
Isolation and Genome Characterization of the Virulent Staphylococcus aureus Bacteriophage SA97




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Bacteriophage Isolation and Propagation
2.3. Transmission Electron Microscopy (TEM)
2.4. Bacteriophage Host Range
2.5. Bacterial Challenge Assay
2.6. Lysogen Formation
2.7. One-Step Growth Curve Assay
2.8. Receptor Analysis
2.9. Adsorption Assay
2.10. Determination of the Frequency of the Bacteriophage-Insensitive Mutants (BIMs)
2.11. Transduction Assay
2.12. Bacteriophage Genomic DNA Purification
2.13. Full-Genome Sequencing of Bacteriophage SA97 and Bioinformatics Analysis
3. Results and Discussion
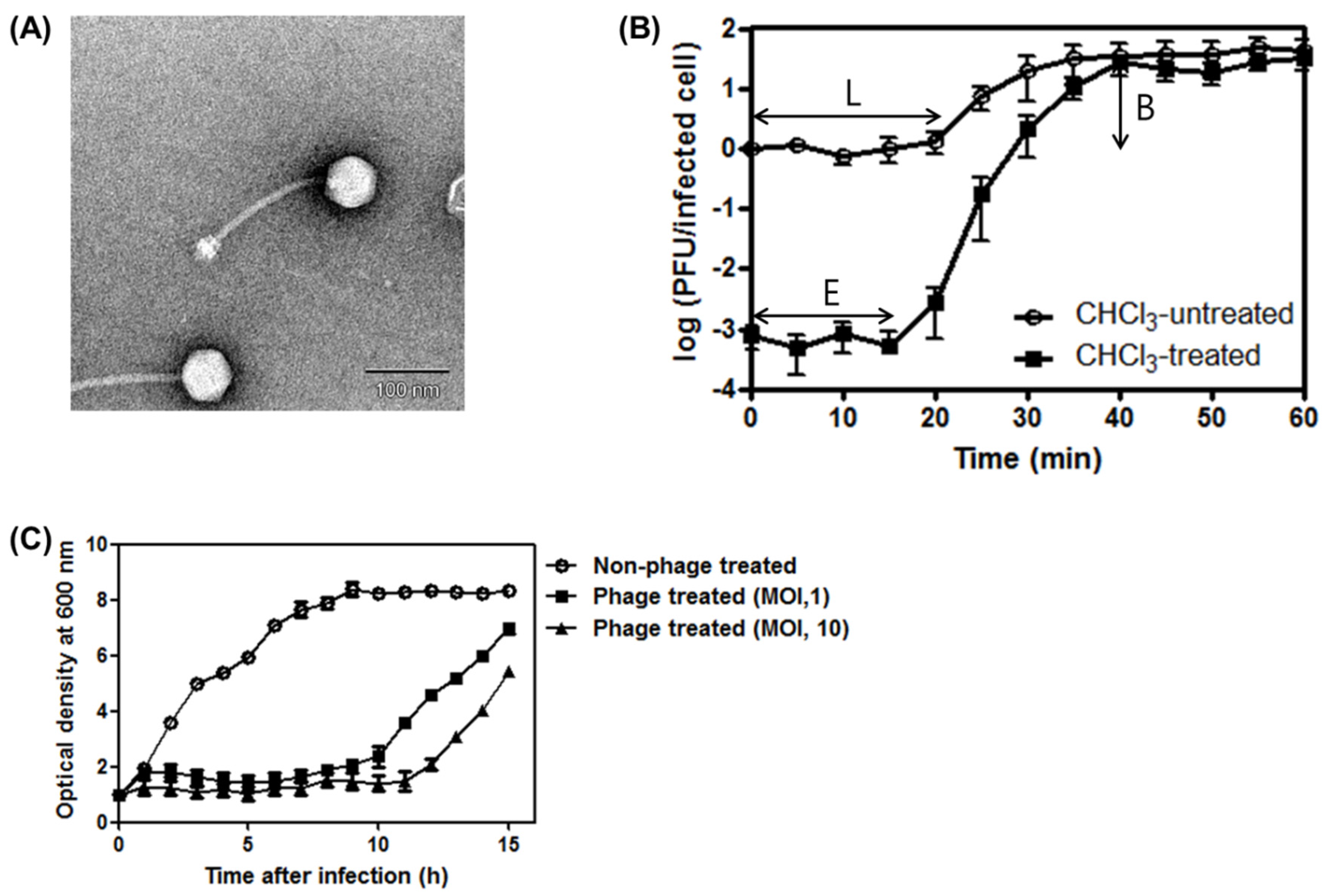
3.1. Phage SA97 Isolation and Its Morphological Analysis
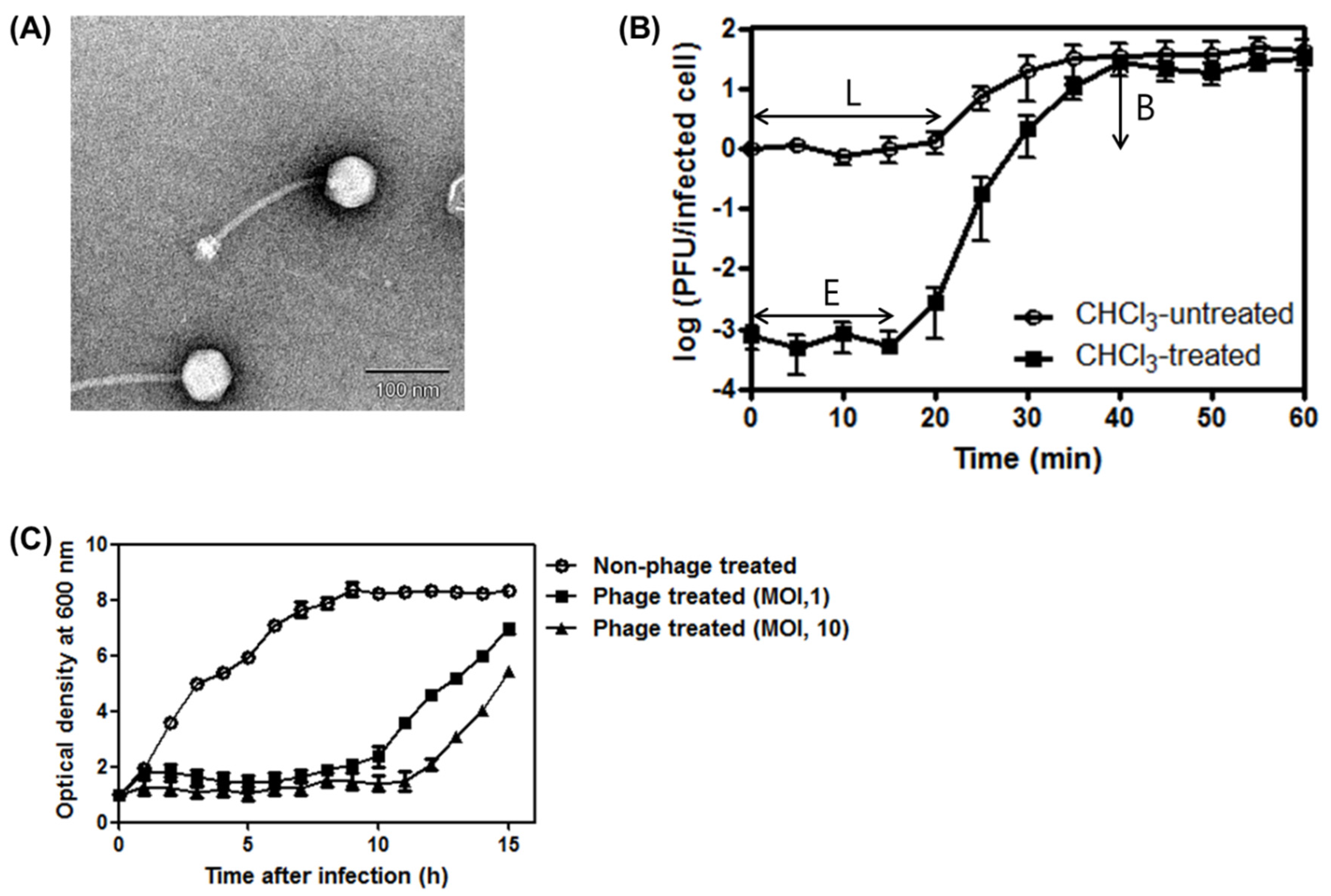
3.2. One-Step Growth Curve of SA97and Bacterial Challenge Assay
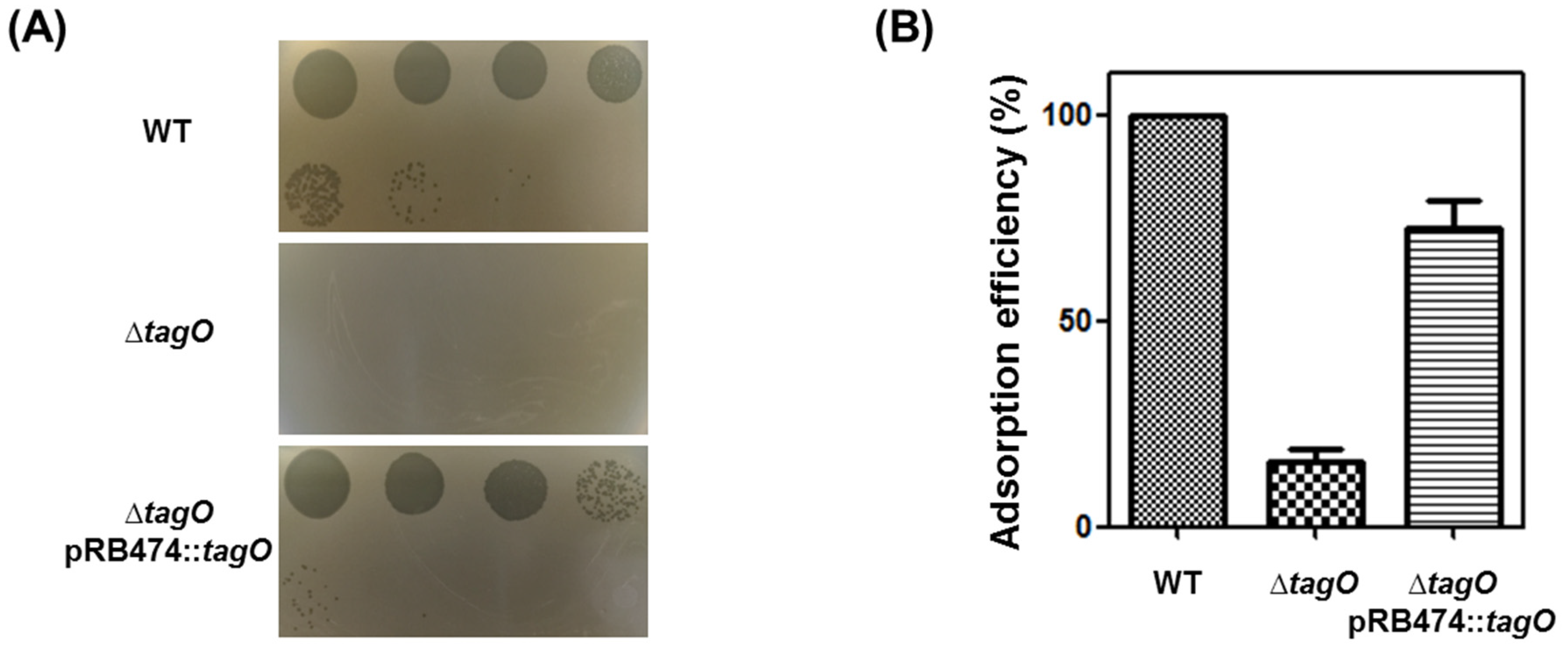
3.3. Receptor Analysis

3.4. Host Range of Bacteriophage SA97
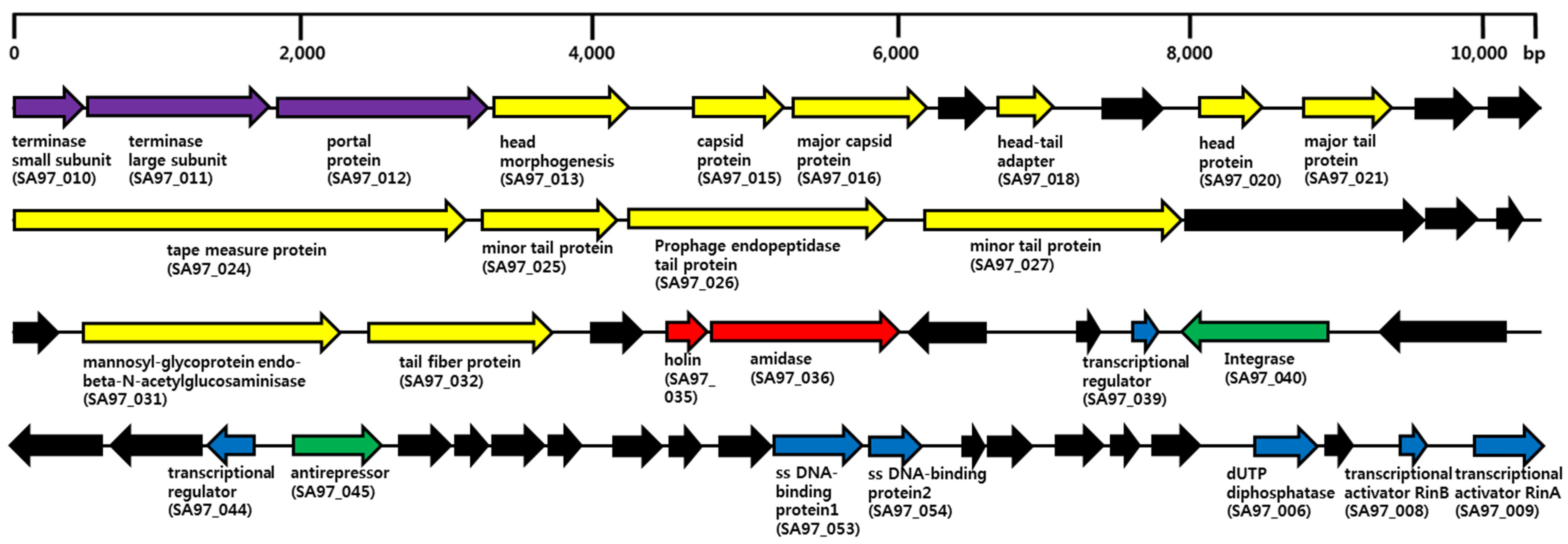
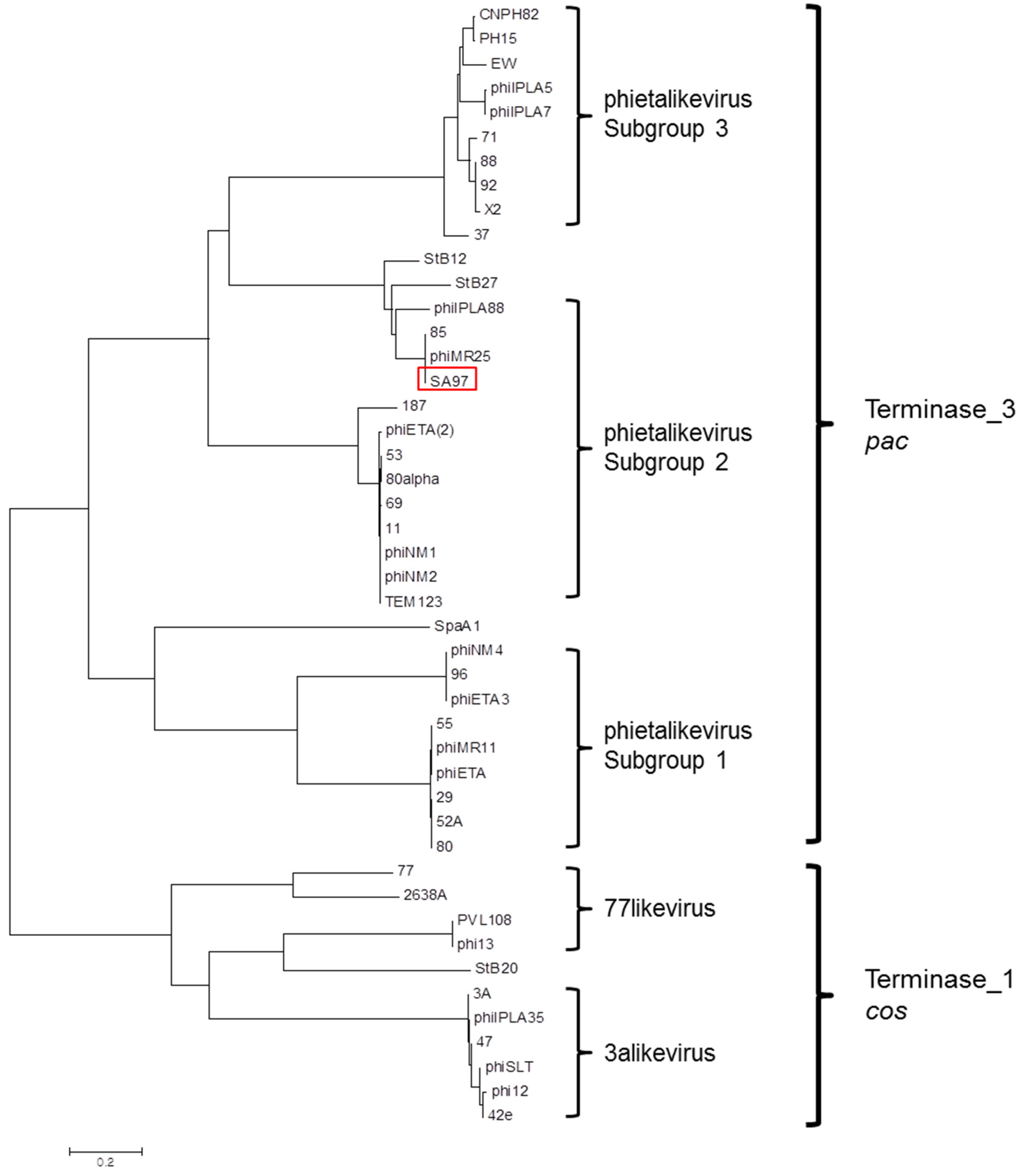
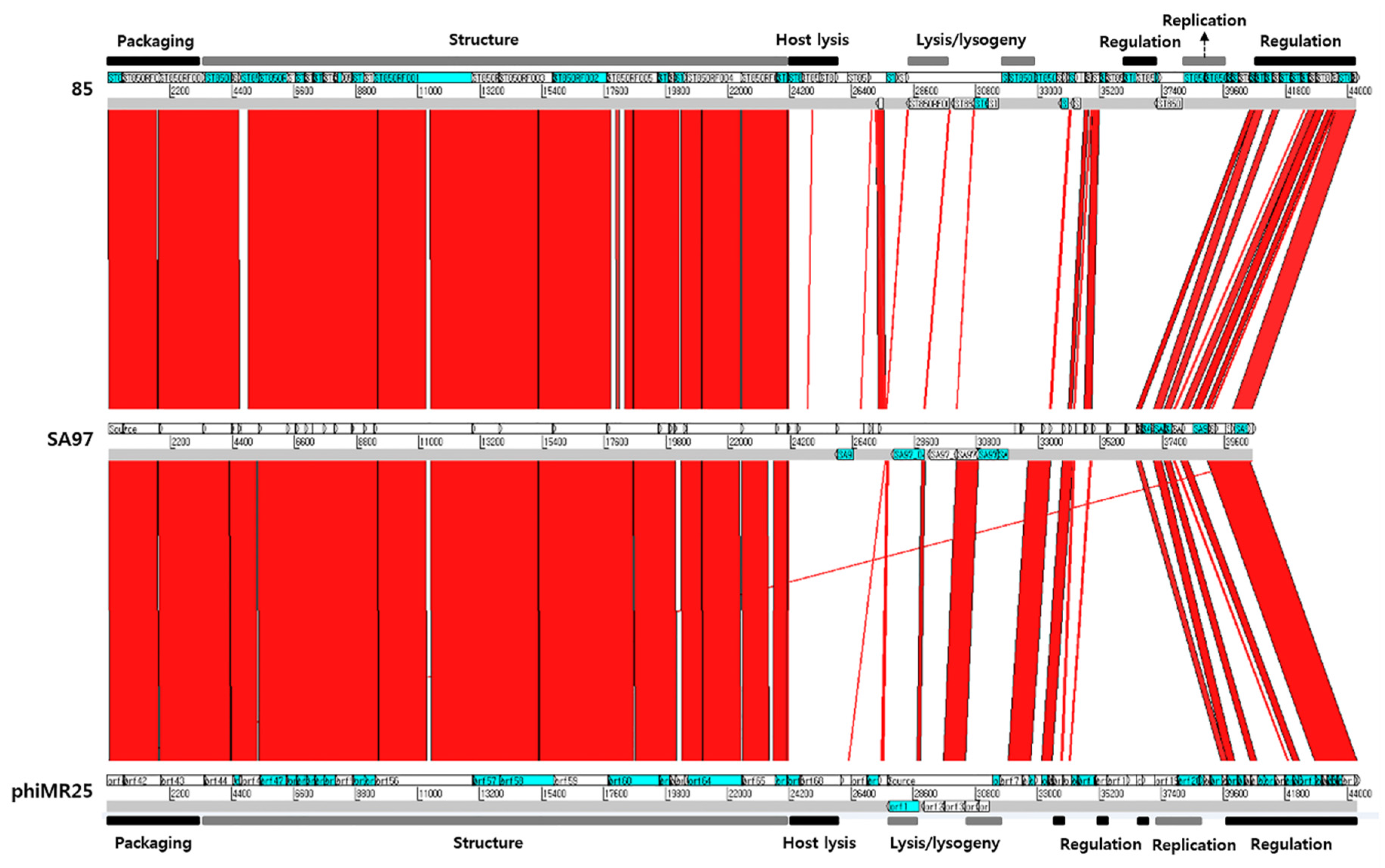
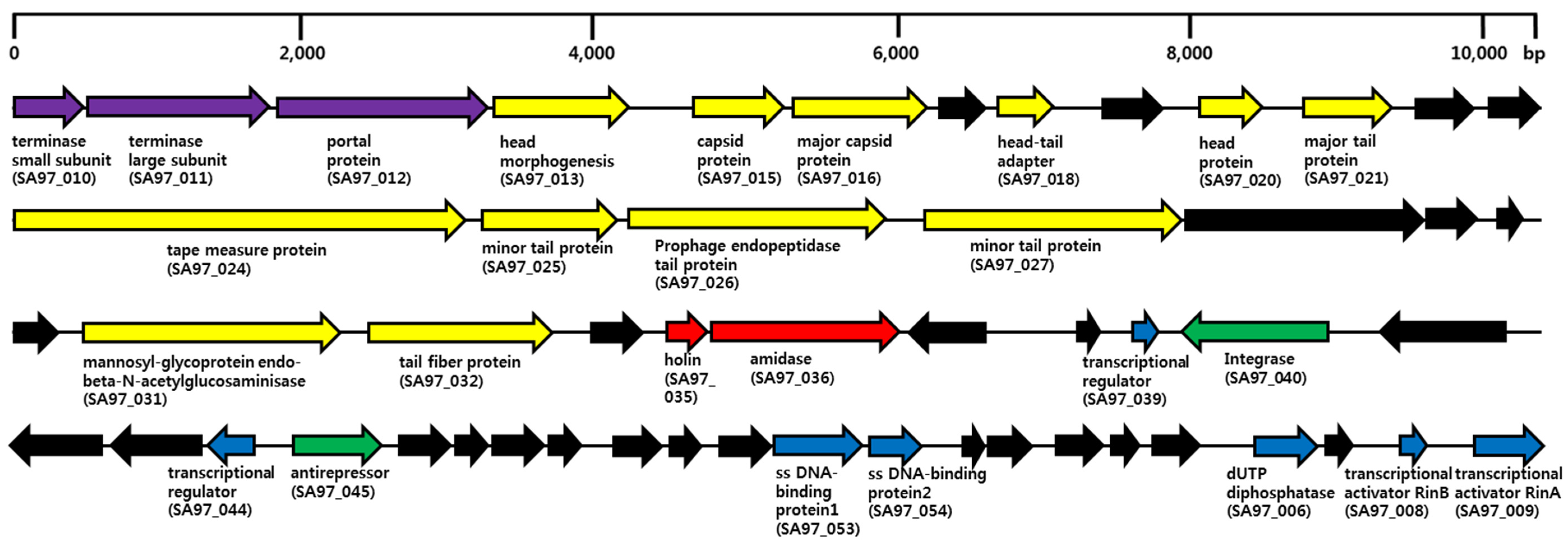
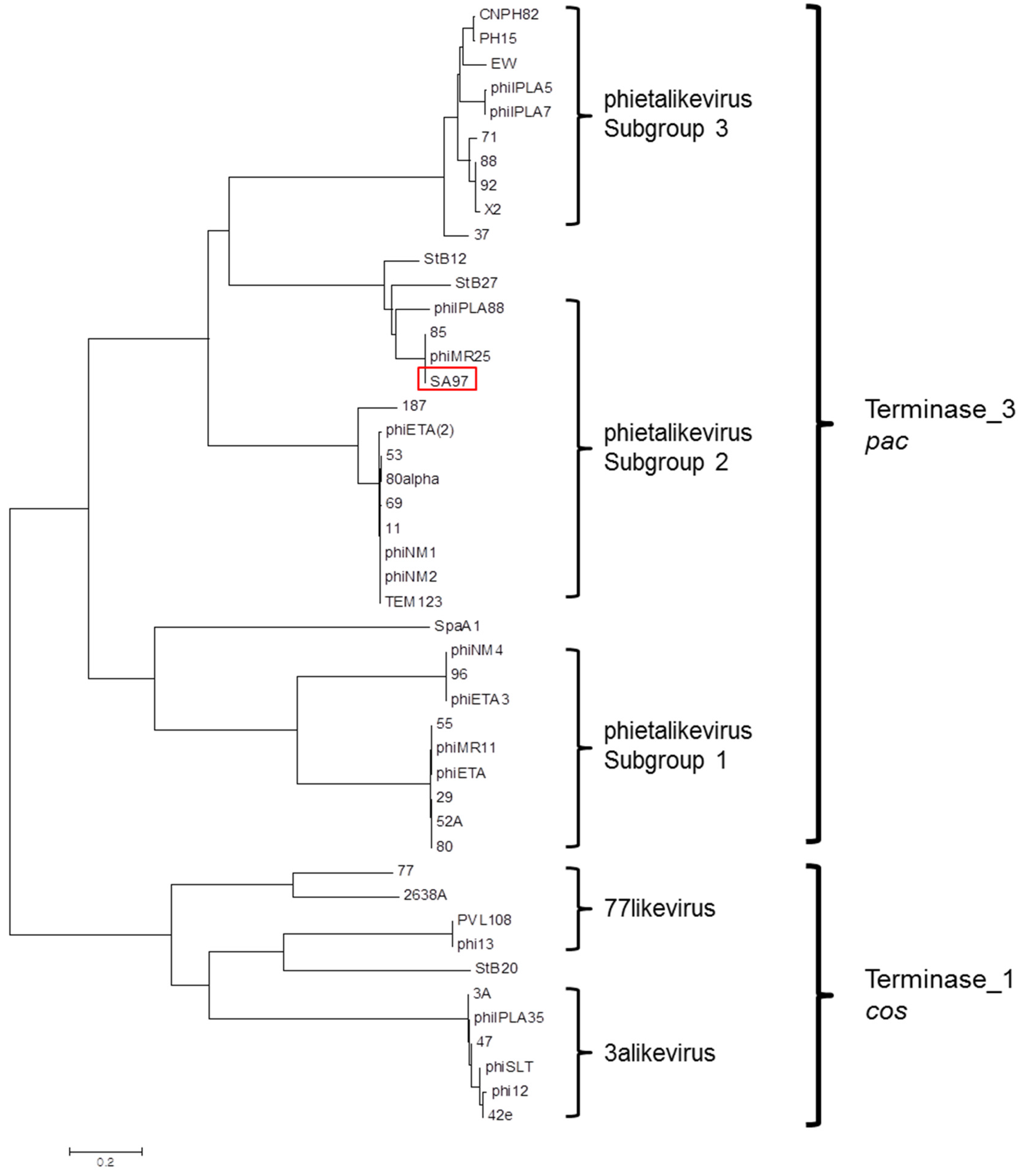
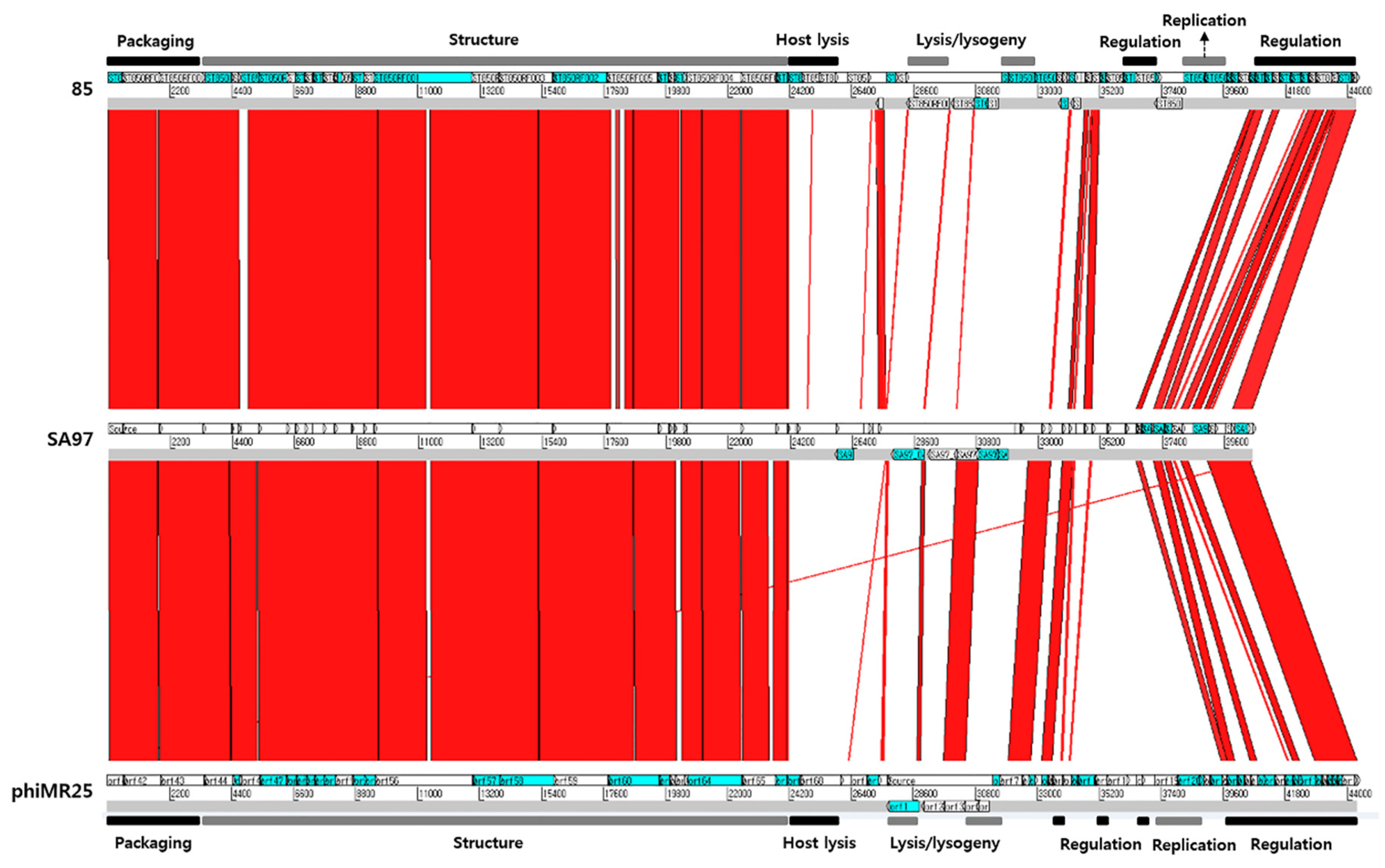
3.5. Bacteriophage SA97 Genomic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Plaque Formation of SA97a | E.O.P b | Reference or Source c |
|---|---|---|---|
| Staphylococcus strains | |||
| Staphylococcus aureus RN4220 | C | 1.0 × 100 ± 0.0 × 100 | [42] |
| Staphylococcus aureus KCTC 1916 | C | 1.1 × 100 ± 1.8 × 10−1 | KCTC |
| Staphylococcus aureus ATCC 29213 | C | 1.7 × 10−4 ± 2.1 × 10−5 | ATCC |
| Staphylococcus aureus ATCC 33593 | C | 1.2 × 100 ± 8.7 × 10−2 | ATCC |
| Methicillin resistant Staphylococcus aureus CCARM 3089 | C | 1.0 × 100 ± 8.1 × 10−2 | CCARM |
| Methicillin resistant Staphylococcus aureus CCARM 3090 | C | 7.4 × 10−2 ± 3.1 × 10−2 | CCARM |
| Staphylococcus aureus food isolate 1 (from vegetable) | C | 4.9 × 10−2 ± 1.6 × 10−2 | This study |
| Staphylococcus aureus food isolate 2 (from pork) | C | 1.2 × 100 ± 1.0 × 10−1 | This study |
| Staphylococcus aureus food isolate 3 (from beef) | C | 1.2 × 100 ± 2.8 × 10−1 | This study |
| Staphylococcus aureus food isolate 4 (from Kimbab) | C | 1.0 × 100 ± 4.0 × 10−1 | This study |
| Staphylococcus aureus clinical isolate 1 | C | 2.0 × 10−4 ± 5.0 × 10−5 | This study |
| Staphylococcus aureus clinical isolate 0055 | C | 1.1 × 10−3 ± 5.5 × 10−5 | GNUH-NCCP |
| Staphylococcus aureus Newman | - | - | [43] |
| Staphylococcus aureus ATCC 6538 | - | - | ATCC |
| Staphylococcus aureus ATCC 23235 | - | - | ATCC |
| Staphylococcus aureus ATCC 25923 | - | - | ATCC |
| Staphylococcus aureus ATCC 12600 | - | - | ATCC |
| Staphylococcus aureus ATCC 13301 | - | - | ATCC |
| Staphylococcus aureus ATCC 27664 | - | - | ATCC |
| Staphylococcus xylosis ATCC 29971 | - | - | ATCC |
| Staphylococcus cohnii ATCC 29974 | - | - | ATCC |
| Staphylococcus warneri ATCC 10209 | - | - | ATCC |
