrium states in the landscape. Figure adapted from [109].
Figure 1.
A conceptual stability landscape for pest-control: Agricultural systems revert to different stability points (open circles) depending on their properties: complex pest-predator food webs (type 1 basin), or crop collapse due to pest outbreaks (type 2 basin). Agricultural systems are often maintained away from points of stability in non-equilibrium states (hatched circles) due to management (orange arrows). Systems can follow diverse pathways between stability points and non-equilibrium states in the landscape. Figure adapted from [109].
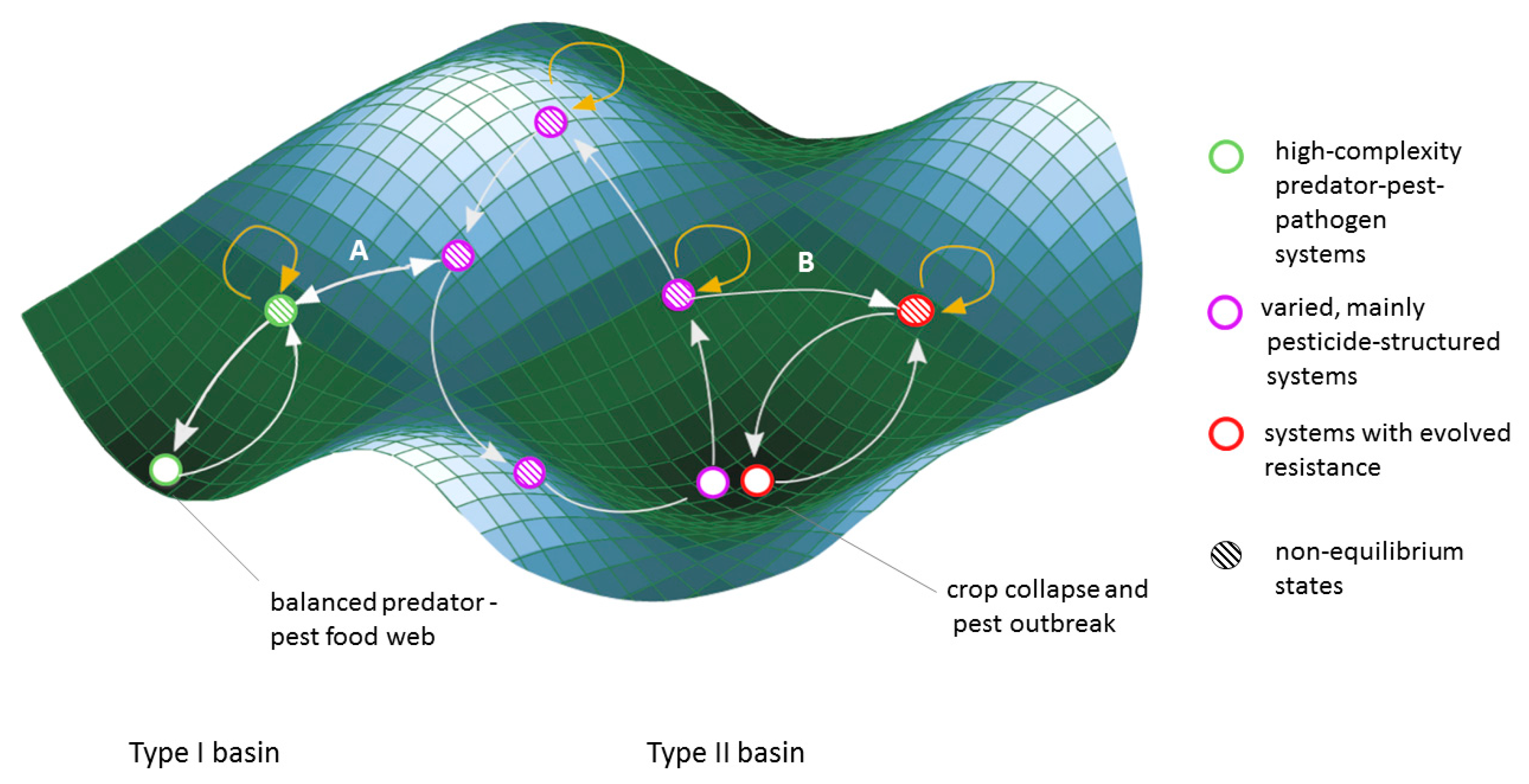

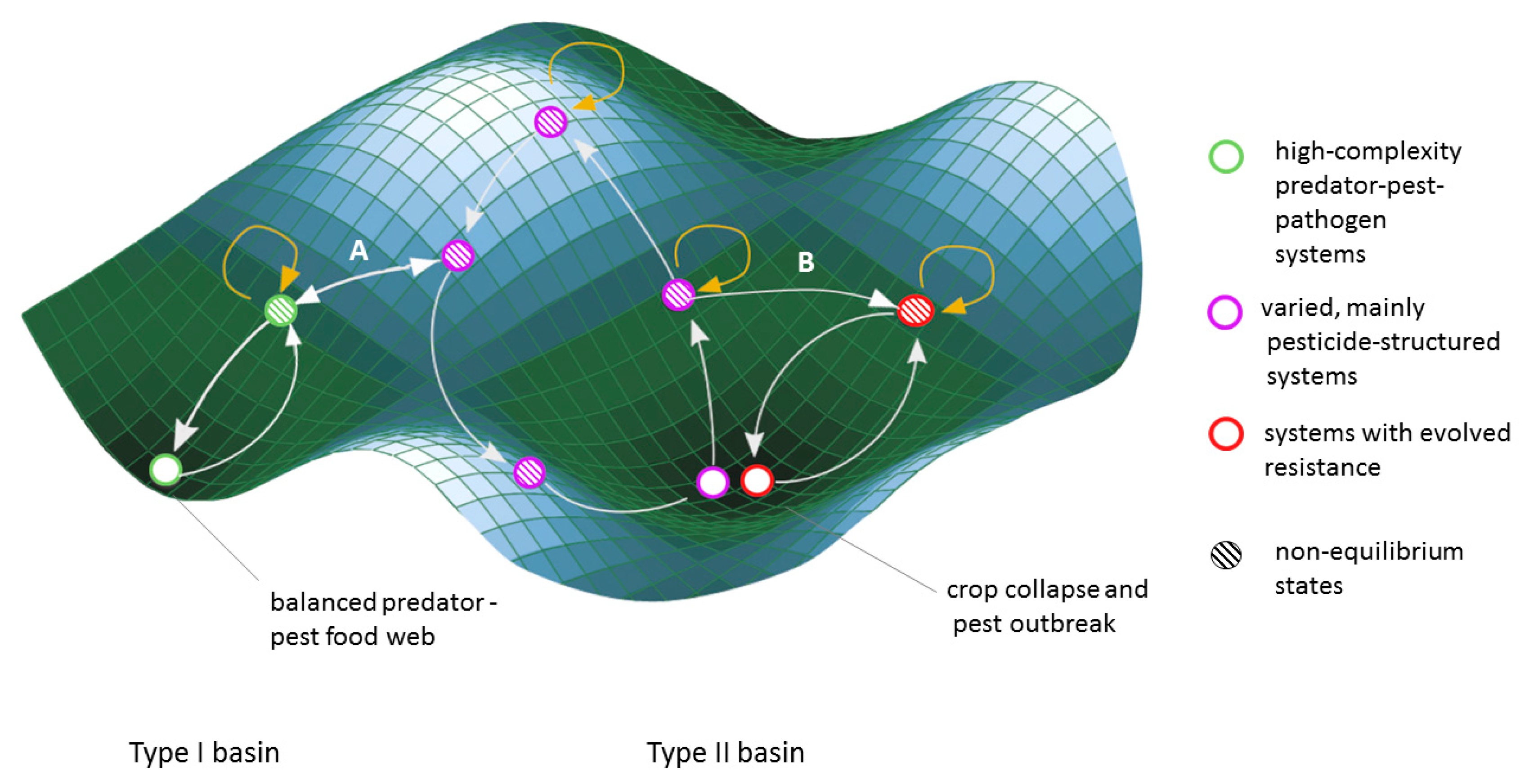{kind=link}
Table 1.
