Analysing the Impacts of Various Environmental Parameters on the Biodiversity Status of Major Habitats




Department of Environmental Technology, Technische Universität Berlin, Berlin 10623, Germany
*
Author to whom correspondence should be addressed.
Sustainability 2017, 9(10), 1775; https://doi.org/10.3390/su9101775
Submission received: 27 July 2017
/
Revised: 21 September 2017
/
Accepted: 26 September 2017
/
Published: 30 September 2017
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Background: Anthropogenic impacts on the environment often result in loss of biodiversity. However, the effects of different parameters (such as soil pH) on biodiversity, which can be influenced by human activities, are poorly investigated on a global scale. The paper at hand proposes an approach to examine the relationship between biodiversity and several parameters, which can be anthropogenically influenced, on a major habitat scale. The goal is to describe the relationship of a single parameter and biodiversity by means of a mathematical model within one major habitat. Methods: Geographic information system (GIS) data for the parameters “mean soil pH”, “mean share of cultivated land” and “mean temperature in June” for the whole world were overlaid with GIS data for the ecoregions proposed by the World Wide Fund for Nature (WWF). By means of the software ArcGIS 10.4 (Esri Deutschland GmbH, Kranzberg, Germany), an average parameter value per ecoregion for all considered parameters was determined. Moreover, the index “biodiversity status” was introduced and determined for every ecoregion by means of the indicators “number of species”, “number of endemic species”, and “mean share of grassland and forest” for each ecoregion. As all ecoregions can be assigned to one of the 14 major habitats (as defined by the WWF), a Kruskal–Wallis test was conducted to analyse whether the parameters and the biodiversity status’ differentiate between the 14 major habitats. A mathematical model, which depicts the impact of the parameters on biodiversity, was established by means of curve fitting. Results: The Kruskal–Wallis test reveals that a significant difference (p-value of 0.000) regarding the considered parameter and the biodiversity status exists between the different major habitats. By means of a regression analysis and curve fitting, mathematical models were developed which describe the relation of the biodiversity status and the parameters for 14 major habitats.
1. Introduction
At the tenth meeting of the conference of parties to the Convention on Biological Diversity (CBD) in 2010 in Japan, the Strategic Plan for Biodiversity 2011–2020 was developed [1]. Governments, intergovernmental and non-governmental organisations and other representatives agreed on five strategic goals and 20 Aichi targets to protect biodiversity. Some years later, the United Nations (UN) acknowledged the protection of biodiversity in two of the published Sustainable Development Goals (SDGs), i.e., number 14 “life on land” and number 15 “life below water” [2]. Protecting biodiversity is important because it affects humanity. It underpins and stabilises [3] the functions of ecological systems that provide goods and services (called ecosystem services (ESS)) securing human life [4]. The ESS include, e.g., the provision of food, regulating of floods or supporting soil formation [4].
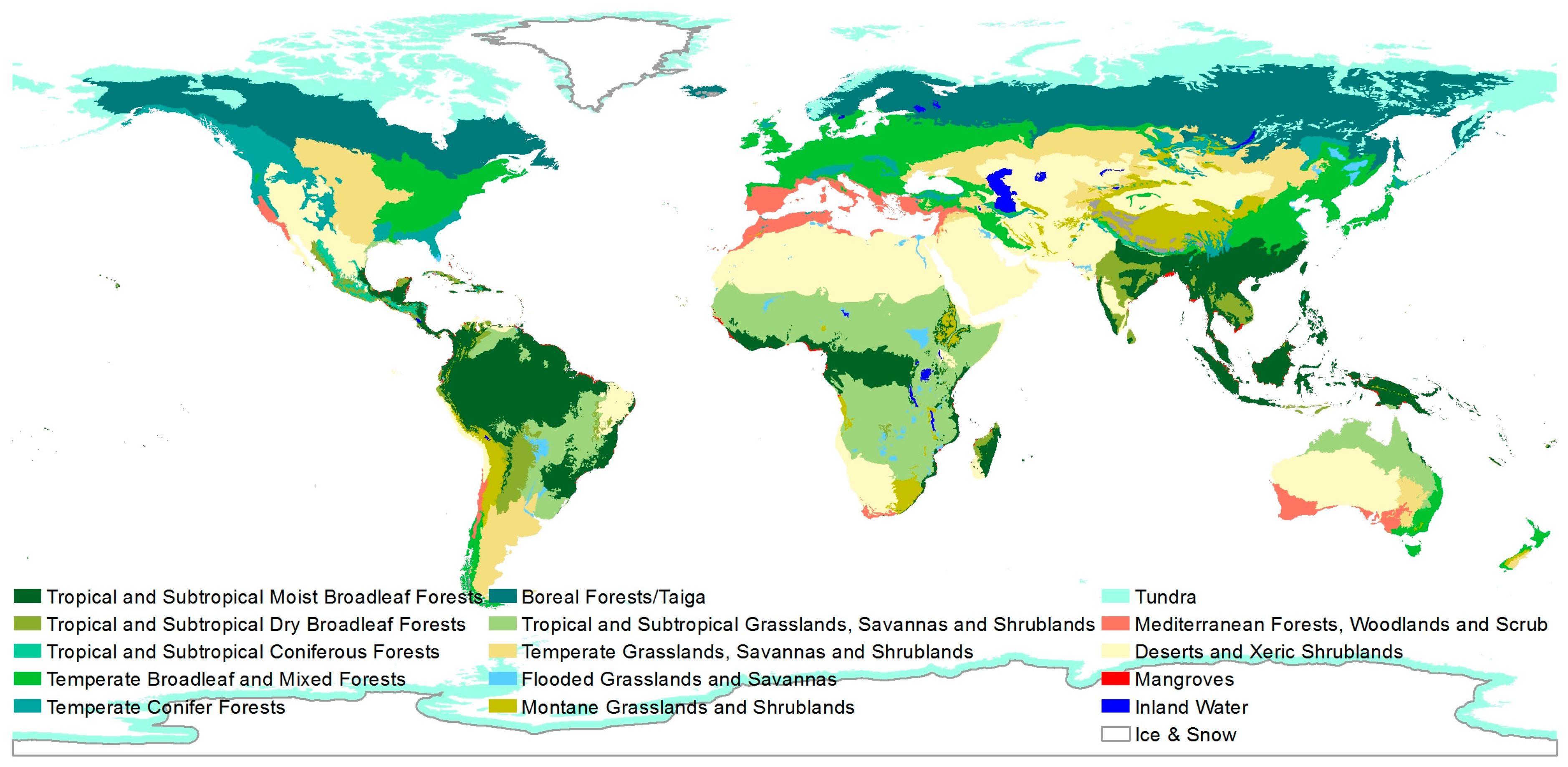
Biodiversity as such includes the genetic, the species and the ecosystem diversity (defined by the UN [5]). It can be described on different spatial resolutions. For example, the World Wild Fund for Nature (WWF) uses a resolution of 14 major habitat types (MHT) (see Figure 1) defined as “areas of the world that share similar environmental conditions, habitat structure, and patterns of biological complexity, and that contain similar communities and species adaptations” [6].
To strengthen the protection of biodiversity, the impacts of anthropogenic pressures on it have to be measurable in order to be preventable. Anthropogenic pressures include, e.g., agriculture (land use, land transformation), emission of toxic substances, greenhouse gas (GHG) emissions, fertilization (i.e., phosphorus and nitrogen emissions), light and noise emissions or destructive harvesting practices [8]. The anthropogenic pressures could have an effect, among others, on different environmental parameters. For instance the pH of soil and water can be affected by the release of acidifying substances [9], the share of cultivated land is affected by agriculture and the mean global temperature can be influenced by man-made GHG emissions [10]. In a second step, these altered environmental parameters can affect biodiversity due to, e.g., loss of species richness.
In recent years, several methods and tools have been developed to determine the impact of human activities (anthropogenic pressures) on biodiversity. One such method which has increasingly been used in the recent years is life cycle assessment (LCA) as set forth by the ISO 14040/14044 standards [11,12]. LCA is a comprehensive method, allowing for the assessment of the potential environmental impacts of products along their entire life cycle from raw material extraction through materials processing, manufacturing, distribution, use, repair and maintenance, to disposal [11]. In LCA’s impact assessment phase, the potential impacts of anthropogenic pressures on, e.g., biodiversity are described at different levels in the cause–effect chain. For example, impacts of GHG emissions are assessed at the middle of the cause–effect chain (global warming potential [13]) and at the end of the cause–effect chain at which damages to human health and biodiversity [13] are quantified.
LCA has improved the possibility to quantify and analyse the potential environmental impacts associated with any given product system (including the impact on certain biodiversity aspects). However, the modelling of cause–effect chains leading, first, from anthropogenic pressures to changes in environmental parameters and, second, leading from altered environmental parameters to biodiversity impacts still needs improvements. Most of the models used in LCA are region specific (because specific species are used to assess the impacts on biodiversity) and consider land use only. Further, they consider one aspects of biodiversity only, namely species diversity. As data for indicators measuring biodiversity (e.g., species richness, genetic diversity) is lacking, the impact assessment is possible for few regions in the world only—but biodiversity should be protected globally. [14].
