Movements of Wild Ruddy Shelducks in the Central Asian Flyway and Their Spatial Relationship to Outbreaks of Highly Pathogenic Avian Influenza H5N1

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Capture, Marking, and Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PTT | Sex | Age | Mass (g) | Capture Date | Last Transmission | Working Days | GPS Locations | Argos Locations | Total Locations |
|---|---|---|---|---|---|---|---|---|---|
| 73042 | F | A | 1,135 | 9/12/2007 | 1/15/2008 | 126 | 314 | 84 | 398 |
| 74808 | F | J | 1,130 | 9/13/2007 | 3/21/2009 | 556 | 3274 | 762 | 4036 |
| 74809 | F | J | 970 | 9/14/2007 | 2/18/2008 | 158 | 800 | 148 | 948 |
| 74810 | M | A | 1,390 | 9/13/2007 | Active | 978 | 2946 | 913 | 3859 |
| 74812 | M | J | 1,510 | 9/13/2007 | 2/1/2008 | 142 | 137 | 16 | 153 |
| 74813 | M | J | 1,350 | 9/14/2007 | 12/10/2008 | 454 | 2207 | 412 | 2619 |
| 74816 | M | A | 1,600 | 9/16/2007 | 12/22/2008 | 464 | 1125 | 317 | 1442 |
| 74817 | M | A | 1,640 | 3/26/2007 | 1/3/2008 | 284 | 1299 | 157 | 1456 |
| 74818 | M | A | 1,270 | 9/13/2007 | 2/15/2008 | 156 | 772 | 92 | 864 |
| 74820 | F | A | 1,185 | 9/13/2007 | 4/21/2008 | 222 | 1675 | 308 | 1983 |
| 74822 | F | A | 1,070 | 9/14/2007 | 2/10/2009 | 516 | 1003 | 167 | 1170 |
| 74823 | F | J | 1,010 | 9/14/2007 | 11/16/2007 | 64 | 159 | 37 | 196 |
| 82095 | F | A | 1,295 | 3/28/2008 | Active | 782 | 3618 | 477 | 4095 |
| 82097 | F | U | 1,515 | 4/3/2008 | 6/6/2009 | 430 | 2919 | 756 | 3675 |
| 82099 | M | A | 1,580 | 4/3/2008 | Active | 776 | 4529 | 1048 | 5577 |
| 82100 | M | A | 1,470 | 4/3/2008 | Active | 776 | 2932 | 656 | 3588 |
| 82116 | F | J | 1,275 | 9/11/2008 | 1/14/2009 | 126 | 970 | 226 | 1196 |