Complex Adaptive Systems Characteristics Definitions and Literature.
| CAS Characteristic | 1. Multi-Causality: There Is No Smoking Gun and No Silver Bullet | 2. Cumulative Impacts: Death by a Thousand Cuts | 3. Regime Shifts: Systems Change in Fits and Spurts, and Can Flip Unexpectedly | 4. Teleconnections: “Transporting” Impacts across Time and Space | 5. Multi-Scalarity: Drivers and Impacts Cross Scales |
|---|---|---|---|---|---|
| Definition | Any given ecological or social pattern is simultaneously the product of many different processes [ 26] | The combined total effect of multiple effects that limit the ability of people to enjoy ecosystem services [ 34] | A system can change non-linearly and non-reversibly between alternative stable states with significantly different components | Links between distant areas that are enabled via larger scale processes | Patterns can emerge at one scale via changes at other scales |
| Associated terms/concepts | Multi-causality | Cumulative impacts | Non-linearity, regime shifts, hysteresis, basins of attraction, attractors, stable states, steady states, bifurcations, oscillations, periodic behavior | Teleconnections, legacy effects, cross-system impacts | Multi-scalarity, shifting baselines, pulse and press disturbances |
| Key Literature | [ 26] | [ 35,36,37] | [ 28,38,39,40] | [ 41,42,43] | [ 26,44] |
Table 2.
Complex Adaptive Systems Characteristics Relevance to Each Case Study.
| Case Study | 1. Multi-Causality: There Is No Smoking Gun and No Silver Bullet | 2. Cumulative Impacts: Death By a Thousand Cuts | 3. Regime Shifts: Systems Change in Fits and Spurts, and Can Flip Unexpectedly | 4. Teleconnections: “Transporting” Impacts across Time and Space | 5. Multi-Scalarity: Drivers and Impacts Cross Scales |
|---|---|---|---|---|---|
| (1) Pollinators | Neonicotinoid pesticides are considered by many to be the cause of pollinator declines. While there is mounting evidence that demonstrates their toxic and sometimes lethal effects on pollinator populations, and important policy and legislative actions have been taken to reduce neonicotinoid use as a result, this one-dimensional view of pollinator challenges is problematic as it can excuse inaction, or obscure the other contributing causes. | There are several contributing causes to decreased resilience of pollinator populations, including compromised immune systems from pesticide exposure, reduced abundance and appropriateness of food sources, decreased natural and semi-natural habitat as nesting sites, increased exposure to pests and parasites, and increased environmental shocks from climate change. The intensification and extensification of agriculture has contributed to all of these causes. | The sudden collapse of honeybee colonies (coined Colony Collapse Disorder) can be considered a micro example of a systems flip. At larger scales, a sudden systems flip or collapse of pollinator populations is also possible as stressors reach critical thresholds that populations are no longer able to withstand. | Local pollinator populations around the world (both wild and managed) are being significantly impacted via the same global drivers. Land conversion of biodiverse areas such as tropical forests to agricultural land in order to feed our growing population, and the increased intensity of management (reduced biodiversity, increased input use) are all contributing to local pollinator declines. | The dynamics and stressors that are contributing to pollinator declines are happening at multiple scales, from in-field biodiversity, to global market conditions influencing farmer decision-making, to widespread habitat destruction from agricultural extensification. In the global agri-food system, agricultural production is often driven by demand from non-local markets, yet critical agro-ecosystem dynamics are limited to smaller scales (e.g., the foraging range of many pollinators is within the scale of a farm-field). |
| (2) PES | One of the benefits of PES is that, as a voluntary program, it allows policymakers to address agricultural impacts without demonstrating proof (as is often expected before prohibitive legislation). | Agricultural impacts are one set among many, but within agriculture, PES have envisioned the actions of separate farmers as separate impacts to be addressed separately by ‘buying’ behavior change (via an incremental addition of an extrinsic motivation). But such an approach misses the point that there are larger system dynamics at play, and it may be possible to intervene in such a way as to change norms, not just individual behaviours. | PES programs are sometimes looked to as solutions to problems in downstream systems. While this recognition of teleconnections and multi-scalar dynamics are welcome, it’s also the case that those downstream systems shouldn’t be expected to change linearly as a result of altered inputs via the PES. | Agricultural intensification and associated environmental impacts are a result of teleconnections from consumer demand and—in some cases—pressure from integrated value-chain retailers (e.g., Walmart). But perhaps what’s needed is to expand PES so as to directly connect improvements in farm management directly to the concerns of consumers, who often value environmental outcomes and demonstrate a willingness to buy accordingly. | PES are an example of recognizing that on-farm actions can have considerable consequences at other scales, e.g., in downstream aquatic ecosystems (Chesapeake, Golden/Tasman Bays, Gulf of Mexico). |