To strengthen LCA in assessing impacts on biodiversity, the central goal of this paper is to analyse the relationship between environmental parameters and biodiversity allowing for a reliable description of cause–effect mechanisms. To meet this aim, three questions have to be answered. First, which parameters should be investigated? Second, how can biodiversity be measured? Third, what is the mathematical relationship between the considered parameters and biodiversity?
2. Method
As biodiversity varies in different regions, the MHTs have been used as a basis to assess the impacts of various parameters on biodiversity. The appropriateness of using the MHTs has been tested after (see Section 2.3).
Usually, the relation of parameters and biodiversity should be considered on a high spatial resolution, e.g., on an ecoregion level as recommended by Coelho and Michelsen (2014) [15]. However, data availability is a limiting factor. Thus, the effect of three parameters has been estimated on a low resolution (i.e., at MHT level) in this study. The data for the parameters and the biodiversity were collected for the 827 ecoregions and analysed with regard to every MHT because each of the 14 MHT includes several ecoregions [16].
To analyse the relationship between the parameters and biodiversity, the following steps were conducted.
2.1. Parameters for Each Ecoregion
Many parameters affect biodiversity, but for most of them data availability on a global level are limited. Therefore, within this study, only parameters were examined for which data is available in a Geographical Information System (GIS) layer for the whole world. The parameters “soil pH”, “share of cultivated land” and “temperature in June” were considered. Data for other parameters, which also might affect the biodiversity, were not available for the whole world or with the resolution which is needed (i.e., on an ecoregion level) for the aim of the study. GIS data for the considered parameters are available as raster data for the whole world from the following sources:
- (a)
- The “soil pH” was extracted from the Harmonized World Soil Database (HWSD) where the data is given with a grid cell resolution of 1 km [17].
- (b)
- The “share of cultivated land” with a grid cell resolution of 10 km is published online as supplementary materials to the HWSD [17].
- (c)
- The “mean temperature in June” with a grid cell resolution of 1 km is published online by Hijmans et al. [18].
Besides the GIS data for the parameters, GIS data for the size, shape and location of the ecoregions proposed by Olsen et al. [16] were collected. These data are available as polygon data at the ecoregion database website [7]. The size of the ecoregions varies from few square kilometres to some thousands square kilometres [7]. After collecting the GIS data, the different GIS layers (for the parameters (raster data) and the ecoregions (polygon data)) were overlaid and merged by means of the zonal statistic function in ArcGIS 10.4 [19]. Thus, all parameter grid cell values which are located in one ecoregion polygon were summed up. This resulted in mean values for the considered parameters for every ecoregion.
2.2. Biodiversity Status (BS) for Each Ecoregion
Several indicators for the estimation of biodiversity are proposed by, e.g., [20,21,22] but for only few of them data are available. To define biodiversity on a more generic level, a new index is defined within this study. The index, called biodiversity status (BS), was developed with indicators for which data are available for all ecoregions. It is meant to present the current situation of the genetic diversity (e.g., gene pool), the species diversity (e.g., species number) and the ecosystem diversity (e.g., landscape) in one ecoregion. Thus, all levels of biodiversity are included. For the BS, the “number of species per km2” (data from the ecoregion database website [7], which includes the taxa amphibians, reptiles, birds and mammals); “number of endemic species per km2” (data from the ecoregion database website [7], which also includes the taxa amphibians, reptiles, birds and mammals); and “mean share of grassland and forest” per ecoregion were used. The last indicator was calculated in the same way as the “mean share of cultivated land” (see Section 2.1 with published data from the HWSD [17].
As there is no definition of “good” and “bad” biodiversity, the index was normalised in order to have a highest BS (which has the value of one) and a lowest BS (which has the value of zero). For this, the individual indicators were normalised, summed up to the BS and normalised again (data and calculation steps are presented in Supplementary Materials (Table S1)). The outcome is a BS between zero and one for every ecoregion. The calculation of the BS is presented in Equation (1).
- SER,n = normalised (n) number of species per km2 per ecoregion (ER);
- ESER,n = normalised (n) number of endemic species per km2 per ecoregion (ER);
- SGFER,n = normalised (n) mean share of grassland and forest per ecoregion (ER);
- min = minimum value of the sum of the indicators of all ecoregions; and
- max = maximum value of the sum of the indicators of all ecoregions.
2.3. Kruskal–Wallis Test
Biodiversity varies even in small ecosystems, but, as mentioned above, data for such a high spatial resolution are not available. To have any differentiation regarding biodiversity and not assuming even distribution of biodiversity all over the world, the MHTs were chosen as the considered regions. A Kruskal–Wallis test was performed using SPSS 24 [23] to prove that this differentiation is appropriate. The Kruskal–Wallis test is a statistical analysis to compare two or more groups (in this case MHTs) and to find significant differences [24]. After this test, a pairwise comparison was performed to highlight which MHTs differentiate from each other.
2.4. Curve Fitting
To analyse the relation of the BS and the three parameters, curve fitting was performed by means of SPSS 24 on a MHT level. This is possible because every MHT consists of several ecoregions. Each ecoregion (i.e., the data for the BS and the parameters) represents one point in the curve estimation models for the respective MHT. The best curve estimation model (depending on the coefficient of determination, the p-value (to characterise the significance) and a reasonable justification) serves as a function which describes the influence of the considered parameter on the BS within the considered MHT.
3. Results
Background data are presented in the Supplementary Materials (Tables S1 and S2), whereas results in the order used in the Method Section are presented herewith.
3.1. Parameters for Each Ecoregion
Overlaying the GIS data of the considered parameters with the GIS data of the ecoregions resulted in: (a) a “mean soil pH”; (b) a “mean share of cultivated land”; and (c) a “mean temperature in June” for every ecoregion.
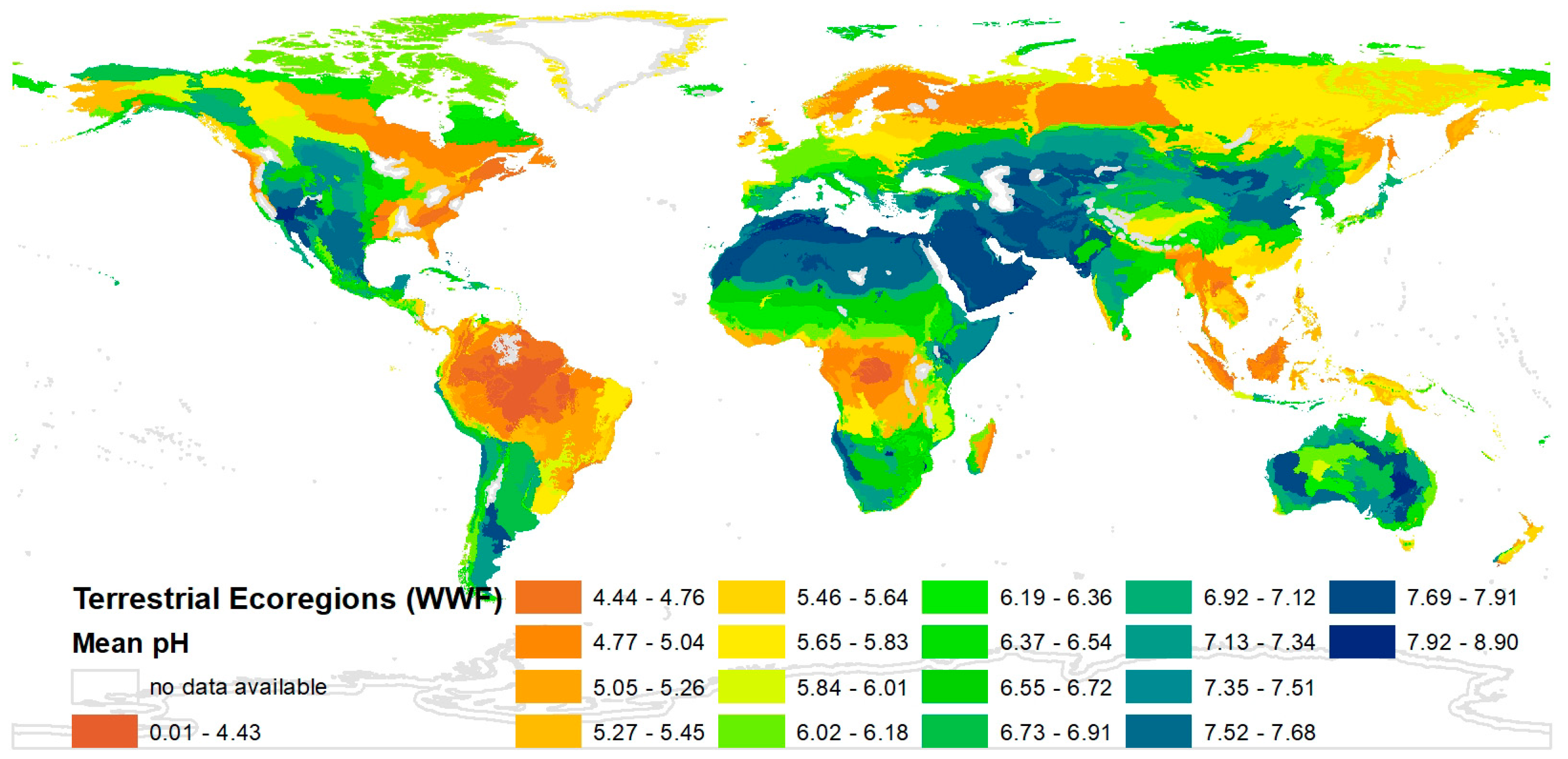
The map for the “mean soil pH” in every ecoregion is presented as an example in Figure 2. The range of the “mean soil pH” in the respective ecoregions reaches from three (orange) to nine (blue), but most ecoregions have a value between five and seven (see Figure 2). For some ecoregions the “mean soil pH” was not calculable (framed in grey). The maps of the parameters “mean share of cultivated land” and “mean temperature in June” are presented in the Supplementary Materials (Documents S3 and S4). It was possible to generate mean values of the parameters for 748 ecoregions.