| 82117 | A | F | 1,300 | 9/11/2008 | Active | 616 | 2530 | 518 | 3048 |
| 82118 | M | A | 1,495 | 9/15/2008 | 11/3/2008 | 50 | 264 | 43 | 307 |
| 82119 | M | A | 1,505 | 9/14/2008 | 7/12/2009 | 302 | 748 | 172 | 920 |
| 82120 | M | J | 1,160 | 9/13/2008 | 11/1/2008 | 50 | 361 | 80 | 441 |
| 82121 | F | A | 1,270 | 9/13/2008 | 6/9/2009 | 270 | 2351 | 506 | 2857 |
| 82122 | M | A | 1,725 | 9/13/2008 | Active | 612 | 3352 | 932 | 4284 |
| 82123 | M | J | 1,395 | 9/13/2008 | 11/21/2008 | 70 | 461 | 64 | 525 |
| 82126 | M | J | 1,155 | 9/12/2008 | Active | 614 | 4876 | 1244 | 6120 |
| 82127 | F | A | 1,210 | 9/13/2008 | 4/16/2010 | 581 | 4476 | 910 | 5386 |
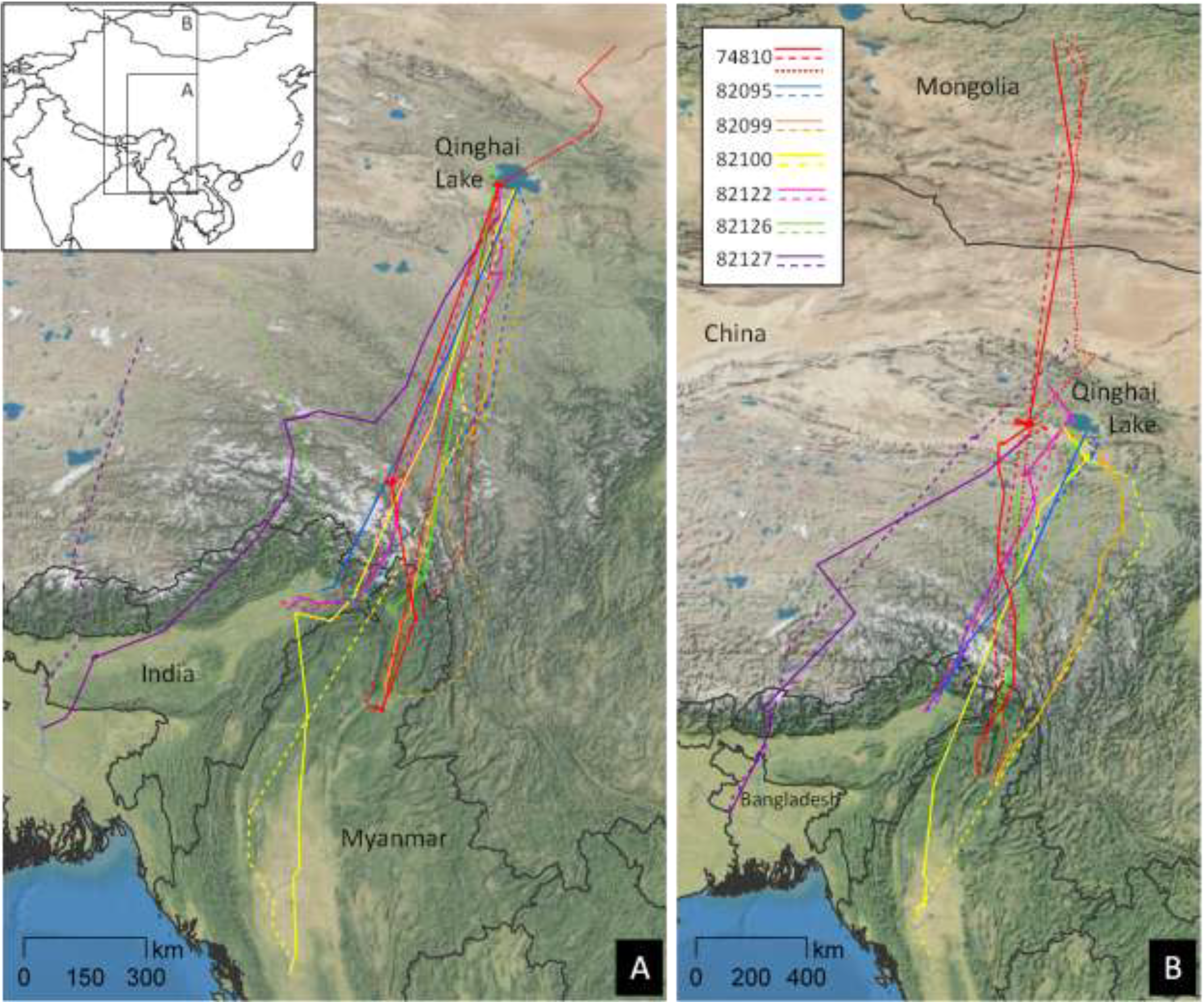
2.2. Migratory Movements
2.2.1. Major Use Areas in the Central Asian Flyway
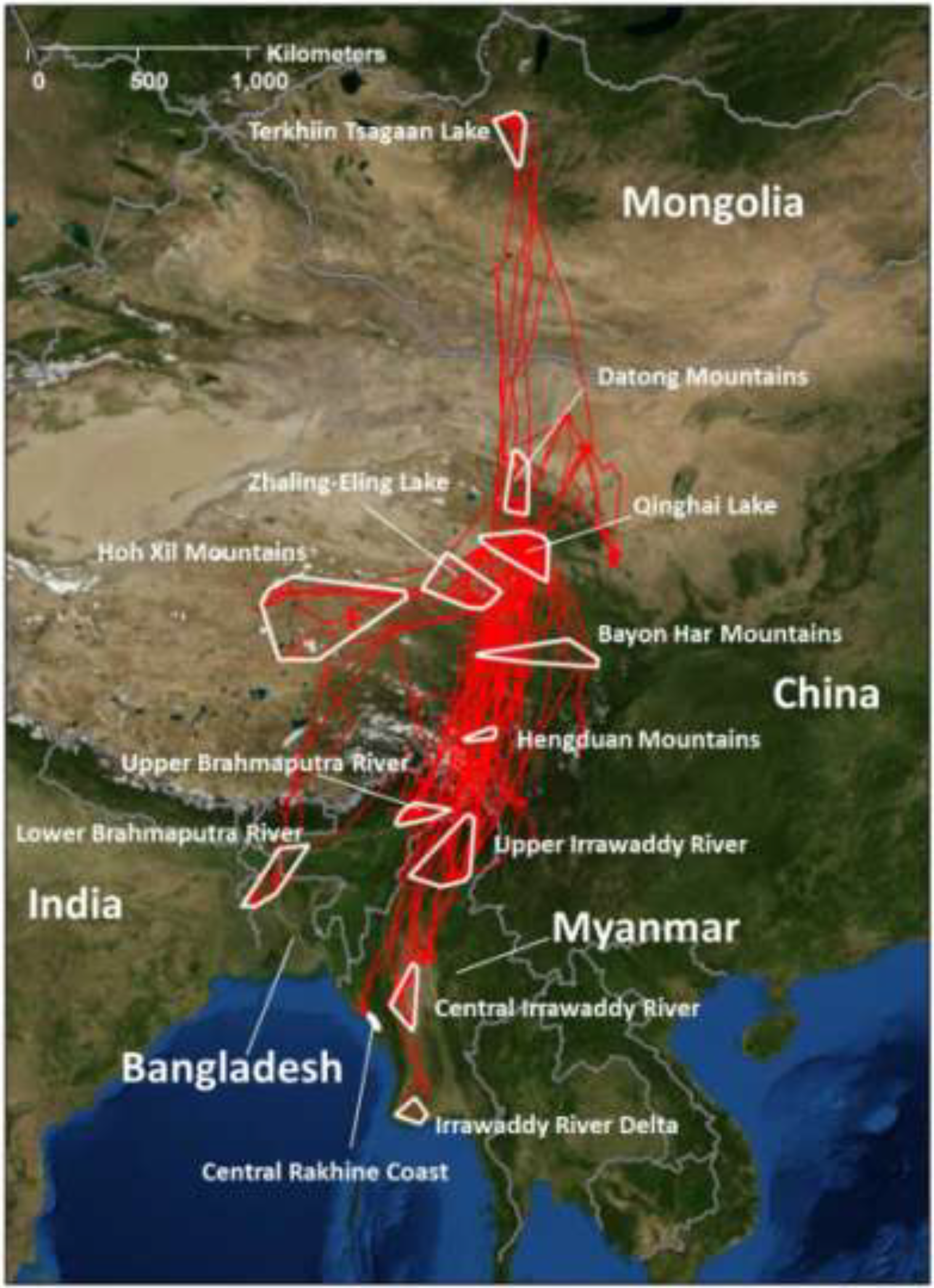
2.2.2. Migration Chronology and Movement Rates

| Site Name | Distance and Direction from Qinghai Lake (km) | Coordinates (Degrees Latitude, Longitude) | Time Period | N | Date Range | Mean Length of Stay (range in days) | Total Locations |
|---|---|---|---|---|---|---|---|
| Terkhiin Tsagaan Lake Region, Arkhangai Province, Mongolia | 1240 N | 48.09 N, 99.92 E | B 08-09 | 2 | 4/22–9/18 | 115 (98–149) | 1615 |
| Hoh Xil Mountain Region, PR China | 570 SW | 34.67 N, 91.60 E | B 08-09 | 3 | 7/8–10/26 | 95 (71–107) | 1952 |
| Datong Mountains Region, PR China | 220 N | 38.85 N, 99.44 E | B 08-09 | 4 | 5/5–7/27 | 59 (37–79) | 864 |
| Qinghai Lake Region, PR China | 0-140 W, S | 36.72 N, 99.02 E | B 07-09 | 10 | 4/2–12/13 | 174 (77–255) | 5360 |
| S 08-10 | 7 | 3/8–4/25 | 13 (2–48) | 259 | |||
| F 07, 09 | 2 | 10/23–12/6 | 13 (8–18) | 17 | |||
| Bayon Har Mountains Region, PR China | 420 S | 33.10 N, 100.52 E | S 08-09 | 4 | 3/12–4/29 | 23 (12–42) | 576 |