| Staphylococcus intermedius ATCC 29663 | - | - | ATCC |
| Other Gram positive bacteria | |||
| Enterococcus faecalis ATCC 29212 | - | - | ATCC |
| Bacillus cereus ATCC 14579 | - | - | ATCC |
| Bacillus subtilis ATCC 23857 | - | - | ATCC |
| Listeria monocytogenes ATCC 19114 | - | - | ATCC |
| Gram negative bacteria | |||
| Salmonella enterica serovar Typhimurium SL1344 | - | - | ATCC |
| Escherichia coli MG1655 ATCC 47076 | - | - | ATCC |
| Escherichia coli O157:H7 ATCC 35150 | - | - | ATCC |
| Cronobacter sakazakii ATCC 29544 | - | - | ATCC |
| Pseudomonas aeruginosa ATCC 27853 | - | - | ATCC |
| Group | Subgroup | Locus_tag a | Function | BLASTP Best Match | Identity (%) b | Accession no. c |
|---|---|---|---|---|---|---|
| packaging | SA97_010 | terminase small subunit | phage terminase small subunit [Staphylococcus aureus subsp. aureus JH1] | 164/164 (100) | YP_001316047.1 | |
| packaging | SA97_011 | terminase large subunit | PBSX family phage terminase large subunit [Staphylococcus aureus subsp. aureus JH9] | 431/431 (100) | YP_001246264.1 | |
| packaging | SA97_012 | portal protein | SPP1 family phage portal protein [Staphylococcus aureus subsp. aureus JH9] | 512/512 (100) | YP_001246265.1 | |
| structure | head | SA97_013 | head morphogenesis protein | phage head morphogenesis protein [Staphylococcus aureus subsp. aureus JH9] | 331/331 (100) | YP_001246266.1 |
| hypothetical | SA97_014 | hypothetical protein | hypothetical protein phiETA2_gp45 [Staphylococcus phage phiETA2] | 56/56 (100) | YP_001004305.1 | |
| structure | head | SA97_015 | capsid protein | phage capsid protein [Staphylococcus aureus] | 206/206 (100) | WP_000392146.1 |
| structure | head | SA97_016 | major capsid protein | Phage major capsid protein [Staphylococcus aureus subsp. aureus DSM 20231] | 324/324 (100) | WP_001836233.1 |
| hypothetical | SA97_017 | hypothetical protein | phage protein [Staphylococcus aureus subsp. aureus DSM 20231] | 95/95 (100) | ELP28728.1 | |
| structure | head-tail | SA97_018 | head-tail adapter protein | head-tail connector protein [Staphylococcus phage SA12] | 110/110 (100) | YP_008241892.1 |
| hypothetical | SA97_019 | hypothetical protein | hypothetical protein NWMN_1023 [Staphylococcus aureus subsp. aureus str. Newman] | 115/115 (100) | YP_001332057.1 | |
| structure | head | SA97_020 | head protein | hypothetical protein SAV0897 [Staphylococcus aureus subsp. aureus Mu50] | 127/127 (100) | NP_371421.1 |
| structure | tail | SA97_021 | major tail protein | phage major tail protein [Staphylococcus phage phiNM] | 193/193 (100) | YP_873998.1 |
| hypothetical | SA97_022 | hypothetical protein | hypothetical protein SaurJH9_0899 [Staphylococcus aureus subsp. aureus JH9] | 121/121 (100) | YP_001246276.1 | |
| hypothetical | SA97_023 | hypothetical protein | hypothetical protein T874_02808, partial [Staphylococcus aureus SJOS6126] | 114/114 (100) | EVG26589.1 | |
