
Differentially-Expressed Pseudogenes in HIV-1 Infection


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. HIV-1 Infection
2.3. RNA Sequencing
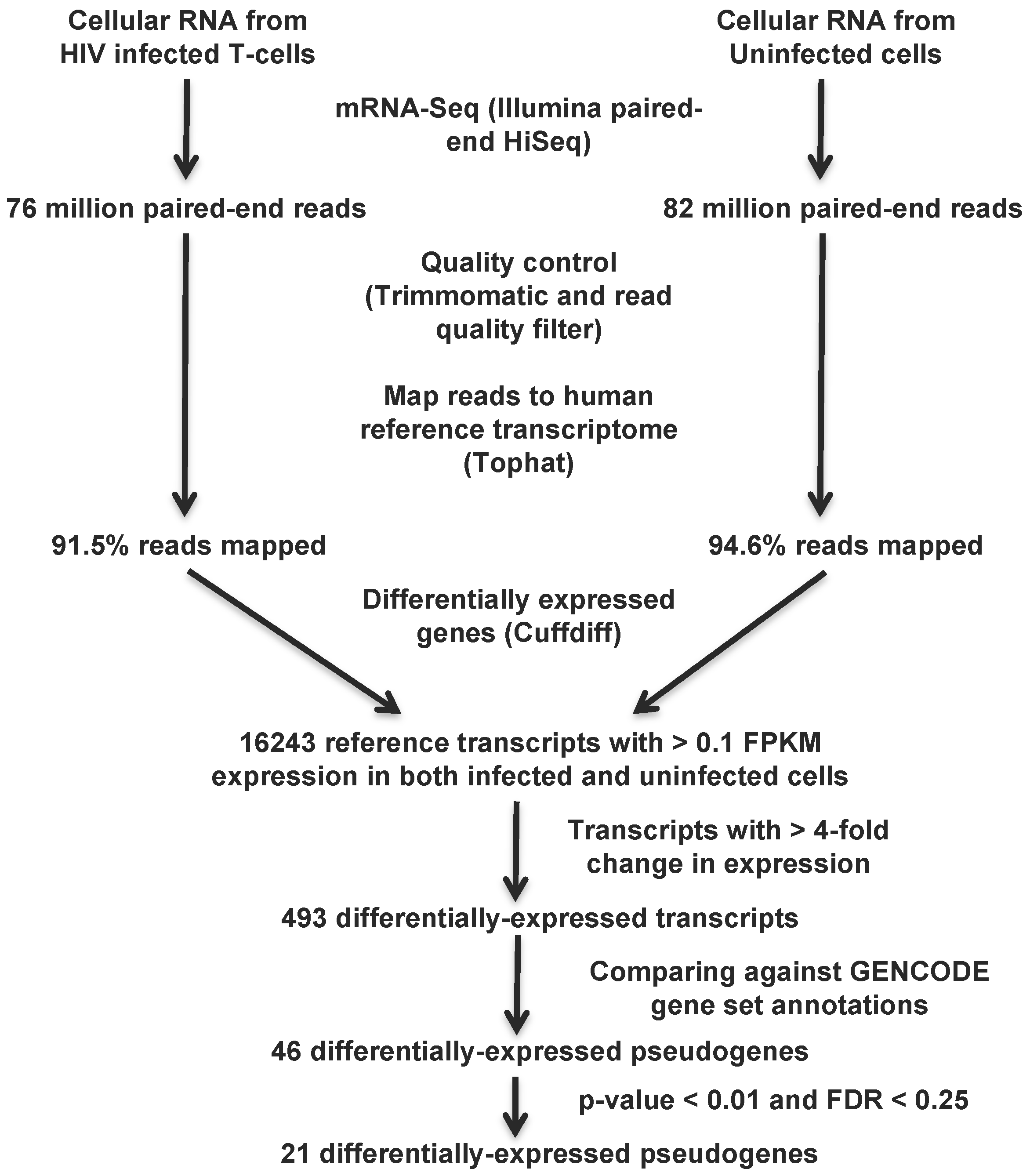
2.4. Pipeline to Identify Differentially-Expressed Pseudogenes
2.5. Bioinformatics Analysis of Candidate Pseudogenes
2.6. Analyses of Publicly-Available Transcriptomic Data
3. Results

{kind=link}
| Pseudogene | Type | Expression in Uninfected T-Cells | Expression in HIV-1 Infected T-Cells | (Fold Change) | p-value | Q-value |
|---|---|---|---|---|---|---|
| A. Over-expressed | ||||||
| DUTP1 | Processed | 0.159 | 2.321 | 3.869 | 1.85e-06 | 0.0012 |
| RP1-89D4.1 | Processed | 0.200 | 1.377 | 2.782 | 0.0006 | 0.0696 |
| RP11-720N19.1 | Processed | 0.102 | 0.676 | 2.722 | 0.0007 | 0.0824 |
| UBE2FP1 | Processed | 1.277 | 7.716 | 2.595 | 0.0013 | 0.1183 |
| RP11-170L3.6 | Processed | 0.312 | 1.887 | 2.595 | 0.0013 | 0.1183 |
| MTATP8P2 | Processed | 2.744 | 16.526 | 2.59 | 0.0013 | 0.1193 |
| RP11-265N6.3 | Processed | 0.106 | 0.617 | 2.536 | 0.0017 | 0.1348 |
| MTND4P15 | Unprocessed | 0.362 | 2.097 | 2.535 | 0.0017 | 0.1348 |
| IFNL3P1 | Unprocessed | 0.128 | 0.632 | 2.305 | 0.0042 | 0.2537 |
| RP11-44M6.3 | Processed | 0.255 | 1.235 | 2.275 | 0.0048 | 0.2693 |
| CTD-2611O12.6 | Processed | 0.183 | 0.882 | 2.266 | 0.0049 | 0.2736 |
| HMGB3P24 | Processed | 0.11 | 0.504 | 2.191 | 0.0065 | 0.3153 |
| AOC4P | Unprocessed | 0.126 | 0.546 | 2.121 | 0.0084 | 0.3688 |
| B. Under-expressed | ||||||
| RP11-380G5.3 | Processed | 1.131 | 0.103 | −3.463 | 2.868e-05 | 0.0103 |
| ZNF137P | Unprocessed | 1.203 | 0.141 | −3.092 | 0.0002 | 0.0353 |
| RP11-114F3.5 | Processed | 0.924 | 0.113 | −3.034 | 0.0003 | 0.0415 |
| SCML2P2 | Processed | 0.955 | 0.119 | −3.003 | 0.0003 | 0.0439 |
| ANTXRLP1 | Unprocessed | 1.507 | 0.207 | −2.865 | 0.0006 | 0.0686 |
| HNRNPA3P6 | Processed | 0.966 | 0.138 | −2.812 | 0.0007 | 0.078 |
| RP1-224A6.8 | Processed | 0.903 | 0.129 | −2.808 | 0.0007 | 0.0787 |
| AC010733.5 | Processed | 3.683 | 0.555 | −2.731 | 0.001 | 0.1026 |
| RP11-411B10.4 | Unprocessed | 0.943 | 0.152 | −2.632 | 0.0015 | 0.1284 |
| RP11-490K7.4 | Processed | 2.187 | 0.358 | −2.613 | 0.0017 | 0.1345 |
| KLHL2P1 | Unprocessed | 0.692 | 0.113 | −2.611 | 0.0017 | 0.1348 |
| RP11-471L13.3 | Processed | 1.6 | 0.263 | −2.608 | 0.0017 | 0.1348 |
| CTD-2008A1.2 | Unprocessed | 1.652 | 0.276 | −2.58 | 0.0019 | 0.1457 |
| GUSBP2 | Unprocessed | 1.124 | 0.217 | −2.375 | 0.0043 | 0.2551 |
| RP11-1166P10.1 | Unprocessed | 0.766 | 0.154 | −2.311 | 0.0055 | 0.2849 |
| AKR7A2P1 | Processed | 0.46 | 0.1 | −2.199 | 0.0083 | 0.3665 |
| ADAMTS7P4 | Unprocessed | 1.06 | 0.234 | −2.182 | 0.0088 | 0.3744 |
3.1. Pseudogenes Derived from Over-Expressed Genes
| Pseudogene | Chromosome | (fc) | BLASTX Hits | Chromosome | Mock | HIV | (fc) |
|---|---|---|---|---|---|---|---|
| A. Over-expressed | |||||||
| DUTP1 | 3 | 3.869 | DUT | 15 | 181.046 | 128.614 | −0.812 |
| UBE2FP1 | 3 | 2.595 | UBE2F | 2 | 17.448 | 20.262 | 0.216 |
| MTATP8P2 | 2 | 2.59 | MT-ATP8 | MT | 1.3e+05 | 1.5e+05 | 0.144 |
