
Changes in Patterns of Seasonality Shown by Migratory Fish under Global Warming: Evidence from Catch Data of Taiwan’s Coastal Fisheries

Abstract
:1. Introduction
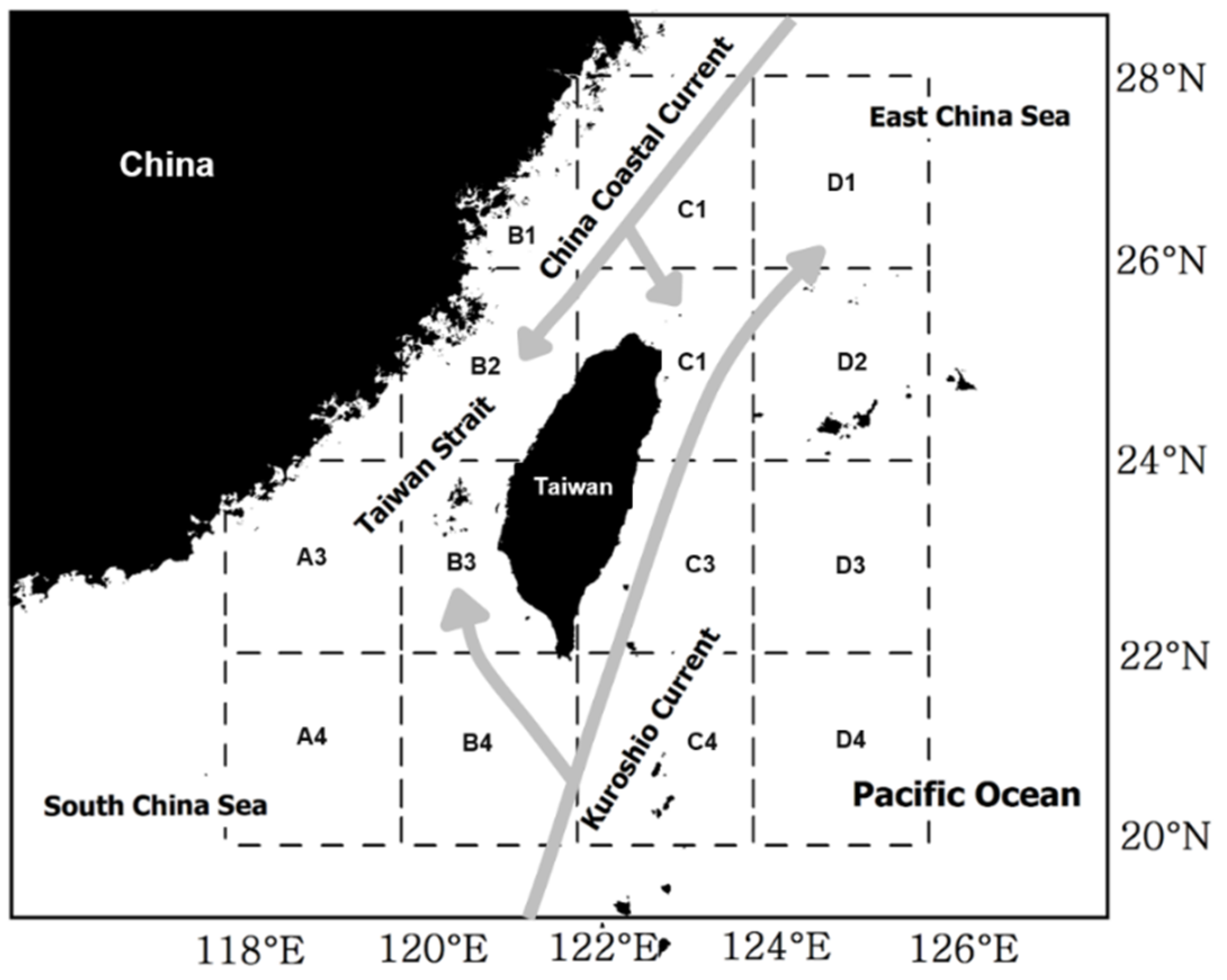
2. Data and Methods
2.1. Catch Data
2.2. Seasonality Analysis of the Catch
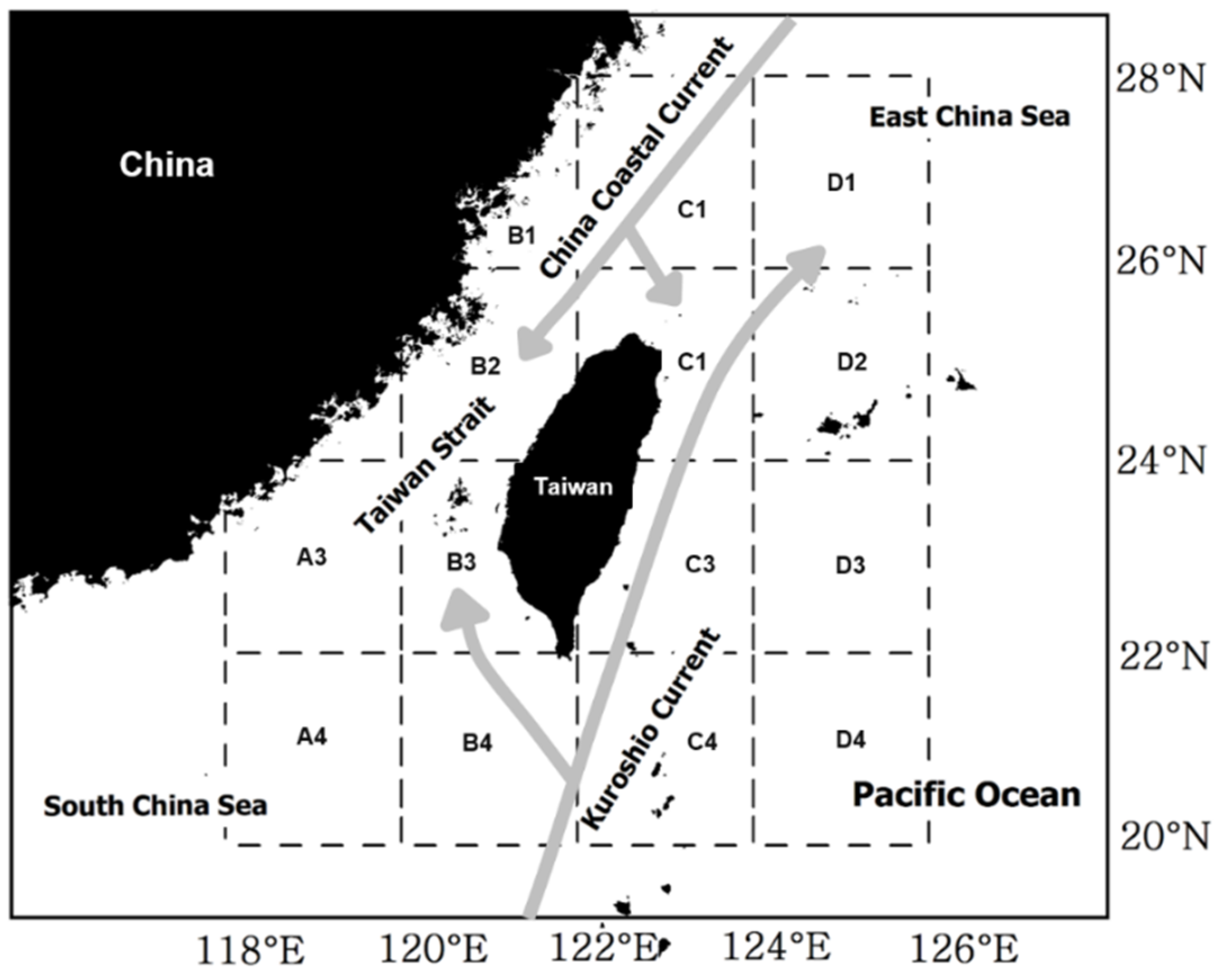
2.3. SST Data Process
2.4. Correlation between Seasonal Catch and Regional SSTA
3. Results
3.1. Data Segregation and Seasonality Determination
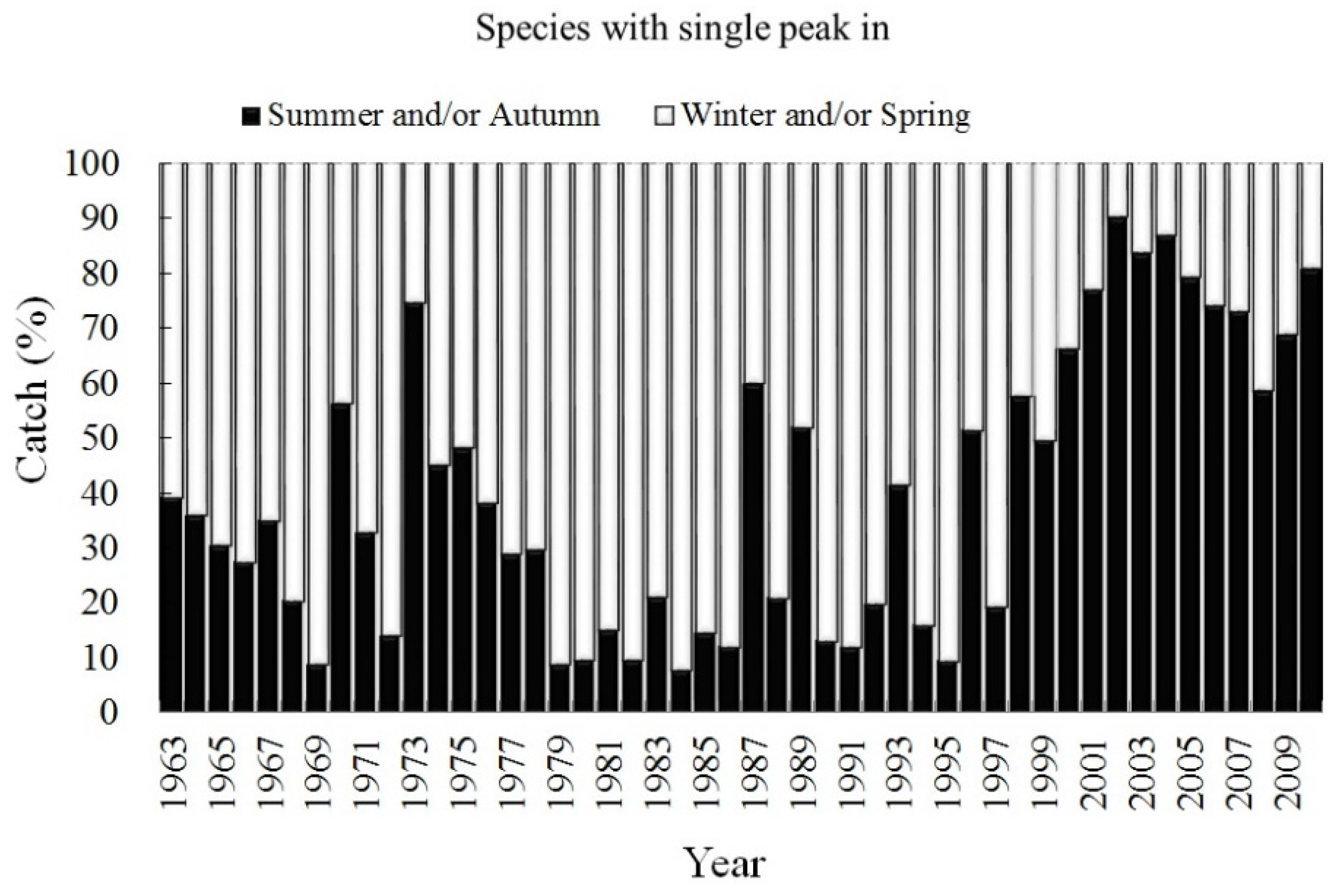
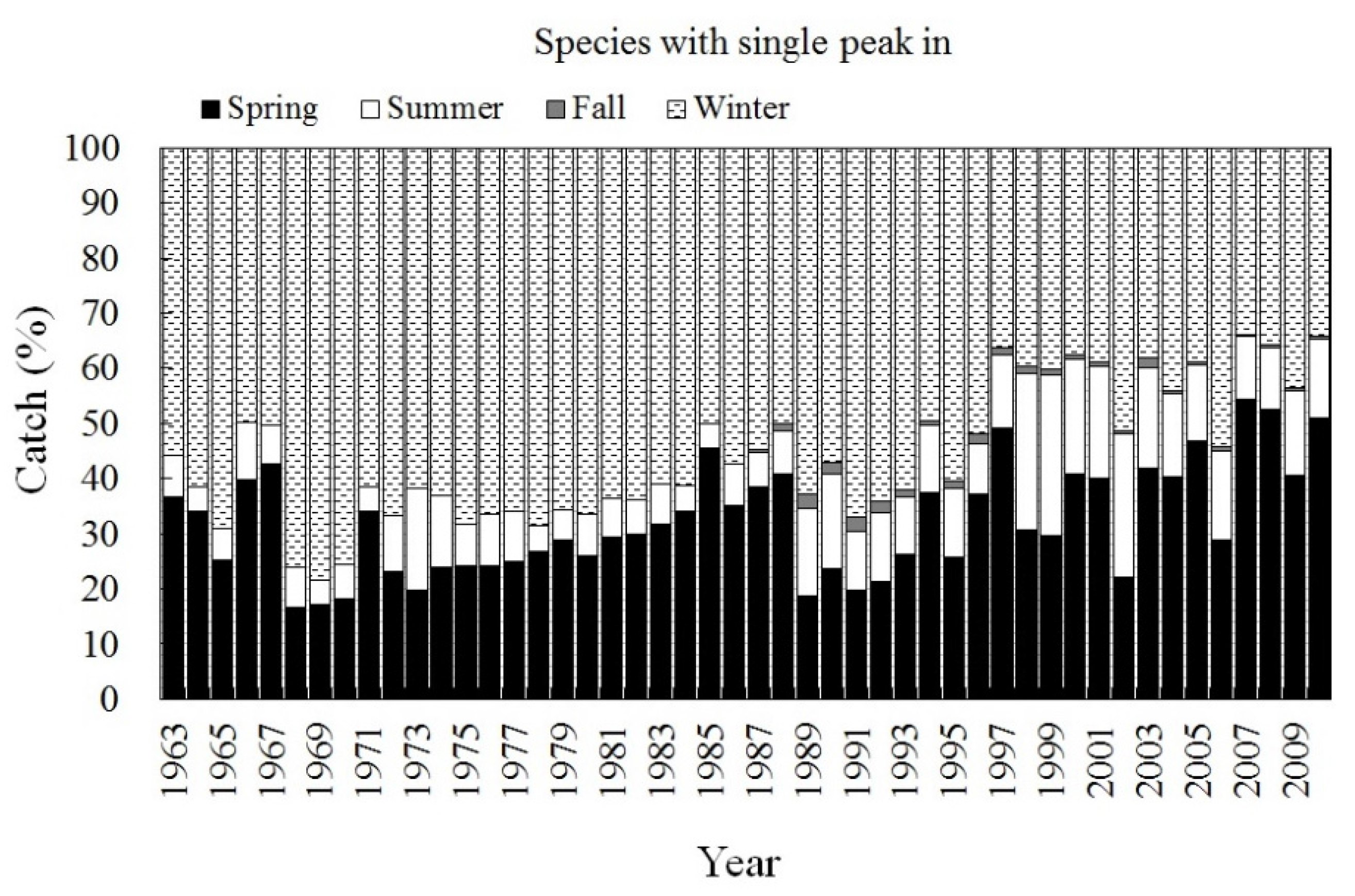
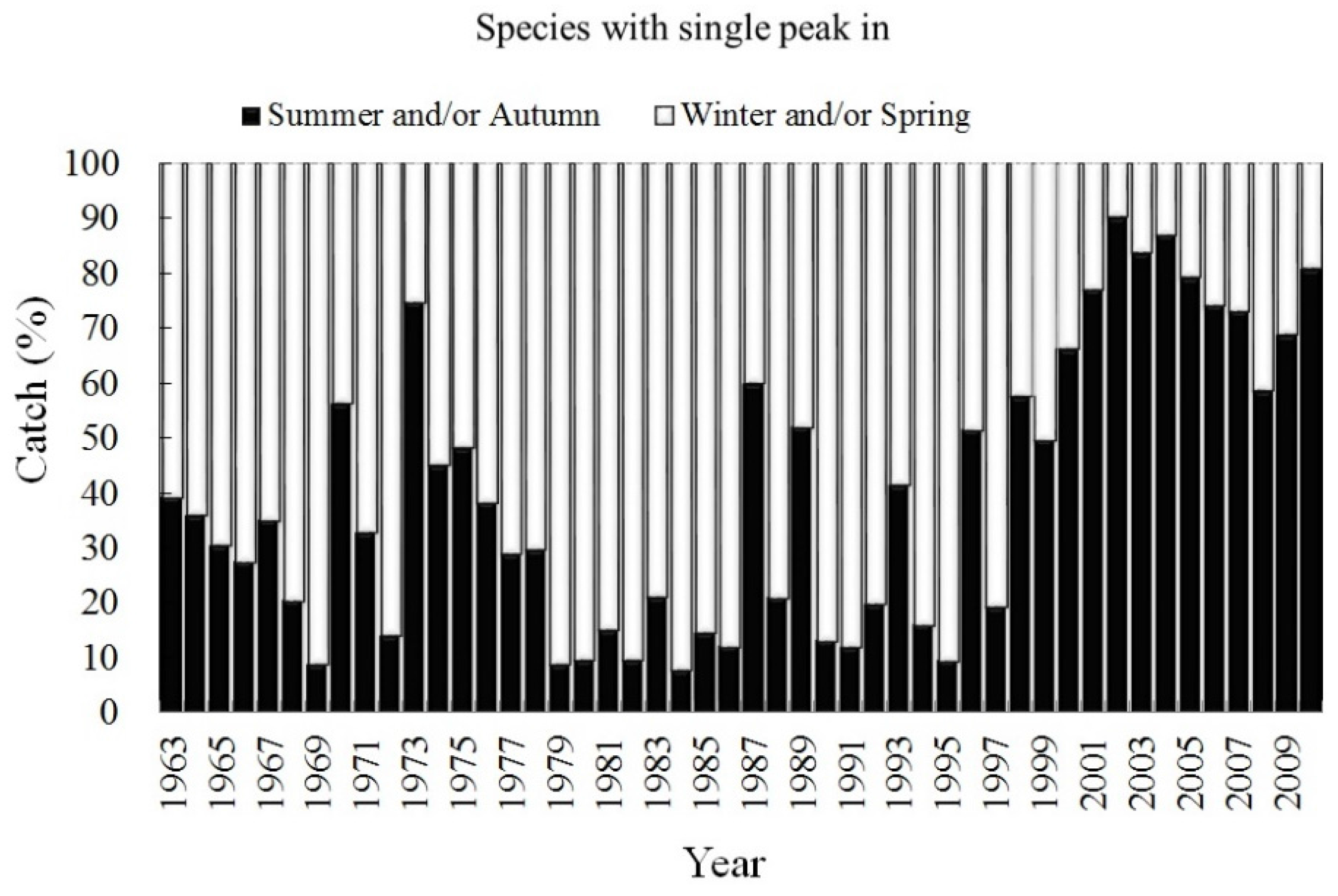
3.2. Trend of Catch Proportion for Single-Season Fish
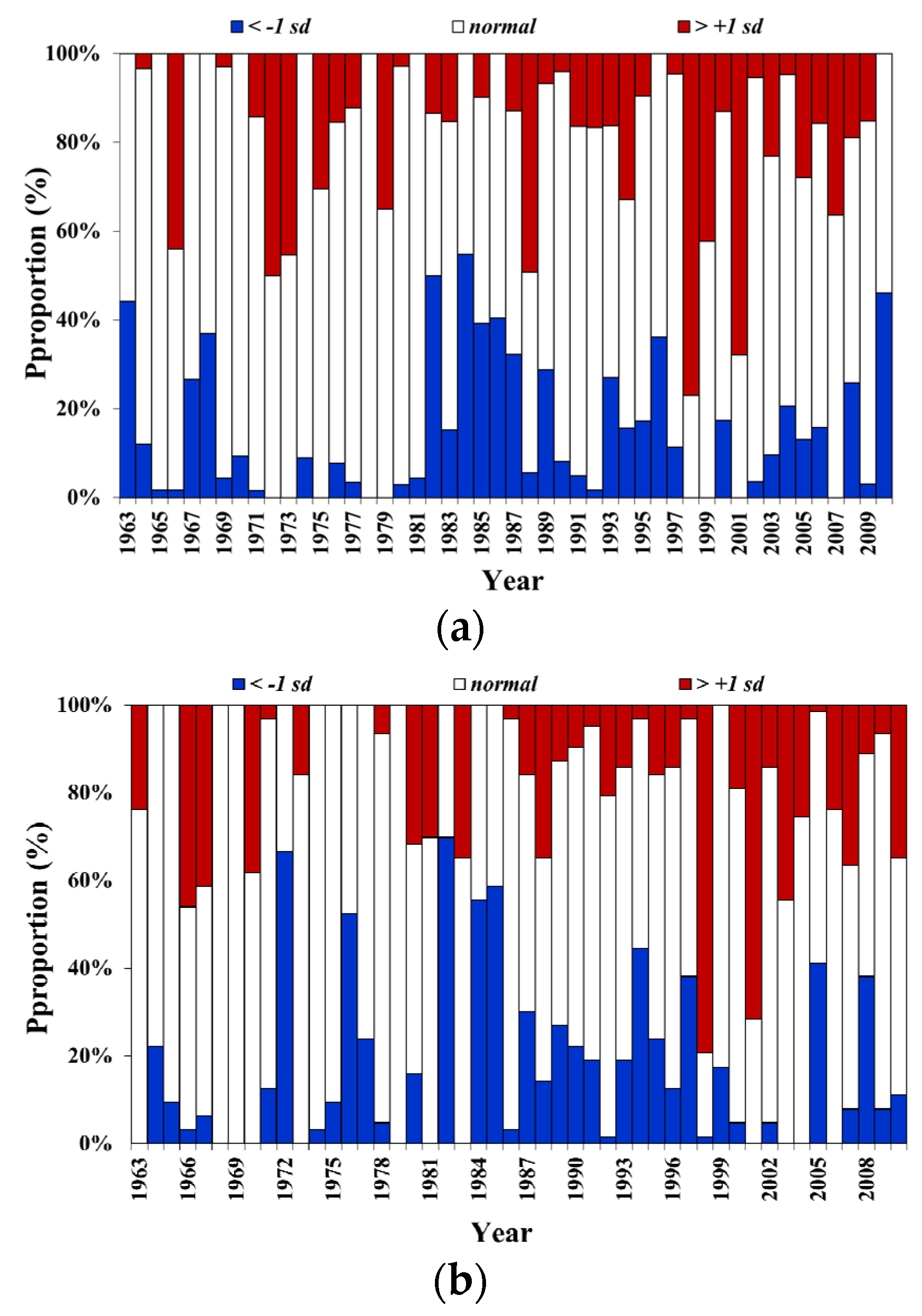
3.3. Trend of Catch Proportion for All Seasonally Migratory Species
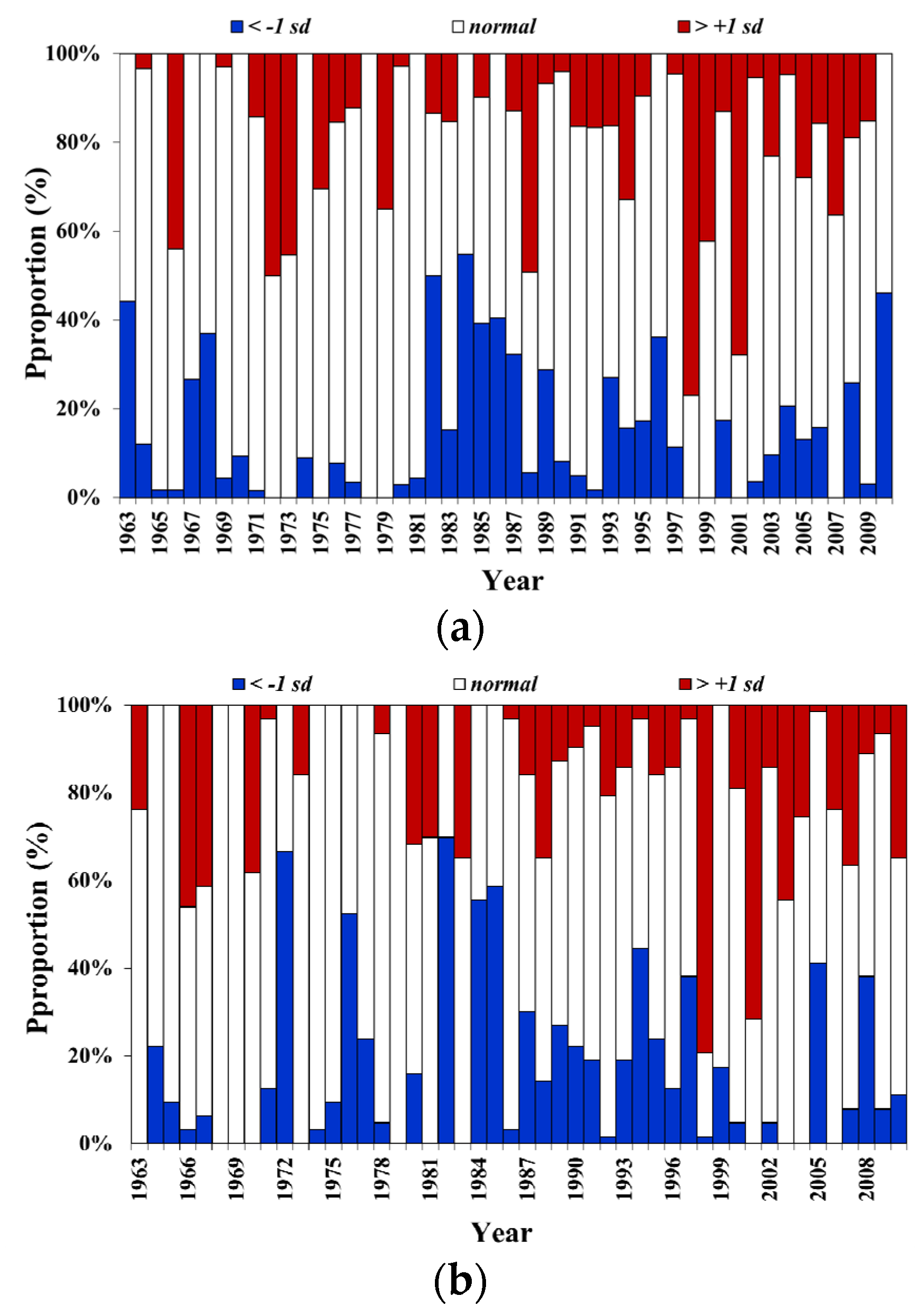