| Zhaling-Eling Lake Region, PR China | 300 SW | 35.42 N, 97.12 E | B 07-09 | 3 | 7/3–10/26 | 45 (28–110) | 1147 |
| S 08-10 | 3 | 3/18–5/2 | 16 (4–22) | 49 | |||
| Hengduan Mountains Region, PR China | 750 SW | 30.28 N, 97.97 E | F 07-08 | 3 | 11/3–12/4 | 9 (2–16) | 132 |
| Upper Brahmaputra River, India | 1140 SW | 27.50 N, 95.03 E | W 07-09 | 3 | 11/2–4/15 | 102 (71–164) | 4086 |
| Lower Brahmaputra River and Ganges River, Bangladesh/India Border | 1630 SW | 25.05 N, 89.51 E | W 08-09 | 1 | 11/13–3/14 | 100 (78–121) | 2446 |
| Upper Irrawaddy River Region, Myanmar | 1220 SW | 26.18 N, 96.81 E | W 07-09 | 5 | 10/27–3/20 | 100 (71–144) | 6154 |
| Central Irrawaddy River Region, Myanmar | 1850 SW | 20.73 N, 94.92 E | F 08 | 1 | 11/3–11/29 | 26 | 358 |
| W 08-09 | 2 | 11/15–3/9 | 84 (78–96) | 2524 | |||
| Irrawaddy River Delta, Myanmar | 1990 SW | 19.75 N, 93.62 E | W 08 | 1 | 12/21–3/8 | 77 | 829 |
| Central Rakhine Coast, Myanmar | 2320 SW | 16.23 N, 95.18 E | W 07 | 1 | 12/21–3/9 | 78 | 828 |
| ID | Capture Date | Fall Migration | Spring Migration | Longest Leg | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depart Breeding | Arrive Wintering | Duration (d) | Stop-overs (N) | Depart Wintering | Arrive Breeding | Duration (d) | Stop-overs (N) | Leg Distance (km) | Time (d) | Rate (km/d) | Total Displacement (km) | ||
| 73042 | 9/12/2007 | 10/31/2007 | 11/2/2007 | 3 | 0 | na | na | na | na | 831.0 | 1.4 | 594 | 724 |
| 74808 | 9/13/2007 | 11/23/2007 | 11/24/2007 | 2 | 0 | 3/12/2008 | 5/2/2008 | 52 | 1 | 1,422.0 | 2.3 | 610 | 2,686 |
| 11/14/2008 | 11/18/2008 | 5 | 0 | 3/20/2009 | na | na | na | 1,331.4 | 4.1 | 326 | |||