3.2. BS for Each Ecoregion
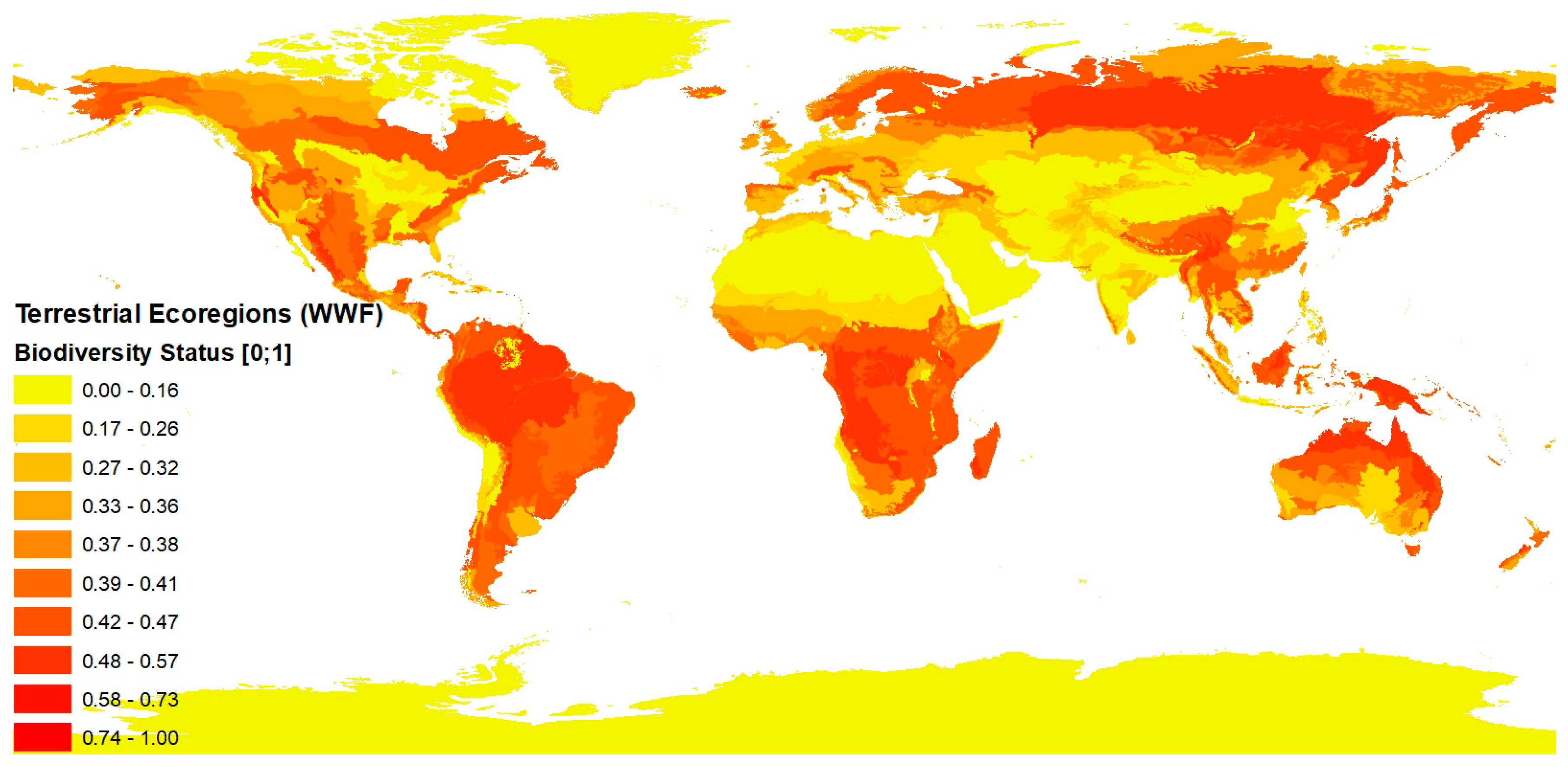
The normalised BS for all ecoregions is presented as map in Figure 3. The more species per km, the more endemic species per km and the more forest and grassland in one ecoregion, the higher is the value of the index, thus, the better is the BS in the respective ecoregion. The optimum of the BS is one; the worst possible BS is zero, as described in Section 2.2.
3.3. Kruskal–Wallis Test
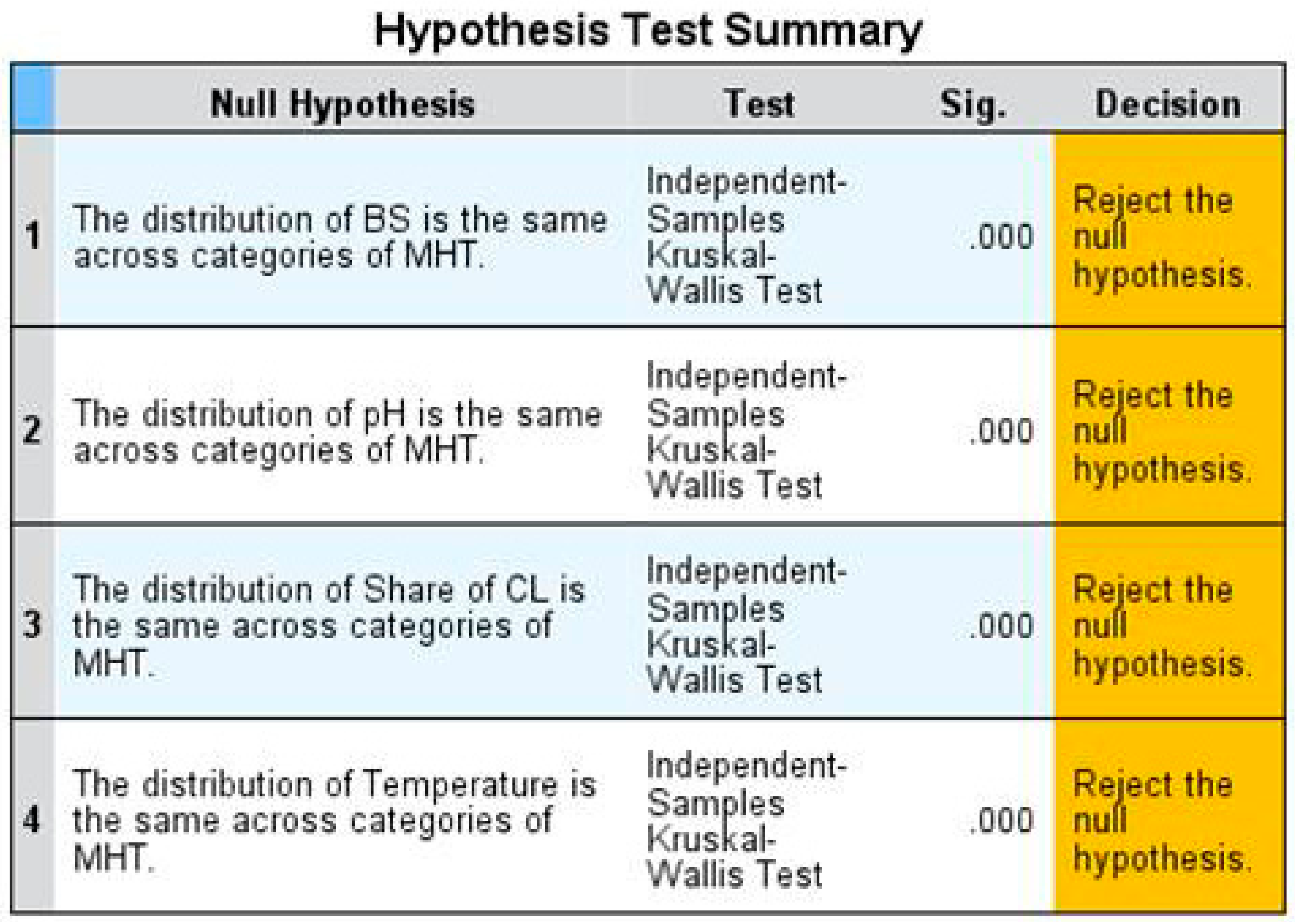
The result of the Kruskal–Wallis test is shown in Figure 4. The null hypothesis, which has to be tested, says that the distribution of the considered parameters and the BS is the same across the MHTs. In our case, the null hypothesis was rejected in all cases. The p-value (displayed in the third column) is 0.000, which means that the chance that the distribution of the parameters and the BS is the same across the MHTs is lower than one per cent.