| MTND4P15 | 9 | 2.535 | MT-ND4 | MT | 3.8e+04 | 3.6e+04 | −0.07 |
| RP1-89D4.1 | 11 | 2.782 | RPS24 | 10 | 6416.6 | 5334.27 | −0.267 |
| RP11-720N19.1 | 17 | 2.722 | MSANTD3 | 9 | 9.694 | 9.455 | −0.036 |
| RP11-170L3.6 | 16 | 2.595 | IGHV4-31 | 14 | 0.806 | 0 | -inf |
| IGHV4-39 | 14 | - | - | - | |||
| RP11-265N6.3 | 15 | 2.536 | MYL12B | 18 | 100.483 | 126.451 | 0.332 |
| MYL12A | 18 | 255.43 | 271.675 | 0.089 | |||
| B. Under-expressed | |||||||
| ZNF137P | 19 | −3.092 | ZNF816 | 19 | 4.178 | 3.376 | −0.307 |
| ZNF813 | 19 | 1.846 | 0.919 | −1.007 | |||
| ZNF845 | 19 | 4.772 | 4.083 | −0.225 | |||
| ZNF83 | 19 | 12.51 | 8.327 | −0.587 | |||
| FKSG61 | 14 | 6.17 | 6.243 | 0.017 | |||
| SCML2P2 | 16 | −3.003 | SCMH1 | 1 | 6.914 | 5.049 | −0.453 |
| ANTXRLP1 | 10 | −2.865 | ANTXRL | 10 | 0.019 | 0 | -inf |
| HNRNPA3P6 | 3 | −2.812 | HNRNPA3 | 2 | 330.731 | 185.908 | −0.831 |
| HNRNPA1 | 12 | 1293.29 | 905.014 | −0.515 | |||
| KLHL2P1 | 4 | −2.611 | TMEM135 | 11 | 17.787 | 15.472 | −0.201 |
| KLHL2 | 4 | 9.699 | 11.424 | 0.236 | |||
| MYB | 6 | 87.15 | 78.048 | −0.159 | |||
| PKN1 | 19 | 25.755 | 27.804 | 0.11 | |||
| CTD-2008A1.2 | 15 | −2.58 | SORD | 15 | 25.16 | 12.187 | −1.046 |
| RP11-380G5.3 | 10 | −3.463 | RPL11 | 1 | 2653.49 | 2219.2 | −0.258 |
| RP11-114F3.5 | 12 | −3.034 | HKR1 | 19 | 17.206 | 12.676 | −0.441 |
| CRLF2 | X | 0.149 | 1.557 | 3.387 | |||
| TSEN2 | 3 | 10.77 | 6.858 | −0.651 | |||
| SEPSECS | 4 | 4.842 | 2.855 | −0.762 | |||
| MRPS25 | 3 | 31.181 | 28.143 | −0.148 | |||
| CLTA | 9 | 24.205 | 34.036 | 0.492 | |||
| RP1-224A6.8 | 1 | −2.808 | MPHOSPH6 | 16 | 22.657 | 22.301 | −0.023 |
| AC010733.5 | 2 | −2.731 | RPS12 | 6 | 7413.97 | 8991.73 | 0.278 |
| RP11-411B10.4 | 18 | −2.632 | VN1R4 | 19 | - | - | - |
| VN1R2 | 19 | - | - | - | |||
| VN1R1 | 19 | 4.273 | 1.546 | −1.466 | |||
| LEPRE1 | 1 | 6.65 | 8.324 | 0.324 | |||
| RP11-490K7.4 | 1 | −2.613 | GTF2A2 | 15 | 108.131 | 143.993 | 0.413 |
| STAP2 | 19 | 19.65 | 20.589 | 0.067 | |||
| STARD10 | 11 | 49.338 | 47.267 | −0.062 | |||
| ADAM10 | 15 | 37.318 | 21.645 | −0.786 | |||
| RP11-471L13.3 | 17 | −2.608 | DYNLT1 | 6 | 204.754 | 140.976 | −0.539 |
| DYNLT3 | X | 16.799 | 20.654 | 0.298 | |||
| Pseudogene | Chromosome | (fc) | BLASTN Hits | Chromosome | Mock | HIV | –(fc) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A. Over-expressed | |||||||||||||
| DUTP1 | 3 | 3.869 | - | - | - | - | - | ||||||
| UBE2FP1 | 3 | 2.595 | - | - | - | - | - | ||||||
| MTATP8P2 | 2 | 2.59 | MT-ATP8 | MT | 1.3e+05 | 1.5e+05 | 0.144 | ||||||
| MT-ATP6 | MT | 133,475 | 147,477 | 0.144 | |||||||||
| MTND4P15 | 9 | 2.535 | - | - | - | - | - | ||||||
| RP1-89D4.1 | 11 | 2.782 | RPS24 | 10 | 6416.6 | 5334.27 | −0.267 | ||||||
| RP11-720N19.1 | 17 | 2.722 | MSANTD3 | 9 | 9.694 | 9.455 | −0.036 | ||||||
| RP11-170L3.6 | 16 | 2.595 | IGHVII-15-1 | 14 | 0 | 0 | 0 | ||||||
| IGHV4-34 | 14 | 0 | 0 | 0 | |||||||||
| RP11-265N6.3 | 15 | 2.536 | MYL12B | 18 | 100.483 | 126.451 | 0.332 | ||||||
| B. Under-expressed | |||||||||||||
| ZNF137P | 19 | −3.092 | PIGL | 17 | 39.2801 | 37.686 | −0.06 | ||||||