| 74809 | 9/14/2007 | 11/24/2007 | 11/26/2007 | 3 | 0 | na | na | na | na | 1,068.9 | 1.8 | 583 | 1,095 |
| 74810 | 9/13/2007 | 11/9/2007 | 12/7/2007 | 29 | 1 | 3/12/2008 | 5/2/2008 | 52 | 1 | 2,431.0 | 2.8 | 859 | 3,461 |
| 11/7/2008 | 11/16/2008 | 10 | 0 | 3/19/2009 | 4/22/2009 | 35 | 2 | 2,240.7 | 8.8 | 256 | |||
| 10/18/2009 | 11/14/2009 | 28 | 1 | 3/4/2010 | 4/28/2010 | 56 | 1 | 1,763.8 | 1.6 | 1,085 | |||
| 74813 | 9/14/2007 | 11/1/2007 | 11/2/2007 | 2 | 0 | 4/7/2008 | 5/4/2008 | 28 | 1 | 1,047.2 | 2.2 | 483 | 1,474 |
| 11/1/2008 | 11/2/2008 | 2 | 0 | na | na | na | na | 888.7 | 0.8 | 1185 | |||
| 74816 | 9/16/2007 | 11/8/2007 | 11/20/2007 | 13 | 1 | 3/15/2008 | 4/9/2008 | 26 | 2 | 1,833.3 | 2.3 | 801 | 1,754 |
| 11/13/2008 | 11/24/2008 | 12 | 1 | na | na | na | na | 770.9 | 2.7 | 289 | |||
| 74817 | 3/26/2007 | 10/28/2007 | 11/1/2007 | 5 | 0 | na | na | na | na | 2,009.6 | 3.5 | 574 | 1,633 |
| 74818 | 9/13/2007 | 10/27/2007 | 10/30/2007 | 4 | 0 | na | na | na | na | 1,141.6 | 3.0 | 381 | 1,205 |
| 74820 | 9/13/2007 | 11/12/2007 | 11/19/2007 | 8 | 1 | 3/28/2008 | na | na | na | 1,881.7 | 2.3 | 808 | 2,372 |
| 74822 | 9/14/2007 | 11/26/2007 | 12/20/2007 | 25 | 1 | 3/17/2008 | 5/1/2008 | 46 | 1 | 979.3 | 6.5 | 151 | 1,144 |
| 82095 | 3/28/2008 | 11/15/2008 | 11/16/2008 | 2 | 0 | 4/8/2009 | 4/10/2009 | 3 | 0 | 1,071.8 | 1.0 | 1,072 | 1,362 |
| 11/14/2009 | 11/19/2009 | 6 | 0 | 3/30/2010 | 4/14/2010 | 16 | 1 | 1,742.5 | 5.0 | 349 | |||
| 82097 | 4/3/2008 | 11/14/2008 | 11/17/2008 | 4 | 0 | 3/22/2009 | 3/25/2009 | 4 | 0 | 2,481.5 | 3.1 | 802 | 2,178 |
| 82099 | 4/3/2008 | 11/29/2008 | 12/10/2008 | 12 | 1 | 3/10/2009 | 4/2/2009 | 24 | 1 | 1,612.8 | 2.5 | 645 | 1,616 |