| structure | tail | SA97_024 | tape measure protein | putative tape measure protein [Staphylococcus phage phiSauS-IPLA88] | 1130/1154 (98) | YP_002332525.1 |
| structure | tail | SA97_025 | putative minor tail protein | putative minor tail protein [Staphylococcus phage TEM123] | 314/315 (99) | YP_ 006382275.1 |
| structure | tail | SA97_026 | prophage endopeptidase tail protein | SGNH hydrolase superfamily [Staphylococcus aureus subsp. aureus 21283] | 632/632 (100) | EHO98287.1 |
| structure | tail | SA97_027 | putative minor tail protein | minor structural protein [Staphylococcus aureus] | 635/636 (99) | WP_000369023.1 |
| hypothetical | SA97_028 | hypothetical protein | hypothetical protein NWMN_0306 [Staphylococcus aureus subsp. aureus str. Newman] | 607/607 (100) | YP_001331340.1 | |
| hypothetical | SA97_029 | hypothetical protein | SLT orf 129-like protein [Staphylococcus aureus subsp. aureus JH1] | 125/125 (100) | YP_001316067.1 | |
| hypothetical | SA97_030 | hypothetical protein | hypothetical protein SAP26_gp20 [Staphylococcus phage SAP-26] | 57/57 (100) | YP_003857088.1 | |
| hypothetical | SA97_031 | hypothetical protein | hypothetical protein phiETA2_gp63 [Staphylococcus phage phiETA2] | 99/99 (100) | YP_001004323.1 | |
| structure | tail | SA97_032 | mannosyl-glycoprotein endo-beta-N-acetylglucosminidase | mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase [Staphylococcus aureus subsp. aureus DSM 20231] | 632/632 (100) | WP_001836236.1 |
| structure | tail | SA97_033 | putative tail fiber protein | tail fiber [Staphylococcus aureus subsp. aureus JH1] | 412/412 (100) | YP_001316071.1 |
| hypothetical | SA97_034 | hypothetical protein | hypothetical protein SASA_03740 [Staphylococcus aureus subsp. aureus DSM 20231] | 131/131 (100) | ELP28745.1 | |
| host lysis | SA97_035 | holin | holin, SPP1 family [Staphylococcus aureus subsp. aureus 21267] | 91/91 (100) | EZH90125.1 | |
| host lysis | SA97_036 | amidase | CHAP domain-containing protein [Staphylococcus aureus subsp. aureus JH9] | 470/470 (100) | YP_001246290.1 | |
| hypothetical | SA97_037 | hypothetical protein | hypothetical protein SaurJH1_0932 [Staphylococcus aureus subsp. aureus JH1] | 185/185 (100) | YP_001316075.1 | |
| hypothetical | SA97_038 | hypothetical protein | hypothetical protein SAOUHSC_02018 [Staphylococcus aureus subsp. aureus NCTC 8325] | 50/50 (100) | YP_500515.1 | |
| regulation | SA97_039 | putative transcriptional regulatory protein | XRE family transcriptional regulator [Staphylococcus aureus] | 61/61 (100) | WP_000382163.1 | |
| lysis/lysogeny | SA97_040 | putative integrase | integrase [Staphylococcus aureus subsp. aureus DSM 20231] | 349/349 (100) | WP_001145725.1 | |
| hypothetical | SA97_041 | hypothetical protein | hypothetical protein SAOUHSC_02087 [Staphylococcus aureus subsp. aureus NCTC 8325] | 301/301 (100) | YP_500579.1 | |
| hypothetical | SA97_042 | hypothetical protein | hypothetical protein phiETA2_gp04 [Staphylococcus phage phiETA2] | 241/241 (100) | YP_001004264.1 | |
