Generation of Recombinant Rabies Virus CVS-11 Expressing eGFP Applied to the Rapid Virus Neutralization Test

{kind=link}
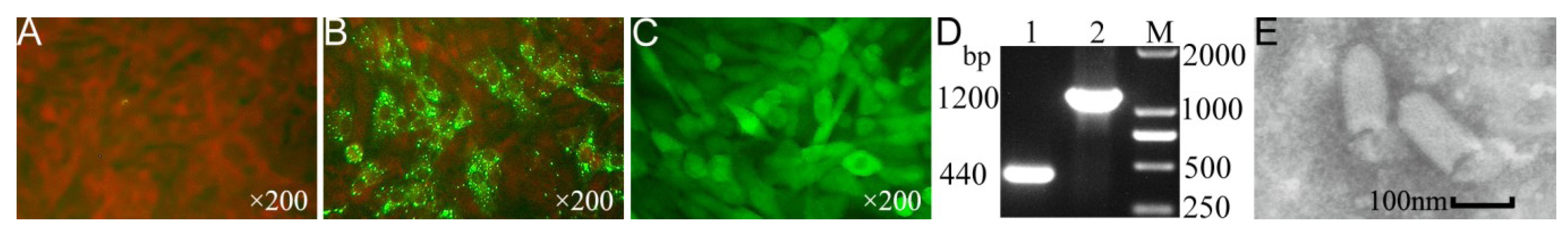{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Viruses and Cells
2.2. Serum Samples
2.3. Construction of the Recombinant rCVS-11-eGFP Strain
2.4. Recovery of the Recombinant rCVS-11-eGFP Strain from Cloned cDNA
2.5. Confirmation of the rCVS-11-eGFP Strain
2.6. Virus Multistep Growth Assay
2.7. Pathogenicity of rCVS-11-eGFP in Mice
2.8. Fluorescent Antibody Virus Neutralization Tests (FAVN)
2.9. The FAVN-eGFP Method
2.10. Data Analyses
3. Results
3.1. Recovery of the rCVS-11-eGFP Strain
3.2. Growth Properties of rCVS-11-eGFP
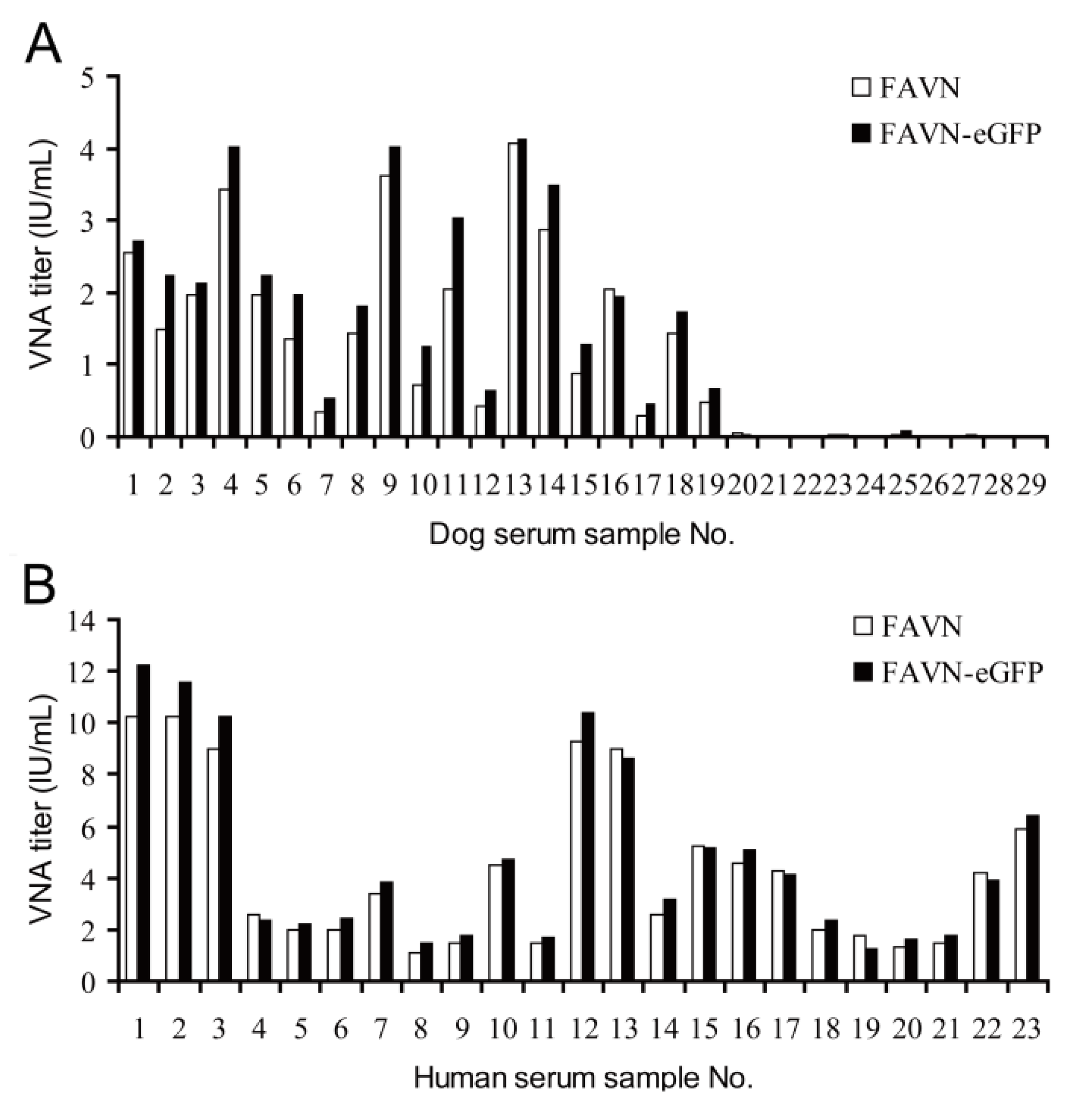
3.3. Comparative Analysis of VNA Titers Determined by Standard FAVN and FAVN-eGFP Methods
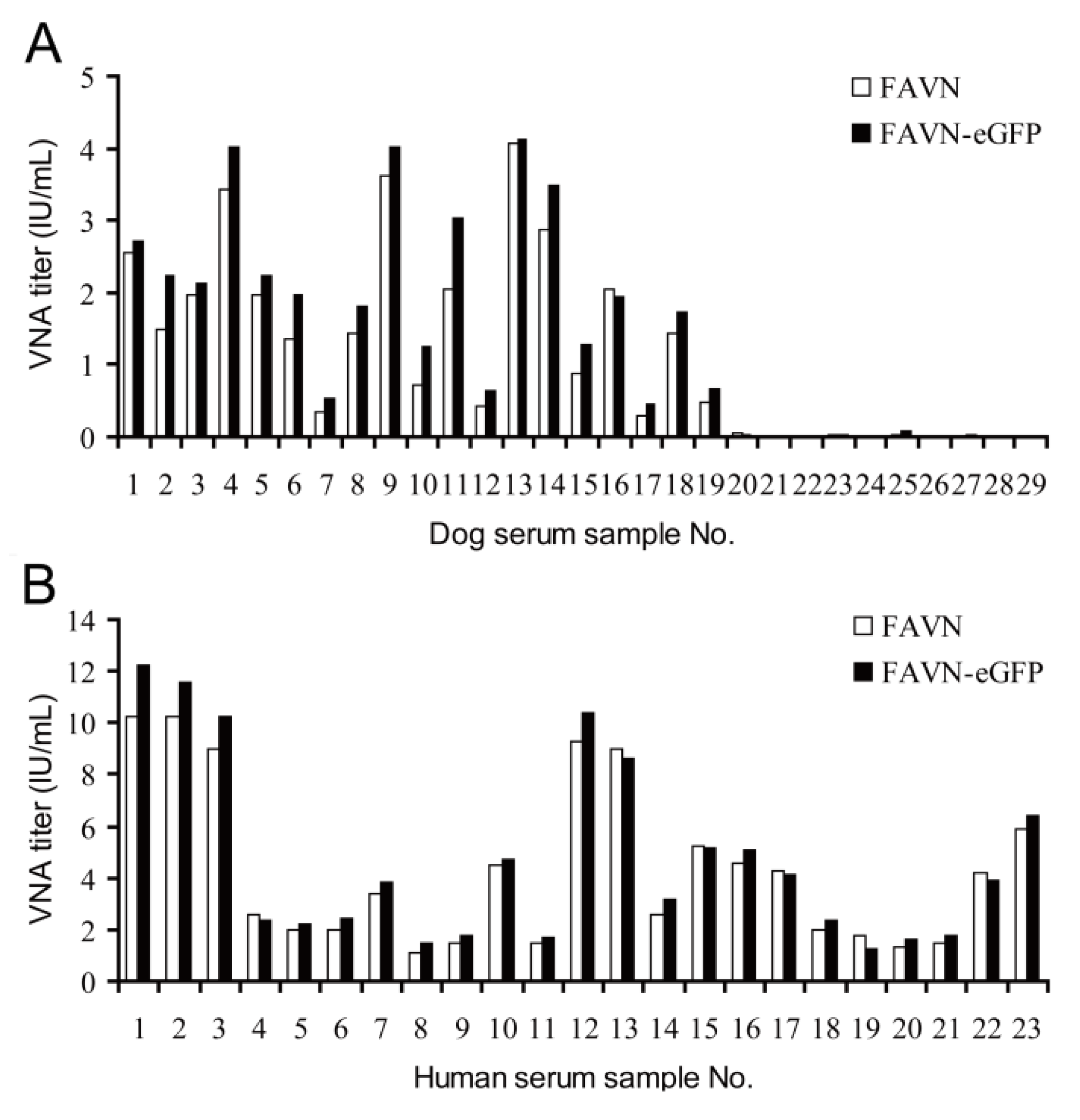
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Dietzschold, B.; Schnell, M.; Koprowski, H. Pathogenesis of rabies. Curr. Top. Microbiol. Immunol. 2005, 292, 45–56. [Google Scholar]
- Zhao, P.; Zhao, L.; Zhang, K.; Feng, H.; Wang, H.; Wang, T.; Xu, T.; Feng, N.; Wang, C.; Gao, Y.; et al. Infection with street strain rabies virus induces modulation of the microRNA profile of the mouse brain. Virol. J. 2012, 9, 159–172. [Google Scholar]
- World Health Organization (WHO). The World Survey for Rabies No. 35 Geneva Document WHO/CDS/CSR/EPH/2002.10; WHO: Geneva, Switzerland, 1999. [Google Scholar]
- Hu, R.; Tang, Q.; Tang, J.; Fooks, A.R. Rabies in China: An update. Vector-Borne Zoonotic Dis. 2009, 9, 1–12. [Google Scholar] [CrossRef]
- Cleaveland, S.; Kaare, M.; Knobel, D.; Laurenson, M.K. Canine vaccination—Providing broader benefits for disease control. Vet. Microbiol. 2006, 117, 43–50. [Google Scholar] [CrossRef]