| 12/14/2009 | 12/16/2009 | 3 | 0 | 2/25/2010 | 2/28/2010 | 4 | 0 | 1,940.1 | 2.2 | 895 | |||
| 82100 | 4/3/2008 | 11/6/2008 | 11/9/2008 | 4 | 0 | 3/25/2009 | 4/11/2009 | 18 | 1 | 2,947.5 | 7.0 | 419 | 2,271 |
| 11/12/2009 | 11/15/2009 | 4 | 0 | 2/19/2010 | 3/28/2010 | 38 | 2 | 2,033.6 | 3.2 | 642 | |||
| 82116 | 9/11/2008 | 11/3/2008 | 11/4/2008 | 2 | 0 | na | na | na | na | 1,288.5 | 1.5 | 859 | 1,347 |
| 82119 | 9/14/2008 | 11/14/2008 | 11/16/2008 | 3 | 0 | 4/8/2009 | 4/12/2009 | 5 | 0 | 2,008.8 | 4.6 | 441 | 1,233 |
| 82121 | 9/13/2008 | 11/1/2008 | 12/1/2008 | 31 | 1 | 3/24/2009 | 4/13/2009 | 21 | 1 | 2,050.1 | 3.9 | 523 | 2,655 |
| 82122 | 9/13/2008 | 11/1/2008 | 11/12/2008 | 12 | 2 | 3/23/2009 | 4/21/2009 | 30 | 2 | 1,759.3 | 2.4 | 746 | 1,546 |
| 10/30/2009 | 11/2/2009 | 4 | 0 | 4/15/2010 | 4/17/2010 | 3 | 0 | 1,647.0 | 1.4 | 1,145 | |||
| 82126 | 9/12/2008 | 11/6/2008 | 11/7/2008 | 2 | 0 | 4/24/2009 | 4/25/2009 | 2 | 0 | 1,112.6 | 1.3 | 890 | 1,415 |
| 10/25/2009 | 10/27/2009 | 3 | 0 | 3/20/2010 | 4/6/2010 | 18 | 1 | 1,089.9 | 3.0 | 363 | |||
| 82127 | 9/13/2008 | 11/14/2008 | 12/8/2008 | 25 | 1 | 4/6/2009 | 4/9/2009 | 4 | 0 | 2,620.3 | 4.1 | 637 | 2,381 |
| 11/9/2009 | 11/13/2009 | 5 | 0 | 3/14/2010 | 4/1/2010 | 19 | 2 | 934.7 | 3.5 | 264 | |||
2.2.3. Space Use and Distribution
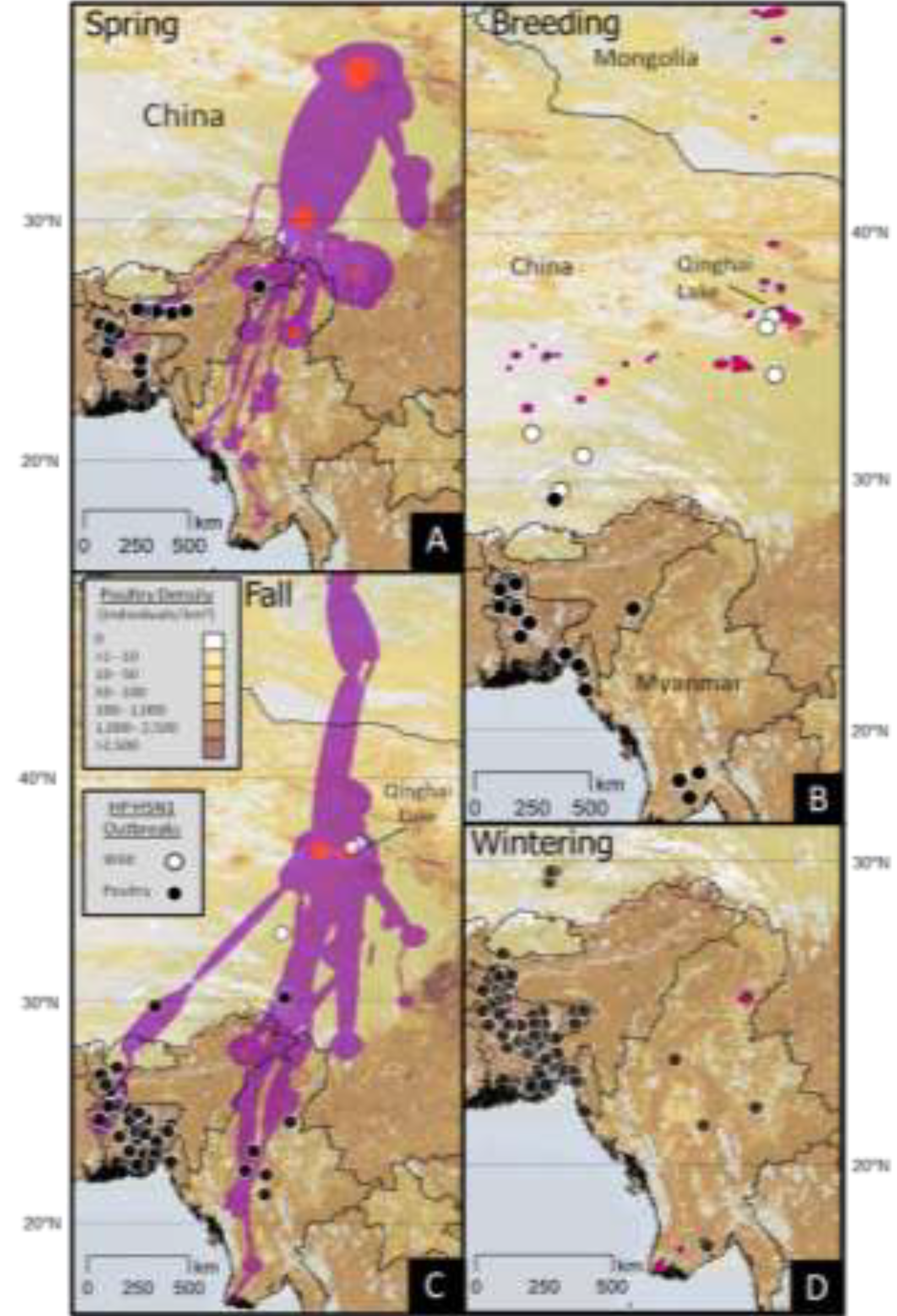

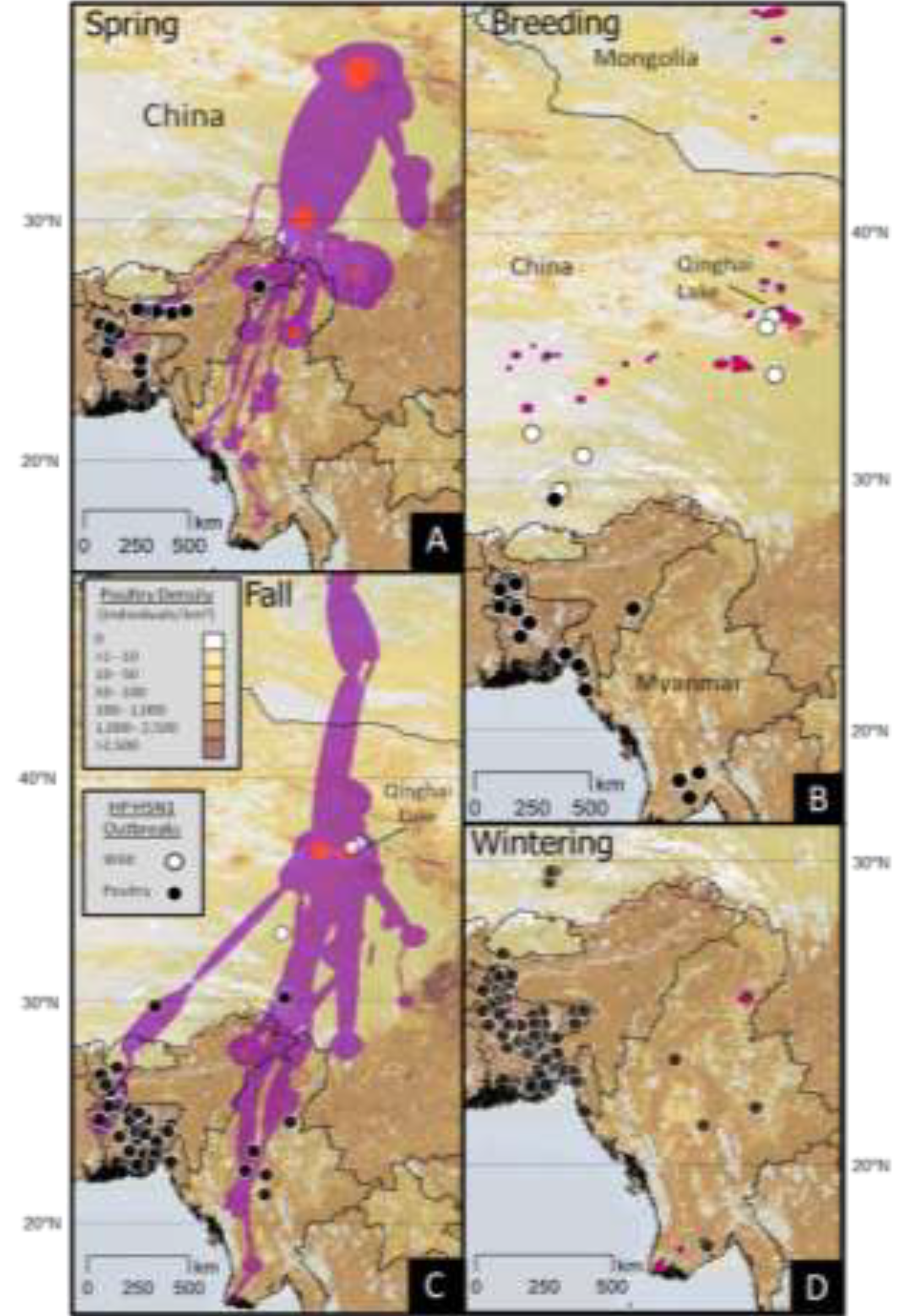
2.3. HPAI H5N1 Outbreaks
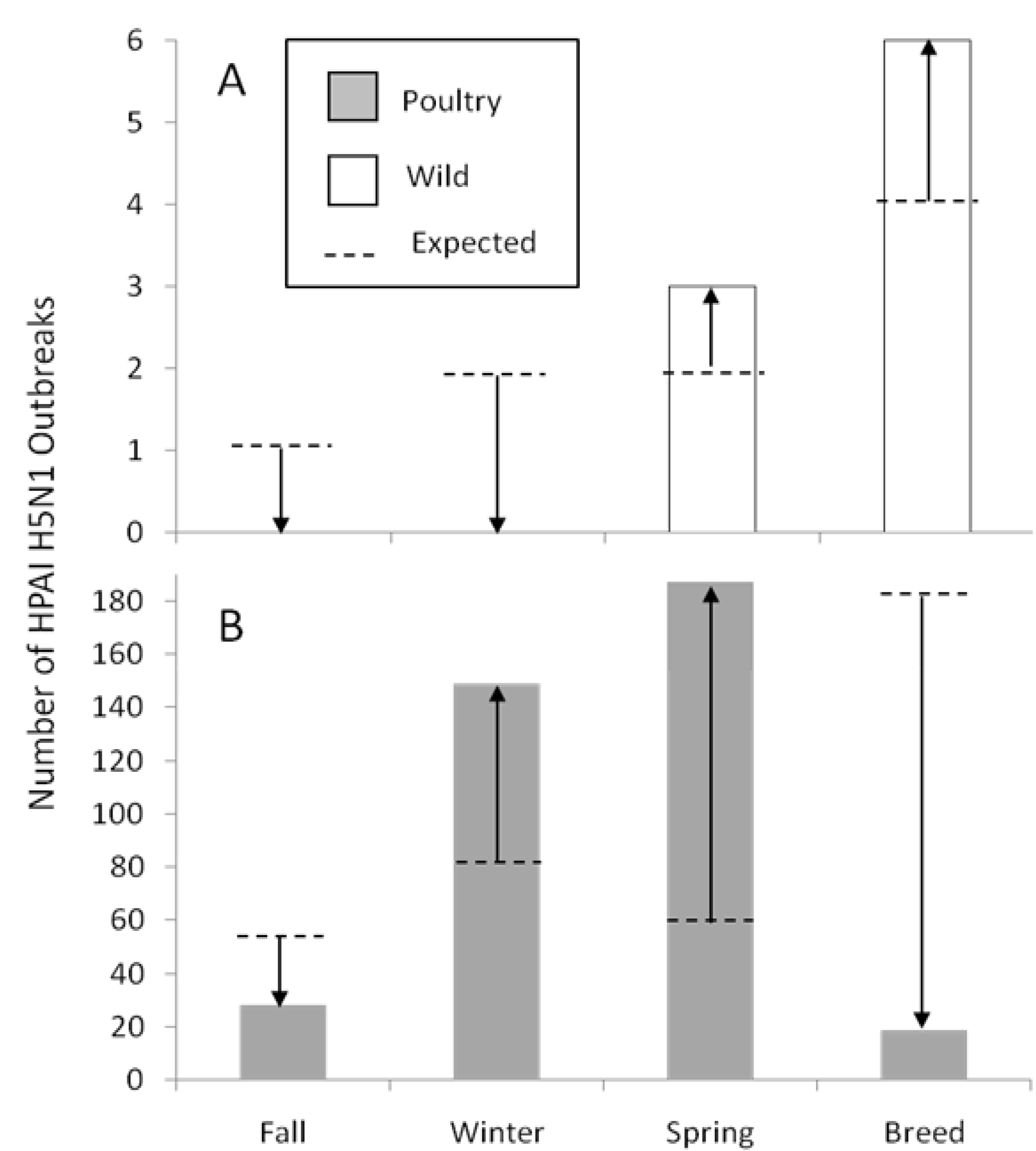
2.3.1. Relation to Ecological Parameters
| Outbreak Type | Model | N | K | −2 Log- likelihood | AICc | ΔAICc | Akaike Weight (%) |
|---|---|---|---|---|---|---|---|
| Domestic | Wetl, Crop, Lat, Dev, Pdens | 4213 | 6 | 1,757.8 | 1,769.8 | 0 | 45.6 |
| Wetl, Crop, Lat, Dev, Pdens, Grass, RUSHUD | 4213 | 8 | 1,757.1 | 1,773.1 | 3.3 | 8.9 | |