| TBC1D9B | 5 | 24.227 | 48.892 | 1.013 | |||||||||
| C2CD3 | 11 | 5.705 | 6.285 | 0.14 | |||||||||
| HLA-DQA1 | 6 | 47.241 | 35.167 | −0.426 | |||||||||
| ORC6 | 16 | 14.991 | 19.302 | 0.365 | |||||||||
| SCML2P2 | 16 | −3.003 | - | - | - | - | - | ||||||
| ANTXRLP1 | 10 | −2.865 | ANTXRL | 10 | 0.019 | 0 | -inf | ||||||
| SOD2 | 6 | 60.695 | 112.151 | 0.886 | |||||||||
| HNRNPA3P6 | 3 | −2.812 | HNRNPA3 | 2 | 330.731 | 185.908 | −0.831 | ||||||
| KLHL2P1 | 4 | −2.611 | KLHL2 | 4 | 9.699 | 11.424 | 0.236 | ||||||
| FOXK1 | 7 | 4.528 | 6.016 | 0.41 | |||||||||
| DNAJC21 | 5 | 25.755 | 17.957 | −0.52 | |||||||||
| BIRC5 | 17 | 35.427 | 21.565 | −0.716 | |||||||||
| TRIM59 | 3 | 70.348 | 43.319 | −0.699 | |||||||||
| CTD-2008A1.2 | 15 | −2.58 | SORD | 15 | 25.16 | 12.187 | −1.046 | ||||||
| RP11-380G5.3 | 10 | −3.463 | DOK1 | 2 | 5.483 | 11.781 | 1.104 | ||||||
| RP11-114F3.5 | 12 | −3.034 | IDS | X | 2.891 | 3.413 | 0.239 | ||||||
| AS3MT | 10 | 20.487 | 16.317 | −0.328 | |||||||||
| THAP6 | 4 | 7.924 | 4.931 | −0.684 | |||||||||
| ZYG11B | 1 | 4.275 | 2.514 | −0.766 | |||||||||
| ABCC10 | 6 | 8.517 | 6.681 | −0.35 | |||||||||
| RP1-224A6.8 | 1 | −2.808 | MPHOSPH6 | 16 | 22.657 | 22.301 | −0.023 | ||||||
| AC010733.5 | 2 | −2.731 | RPS12 | 6 | 7413.97 | 8991.73 | 0.278 | ||||||
| RP11-411B10.4 | 18 | −2.632 | ADC | 1 | 0.768 | 1.148 | 0.58 | ||||||
| SLC2A5 | 1 | 0.498 | 0.361 | −0.464 | |||||||||
| SETD2 | 3 | 12.064 | 9.976 | −0.274 | |||||||||
| RP11-490K7.4 | 1 | −2.613 | GTF2A2 | 15 | 108.131 | 143.993 | 0.413 | ||||||
| RP11-471L13.3 | 17 | −2.608 | DYNLT1 | 6 | 204.754 | 140.976 | −0.539 | ||||||
3.2. Pseudogenes with Antagonistic Expression to the Parent Genes
| Pseudogene | 12 h | 24 h | 7 d | Parent Gene | 12 h | 24 h | 7 d |
|---|---|---|---|---|---|---|---|
| DUTP1 | −0.908 | on | 3.869 | DUT | 0.245 | −0.527 * | −0.812 |
| RP11-265N6.3 | 1.082 | on | 2.536 | MYL12A | 0.619 * | −0.624 * | 0.089 |
| ZNF137P | −0.031 | −1.007 | −3.092 | TBC1D9B | 0.229 | 0.18 * | 1.013 |
| C2CD3 | −0.6 | −0.21 * | 0.14 | ||||
| ZNF813 | −0.212 | −0.298 * | −1.007 | ||||
| ZNF83 | 0.062 | 0.62 * | −0.587 | ||||
| ANTXRLP1 | off | −3.75 | −2.865 | ANTXRL | - | −1.086 | off |
| HNRNPA3P6 | 0.406 | −3.517 | −2.812 | HNRNPA3 | −0.231 | −0.962 * | −0.831 |
| HNRNPA1 | −0.14 | −0.903 * | −0.515 | ||||
| KLHL2P1 | on | 0.017 | −2.611 | KLHL2 | −0.043 | 0.79 | 0.236 |
| FOXK1 | −0.68 * | −1.075 * | 0.41 | ||||
| BIRC5 | −0.238 | −1.163 * | −0.716 | ||||
| TRIM59 | −0.1 | −0.22 | −0.699 | ||||
| MYB | −0.694 * | −1.915 * | −0.159 | ||||
| CTD-2008A1.2 | 0.131 | −0.319 | −2.58 | SORD | 0.055 | −1.928 * | −1.046 |
| RP11-380G5.3 | −0.083 | 0.285 | −3.463 | DOK1 | 0.494 | −0.252 * | 1.104 |
| RP11-114F3.5 | −0.438 | −0.0628 | −3.034 | CRLF2 | −1.08 | on * | 3.387 |
| TSEN2 | −0.423 * | −1.647 * | −0.651 | ||||
| SEPSECS | −0.003 | −0.213 * | −0.762 | ||||
| CLTA | −0.05 | −1.255 * | 0.492 | ||||
| THAP6 | 0.474 * | 0.806 * | −0.684 | ||||
| ZYG11B | −0.099 | −0.093 | −0.766 | ||||
| ABCC10 | −0.594 * | 0.419 * | −0.35 | ||||