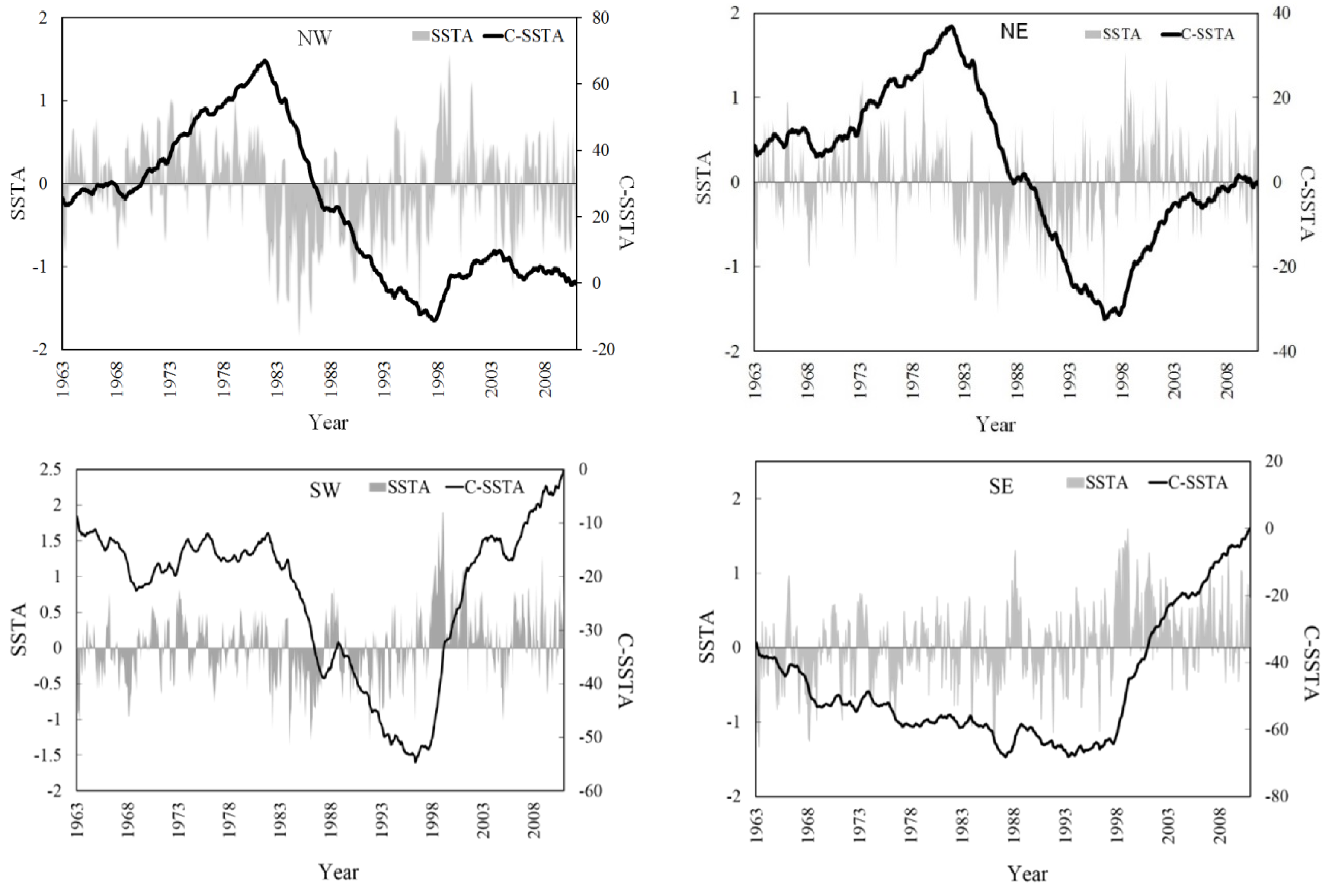
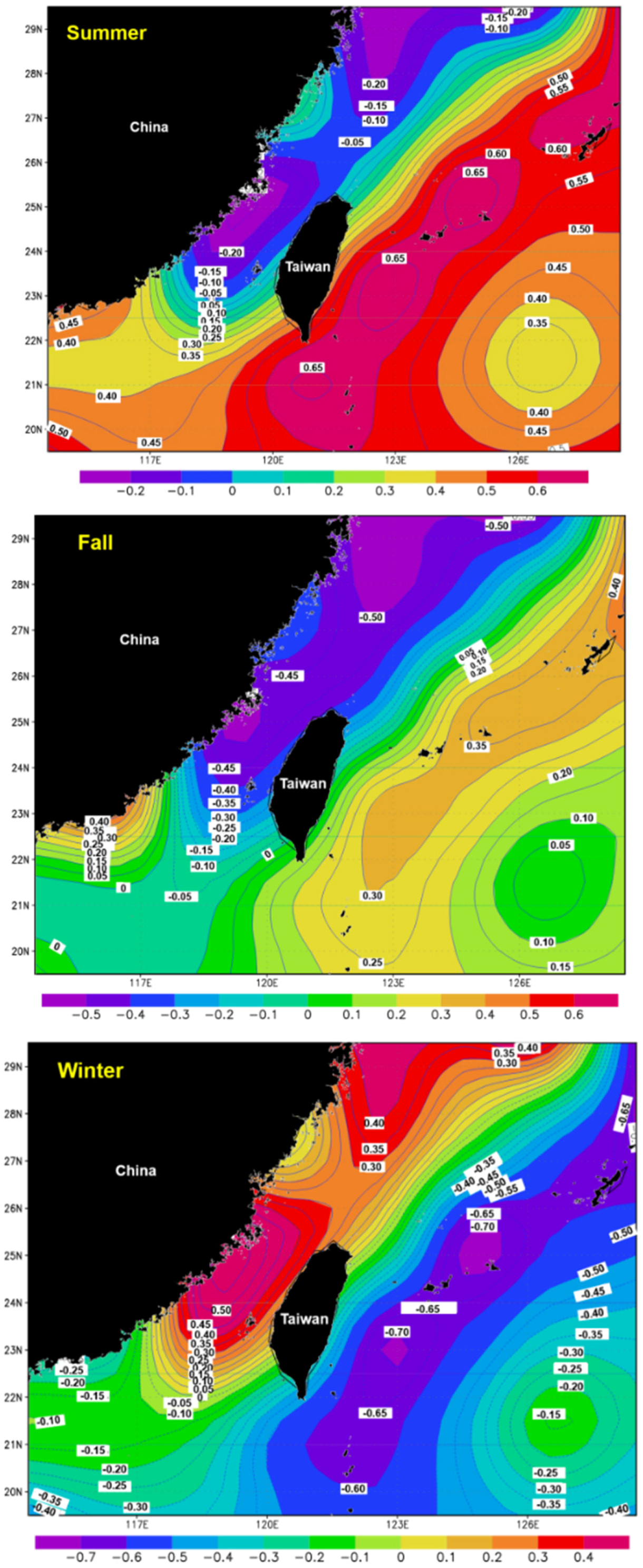
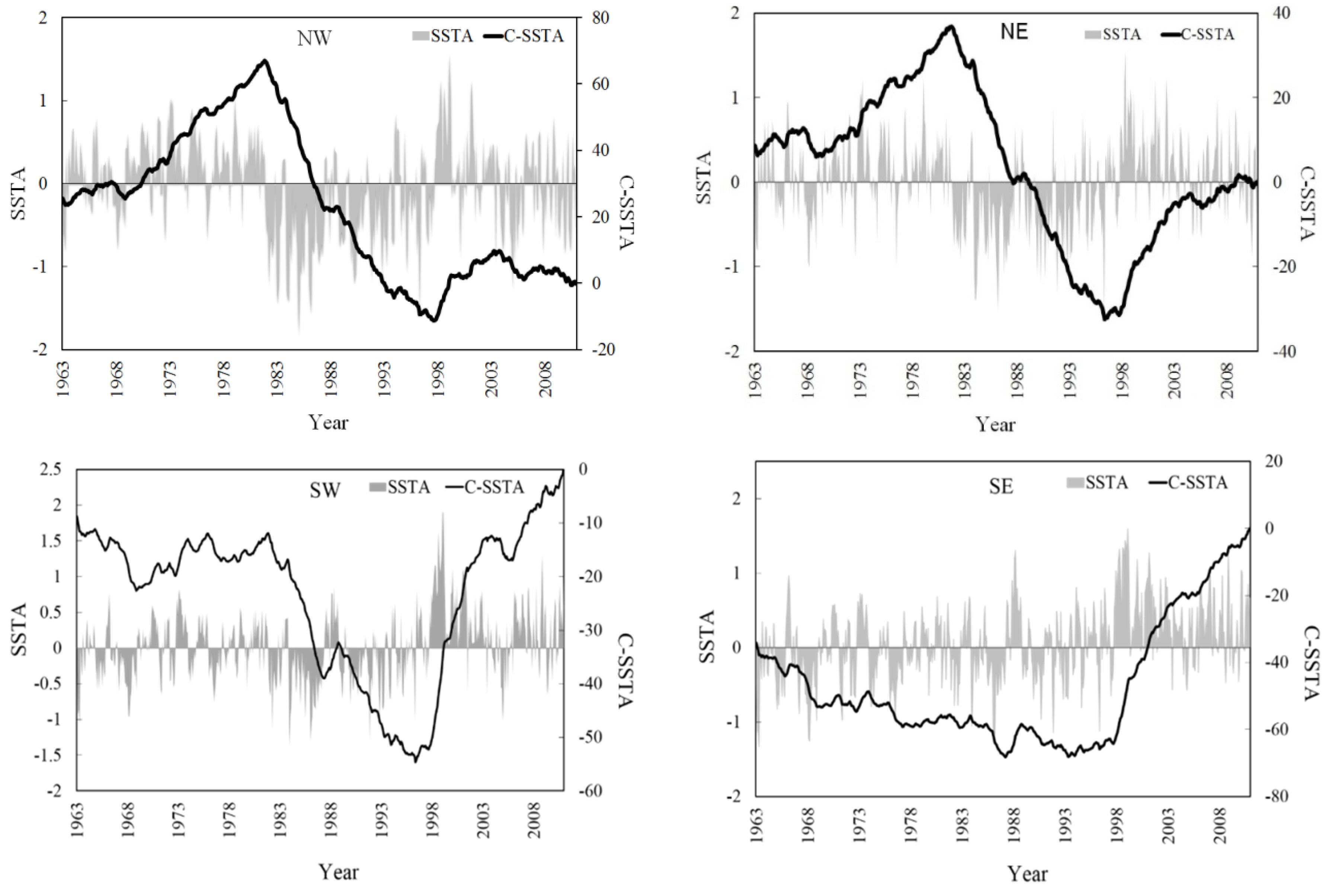
3.4. SST Change Trends in the Surrounding Waters of Taiwan
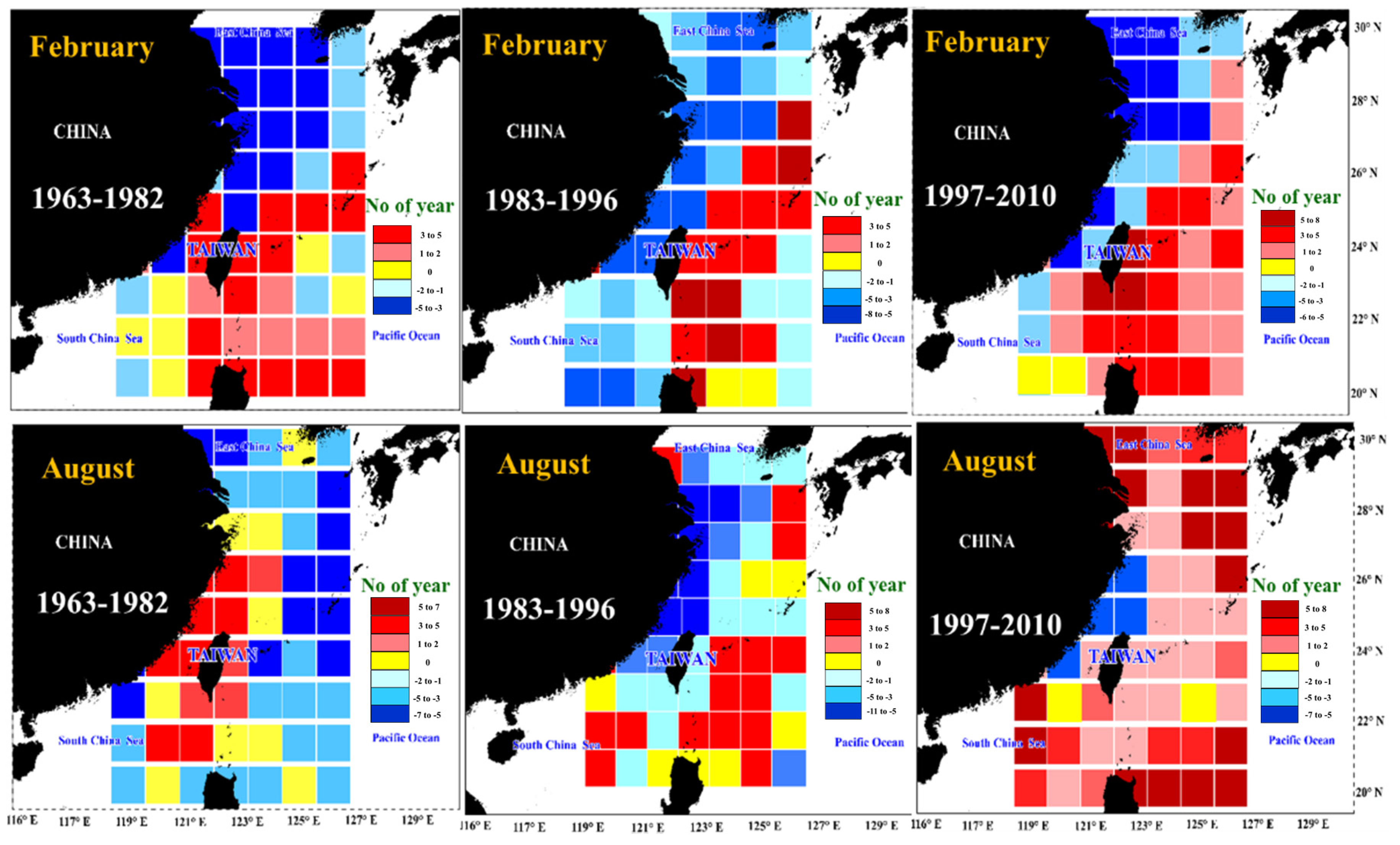
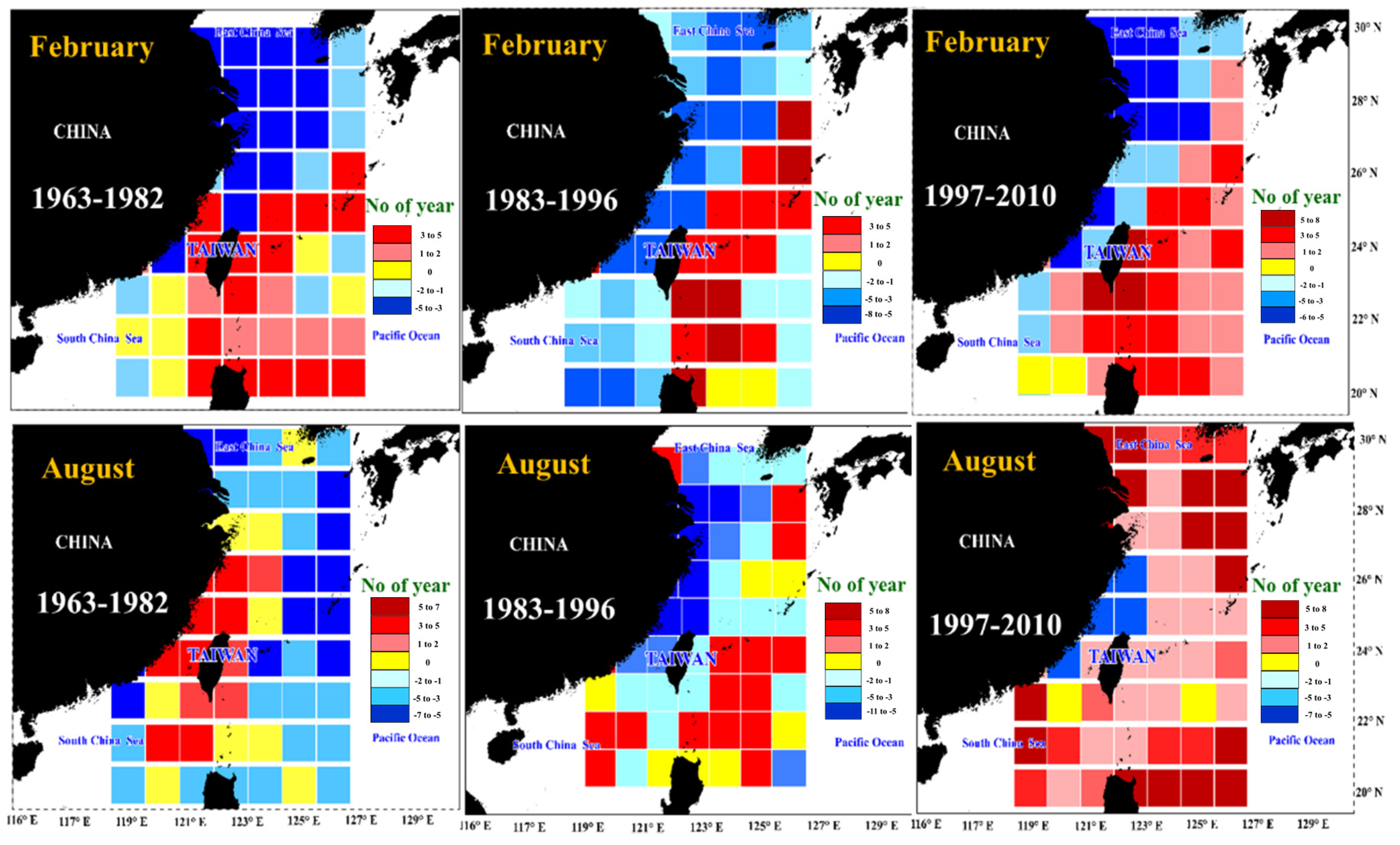
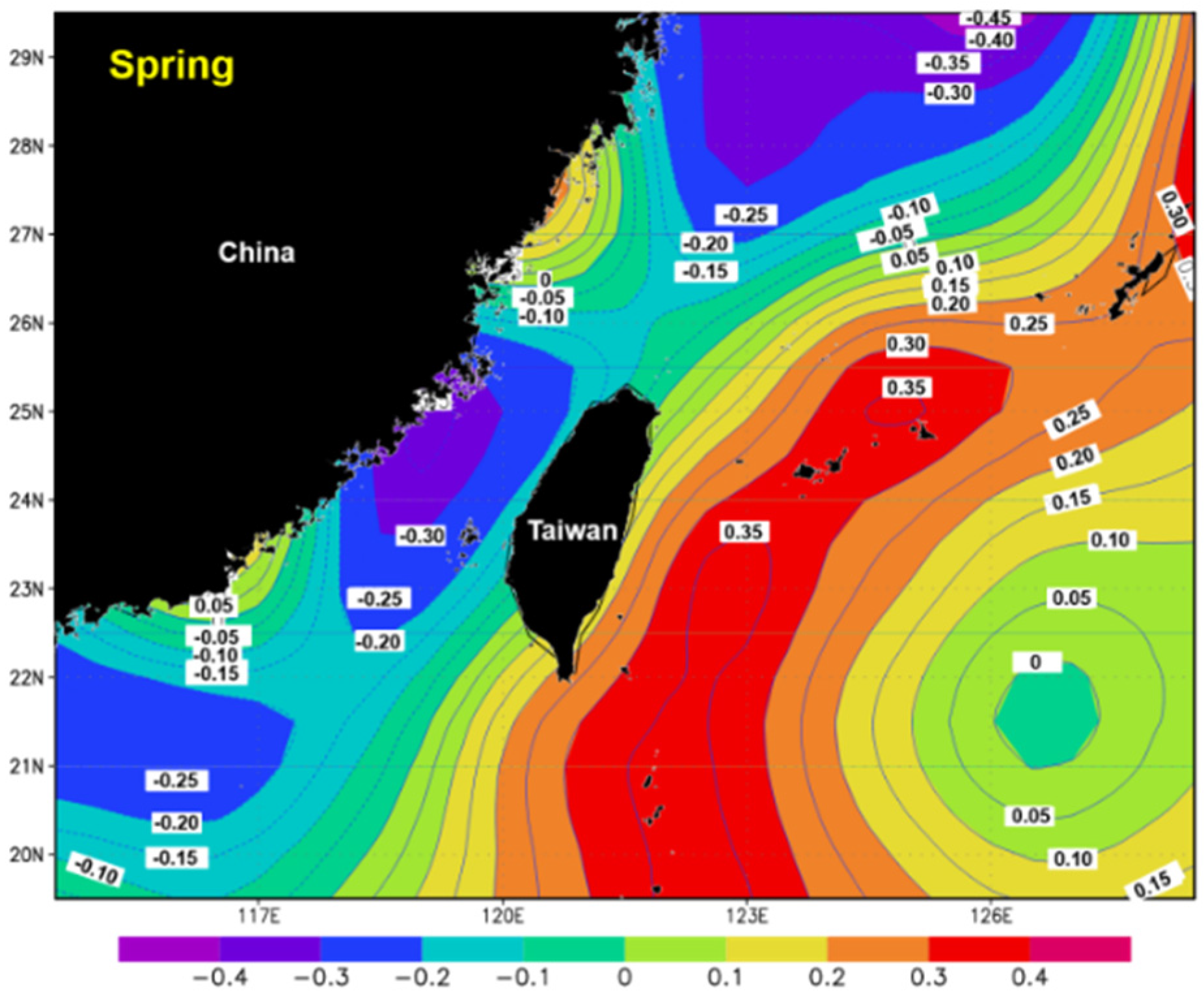
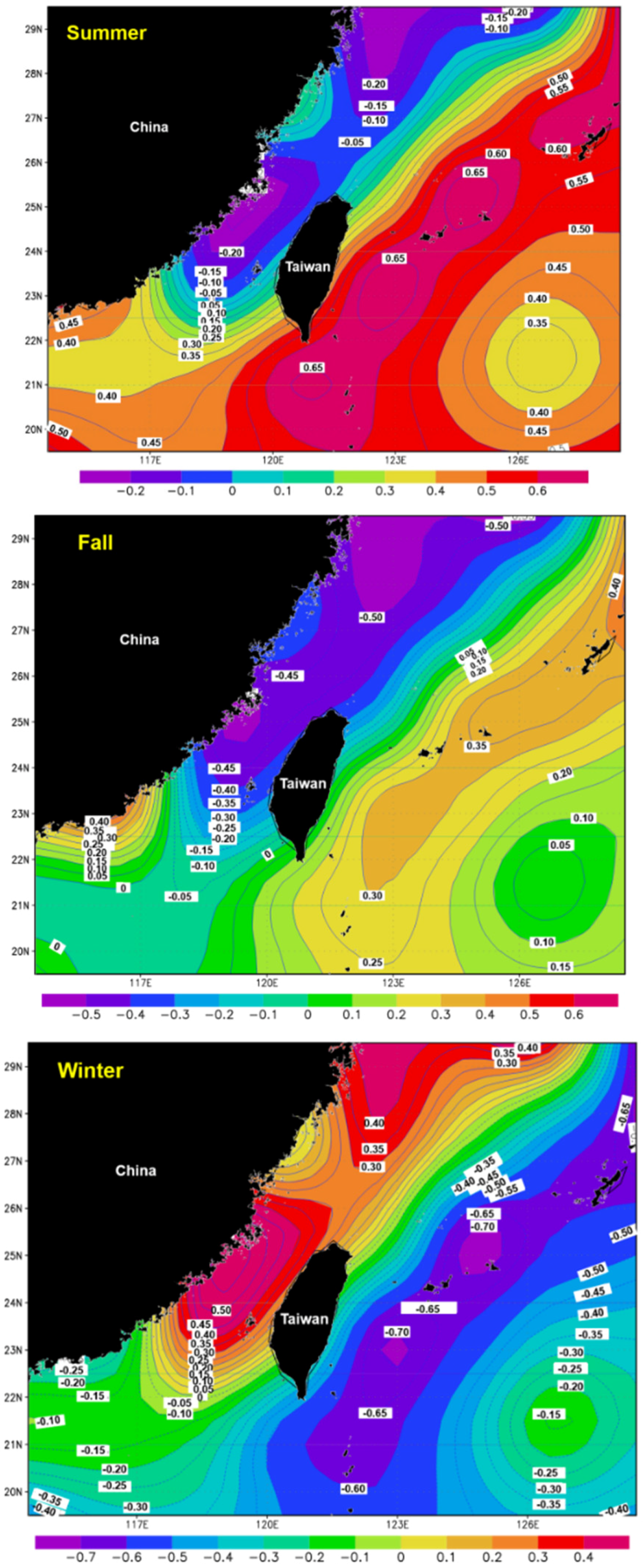
3.5. Spatial Distribution of the Correlation Coefficients between Catch and SST
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brander, K.; Neuheimer, A.; Andersen, K.H.; Hartvig, M. Overconfidence in model projections. ICES J. Mar. Sci. 2013, 70, 1065–1068. [Google Scholar] [CrossRef]
- Brander, K. Improving the Reliability of Fishery Predictions Under Climate Change. Curr. Clim. Chang. Rep. 2015. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Lam, V.W.Y.; Sarmiento, J.L.; Kearney, K.; Watson, R.; Pauly, D. Projecting global marine biodiversity impacts under climate change scenarios. Fish Fish. 2009, 10, 235–251. [Google Scholar] [CrossRef]
- Cheung, W.W.L.; Lam, V.W.; Sarmiento, K.K.; Watson, R.; Zeller, D.; Pauly, D. Large-scale redistribution of maximum fisheries catch potential in the global ocean under climate change. Glob. Chang. Biol. 2010, 16, 24–35. [Google Scholar] [CrossRef]
- Hobday, A.J.; Pecl, G.T. Identification of global marine hotspots: Sentinels for change and vanguards for adaptation action. Rev. Fish Biol. Fish. 2014, 24, 415–425. [Google Scholar] [CrossRef]
- Lu, J.H.; Lee, S.L. Observations of changes in the fish species composition in the coastal zone at the Kuroshio Current and China Coastal Current front during climate change using set-net fishery (1993–2011). Fish. Res. 2014, 155, 103–113. [Google Scholar] [CrossRef]
- Tang, T.Y.; Yang, Y.J. Low frequency current variability on the shelf breaknortheast of Taiwan. J. Oceanogr. 1993, 49, 193–210. [Google Scholar] [CrossRef]
- Chuang, W.S.; Liang, W.D. Seasonal variability of intrusion of the KuroshioWater across the continental shelf northeast of Taiwan. J. Oceanogr. 1994, 50, 531–542. [Google Scholar] [CrossRef]
- Liu, Z.; Gan, J. Variability of the Kuroshio in the East China Sea derived fromsatellite altimetry data. Deep Sea Res. I Oceanogr. Res. Pap. 2012, 59, 25–36. [Google Scholar] [CrossRef]
- Igor, M.B.; Lee, M.A. Long-term variability of sea surface temperature in Taiwan Strait. Clim. Chang. 2014, 124, 821–834. [Google Scholar]