| Null Model (intercept only) | 4213 | 0 | 2,566.9 | 2,566.9 | 797.0 | 0.0 | |
| Wild | Lat, Dev | 99 | 3 | 33.5 | 39.5 | 0 | 49.6 |
| Wetl, Crop, Lat, Dev, Pdens, Grass, RUSHUD | 99 | 8 | 30.2 | 46.2 | 8.1 | 0.9 | |
| Null Model (intercept only) | 99 | 0 | 60.3 | 60.3 | 20.6 | 0.0 |
| Model | Parameter | Β | SE | Wald χ2 | Pr > χ2 |
|---|---|---|---|---|---|
| Poultry Outbreaks | Intercept | −0.2415 | 0.6307 | 0.1466 | 0.7018 |
| Poultry Density | 0.0001 | 0.0001 | 2.7242 | 0.0988 | |
| Wetland | 2.1 | 0.5557 | 14.2805 | 0.0002 | |
| Cropland | 3.1258 | 0.1953 | 256.1308 | <0.0001 | |
| Latitude | −0.1637 | 0.0261 | 39.4072 | <0.0001 | |
| Urban | 15.7376 | 1.3640 | 133.1154 | <0.0001 | |
| Wild Outbreaks | Intercept | −25.2017 | 7.3469 | 11.7665 | 0.0006 |
| Latitude | 0.7044 | 0.2158 | 10.6519 | 0.0011 | |
| Urban | 21.0892 | 13.0902 | 2.5955 | 0.1072 |
2.3.2. Correlation and Regression Tree

3. Experimental Section
3.1. Study Area
3.2. Capture, Virology Sampling, and Marking
3.3. Telemetry and Movements
3.4. H5N1 Outbreaks and Risk Factors
3.5. Statistical Analyses
3.6. Outbreak Characteristics
4. Discussion and Conclusions
Acknowledgments
Conflicts of Interest
References and Notes
- Liu, J.; Xiao, H.; Lei, F.; Zhu, Q.; Qin, K.; Zhang, X.W.; Zhang, X.; Zhao, D.; Wang, G.; Feng, Y.; et al. Highly pathogenic H5N1 influenza virus infection in migratory birds. Science 2005, 309, 1206–1206. [Google Scholar] [CrossRef]
- Chen, H.; Smith, G.J.D.; Zhang, S.Y.; Qin, K.; Wang, J.; Li, K.S.; Webster, R.G.; Peiris, J.S.M.; Guan, Y. Avian flu: H5N1 virus outbreak in migratory waterfowl. Nature 2005, 436, 191–192. [Google Scholar] [CrossRef]