| RP1-224A6.8 | 0.096 | on | −2.808 | MPHOSPH6 | 0.123 | −0.618 * | −0.023 |
| RP11-411B10.4 | 0.02 | −0.53 | −2.632 | ADC | 0.24 | 3.235 * | 0.58 |
| SLC2A5 | −1.698 | 1.908 * | −0.464 | ||||
| VN1R1 | 0.296 * | −0.425 * | −1.466 | ||||
| RP11-490K7.4 | 0.346 | 0.069 | −2.613 | GTF2A2 | 0.031 | −0.83* | 0.413 |
| ADAM10 | −0.115 | −0.12 | −0.786 | ||||
| STAP2 | −0.358 | 0.704 | 0.067 | ||||
| RP11-471L13.3 | on | 0.839 | −2.608 | DYNLT1 | 0.323 * | 0.626 * | −0.539 |
| DYNLT3 | −0.099 | −0.67 * | 0.298 |
3.3. Pseudogenes with Synergistic Expression to the Parent Genes
3.4. Pseudogenes and Their Parent Genes with Modulating Gene Expression in Early and Late HIV-1 Infection
4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflict of Interest
References
- Pink, R.C.; Wicks, K.; Caley, D.P.; Punch, E.K.; Jacobs, L.; Carter, D.R.F. Pseudogenes: Pseudo-functional or key regulators in health and disease? RNA 2011, 17, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Yano, Y.; Saito, R.; Yoshida, N.; Yoshiki, A.; Wynshaw-Boris, A.; Tomita, M.; Hirotsune, S. A new role for expressed pseudogenes as ncRNA: Regulation of mRNA stability of its homologous coding gene. J. Mol. Med. 2004, 82, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Frankish, A.; Baertsch, R.; Kapranov, P.; Reymond, A.; Choo, S.W.; Lu, Y.; Denoeud, F.; Antonarakis, S.E.; Snyder, M.; et al. Pseudogenes in the ENCODE regions: Consensus annotation, analysis of transcription, and evolution. Genome Res. 2007, 17, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.P.; Munjaal, R.P.; Lagace, L.; Lai, E.C.; O’Malley, B.W.; Means, A.R. Tissue-specific expression of a chicken calmodulin pseudogene lacking intervening sequences. Proc. Natl. Acad. Sci. USA 1983, 80, 6485–6489. [Google Scholar] [CrossRef] [PubMed]
- Grunau, C.; Hindermann, W.; Rosenthal, A. Large-scale methylation analysis of human genomic DNA reveals tissue-specific differences between the methylation profiles of genes and pseudogenes. Hum. Mol. Genet. 2000, 9, 2651–2663. [Google Scholar] [CrossRef] [PubMed]
- Kalyana-Sundaram, S.; Kumar-Sinha, C.; Shankar, S.; Robinson, D.R.; Wu, Y.M.; Cao, X.; Asangani, I.A.; Kothari, V.; Prensner, J.R.; Lonigro, R.J.; et al. Expressed pseudogenes in the transcriptional landscape of human cancers. Cell 2012, 149, 1622–1634. [Google Scholar] [CrossRef] [PubMed]
- Wezel, F.; Pearson, J.; Kirkwood, L.A.; Southgate, J. Differential expression of Oct4 variants and pseudogenes in normal urothelium and urothelial cancer. Am. J. Pathol. 2013, 183, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Costa, V.; Esposito, R.; Aprile, M.; Ciccodicola, A. Non-coding RNA and pseudogenes in neurodegenerative diseases: “The (un)Usual Suspects”. Front. Genet. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.L.; Shlien, A.; Marshall, J.; Pipinikas, C.P.; Martincorena, I.; Tubio, J.M.C.; Li, Y.; Menzies, A.; Mudie, L.; Ramakrishna, M.; et al. Processed pseudogenes acquired somatically during cancer development. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poliseno, L. Pseudogenes: Newly discovered players in human cancer. Sci. Signal. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Yuan, Y.; Zheng, S.; Yang, Y.; Li, J.; Edgerton, M.E.; Diao, L.; Xu, Y.; Verhaak, R.G.W.; Liang, H. The Pan-Cancer analysis of pseudogene expression reveals biologically and clinically relevant tumour subtypes. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.L.; Yuo, C.Y.; Yang, W.K.; Hung, S.Y.; Chang, Y.S.; Chiu, C.C.; Yeh, K.T.; Huang, H.D.; Chang, J.G. Transcribed pseudogene ψPPM1K generates endogenous siRNA to suppress oncogenic cell growth in hepatocellular carcinoma. Nucleic Acids Res. 2013, 41, 3734–3747. [Google Scholar] [CrossRef] [PubMed]
- Korneev, S.A.; Park, J.H.; O’Shea, M. Neuronal expression of neural nitric oxide synthase (nNOS) protein is suppressed by an antisense RNA transcribed from an NOS pseudogene. J. Neurosci. 1999, 19, 7711–7720. [Google Scholar] [PubMed]
- Hawkins, P.G.; Morris, K.V. Transcriptional regulation of Oct4 by a long non-coding RNA antisense to Oct4-pseudogene 5. Transcription 2010, 1, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Tam, O.H.; Aravin, A.A.; Stein, P.; Girard, A.; Murchison, E.P.; Cheloufi, S.; Hodges, E.; Anger, M.; Sachidanandam, R.; Schultz, R.M.; et al. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 2008, 453, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Karreth, F.A.; Ala, U.; Provero, P.; Pandolfi, P.P. Pseudogenes as competitive endogenous RNAs: Target prediction and validation. Methods Mol. Biol. 2014, 1167, 199–212. [Google Scholar] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Chiefari, E.; Iiritano, S.; Paonessa, F.; le Pera, I.; Arcidiacono, B.; Filocamo, M.; Foti, D.; Liebhaber, S.A.; Brunetti, A. Pseudogene-mediated posttranscriptional silencing of HMGA1 can result in insulin resistance and type 2 diabetes. Nat. Commun. 2010, 1. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar] [CrossRef] [PubMed]
- Ala, U.; Karreth, F.A.; Bosia, C.; Pagnani, A.; Taulli, R.; Léopold, V.; Tay, Y.; Provero, P.; Zecchina, R.; Pandolfi, P.P. Integrated transcriptional and competitive endogenous RNA networks are cross-regulated in permissive molecular environments. Proc. Natl. Acad. Sci. USA 2013, 110, 7154–7159. [Google Scholar] [CrossRef] [PubMed]
- Vanin, E.F. Processed pseudogenes. Characteristics and evolution. Biochim. Biophys. Acta 1984, 782, 231–241. [Google Scholar] [CrossRef]
- Zhang, Z.; Harrison, P.M.; Liu, Y.; Gerstein, M. Millions of years of evolution preserved: A comprehensive catalog of the processed pseudogenes in the human genome. Genome Res. 2003, 13, 2541–2558. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Hattori, M.; Yada, T.; Gojobori, T.; Sakaki, Y.; Okada, N. Whole-genome screening indicates a possible burst of formation of processed pseudogenes and Alu repeats by particular L1 subfamilies in ancestral primates. Genome Biol. 2003, 4. [Google Scholar] [CrossRef] [PubMed]
- Esnault, C.; Maestre, J.; Heidmann, T. Human LINE retrotransposons generate processed pseudogenes. Nat. Genet. 2000, 24, 363–367. [Google Scholar] [PubMed]