- Tian, Y.; Ueno, Y.; Suda, M.; Akamine, T. Decadal variability in the abundance of Pacific saury and its response to climatic/oceanic regime shifts in the northwestern subtropical Pacific during the last half century. J. Mar. Syst. 2004, 52, 235–257. [Google Scholar] [CrossRef]
- Tian, Y.J.; Kidokoro, H.; Watanabe, T. Long-term changes in the fish community structure from the Tsushima warm current region of the Japan/East Sea with an emphasis on the impacts of fishing and climate regime shift over the last four decades. Prog. Oceanogr. 2006, 68, 217–237. [Google Scholar] [CrossRef]
- Tian, Y.; Kidokoro, H.; Fujino, T. Interannual-decadal variability of demersal fish community in the Japan Sea: Impacts of climate regime shifts and trawl fishing with implications for ecosystem-based management. Fish. Res. 2011, 112, 140–153. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Babcock, R.C.; Butler, A.; Hobday, A.J.; HoeghGuldberg, O.; Kunz, T.J.; Matear, R.; Milton, D.; Okey, T.A.; Richardson, A.J. Climate change and Australian marine life. Oceanogr. Mar. Biol. Annu. Rev. 2007, 45, 409–480. [Google Scholar]
- Brierley, A.S.; Kingsford, M.J. Impacts of climate change on marine organisms and ecosystems. Curr. Biol. 2009, 19, R602–R614. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.J.; Fulton, E.A.; Hobday, A.J.; Matear, R.J.; Possingham, H.; Bulman, C.; Christensen, V.; Forrest, R.; Gehrke, P.; Gribble, N.; et al. Effects of climate-driven primary production change on marine food webs: Implications for fisheries and conservation. Glob. Chang. Biol. 2009, 16, 1194–1212. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Ke, H.M.; Lin, W.W.; Kao, H.W. Genetic diversity and differentiation of grey mullet (Mugil cephalus) in the coastal waters of Taiwan. Zool. Sci. 2009, 26, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Lee, M.A.; Lee, K.T.; Shao, K.S. Adaptation of fisheries and mariculture management to extreme oceanic environmental changes and climate variability in Taiwan. Mar. Pol. 2013, 38, 476–482. [Google Scholar] [CrossRef]
- Lan, K.W.; Lee, M.A.; Zhang, C.I.; Wang, P.Y.; Wu, L.J.; Lee, K.T. Effects of climate variability and climate change on the fishing conditions for grey mullet (Mugil cephalus L.) in the Taiwan Strait. Clim. Change 2014. [Google Scholar] [CrossRef]
- Shao, K.T.; Chen, J.Y. Coastal Fishes of Taiwan. 2004. Available online: http://fishdb.sinica.edu.tw/chi/home.php (accessed on 15 March 2016).