| (3) Pest control | Pesticide resistance is driven by multiple factors, including pressure from pest predators, availability of habitat fragments without pesticide pressure and farm and landscape scale diversity. | If pesticides have acquired resistance to multiple pests, viable pest-control options become increasingly difficult to find, and the alternative is a more severe collapse in production. | Pest control systems with multiple pest predators and competitors exist in the basin of attraction of a self-regulating, functioning food web. Intensive pesticide use can shift these systems to a different basin of attraction where crop collapse results in the event of pesticide failure. | Once pests evolve genetic pesticide resistance, the trait can travel through metapopulations and spread to areas where the resistance mutation would not have arisen. On the positive side, meta-populations of beneficial insects can help repopulate depleted areas if management changes to support their survival and connectivity. | Changes to the ecological community on a local scale (the absence of pest predators and competitors) and local selective forces (intensive pesticide use) drive the acquisition of permanent genetic changes with large scale impacts (both in time and space). |
| (4) Sediment in Tasman bay | While sedimentation is seen as a major issue in Tasman bay, it has many contributing causes. A further complication is that historical fishing practices have led to the context where sediment is as problematic as it is today. | Future climate change may reinforce current feedbacks as more intense storms may increase sediment runoff to the bay. | Changing the benthic community from a three dimensional, high bivalve biomass floor to a system with a flat silty bottom through historic fishing practices has exposed Tasman bay to sedimentation and sediment resuspension, as there does not exist the density of filter feeders to help sediment settle out. | Demand for New Zealand dairy, meat and other agricultural products in other continents can shape land use practices around Tasman bay, resulting in changes to sediment input into the bay. | Land use around the bays that are sources of sediment are regional in scale. Because New Zealand agriculture is fully subject to global markets, global demand affects land use (global scale). Ships (commercial and fishing) in the bays also resuspend sediment at a local scale. |
| (5) Scales in Puget Sound | While loss, fragmentation and destruction of salmon habitat is considered to be the limiting factor (aka smoking gun) for salmon returns, the many other impacts to salmon, including dams, climate change, and non-point source pollution as well as fisheries related impacts allows different actors to point blame elsewhere. Even if habitat is greatly improved, it is no silver bullet as the other factors could still combine to impede recovery. | Salmon face multiple impacts including from land use, dams, fisheries, and indirect global impacts such as climate change. Impacts from individual farms such as run-off and lack of riparian habitat are individually small, yet cumulatively important. | Potential collapses in salmon runs could have dramatic social, cultural and ecological consequences. | Ocean currents and migration mean that salmon may face impacts generated in distant places, including changes in ocean temperature and acidity driven by distant emissions. | Restoration projects are completed on the parcel scale and in relatively short term contracts yet seek to have large spatial scale impacts on salmon returns. Historical impacts have dramatically altered the baseline expectations of many Puget Sound residents. Yet Treaty Tribes employ a much longer term baseline going back prior to colonization and the resultant widespread landscape changes. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chapman, M.; Klassen, S.; Kreitzman, M.; Semmelink, A.; Sharp, K.; Singh, G.; Chan, K.M.A. 5 Key Challenges and Solutions for Governing Complex Adaptive (Food) Systems. Sustainability 2017, 9, 1594. https://doi.org/10.3390/su9091594
AMA Style
Chapman M, Klassen S, Kreitzman M, Semmelink A, Sharp K, Singh G, Chan KMA. 5 Key Challenges and Solutions for Governing Complex Adaptive (Food) Systems. Sustainability. 2017; 9(9):1594. https://doi.org/10.3390/su9091594
Chicago/Turabian StyleChapman, Mollie, Susanna Klassen, Maayan Kreitzman, Adrian Semmelink, Kelly Sharp, Gerald Singh, and Kai M. A. Chan. 2017. "5 Key Challenges and Solutions for Governing Complex Adaptive (Food) Systems" Sustainability 9, no. 9: 1594. https://doi.org/10.3390/su9091594
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.