- Pei, B.; Sisu, C.; Frankish, A.; Howald, C.; Habegger, L.; Mu, X.J.; Harte, R.; Balasubramanian, S.; Tanzer, A.; Diekhans, M.; et al. The GENCODE pseudogene resource. Genome Biol. 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Linial, M. Creation of a processed pseudogene by retroviral infection. Cell 1987, 49, 93–102. [Google Scholar] [CrossRef]
- Carlton, M.B.; Colledge, W.H.; Evans, M.J. Generation of a pseudogene during retroviral infection. Mamm. Genome 1995, 6, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Lazar, D.C.; Morris, K.V.; Saayman, S.M. The emerging role of long non-coding RNAs in HIV infection. Virus Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Adoro, S.; Cubillos-Ruiz, J.R.; Chen, X.; Deruaz, M.; Vrbanac, V.D.; Song, M.; Park, S.; Murooka, T.T.; Dudek, T.E.; Luster, A.D.; et al. IL-21 induces antiviral microRNA-29 in CD4 T cells to limit HIV-1 infection. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.L.; O’Brien, S.J.; Gilbert, D.A.; Reid, Y.; Popovic, M.; Read-Connole, E.; Gallo, R.C.; Gazdar, A.F. Origin of the HIV-susceptible human CD4+ cell line H9. AIDS Res. Hum. Retroviruses 1989, 5, 253–255. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Hendrickson, D.G.; Sauvageau, M.; Goff, L.; Rinn, J.L.; Pachter, L. Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat. Biotechnol. 2013, 31, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Deng, S.; Lu, H.; Zheng, Y.; Yang, G.; Kim, D.; Cao, Q.; Wu, J.Q. RNA-seq characterization of spinal cord injury transcriptome in acute/subacute phases: A resource for understanding the pathology at the systems level. PLoS ONE 2013, 8, e72567. [Google Scholar] [CrossRef] [PubMed]
- Marinov, G.K.; Williams, B.A.; McCue, K.; Schroth, G.P.; Gertz, J.; Myers, R.M.; Wold, B.J. From single-cell to cell-pool transcriptomes: Stochasticity in gene expression and RNA splicing. Genome Res. 2014, 24, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Flegel, C.; Manteniotis, S.; Osthold, S.; Hatt, H.; Gisselmann, G. Expression profile of ectopic olfactory receptors determined by deep sequencing. PLoS ONE 2013, 8, e55368. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Sova, P.; Green, R.R.; Thomas, M.J.; Korth, M.J.; Proll, S.; Xu, J.; Cheng, Y.; Yi, K.; Chen, L.; et al. Deep sequencing of HIV-infected cells: Insights into nascent transcription and host-directed therapy. J. Virol. 2014, 88, 8768–8782. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, A.; Cabau, C.; Bouchez, O.; Sarry, J.; Marsaud, N.; Foissac, S.; Woloszyn, F.; Mulsant, P.; Mandon-Pepin, B. An overview of gene expression dynamics during early ovarian folliculogenesis: Specificity of follicular compartments and bi-directional dialog. BMC Genomics 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gerstein, M. Large-scale analysis of pseudogenes in the human genome. Curr. Opin. Genet. Dev. 2004, 14, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Hodges, A.; Sharrocks, K.; Edelmann, M.; Baban, D.; Moris, A.; Schwartz, O.; Drakesmith, H.; Davies, K.; Kessler, B.; McMichael, A.; et al. Activation of the lectin DC-SIGN induces an immature dendritic cell phenotype triggering Rho-GTPase activity required for HIV-1 replication. Nat. Immunol. 