| hypothetical | SA97_043 | hypothetical protein | hypothetical protein SaurJH1_0868 [Staphylococcus aureus subsp. aureus JH1] | 224/224 (100) | YP_001316011.1 | |
| regulation | SA97_044 | putative transcriptional regulator | XRE family transcriptional regulator [Staphylococcus aureus] | 98/110 (89) | WP_001055143.1 | |
| lysis/lysogeny | SA97_045 | anti-repressor | anti-repressor protein [Staphylococcus phage phiETA2] | 255/255 (100) | YP_001004268.1 | |
| hypothetical | SA97_046 | hypothetical protein | hypothetical protein WWS_01005 [Staphylococcus aureus M1510] | 154/155 (99) | ENN21654.1| | |
| hypothetical | SA97_047 | hypothetical protein | hypothetical protein SAV0856 [Staphylococcus aureus subsp. aureus Mu50] | 74/74 (100) | NP_371380.1 | |
| hypothetical | SA97_048 | hypothetical protein | hypothetical protein phiETA2_gp10 [Staphylococcus phage phiETA2] | 148/149 (99) | YP_001004270.1 | |
| hypothetical | SA97_049 | hypothetical protein | hypothetical protein phiETA3_gp11 [Staphylococcus phage phiETA3] | 73/73 (100) | YP_001004340.1 | |
| hypothetical | SA97_050 | hypothetical protein | hypothetical protein SaurJH1_0876 [Staphylococcus aureus subsp. aureus JH1] | 100/100 (100) | YP_001316019.1 | |
| hypothetical | SA97_051 | hypothetical protein | hypothetical protein SASA_03250 [Staphylococcus aureus subsp. aureus DSM 20231] | 86/86 (100) | ELP28696.1 | |
| hypothetical | SA97_052 | hypothetical protein | hypothetical protein phiSLTp16 [Staphylococcus phage phiSLT] | 177/178 (99) | NP_075479.1 | |
| regulation | SA97_053 | single-stranded DNA-binding protein 1 | ssDNA-binding protein St aphylococcus aureus SA_ST125_MupR] | 216/216 (100) | EPR23947.1 | |
| regulation | SA97_054 | single-stranded DNA-binding protein 2 | ssDNA-binding protein [Staphylococcus phage phiETA3] | 104/105 (99) | YP_001004346.1 | |
| hypothetical | SA97_001 | hypothetical protein | hypothetical protein U622_02779, partial [Staphylococcus aureus F53399] | 61/61 (100) | EWV07862.1 | |
| hypothetical | SA97_002 | hypothetical protein | hypothetical protein SaurJH9_0873 [Staphylococcus aureus subsp. aureus JH9] | 101/101 (100) | YP_001246250.1 | |
| hypothetical | SA97_003 | hypothetical protein | hypothetical protein phiETA3_gp28 [Staphylococcus phage phiETA3] | 118/118 (100) | YP_001004357.1 | |
| hypothetical | SA97_004 | hypothetical protein | hypothetical protein SASA_03390 [Staphylococcus aureus subsp. aureus DSM 20231] | 80/80 (100) | ELP28710.1 | |
| hypothetical | SA97_005 | hypothetical protein | phage protein [Staphylococcus aureus M0157] | 110/110 (100) | EUU37240.1 | |
| regulation | SA97_006 | dUTP diphosphatase | dUTP diphosphatase [Staphylococcus aureus A9765] | 175/176 (99) | WP_001797468.1 | |
| hypothetical | SA97_007 | hypothetical protein | hypothetical protein [Staphylococcus phage phiMR25] | 67/68 (99) | YP_001949833.1 | |