- Kitala, P.M.; McDermott, J.J.; Coleman, P.G.; Dye, C. Comparison of vaccination strategies for the control of dog rabies in Machakos District, Kenya. Epidemiol. Infect. 2002, 129, 215–222. [Google Scholar]
- Hampson, K.; Dushoff, J.; Cleaveland, S.; Haydon, D.T.; Kaare, M.; Packer, C.; Dobson, A. Transmission dynamics and prospects for the elimination of Canine Rabies. PLoS Biol. 2009, 7, 462–471. [Google Scholar]
- Coleman, P.G.; Dye, C. Immunization coverage required to prevent outbreaks of dog rabies. Vaccine 1996, 14, 185–186. [Google Scholar] [CrossRef]
- Li, J.; Ertel, A.; Portocarrero, C.; Barkhouse, D.A.; Dietzschold, B.; Hooper, D.C. Faber M.Postexposure treatment with the live-attenuated rabies virus (RV) vaccine TriGAS triggers the clearance of wild-type RV from the Central Nervous System (CNS) through the rapid induction of genes relevant to adaptive immunity in CNS tissues. J. Virol. 2012, 86, 3200–3210. [Google Scholar] [CrossRef]
- Liao, P.H.; Yang, H.H.; Chou, P.T.; Wang, M.H.; Chu, P.C.; Liu, H.L.; Chen, L.K. Sufficient virus-neutralizing antibody in the central nerve system improves the survival of rabid rats. J. Biomed. Sci. 2012, 19, 61–71. [Google Scholar] [CrossRef]
- Ondrejkova, A.; Suli, J.; Ondrejka, R.; Benisek, Z.; Franka, R.; Svrcek, S. Comparison of the detection and quantification of rabies antibodies in canine sera. Vet. Med. 2002, 47, 218–221. [Google Scholar]
- Briggs, D.J.; Smith, J.S.; Mueller, F.L.; Schwenke, J.; Davis, R.D.; Gordon, C.R.; Schweitzer, K.; Orciari, L.A.; Yager, P.A.; Rupprecht, C.E. A Comparison of two serological methods for detecting the immune response after rabies vaccination in dogs and cats being exported to rabies-free areas. Biologicals 1998, 26, 347–355. [Google Scholar] [CrossRef]