2007, 8, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Del Real, G.; Jiménez-Baranda, S.; Mira, E.; Lacalle, R.A.; Lucas, P.; Gómez-Moutón, C.; Alegret, M.; Peña, J.M.; Rodríguez-Zapata, M.; Alvarez-Mon, M.; et al. Statins inhibit HIV-1 infection by down-regulating Rho activity. J. Exp. Med. 2004, 200, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Sklan, E.H.; Staschke, K.; Oakes, T.M.; Elazar, M.; Winters, M.; Aroeti, B.; Danieli, T.; Glenn, J.S. A Rab-GAP TBC domain protein binds hepatitis C virus NS5A and mediates viral replication. J. Virol. 2007, 81, 11096–11105. [Google Scholar] [CrossRef] [PubMed]
- Ferri, K.F.; Jacotot, E.; Blanco, J.; Esté, J.A.; Kroemer, G. Mitochondrial control of cell death induced by HIV-1-encoded proteins. Ann. N. Y. Acad. Sci. 2000, 926, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Bronte, V.; Zanovello, P. Regulation of immune responses by L-arginine metabolism. Nat. Rev. Immunol. 2005, 5, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.; Nasr-Esfahani, S.; Tan, C.H.; O’Brien, K.; Howard, J.L.; Jans, D.A.; Purcell, D.F.J.; Stoltzfus, C.M.; Sonza, S. HIV-1 infection induces changes in expression of cellular splicing factors that regulate alternative viral splicing and virus production in macrophages. Retrovirology 2008, 5. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, J.A.; Caputi, M. Role of cellular RNA processing factors in human immunodeficiency virus type 1 mRNA metabolism, replication, and infectivity. J. Virol. 2009, 83, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Ambrosini, G.; Adida, C.; Altieri, D.C. A novel anti-apoptosis gene, survivin, expressed in cancer and lymphoma. Nat. Med. 1997, 3, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Ozato, K.; Shin, D.M.; Chang, T.H.; Morse, H.C., 3rd. TRIM family proteins and their emerging roles in innate immunity. Nat. Rev. Immunol. 2008, 8, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Saikumar, P.; Reddy, C.D.; Reddy, E.P. Myb protein binds to human immunodeficiency virus 1 long terminal repeat (LTR) sequences and transactivates LTR-mediated transcription. Proc. Natl. Acad. Sci. USA 1990, 87, 8090–8094. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, B.M.; Murray, J.L.; Li, G.; Sheng, J.; Hodge, T.W.; Rubin, D.H.; O’Brien, W.A.; Ferguson, M.R. A functional role for ADAM10 in human immunodeficiency virus type-1 replication. Retrovirology 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Morou, A.K.; Porichis, F.; Krambovitis, E.; Sourvinos, G.; Spandidos, D.A.; Zafiropoulos, A. The HIV-1 gp120/V3 modifies the response of uninfected CD4 T cells to antigen presentation: Mapping of the specific transcriptional signature. J. Transl. Med. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; O’Day, E.; Wheeler, L.A.; Engelman, A.; Lieberman, J. HIV DNA is heavily uracilated, which protects it from autointegration. Proc. Natl. Acad. Sci. USA 2011, 108, 9244–9249. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Liu, B.; Wu, J.Q.; Soriano, V.; Vispo, E.; Carroll, A.P.; Goldie, B.J.; Cairns, M.J.; Saksena, N.K. Genome-wide mRNA and miRNA analysis of peripheral blood mononuclear cells (PBMC) reveals different miRNAs regulating HIV/HCV co-infection. Virology 2014, 450–451, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.P.; Koyuncu, O.O.; Enquist, L.W. Subversion of the actin cytoskeleton during viral infection. Nat. Rev. Microbiol. 2011, 9, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.; Finlay, B.; Scott, M.G.; Bowdish, D.; Rosenberger, C.; Powers, J.P.S.; Yu, J.; Mookherjee, N. Effectors of innate immunity. U.S. Patent Application 11/400,411, 2006. [Google Scholar]
- Li, S.; Wang, L.; Berman, M.; Kong, Y.Y.; Dorf, M.E. Mapping a dynamic innate immunity protein interaction network regulating type I interferon production. Immunity 2011, 35, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Fontenot, D.; He, H.; Hanabuchi, S.; Nehete, P.N.; Zhang, M.; Chang, M.; Nehete, B.; Wang, Y.H.; Wang, Y.H.; Ma, Z.M.; et al. TSLP production by epithelial cells exposed to immunodeficiency virus triggers DC-mediated mucosal infection of CD4+ T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 16776–16781. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.S.; Hopper-Borge, E.; Belinsky, M.G.; Shchaveleva, I.; Kotova, E.; Kruh, G.D. Characterization of the transport properties of human multidrug resistance protein 7 (MRP7, ABCC10). Mol. Pharmacol. 2003, 63, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Pu, H.; Andras, I.E.; Eum, S.Y.; Yamauchi, A.; Hennig, B.; Toborek, M. HIV-TAT protein upregulates expression of multidrug resistance protein 1 in the blood-brain barrier. J. Cereb. Blood Flow Metab. 2006, 26, 1052–1065. [Google Scholar] [CrossRef] [PubMed]
- Merino-Gracia, J.; García-Mayoral, M.F.; Rodríguez-Crespo, I. The association of viral proteins with host cell dynein components during virus infection. FEBS J. 2011, 278, 2997–3011. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Gojobori, T.; Nei, M. Pseudogenes as a paradigm of neutral evolution. Nature 1981, 292, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Podlaha, O.; Zhang, J. Nonneutral evolution of the transcribed pseudogene Makorin1-p1 in mice. Mol. Biol. Evol. 2004, 21, 2202–2209. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, J.; White, K.P. RNA-seq differential expression studies: More sequence or more replication? Bioinformatics 2014, 30, 301–304. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, A.; Brown, C.T.; Zheng, Y.-H.; Adami, C. Differentially-Expressed Pseudogenes in HIV-1 Infection. Viruses 2015, 7, 5191-5205. https://doi.org/10.3390/v7102869
Gupta A, Brown CT, Zheng Y-H, Adami C. Differentially-Expressed Pseudogenes in HIV-1 Infection. Viruses. 2015; 7(10):5191-5205. https://doi.org/10.3390/v7102869
Chicago/Turabian StyleGupta, Aditi, C. Titus Brown, Yong-Hui Zheng, and Christoph Adami. 2015. "Differentially-Expressed Pseudogenes in HIV-1 Infection" Viruses 7, no. 10: 5191-5205. https://doi.org/10.3390/v7102869