- Stallknecht, D.; Shane, S. Host range of avian influenza virus in free-living birds. Vet. Res. Commun. 1988, 12, 125–141. [Google Scholar] [CrossRef]
- Takekawa, J.Y.; Prosser, D.J.; Newman, S.H.; Muzaffar, S.B.; Hill, N.J.; Yan, B.; Xiao, X.; Lei, F.; Li, T.; Schwarzbach, S.E.; et al. Victims and vectors: Highly pathogenic avian influenza H5N1 and the ecology of wild birds. Avian Biol. Res. 2010, 3, 51–73. [Google Scholar] [CrossRef]
- Fergus, R.; Fry, M.; Karesh, W.B.; Marra, P.P.; Newman, S.; Paul, E. Migratory Birds and Avian Flu. Science 2006, 312, 845–846. [Google Scholar]
- Normile, D. Evidence points to migratory birds in H5N1 spread. Science 2006, 311, 1225. [Google Scholar] [CrossRef]
- Gaidet, N.; Cattoli, G.; Hammoumi, S.; Newman, S.H.; Hagemeijer, W.; Takekawa, J.Y.; Cappelle, J.; Dodman, T.; Joannis, T.; Gil, P.; et al. Evidence of infection by H5N2 highly pathogenic avian influenza viruses in healthy wild waterfowl. PLoS Pathog. 2008, 4, e1000127. [Google Scholar] [CrossRef]
- Newman, S.H.; Hill, N.J.; Spragens, K.A.; Janies, D.; Voronkin, I.O.; Prosser, D.J.; Yan, B.; Lei, F.; Batbayar, N.; Natsagdorj, T.; et al. Eco-virological approach for assessing the role of wild birds in the spread of avian influenza H5N1 along the Central Asian Flyway. PLoS One 2012, 7, e30636. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Zhang, Y.; Duan, Z.; Tian, G.; Zeng, X.; Shi, J.; Zhang, L.; Chen, H. New Avian Influenza Virus (H5N1) in Wild Birds, Qinghai, China. Emerg. Infect. Dis. 2011, 17, 265–267. [Google Scholar] [CrossRef]
- Food and Agricultural Organization of the United Nations, FAO Food Outlook: Global Market Analysis; Food and Agricultural Organization of the United Nations: Rome, Italy, 2012.