- Yu, P.; Lv, X.; Shen, X.; Tang, Q.; Liang, G. Establishment and preliminary application of a rapid fluorescent focus inhibition test (RFFIT) for rabies virus. Virol. Sin. 2013, 28, 223–227. [Google Scholar] [CrossRef]
- Wu, X.; Rupprecht, C.E. Glycoprotein gene relocation in rabies virus. Virus Res. 2008, 131, 95–99. [Google Scholar] [CrossRef]
- Wu, X.; Rupprecht, C.E. Rescue of recombinant peste des petits ruminants virus: Creation of a GFP-expressing virus and application in rapid virus neutralization test. Vet. Res. 2012, 43, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Tang, Q.; Nadin-Davis, S.A.; Zhang, S.; Hooper, C.D.; Ming, P.; Du, J.; Tao, X.; Hu, R.; Liang, G. Development of a reverse genetics system for a human rabies virus vaccine strain employed in China. Virus Res. 2010, 149, 28–35. [Google Scholar]
- Van den Pol, A.N.; Dalton, K.P.; Rose, J.K. Relative neurotropism of a recombinant rhabdovirus expressing a green fluorescent envelope glycoprotein. J. Virol. 2002, 76, 1309–1327. [Google Scholar] [CrossRef]
- Finke, S.; Brzozka, K.; Conzelmann, K.K. Tracking fluorescence-labeled rabies virus: Enhanced green fluorescent protein-tagged phosphoprotein P supports virus gene expression and formation of infectious particles. J. Virol. 2004, 78, 12333–12343. [Google Scholar] [CrossRef]
- Xue, X.; Zheng, X.; Gai, W.; Liang, H.; Ma, J.; Li, L.; Wang, T.; Feng, N.; Huang, G.; Zhao, Y.; et al. Sequencing the complete genome of rabies virus CVS-11 strain and constructing its full-length infectious cDNA clone. Acta Microbiol. Sin. 2013, 4, 409–415. [Google Scholar]