| regulation | SA97_008 | transcriptional activator RinB | transcriptional activator RinB [Staphylococcus aureus subsp. aureus CM05] | 57/57 (100) | WP_001657250.1 | |
| regulation | SA97_009 | transcriptional activator RinA | transcriptional activator RinA [Staphylococcus phage phiETA3] | 140/140 (100) | YP_001004368.1 |
3.6. Determination of the Frequency of BIMs and Horizontal Gene Transfer by SA97
| Recipient Strain | Transduction | Frequency |
|---|---|---|
| SA97 | SA12 | |
| S. aureus RN4220 | 0.0 ± 0.0 | 1.7 × 10−8± 1.2 × 10−8 |
| S. aureus food isolate 1 | 0.0 ± 0.0 | 2.6 × 10−7± 1.5 × 10−7 |
| S. aureus food isolate 2 | 0.0 ± 0.0 | 6.0 × 10−7± 1.6 × 10−7 |
| S. aureus food isolate 3 | 0.0 ± 0.0 | 5.1 × 10−8± 1.2 × 10−8 |
| S. aureus clinical isolate 1 | 0.0 ± 0.0 | 1.3 × 10−7± 0.5 × 10−7 |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mahony, J.; McAuliffe, O.; Ross, R.P.; van Sinderen, D. Bacteriophages as biocontrol agents of food pathogens. Curr. Opin. Biotech. 2011, 22, 157–163. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar]
- Bal, A.M.; Gould, I.M. Antibiotic resistance in Staphylococcus aureus and its relevance in therapy. Expert Opin. Pharmacother. 2005, 6, 2257–2269. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, P.C. The emergence of vancomycin-intermediate and vancomycin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2006, 12, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G., Jr. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Lang, L.H. FDA approves use of bacteriophages to be added to meat and poultry products. Gastroenterology 2006, 131, 1370. [Google Scholar] [CrossRef] [PubMed]
- Guenther, S.; Huwyler, D.; Richard, S.; Loessner, M.J. Virulent bacteriophage for efficient biocontrol of Listeria monocytogenes in ready-to-eat foods. Appl. Environ. Microb. 2009, 75, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Yasuda, M.; Nishikawa, H.; Kuroda, M.; Ujihara, T.; Shuin, T.; Shen, Y.; Jin, Z.; Fujimoto, S.; Nasimuzzaman, M.D.; et al. Experimental protection of mice against lethal Staphylococcus aureus infection by novel bacteriophage phiMR11. J. Infect. Dis. 2003, 187, 613–624. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Ross, R.P.; Meaney, W.; Fitzgerald, G.F.; Elbreki, M.F.; Coffey, A. Potential of the polyvalent anti-Staphylococcus bacteriophage K for control of antibiotic-resistant staphylococci from hospitals. Appl. Environ. Microbiol. 2005, 71, 1836–1842. [Google Scholar] [CrossRef] [PubMed]
- O'Flaherty, S.; Ross, R.P.; Flynn, J.; Meaney, W.J.; Fitzgerald, G.F.; Coffey, A. Isolation and characterization of two anti-staphylococcal bacteriophages specific for pathogenic Staphylococcus aureus associated with bovine infections. Lett. Appl. Microbiol. 2005, 41, 482–486. [Google Scholar] [CrossRef]