To be more specific with regard to the differences of the MHTs, a pairwise comparison was performed. These results are displayed in the Supplementary Materials (S1). The most differences across the MHTs are regarding the parameters “mean share of cultivated land” and “mean temperature in June”. The BS is the variable that differentiates the least across the MHTs.
3.4. Curve Fitting
Based on the abovementioned results, it is reasonable to apply curve fitting for the parameters “mean soil pH”, “mean share of cultivated land” and “mean temperature in June” in the respective MHTs.
3.4.1. Curve Fitting for all MHTs
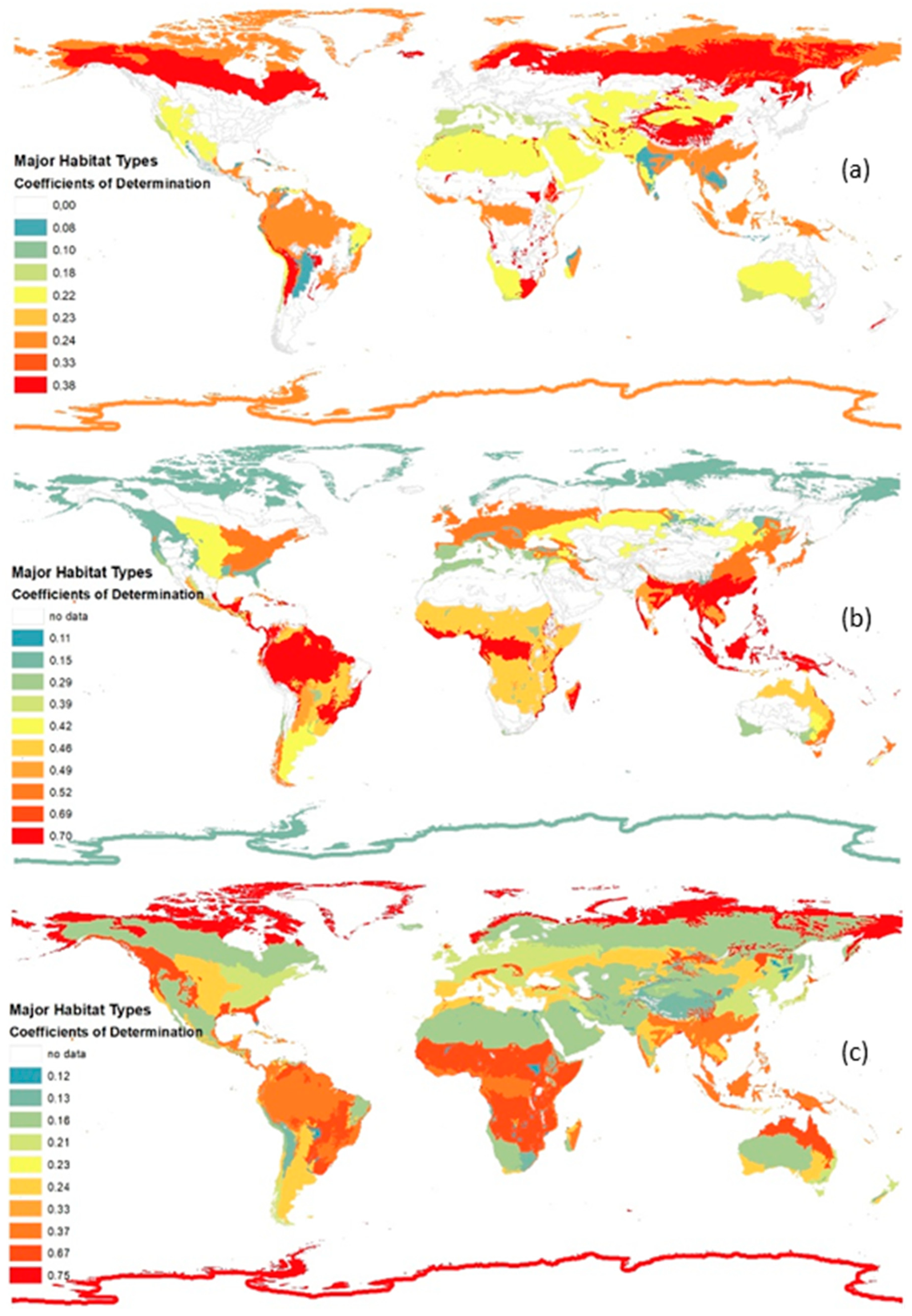
Different curve estimation models (cubic, quadratic, linear, etc.) are applied to model the relation of the BS and the individual parameters in each MHT. A coefficient of determination is calculated for each curve estimation model. The coefficients of determination for the best fitting curve estimation models of the BS and the parameters for each MHT are presented in Figure 5.
The results of the curve fitting for the different MHTs differentiate substantially. The coefficients of determination vary from low (e.g., lower than 0.013, see Supplementary Materials (S2) MHT8) to high (e.g., 0.747, see MHT11). The quadratic model provides the highest coefficient of determination for most of the MHTs. The models that fit the best to the different MHTs are marked red in the Supplementary Materials (S2).
In this paper, two examples are displayed in more detail (for MHT1 “Tropical and Subtropical Moist Broadleaf Forests” and MHT11 “Tundra”).
3.4.2. Curve Fitting MHT1—Tropical and Subtropical Moist Broadleaf Forests
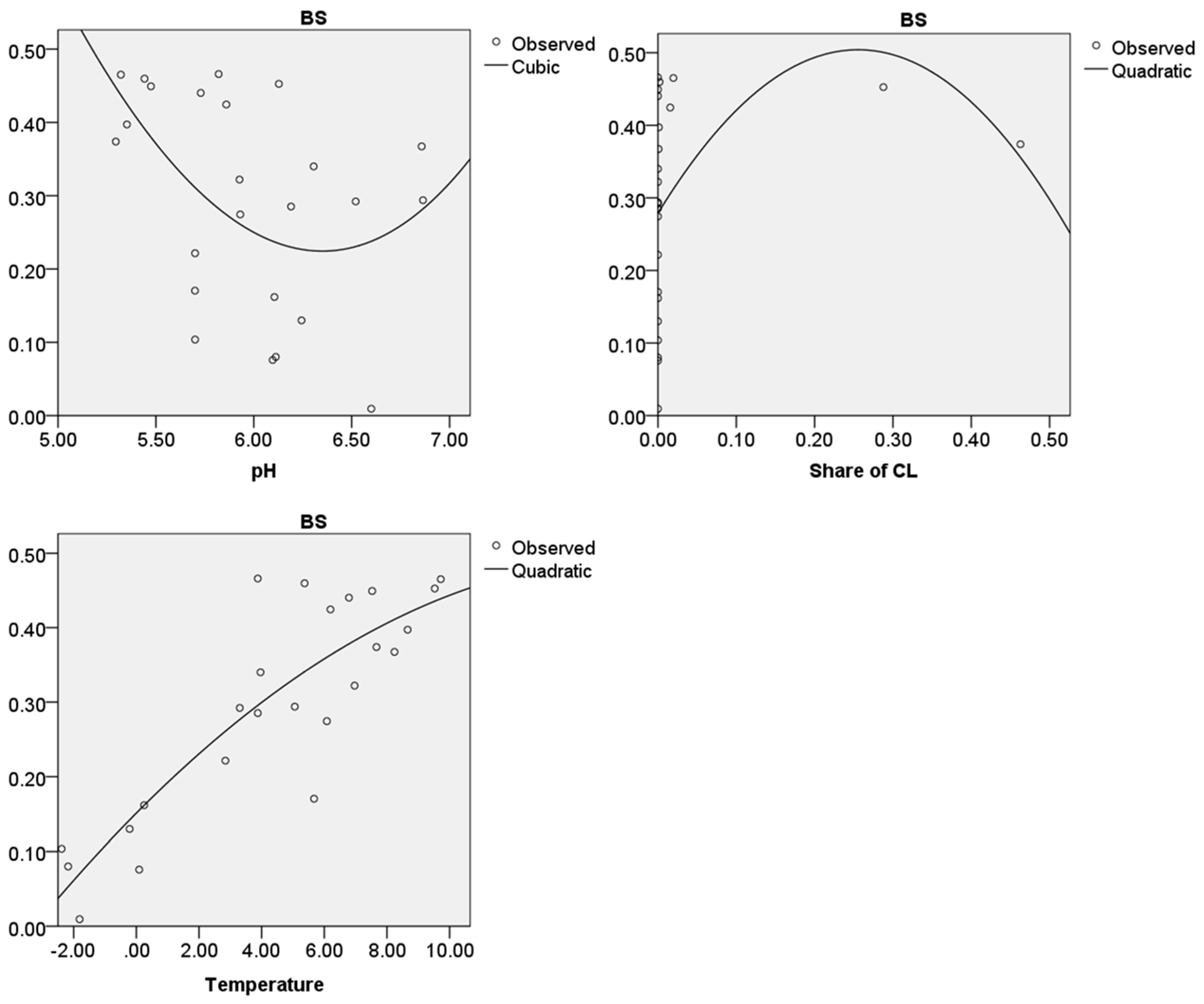
For 204 ecoregions, which are located in MHT1 (presented as “observed” in Figure 6), a “mean soil pH”, a “mean share of cultivated land”, a “mean temperature in June” and a BS could be calculated. The results for the best curve estimation models of the parameters and the BS are presented in Figure 6. The coefficients of determination, the other curve estimation models and the regression coefficients for the different models are presented in the Supplementary Materials (S2).