- Froese, R.; Pauly, D. FishBase. 2012. Available online: http://www.fishbase.org (accessed on 15 March 2016).
- Sea Around Us. 2012. Available online: http://www.seaaroundus.org (accessed on 15 March 2016).
- Oey, L.Y.; Chang, M.C.; Chang, Y.L.; Lin, Y.C.; Xu, F.H. Decadal warming of coastal China Seas and coupling with winter monsoon and currents. Geophys. Res. Lett. 2013, 40, 6288–6292. [Google Scholar] [CrossRef]
- Zhang, C.I.; Lee, J.B.; Kim, S.; Oh, J.H. Climatic regime shifts and their impacts on marine ecosystems and fisheries resources in Korean waters. Progr. Oceanogr. 2000, 47, 171–190. [Google Scholar] [CrossRef]
- Zhang, C.I.; Lee, J.B.; Seo, Y.I.; Yoon, S.C.; Kim, S. Variations in the abundance of fisheries resources and ecosystem structure in the Japan/East Sea. Prog. Oceanogr. 2004, 61, 245–265. [Google Scholar] [CrossRef]
- Jung, S.; Pang, I.C.; Lee, J.H.; Choi, I.; Cha, H.K. Latitudinal shifts in the distribution of exploited fishes in Korean waters during the last 30 years: A consequence of climate change. Rev. Fish Biol. Fish. 2014, 24, 443–462. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Species Name | Associated Current | p1 | p2 | Seasonality |
|---|---|---|---|---|---|
| Black pomfret | Parastromateus niger | CCC | 0.01 | 0.01 | Winter |
| Black sea bream | Acanthopagrus schlegelii | - | 0.01 | 0.02 | Winter |
| East Asian four-finger threadfin | Eleutheronema rhadinum | KC | 0.01 | 0.01 | Winter |
| Giant black marlin | Makaira indica | KC | 0.01 | 0.01 | Winter |
| Japanese butterfish | Psenopsis anomala | CCC | 0.01 | 0.01 | Winter |
| Largehead hairtail | Trichiurus lepturus | CCC | 0.01 | 0.01 | Winter |
| Mullet | Mugil cephalus | CCC | 0.01 | 0.01 | Winter |
| Red stingray | Dasyatis akajei | CCC | 0.01 | 0.01 | Winter |
| Cattlefish | Sepia pharaonis | - | 0.01 | 0.02 | Spring |
| Jack mackerel | Trachurus japonicus | CCC | 0.01 | 0.01 | Spring |
| Sea barbel | Arius maculatus | KC | 0.01 | 0.04 | Spring |
| Slender snoek | Thyrsitoides marleyi | KC | 0.01 | 0.04 | Spring |
| Southern mackerel | Scomber australasicus | CCC | 0.01 | 0.02 | Spring |
| Black croaker | Atrobucca nibe | KC | 0.01 | 0.01 | Summer |
| Octopus | Octopus vulgare | - | 0.01 | 0.04 | Summer |
| Oval grouper | Triso dermopterus | CCC | 0.01 | 0.01 | Summer |
| Pacific sailfish | Istiophorus platypterus | KC | 0.01 | 0.01 | Summer |
| Pike conger | Muraenesox cinereus | KC | 0.01 | 0.02 | Summer |
| Red sea bream | Dentex hypselosomus | KC | 0.01 | 0.01 | Summer |
| Silver sillago | Sillago sihama | - | 0.01 | 0.01 | Fall |
| Common Name | Species Name | Associated Current | p1 | p2 | Seasonality |
|---|---|---|---|---|---|
| Bullet tuna | Auxis rochei rochei | KC | 0.03 | 0.14 | Spring & Winter |
| Pacific blue marlin | Makaira nigricans | KC | 0.01 | 0.07 | Spring & Winter |
| Silver pomfret | Pampus argenteus | KC | 0.01 | 0.06 | Spring & Winter |
| Slender lizard fish | Saurida elongata | KC | 0.01 | 0.49 | Spring & Winter |
| Spaniard | Scomberomorus niphonius | CCC | 0.01 | 0.37 | Spring & Winter |
| Yellowfin tuna | Thunnus albacares | KC | 0.01 | 0.4 | Spring & Winter |
| Dolphinfish | Coryphaena hippurus | KC | 0.01 | 0.40 | Summer & Winter |
| Silver sea meagre | Pennahia argentata | - | 0.01 | 0.37 | Summer & Winter |
| Mi-iuy croaker | Miichthys miiuy | KC | 0.02 | 0.30 | Spring & Summer |
| Sword tip squid | Loligo edulis | - | 0.01 | 0.20 | Summer & Fall |
| Tilefish | Branchiostegus albus | - | 0.04 | 0.08 | Summer & Fall |
| Common Name | Species Name | Associated Current | p1 | p2 | Seasonality |
|---|---|---|---|---|---|
| Bullet mackerel | Auxis thazard thazard | KC | 0.34 | 0.14 | NS |
| Croceine croaker | Larimichthys crocea | - | 0.23 | 0.34 | NS |
| Golden threadfin bream | Nemipterus virgatus | - | 0.51 | 0.43 | NS |
| Large yellow croaker | Larimichthys crocea | - | 0.95 | 0.4 | NS |
| Moonfish | Mene maculata | - | 0.07 | 0.22 | NS |
| Pacific striped marlin | Kajikia audax | KC | 0.17 | 0.21 | NS |
| Rainbow sardine | Dussumieria elapsoides | - | 0.23 | 0.4 | NS |
| Red bigeye fish | Priacanthus macracanthus | - | 0.96 | 0.34 | NS |
| Red bokako | Argyrops spinifer | - | 0.43 | 0.34 | NS |
| Red seabream | Pagrus major | - | 0.25 | 0.06 | NS |
| Redgill emperor | Lethrinus rubrioperculatus | - | 0.1 | 0.4 | NS |
| Skipjack tuna | Katsuwonus pelamis | CCC | 0.55 | 0.44 | NS |
| Small-mouthed nannygai | Lutjanus erythropterus | - | 0.69 | 0.31 | NS |
| Spotted sardine | Konosirus punctatus | - | 0.43 | 0.24 | NS |
| Swordfish | Xiphias gladius | KC | 0.16 | 0.38 | NS |
| White lady carp | Cirrhinus Molitorella | - | 0.11 | 0.13 | NS |
| Yellow band goatfish | Parupeneus chrysopleuron | - | 0.24 | 0.31 | NS |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, C.-H.; Lu, H.-J.; He, J.-S.; Lan, K.-W.; Chen, J.-L. Changes in Patterns of Seasonality Shown by Migratory Fish under Global Warming: Evidence from Catch Data of Taiwan’s Coastal Fisheries. Sustainability 2016, 8, 273. https://doi.org/10.3390/su8030273
Ho C-H, Lu H-J, He J-S, Lan K-W, Chen J-L. Changes in Patterns of Seasonality Shown by Migratory Fish under Global Warming: Evidence from Catch Data of Taiwan’s Coastal Fisheries. Sustainability. 2016; 8(3):273. https://doi.org/10.3390/su8030273
Chicago/Turabian StyleHo, Ching-Hsien, Hsueh-Jung Lu, Jia-Sin He, Kuo-Wei Lan, and Jyun-Long Chen. 2016. "Changes in Patterns of Seasonality Shown by Migratory Fish under Global Warming: Evidence from Catch Data of Taiwan’s Coastal Fisheries" Sustainability 8, no. 3: 273. https://doi.org/10.3390/su8030273