- Takemura-Uchiyama, I.; Uchiyama, J.; Kato, S.; Inoue, T.; Ujihara, T.; Ohara, N.; Daibata, M.; Matsuzaki, S. Evaluating efficacy of bacteriophage therapy against Staphylococcus aureus infections using a silkworm larval infection model. FEMS Microbiol. Lett. 2013, 347, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Goerke, C.; Pantucek, R.; Holtfreter, S.; Schulte, B.; Zink, M.; Grumann, D.; Broker, B.M.; Doskar, J.; Wolz, C. Diversity of prophages in dominant Staphylococcus aureus clonal lineages. J. Bacteriology 2009, 191, 3462–3468. [Google Scholar] [CrossRef]
- Blair, J.E.; Carr, M. Lysogeny in staphylococci. J. Bacteriol. 1961, 82, 984–993. [Google Scholar]
- Simatake, H.; Rosenberg, M. Purified lambda regulatory protein CII positively activates promoters for lysogenic development. Nature 1981, 292, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Ptashne, M.; Hopkins, N. The operators controlled by the lambda phage repressor. Proc. Natl. Acad. Sci. USA 1968, 60, 1282–1287. [Google Scholar] [CrossRef] [PubMed]
- Groth, A.C.; Calos, M.P. Phage integrases: Biology and applications. J. Mol. Biol. 2004, 335, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Lee, J.H.; Shin, H.; Heu, S.; Ryu, S. Characterization and complete genome sequence analysis of Staphylococcus aureus bacteriophage SA12. Virus Genes 2013, 47, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Kwiatek, M.; Parasion, S.; Mizak, L.; Gryko, R.; Bartoszcze, M.; Kocik, J. Characterization of a bacteriophage, isolated from a cow with mastitis, that is lytic against Staphylococcus aureus strains. Arch. Virol. 2012, 157, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Rodhain, F. Classification and Nomenclature of Viruses—5th Report of the International Committee for Virus Taxonomy, 5th ed.; Francki, R.I.B., Fauquet, C.M., Knudson, D.I., Brown, F., Eds.; Springer: Verlag Wien, Austria, 1995; p. 95. [Google Scholar]
- Park, M.; Lee, J.H.; Shin, H.; Kim, M.; Choi, J.; Kang, D.H.; Heu, S.; Ryu, S. Characterization and comparative genomic analysis of a novel bacteriophage, SFP10, simultaneously inhibiting both Salmonella enterica and Escherichia coli O157:H7. Appl. Environ. Microb. 2012, 78, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Breidt, F., Jr.; Fleming, H.P.; Altermann, E.; Klaenhammer, T.R. Isolation and characterization of a Lactobacillus plantarum bacteriophage, phiJL-1, from a cucumber fermentation. Int. J. Food Microbiol. 2003, 84, 225–235. [Google Scholar] [CrossRef]
- Kreiswirth, B.N.; Lofdahl, S.; Betley, M.J.; O’Reilly, M.; Schlievert, P.M.; Bergdoll, M.S.; Novick, R.P. The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 1983, 305, 709–712. [Google Scholar] [CrossRef] [PubMed]
- Wann, E.R.; Dassy, B.; Fournier, J.M.; Foster, T.J. Genetic analysis of the cap5 locus of Staphylococcus aureus. FEMS Microbiol. Lett. 1999, 170, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Swoboda, J.G.; Campbell, J.; Meredith, T.C.; Walker, S. Wall teichoic acid function, biosynthesis, and inhibition. Chembiochem 2010, 11, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, R. A series of shuttle vectors for Bacillus subtilis and Escherichia coli. Gene 1992, 122, 187–192. [Google Scholar] [CrossRef]