For the BS and the “mean soil pH” the curve, estimation model with the highest coefficient of determination (0.226) is the cubic model within the interval of pH 3–8. The function calculated by SPSS which fits the best in order to present the relation of the BS and the pH is shown in Equation (2). “x“ presents the parameter “mean soil pH”.
Regarding the BS and the “mean share of cultivated land” the curve estimation model with the highest coefficient of determination (0.700) is the quadratic model within the interval of 0–60% of cultivated land. The function is presented in Equation (3). “x” presents the parameter “mean share of cultivated land”.
The curve estimation model with the highest coefficient of determination (0.330) for the BS and the “mean temperature in June” is the cubic model within the interval of 10–35 °C. The related function is presented in Equation (4). “x” presents the parameter “mean temperature in June”.
The p-values describing the significance (available in the Supplementary Materials (S2)) for all suggested models are the same, namely 0.000. This means that the chance that the models do not explain the relation between the considered parameter and the BS is lower than one per cent.
3.4.3. Curve Fitting MHT11—Tundra
Twenty-four ecoregions are located in the MHT “Tundra”. The results for the best curve estimation models of the parameters and the BS are presented in Figure 7. The coefficients of determination and the other curve estimation models are presented in the Supplementary Materials (S2).
In the interval of pH 5–7, the curve estimation model with the highest coefficient of determination (0.244) is the cubic model. The related function is presented in Equation (5). “x” presents the parameter “mean soil pH”. The p-value for this model is 0.053. This means that the chance that the relation of the BS and “mean soil pH” cannot be explained via the suggested model is 5.3%.
The relation of the BS and the “mean share of cultivated land” can be displayed as a quadratic model (with a coefficient of determination of 0.108) by means of the function presented in Equation (6) in the interval of 0–50%. The p-value for this model is 0.301. Thus, there is a 30.1% chance the model does not explain the relation of the BS and the “mean share of cultivated land”.
By means of a quadratic model the relation of the BS and the “mean temperature in June” can be explained in the interval from −3 to 10 °C. The coefficient of determination for the suggested function (see Equation (7)) is 0.747. The p-value for this model is 0.000.
4. Discussion and Conclusions
4.1. Parameters for Each Ecoregion
The “mean soil pH” varies a lot in the different regions, but it can be seen in Figure 2 that usually neighbouring areas have similar soil pH values. As a tendency, it can be seen that the higher the population density the higher the “mean share of cultivated land” (see Figure S2) in different areas, which is not surprising. Data for the “mean temperature” were available for all months. It would not be meaningful to use the average data for one year or months for which the temperature lies in between maxima and minima of the yearly temperature (e.g., months in autumn and spring) because the differences are not high all over the world. Thus, the month June was chosen. It would be possible to use other months. A comparison of the results of a correlation analysis between the mean temperature and the BS with data for January and for June shows only insignificant differences (the correlation coefficient is |0.206| for January and |0.216| for June).
The accuracy of the mean parameter values per ecoregion can be improved by using raster data with higher resolutions. This would mean to have more exact values for different region. However, such data were not available at the time the study was conducted.
4.2. Measuring Biodiversity
The BS presented in Figure 3 shows biodiversity hotspots all over the world. The red marked ecoregions (i.e., high BS which means high biodiversity) are located, e.g., in South America and south-central China. The hotspots overlap with the hotspots identified by Myers et al. (2000) [25].
As already mentioned, the UN defined biodiversity as genetic, species and ecosystem diversity [5]. Within this study, these levels were implicitly considered (as completely as possible for the estimation of the BS) by selecting the appropriate indicators. Species diversity, for some taxa only, is covered by the “number of species per km” and the “number of endemic species per km”, while ecosystem diversity is covered by the “share of grassland and forest”. Thus far, genetic diversity is not covered explicitly, but is included implicitly in the indicators for species and ecosystem diversity because it is assumed that the genetic diversity increases with increasing number of (endemic) species and increasing number of ecosystems (i.e., grassland and forest). However, as this is not proven and only an assumption, it would be more scientifically robust to use an indicator for genetic diversity directly, but data are not yet available for such an indicator [26]. Within the index of the BS equal weighting of all indicators was applied because there is no consensus within the research community which level of biodiversity is more important [27].
4.3. Kruskal–Wallis Test
The Kruskal–Wallis test shows that there is a statistically significant difference in the distribution of the BS, the “mean soil pH”, the “share of cultivated land” and the “mean temperature in June” across all MHTs. Even though the pairwise comparison of the MHTs regarding the respective variables (i.e., the BS and the parameters) reveals that the distribution of the BS and the parameters is the same in few MHTs (see Supplementary Materials S1), it is relevant to differentiate the BS and the parameters on a MHT and not to analyse the impacts on a global level without any differentiation.
4.4. Curve Fitting
In most MHTs the parameter “mean share of cultivated land” has the highest coefficient of determination (see Figure 5). It seems that this parameter has the most effect on biodiversity (of the considered parameters), which supports the hypothesis that land use can be regarded as the main pressure on biodiversity [28]. However, in some MHTs, the parameters “mean temperature in June” and “mean soil pH” have a moderate to high coefficient of determination (e.g., 0.747 in MHT11 and 0.383 MHT10, respectively). Thus, these two parameters also have a moderate to strong relation to biodiversity.
In general, most of the obtained coefficients of determination are lower than 0.700 (see Figure 5). For these models the strength of relationship cannot be stated as very strong. However, it is still justifiable to use the different models to explain the relation of the BS and the considered parameters. In this study the relation of the BS and one parameter only, e.g., “mean soil pH”, was considered. In reality, many parameters affect the biodiversity in a cumulative manner. However, so far it has not been possible to analyse the interrelation of different parameters and their cumulated impacts on biodiversity because of a lack of data. As the study at hand analysed the impact of individual parameters on the BS, the coefficient of determination is not expected to be high. Therefore, the models are still valuable. Additionally, the p-value of the chosen models is mostly lower than 0.05, i.e., the chance that the respective model does not explain the relation of the considered parameter and the BS is lower than five per cent. Thus, most of the developed models are significant. However, for some models, the p-value is higher (see MHT11 BS and “mean share of cultivated land”). For these models, one has to decide by means of the coefficient of determination if it is reasonable to use the model to explain the relation of the considered parameter and the BS or not. In general, for some MHTs and parameters no curve estimation model fits because neither the coefficient of determination is high, nor the p-value is low (e.g., see Supplementary Materials (S2) MHT5 BS and “mean soil pH”). One example is the relation of the BS and the “mean share of cultivated land” in MHT11 (displayed in Figure 7). The coefficient of determination is relatively low (0.108) while the p-value is high (0.301) for the quadratic model. Thus, the curve estimation model does not appear reliable. The other models for this relation have even worse coefficients of determination and p-values or do not seem to be reasonable (see Supplementary Materials (S2)). A case specific decision has to be made whether a curve estimation model is useful or not.
The choice of the best curve estimation model for functionalising of the relation of the parameter and the BS has been made depending on the coefficient of determination, the significance and a reasonable justification. For example, even though the coefficient of determination of the cubic model for the relation of the BS and the “mean share of cultivated land” in MHT11 is higher (0.222) than the coefficient of determination for the quadratic model (0.108), it is reasonable to use the quadratic model. The cubic model would imply that, from a mean share of cultivated land of 40%, the BS increases with increasing mean share of cultivated land. As this is not justifiable, it was decided by the authors of this study that the quadratic model is more reasonable because the BS decreases from a mean share of cultivated land of 30%, which seems to be more reasonable.
All curve estimation models are restricted to a specific interval for the considered parameter. This is because the values of the considered parameters lie usually in specific intervals depending on the considered MHT. Thus, the calculation of the BS in dependence of the considered parameter in a specific MHT is only possible in the respective interval. For example, the curve estimation models of the parameter “mean temperature in June” differentiate regarding their temperature intervals in the different MHTs (e.g., MHT6, 4–14 °C; and MHT9, 10–35 °C).
The results of the curve fitting differentiate greatly between the different MHTs because of the differences in the number of ecoregions which are located in the MHTs. The number varies from 16 (see Supplementary Materials (S2) MHT3) to 204 (see MHT1). The more ecoregions, the more data are available and, thus, the more meaningful is the regression analysis and the curve fitting.
4.5. Limitations and Conclusions
Many different pressures on biodiversity exist, but data availability was decisive for the selection of the parameters for this study. Additionally, the data used for this study (i.e., data for the considered parameters and data for the indicators defining the BS) are secondary (i.e., they were not collected by the authors of this paper). For these data, uncertainty exists. Such uncertainty cannot be decreased by the user and has to be accepted as is. Moreover, the time representativeness of the data is different. For instance, the data for the “mean soil pH” are from 2012, whereas the data for the “mean temperature in June” are for a period from 1950 to 2000.