- Inoue, K.; Shoji, Y.; Kurane, I.; Iijima, T.; Sakai, T.; Morimoto, K. An improved method for recovering rabies virus from cloned cDNA. J. Virol. Methods 2003, 107, 229–236. [Google Scholar] [CrossRef]
- Cliquet F, A.M.; Sagne, L. Development of a fluorescent antibody virus neutralisation test (FAVN test) for the quantification of rabies neutralising antibody. J. Immunol. Methods 1998, 212, 79–87. [Google Scholar] [CrossRef]
- Fabian Käsermann, C.K. Photodynamic inactivation of enveloped viruses by buckminsterfullerene. Antivir. Res. 1997, 34, 65–70. [Google Scholar]
- Statistical Program for Social Sciences (SPSS), version 13.0; SPSS Inc.: Chicago, IL, USA, 2005.
- Khawplod, P.; Inoue, K.; Shoji, Y.; Wilde, H.; Ubol, S.; Nishizono, A.; Kurane, I.; Morimoto, K. A novel rapid fluorescent focus inhibition test for rabies virus using a recombinant rabies virus visualizing a green fluorescent protein. J. Virol. Methods 2005, 125, 35–40. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xue, X.; Zheng, X.; Liang, H.; Feng, N.; Zhao, Y.; Gao, Y.; Wang, H.; Yang, S.; Xia, X. Generation of Recombinant Rabies Virus CVS-11 Expressing eGFP Applied to the Rapid Virus Neutralization Test. Viruses 2014, 6, 1578-1589. https://doi.org/10.3390/v6041578
Xue X, Zheng X, Liang H, Feng N, Zhao Y, Gao Y, Wang H, Yang S, Xia X. Generation of Recombinant Rabies Virus CVS-11 Expressing eGFP Applied to the Rapid Virus Neutralization Test. Viruses. 2014; 6(4):1578-1589. https://doi.org/10.3390/v6041578
Chicago/Turabian StyleXue, Xianghong, Xuexing Zheng, Hongru Liang, Na Feng, Yongkun Zhao, Yuwei Gao, Hualei Wang, Songtao Yang, and Xianzhu Xia. 2014. "Generation of Recombinant Rabies Virus CVS-11 Expressing eGFP Applied to the Rapid Virus Neutralization Test" Viruses 6, no. 4: 1578-1589. https://doi.org/10.3390/v6041578