- Yoon, H.; Yun, J.; Lim, J.A.; Roh, E.; Jung, K.S.; Chang, Y.; Ryu, S.; Heu, S. Characterization and genomic analysis of two Staphylococcus aureus bacteriophages isolated from poultry/livestock farms. J. Gen. Virol. 2013, 94, 2569–2576. [Google Scholar] [CrossRef]
- Garcia, P.; Madera, C.; Martinez, B.; Rodriguez, A. Biocontrol of Staphylococcus aureus in curd manufacturing processes using bacteriophages. Int. Dairy J. 2007, 17, 1232–1239. [Google Scholar] [CrossRef]
- Kiriukhin, M.Y.; Debabov, D.V.; Shinabarger, D.L.; Neuhaus, F.C. Biosynthesis of the glycolipid anchor in lipoteichoic acid of Staphylococcus aureus RN4220: Role of YpfP, the diglucosyldiacylglycerol synthase. J. Bacteriol. 2001, 183, 3506–3514. [Google Scholar] [CrossRef] [PubMed]
- Welker, N.E. Transduction in Bacillus stearothermophilus. J. Bacteriol. 1988, 170, 3761–3764. [Google Scholar]
- Wilcox, S.A.; Toder, R.; Foster, J.W. Rapid isolation of recombinant lambda phage DNA for use in fluorescence in situ hybridization. Chromosome Res. 1996, 4, 397–398. [Google Scholar] [CrossRef] [PubMed]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Lukashin, A.V.; Borodovsky, M. GeneMark.hmm: New solutions for gene finding. Nucleic Acids Res. 1998, 26, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Ziedaite, G.; Daugelavicius, R.; Bamford, J.K.; Bamford, D.H. The Holin protein of bacteriophage PRD1 forms a pore for small-molecule and endolysin translocation. J. Bacteriol. 2005, 187, 5397–5405. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Carver, T.; Berriman, M.; Tivey, A.; Patel, C.; Bohme, U.; Barrell, B.G.; Parkhill, J.; Rahandream, M.A. Artemis and ACT: Viewing, annotating and comparing sequences stored in a relational database. Bioinformatics 2008, 24, 2672–2676. [Google Scholar] [CrossRef] [PubMed]
- Deghorain, M.; Bobay, L.M.; Smeesters, P.R.; Bousbata, S.; Vermeersch, M.; Perez-Morga, D.; Dreze, P.A.; Rocha, E.P.; Touchon, M.; van Melderen, L. Characterization of novel phages isolated in coagulase-negative staphylococci reveals evolutionary relationships with Staphylococcus aureus phages. J. Bacteriol. 2012, 194, 5829–5839. [Google Scholar] [CrossRef] [PubMed]
- Guoqing, X.; Christiane, W. Phages of Staphylococcus aureus and their impact on host evolution. Infect. Genet. Evol. 2014, 21, 593–601. [Google Scholar]
- Han, J.E.; Kim, J.H.; Hwang, S.Y.; Choresca, C.H., Jr.; Shin, S.P.; Jun, J.W.; Chai, J.Y.; Park, Y.H.; Park, S.C. Isolation and characterization of a Myoviridae bacteriophage against Staphylococcus aureus isolated from dairy cows with mastitis. Res. Vet. Sci. 2013, 95, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Corrigan, R.M.; Winstel, V.; Goerke, C.; Grundling, A.; Peschel, A. Wall teichoic Acid-dependent adsorption of staphylococcal siphovirus and myovirus. J. Bacteriol. 2011, 193, 4006–4009. [Google Scholar] [CrossRef]