Usually an ecosystem depends on so-called limiting factors (e.g., water or nutrients). These are decisive factors that are crucial for the development of an ecosystem. Within this study, it was not investigated whether a parameter could be decisive or not because of a lack of data. However, it would be valuable to analyse the influence of decisive factors on a MHT level, too.
The analysis on a MHT level is controversial and impacts on biodiversity are usually considered on a higher resolution (e.g., ecosystems). An assessment on such a low resolution implies great simplifications and assumptions. Even on an ecoregion level, huge differences exist regarding biodiversity [15]. However, data on such a high resolution are not available. Thus, assumptions have to be made to assess impacts on biodiversity all over the world. The approach suggested in this paper is a simplification for estimating the influence of various parameters on biodiversity, but it is useful, e.g., to assess and compare impacts on a global level, as it is done in LCA. Two other approaches were also considered in this study (no differentiation at all and differentiation on a continent level). Firstly, as global data are available, an analysis could be done without differentiation on a MHT level. However, the results of the Kruskal–Wallis test and the curve fitting show obvious differences across the MHTs. For example, the range and the optimum values of the parameters differentiate a lot in the different MHTs. Additionally, since the MHT distinction is already a simplification, the analysis on a global level would not be reasonable. Secondly, it was considered to distinguish between the MHTs and the continents in combination, i.e., analysing the sections of a MHT located on different continents individually. However, for this, not enough data points are available because some areas would contain only few ecoregions. Thus, fragmented MHTs were considered as one MHT.
Despite the uncertainty of the suggested approach, this study provides a concept of how to assess impacts of several parameters, which can be anthropogenically influenced, on biodiversity on a MHT level for the whole world. The results (i.e., curve estimation models) can be used, e.g., in life cycle impact assessment. By describing changes in biodiversity as a consequence of changes in (anthropogenically influenced) parameters considered here, the curve estimation models can be a central part of impact assessment models (i.e., they can be used to develop the needed cause effect chain), which are urgently needed to comprehensively consider biodiversity impacts in LCA. It would help to identify and improve impact hotspots of production processes on biodiversity on a global level. However, a temporal resolution is not considered. The approach can be used, e.g., by policy makers for the identification of current pressures on biodiversity and, therefore, for their prevention. The strategic goal to “reduce the direct pressures on biodiversity and promote sustainable use” from the Strategic Plan for Biodiversity 2011–2020 [1] could be addressed.
Further research should be undertaken in several directions. First, data for other parameters, such as nitrogen concentration or water content, which affect biodiversity, should be generated. Second, more data points with regard to the parameters already used should be provided to enhance the certainty of the curve estimation models. Third, the BS should be developed further. Research on biodiversity indicators is currently developing towards the use of functional biodiversity [29,30] and ESS instead of the levels of biodiversity defined by the UN [5]. However, as this research is in an early stage, and data are not available, in this study, the common understanding of biodiversity (i.e., the three levels of biodiversity) was used as a basis for the development of the BS. Fourth, the interrelation of the different parameters and their cumulative impact on biodiversity should be investigated. For this a multi regression analysis can be used. The main problem that has to be solved is to determine the mathematical relationship of the parameters. This step will be undertaken in further research by the authors.
Supplementary Materials
The following supplementary materials are available online at www.mdpi.com/2071-1050/9/10/1775/s1, Excel Sheet S1: Calculation of the BS and results of the Kruskal–Wallis test, Excel Sheet S2: Results for the regression analysis and curve estimation models, Word Documents S3 and S4: GIS maps and explanations for the parameters “mean share of cultivated land” and “mean temperature in June”.
Acknowledgments
The authors thank the “Berlin Funding for Graduates” for their financial support. We acknowledge support by the German Research Foundation and the Open Access Publication Funds of Technische Universität Berlin.
Author Contributions
L.W. did the literature/data review, the data analysis and the method development. M.B., N.M. and M.F. contributed to the method development, the discussion of the results and to the design of the study. All authors proofread and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Secretary of the Convention on Biological Diversity (SCBD). Global Biodiversity Outlook 4; Secretary of the Convention on Biological Diversity: Montréal, QC, Canada, 2014. [Google Scholar]
- United Nations (UN). Sustainable Development Goals. Available online: https://sustainabledevelopment.un.org/topics/sustainabledevelopmentgoals (accessed on 2 June 2017).
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Naewani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A. Corrigendum: Biodiversity loss and its impact on humanity. Nature 2012, 489, 326. [Google Scholar] [CrossRef]
- Millennium Ecosystem Assessment (MEA). Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- United Nations (UN). Convention on Biological Diversity. Conf. Adopt. Conv. Biol. Divers. Rio de Janeiro, Brazil; 1992. Report No.: 1760 UNTS 79; 31 ILM 818. Available online: https://www.cbd.int/convention/text/ (accessed on 7 November 2013).
- World Wide Fund For Nature (WWF). Ecoregions. Available online: http://www.worldwildlife.org/biomes (accessed on 16 June 2016).
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World, Database. 2001. Available online: http://www.worldwildlife.org/publications/terrestrial-ecoregions-of-the-world (accessed on 3 March 2016).
- Secretary of the Convention on Biological Diversity (SCBD). Global Biodiversity Outlook 3; Secretary of the Convention on Biological Diversity: Montréal, QC, Canada, 2010. [Google Scholar]
- Bouwman, A.F.; Van Vuuren, D.P.; Derwent, R.G.; Posch, M. A global analysis of acidification and eutrophication of terrestrial ecosystems. Water Air Soil Pollut. 2002, 141, 349–382. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M.; et al. Technical Summary. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- International Organization for Standardization (ISO). ISO 14040, Environmental Management—Life Cycle Assessment—Principles and Framework (ISO 14040:2006); ISO: Brussels, Belgium, 2006. [Google Scholar]
- International Organization for Standardization (ISO). ISO 14044, Environmental Management—Life Cycle Assessment—Requirements and Guidelines (ISO 14044:2006); ISO: Brussels, Belgium, 2006. [Google Scholar]
- Klöpffer, W.; Grahl, B. Life Cycle Assessment; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2014. [Google Scholar]
- Winter, L.; Lehmann, A.; Finogenova, N.; Finkbeiner, M. Including biodiversity in life cycle assessment—State of the art, gaps and research needs. Environ. Impact Assess. Rev. 2017, 67, 88–100. [Google Scholar] [CrossRef]
- Coelho, C.R.V.; Michelsen, O. Land use impacts on biodiversity from kiwifruit production in New Zealand assessed with global and national datasets. Int. J. Life Cycle Assess. 2014, 19, 285–296. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- FAO/IIASA/ISRIC/ISS-CAS/JRC. Harmonized World Soil Database, version 1.2; FAO: Rome, Italy, 2012. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- ArcGIS. Esri. 2017. Available online: https://www.esri.de/landingpages/arcgis-10-4 (accessed on 24 July 2017).
- Joint Nature Conservation Committee. Indicators for Biodiversity. 2015. Available online: http://jncc.defra.gov.uk/page-4233 (accessed on 7 December 2015).
- Bundesamt für Umwelt Schweiz. Indicators for Biodiversity. 2015. Available online: http://www.bafu.admin.ch/umwelt/indikatoren/index.html?lang=de (accessed on 7 April 2015).
- Biodiversity Indicators Partnership (BIP). Indicators for Biodiversity. 2015. Available online: http://www.bipindicators.net/globalindicators (accessed on 7 December 2015).
- SPSS. IBM. 2017. Available online: https://www-01.ibm.com/software/de/stats24/ (accessed on 17 July 2017).
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Penman, T.D.; Law, B.S.; Ximenes, F. A proposal for accounting for biodiversity in life cycle assessment. Biodivers. Conserv. 2010, 19, 3245–3254. [Google Scholar] [CrossRef]
- Failing, L.; Gregory, R. Ten common mistakes in designing biodiversity indicators for forest policy. J. Environ. Manag. 2003, 68, 121–132. [Google Scholar] [CrossRef]
- Haines-Young, R. Land use and biodiversity relationships. Land Use Policy 2009, 26, S178–S186. [Google Scholar] [CrossRef]
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user’s guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef]
- Stuart-Smith, R.D.; Bates, A.E.; Lefcheck, J.S.; Duffy, J.E.; Baker, S.C.; Thomson, R.J.; Stuart-Smith, J.F.; Hill, N.A.; Kinimonth, S.J.; Airoldi, L.; et al. Integrating abundance and functional traits reveals new global hotspots of fish diversity. Nature 2013, 501, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Major habitat types proposed by the World Wild Fund for Nature (WWF) [7].
Figure 1.
Major habitat types proposed by the World Wild Fund for Nature (WWF) [7].