- Oku, Y.; Kurokawa, K.; Matsuo, M.; Yamada, S.; Lee, B.L.; Sekimizu, K. Pleiotropic roles of polyglycerolphosphate synthase of lipoteichoic acid in growth of Staphylococcus aureus cells. J. Bacteriol. 2009, 191, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Kurokawa, K.; Zheng, L.; Jung, D.J.; Tateishi, K.; Jin, J.O.; Ha, N.C.; Kang, H.J.; Matsushita, M.; Kwak, J.Y.; et al. Human serum mannose-binding lectin senses wall teichoic acid Glycopolymer of Staphylococcus aureus, which is restricted in infancy. J. Biol. Chem. 2010, 285, 27167–27175. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Bae, T.; Schneewind, O.; Takeuchi, F.; Hiramatsu, K. Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes: Polymorphism and evolution of two major pathogenicity islands. J. Bacteriol. 2008, 190, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Loessner, M.J. Bacteriophage endolysins-current state of research and applications. Curr. Opin. Microbiol. 2005, 8, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rubio, L.; Martinez, B.; Donovan, D.M.; Rodriguez, A.; Garcia, P. Bacteriophage virion-associated peptidoglycan hydrolases: Potential new enzybiotics. Crc. Cr. Rev. Microbiol. 2013, 39, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, D.; Adriaenssens, E.M.; Martinez, B.; Rodriguez, A.; Lavigne, R.; Kropinski, A.M.; Garcia, P. Three proposed new bacteriophage genera of staphylococcal phages: “3alikevirus”, “77likevirus”, “Phietalikevirus”. Arch. Virol. 2014, 159, 389–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, T.; Liu, J.; DuBow, M.; Gros, P.; Pelletier, J. The complete genomes and proteomes of 27 Staphylococcus aureus bacteriophages. Proc. Natl. Acad. Sci. USA 2005, 102, 5174–5179. [Google Scholar] [CrossRef] [PubMed]
- Hoshiba, H.; Uchiyama, J.; Kato, S.; Ujihara, T.; Muraoka, A.; Daibata, M.; Wakiguchi, H.; Matsuzaki, S. Isolation and characterization of a novel Staphylococcus aureus bacteriophage, phiMR25, and its therapeutic potential. Arch. Virol. 2010, 155, 545–552. [Google Scholar]
- Chai, S.; Bravo, A.; Luder, G.; Nedlin, A.; Trautner, T.A.; Alonso, J.C. Molecular analysis of the Bacillus subtilis bacteriophage SPP1 region encompassing genes 1 to 6. J. Mol. Biol. 1992, 224, 87–102. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.; Shin, H.; Lee, J.-H.; Park, C.J.; Paik, S.-Y.; Ryu, S. Isolation and Genome Characterization of the Virulent Staphylococcus aureus Bacteriophage SA97. Viruses 2015, 7, 5225-5242. https://doi.org/10.3390/v7102870
Chang Y, Shin H, Lee J-H, Park CJ, Paik S-Y, Ryu S. Isolation and Genome Characterization of the Virulent Staphylococcus aureus Bacteriophage SA97. Viruses. 2015; 7(10):5225-5242. https://doi.org/10.3390/v7102870
Chicago/Turabian StyleChang, Yoonjee, Hakdong Shin, Ju-Hoon Lee, Chul Jong Park, Soon-Young Paik, and Sangryeol Ryu. 2015. "Isolation and Genome Characterization of the Virulent Staphylococcus aureus Bacteriophage SA97" Viruses 7, no. 10: 5225-5242. https://doi.org/10.3390/v7102870