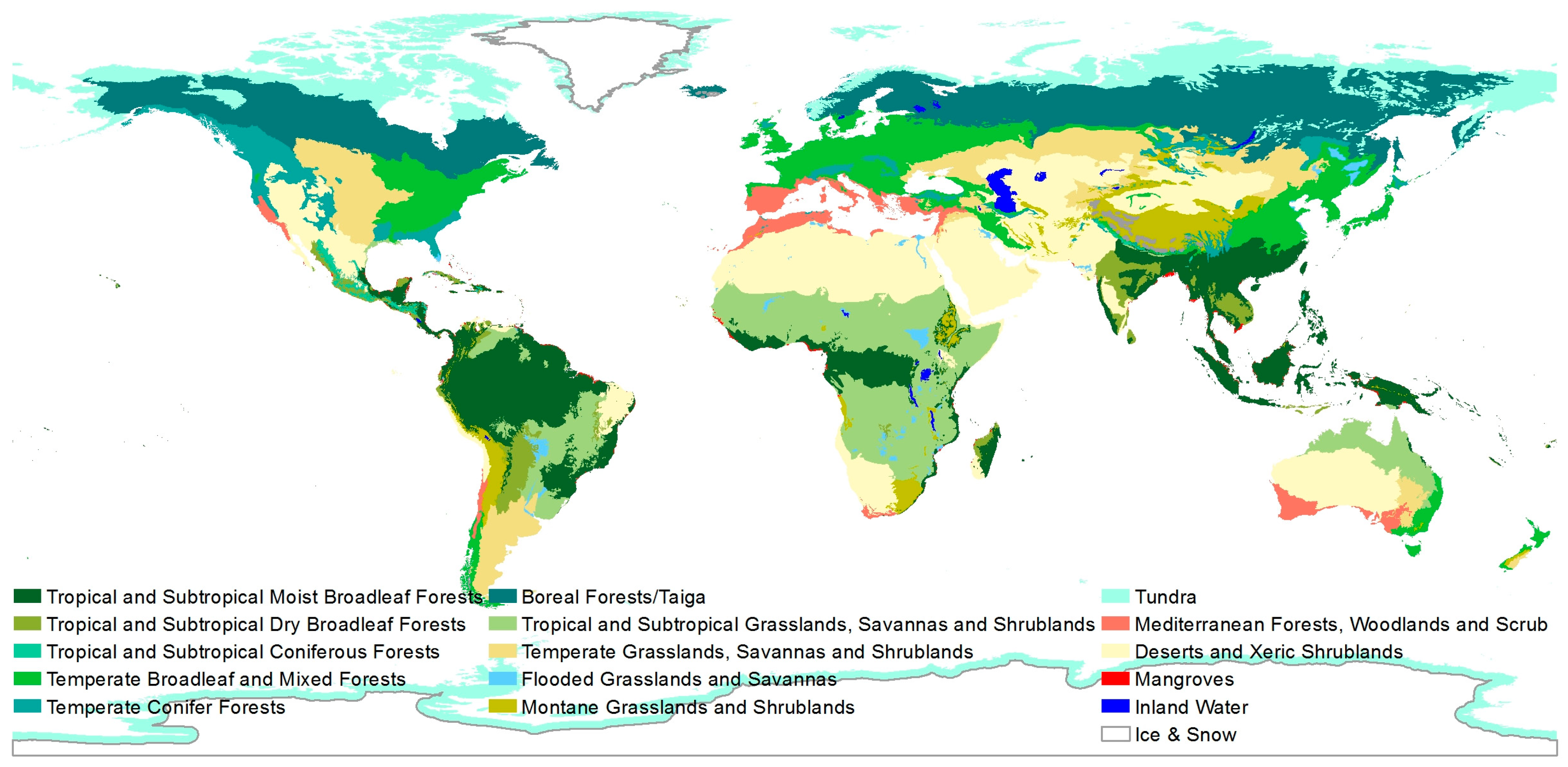
Figure 2.
Mean soil pH in every ecoregion.
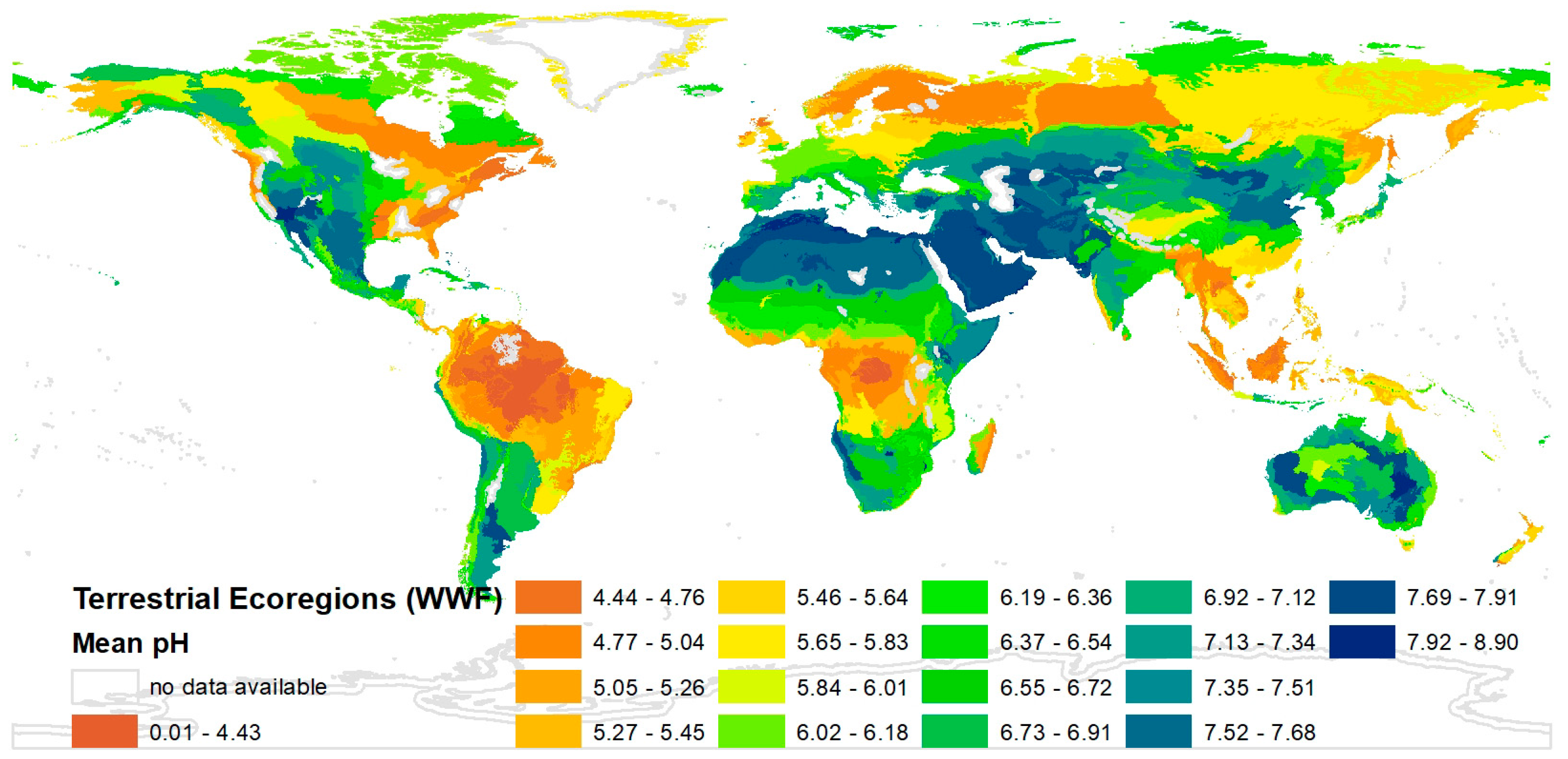
Figure 3.
Normalised biodiversity status of the ecoregions.

Figure 4.
Results of the Kruskal–Wallis test performed with SPSS 24 [23].
Figure 4.
Results of the Kruskal–Wallis test performed with SPSS 24 [23].
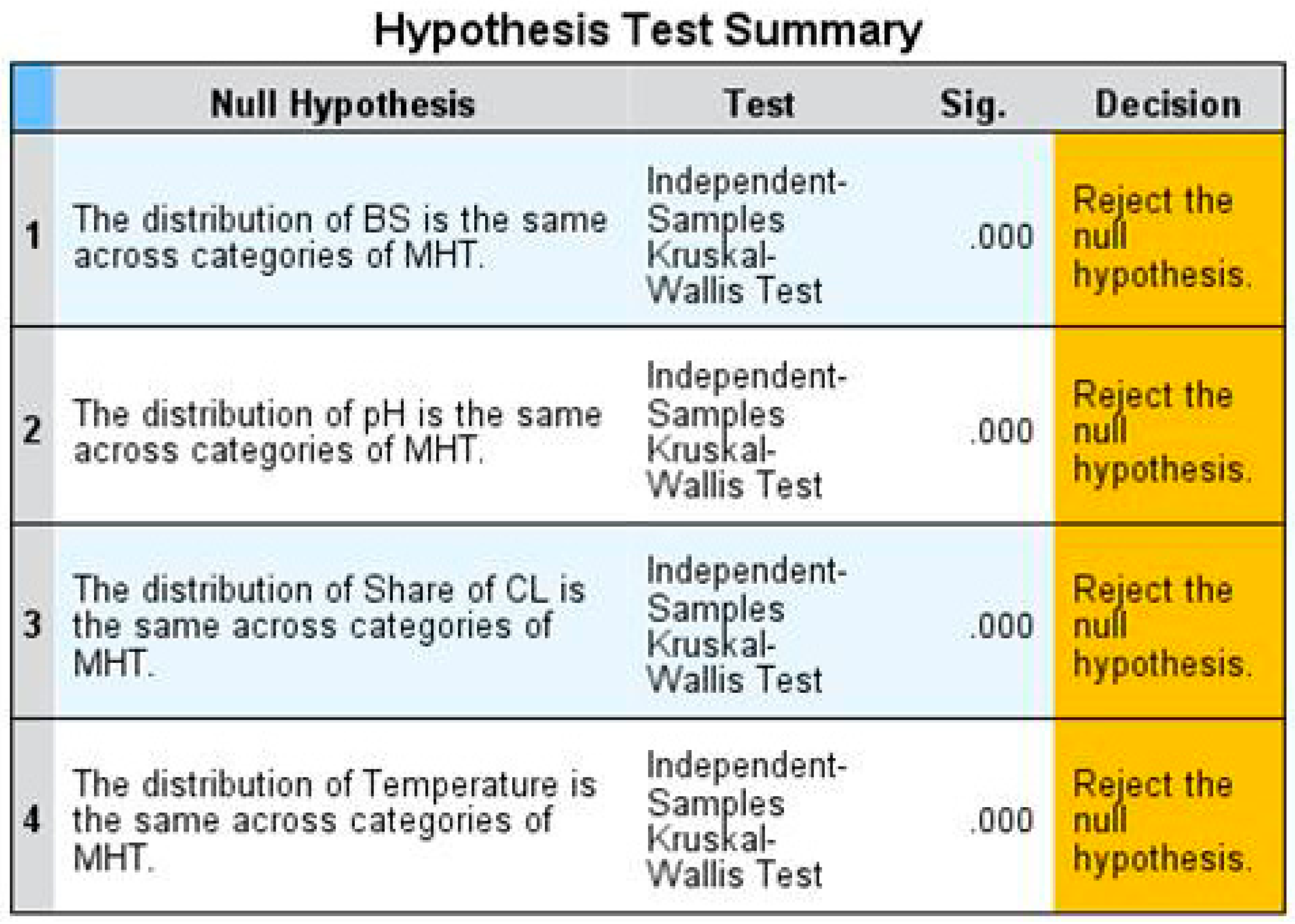
Figure 5.
Coefficients of determination for the best fitting curve estimation models regarding the relation of the biodiversity status (BS) and the “mean soil pH” (a); the BS and the “mean share of cultivated land” (b); and the BS and the “mean temperature in June” (c) in the respective major habitat type (MHT).
Figure 5.
Coefficients of determination for the best fitting curve estimation models regarding the relation of the biodiversity status (BS) and the “mean soil pH” (a); the BS and the “mean share of cultivated land” (b); and the BS and the “mean temperature in June” (c) in the respective major habitat type (MHT).
Figure 6.
Regression analysis and curve fitting results for the major habitat type “Tropical and Subtropical Moist Broadleaf Forests” for the biodiversity status and the parameters “mean soil pH” (pH), “mean share of cultivated land” (Share of CL) in (%) and “mean temperature in June” (Temperature) in (°C). The best curve fitting model is displayed. The y-axis presents the biodiversity status (BS), the x-axis presents the considered parameter.
Figure 6.
Regression analysis and curve fitting results for the major habitat type “Tropical and Subtropical Moist Broadleaf Forests” for the biodiversity status and the parameters “mean soil pH” (pH), “mean share of cultivated land” (Share of CL) in (%) and “mean temperature in June” (Temperature) in (°C). The best curve fitting model is displayed. The y-axis presents the biodiversity status (BS), the x-axis presents the considered parameter.
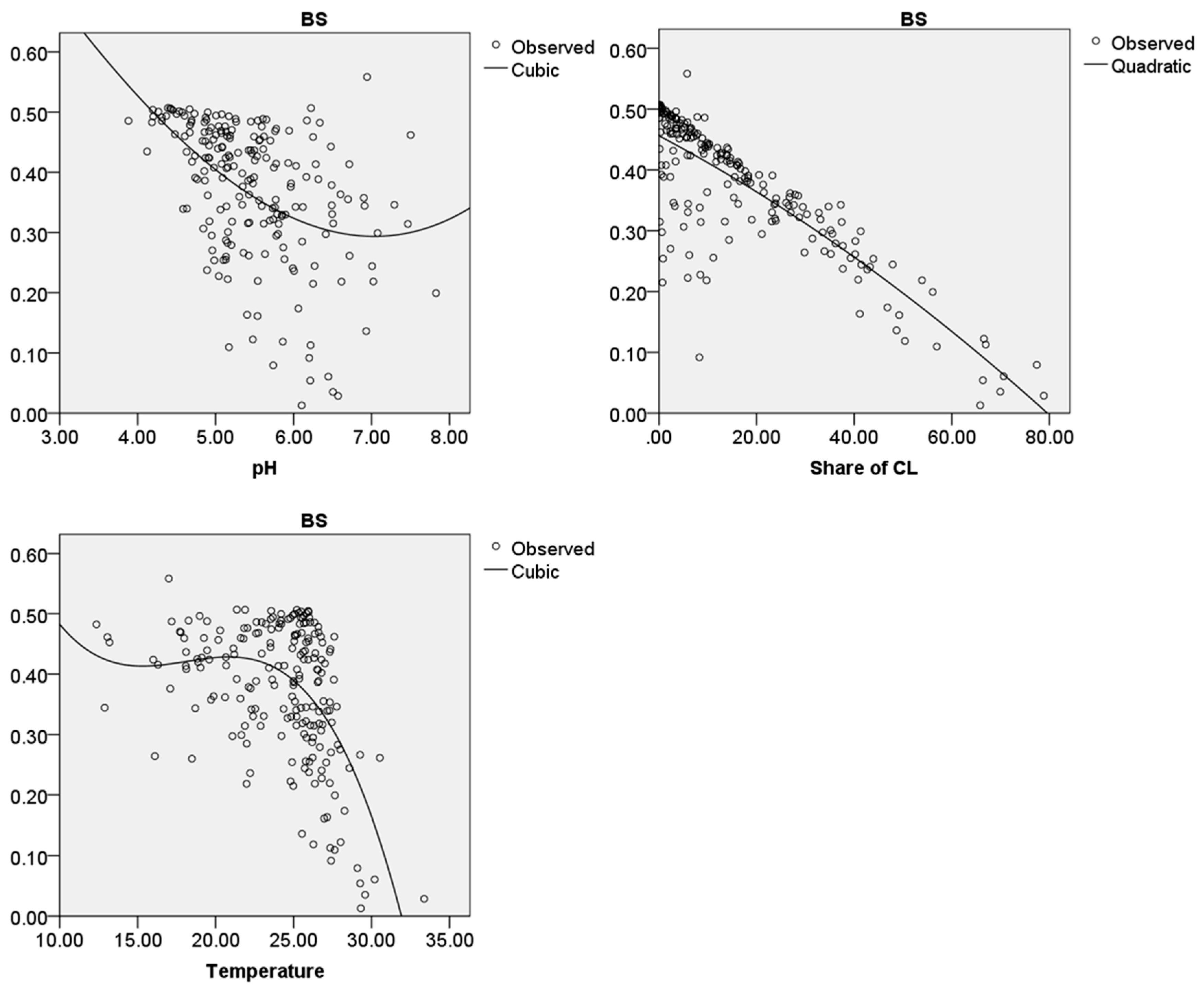
Figure 7.
Regression analysis and curve fitting results for the major habitat type “Tundra” for the biodiversity status and the parameters “mean soil pH” (pH), “mean share of cultivated land” (Share of CL) in (%) and “mean temperature in June” (Temperature) in (°C). The best curve fitting model is displayed. The y-axis presents the biodiversity status (BS), the x-axis presents the considered parameter.
Figure 7.
Regression analysis and curve fitting results for the major habitat type “Tundra” for the biodiversity status and the parameters “mean soil pH” (pH), “mean share of cultivated land” (Share of CL) in (%) and “mean temperature in June” (Temperature) in (°C). The best curve fitting model is displayed. The y-axis presents the biodiversity status (BS), the x-axis presents the considered parameter.

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Winter, L.; Berger, M.; Minkov, N.; Finkbeiner, M. Analysing the Impacts of Various Environmental Parameters on the Biodiversity Status of Major Habitats. Sustainability 2017, 9, 1775. https://doi.org/10.3390/su9101775
AMA Style
Winter L, Berger M, Minkov N, Finkbeiner M. Analysing the Impacts of Various Environmental Parameters on the Biodiversity Status of Major Habitats. Sustainability. 2017; 9(10):1775. https://doi.org/10.3390/su9101775
Chicago/Turabian StyleWinter, Lisa, Markus Berger, Nikolay Minkov, and Matthias Finkbeiner. 2017. "Analysing the Impacts of Various Environmental Parameters on the Biodiversity Status of Major Habitats" Sustainability 9, no. 10: 1775. https://doi.org/10.3390/su9101775
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.