- Chen, H. H5N1 avian influenza in China. Sci. China C Life Sci. Chinese Acad. of Sci. 2009, 52, 419–427. [Google Scholar] [CrossRef]
- Chen, H.; Li, Y.; Li, Z.; Shi, J.; Shinya, K.; Deng, G.; Qi, Q.; Tian, G.; Fan, S.; Zhao, H.; Sun, Y.; Kawaoka, Y. Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in western China. J. Virol. 2006, 80, 5976–5983. [Google Scholar] [CrossRef]
- Namgail, T.; Takekawa, J.Y.; Sivananinthaperumal, B.; Areendran, G.; Sathiyaselvam, P.; Mundkur, T.; McCracken, T.; Newman, S. Ruddy Shelduck Tadorna ferruginea home range and habitat use during the non-breeding season in Assam, India. Wildfowl 2011, 61, 182–193. [Google Scholar]
- Ali, S.; Ripley, S.D. Handbook of the Birds of India and Pakistan: Together with Those of Bangladesh, Nepal, Sikkim, Bhutan and Sri Lanka; Oxford University Press: Delhi, India, 1978; p. 247. [Google Scholar]
- Kumar, A.; Sati, J.P.; Tak, P.C.; Alfred, J.R.B. Handbook on Indian Wetland Birds and Their Conservation; Shiva Printing Press: Dehradun, India, 2005. [Google Scholar]
- BirdLife International. Species factsheet: Tadorna ferruginea. Available online: http://www.birdlife.org/datazone/speciesfactsheet.php?id=397 (accessed on 17 June 2010).
- Popovkina, A.B. Conflicting trends in Ruddy Shelduck Tadorna ferruginea populations: A myth or reality? In Waterbirds Around the World; Boere, G.C., Gailbraith, C.A., Stroud, D.A., Eds.; The Stationery Office: Edinburgh, UK, 2006; pp. 480–481. [Google Scholar]
- Xu, H.; Hou, Z.H.; Ai, L.; Tan, L.C. Precipitation at Lake Qinghai, NE Qinghai-Tibet Plateau, and its relation to Asian summer monsoons on decadal/interdecadal scales during the past 500 years. Palaeogeogr. Palaeocl. 2007, 254, 541–549. [Google Scholar] [CrossRef]
- Rhode, D.; Haizhou, M.; Madsen, D.B.; Brantingham, P.J.; Forman, S.L.; Olsen, J.W. Paleoenvironmental and archaeological investigations at Qinghai Lake, western China: Geomorphic and chronometric evidence of lake level history. Quatern. Int. 2010, 218, 29–44. [Google Scholar] [CrossRef]
- Ramsar Convention Secretariat, Designating Ramsar Sites: The Strategic Framework and Guidelines for the Future Development of the List of Wetlands of International Importance. Ramsar Handbooks for the Wise Use of Wetlands, 3rd ed.Ramsar Convention Secretariat: Gland, Switzerland, 2007.
- Li, L. Survey results and new records of waterfowl at Qinghai Lake. China Crane News 2001, 5, 36–37. [Google Scholar]
- L’vov, D.; Shchelkanov, M.I.; Deriabin, P.; Grebennikova, T.; Prilipov, A.; Nepoklonov, E.; Onishchenko, G.; Vlasov, N.; Aliper, T.; Zaberezhnyĭ, A.; et al. Isolation of influenza A/H5N1 virus strains from poultry and wild birds in West Siberia during epizooty (July 2005) and their depositing to the state collection of viruses (August 8, 2005). Vop. Virusol. 2006, 51, 11–14. [Google Scholar]
- Dzubin, A.; Cooch, E.G. Measurements of Geese: General Field Methods; California Waterfowl Association: Sacramento, CA, USA, 1992.
- FAO Animal Production and Health Manual; Whitworth, D.; Newman, S.H.; Mundkur, T.; Harris, P. (Eds.) FAO: Rome, Italy, 2007.
- Fouchier, R.A.; Bestebroer, T.M.; Herfst, S.; Van Der Kemp, L.; Rimmelzwaan, G.F.; Osterhaus, A.D. Detection of influenza A viruses from different species by PCR amplification of conserved sequences in the matrix gene. J. Clin. Microbiol. 2000, 38, 4096–4101. [Google Scholar]
- OIE Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organization of Animal Health: Paris, France, 2004; pp. 465–481.
- Kou, Z.; Li, Y.; Yin, Z.; Guo, S.; Wang, M.; Gao, X.; Li, P.; Tang, L.; Jiang, P.; Luo, Z.; et al. The survey of H5N1 flu virus in wild birds in 14 provinces of China from 2004 to 2007. PLoS One 2009, 4, e6926. [Google Scholar]
- Douglas, D.C.; Weinzierl, R.; Davidson, S.C.; Kays, R.; Wikelski, M.; Bohrer, G. Moderating Argos location errors in animal tracking data. Meth. Ecol. Evol. 2012, 3, 999–1007. [Google Scholar]
- Douglas, D.C. Douglas Filter Version 7.03. Available online: http://alaska.usgs.gov/science/biology/spatial (accessed on 16 June 2010).
- Beyer, H.L. Hawth’s Analysis Tools for ArcGIS. Available online: http://www.spatialecology.com/htools/tooldesc.php (accessed on 22 June 2010).
- MOA Ministry of Agriculture of the People’s Republic of China. Prevention and Control of Avian Influenza. Available online: http://www.agri.gov.cn/ztzl/fkqlg/yqfb/ (accessed on 16 January 2010).
- Prosser, D.J.; Cui, P.; Takekawa, J.Y.; Tang, M.; Hou, Y.; Collins, B.M.; Yan, B.; Hill, N.J.; Li, T.; Li, Y.; et al. Wild bird migration across the Qinghai-Tibetan plateau: A transmission route for highly pathogenic H5N1. PLoS One 2011, 6, e17622. [Google Scholar]
- FAO. Geonetwork. Available online: Available online: www.fao.org/geonetwork (accessed on 30 May 2010).
- Wint, W.; Robinson, T. Gridded Livestock of the World; FAO: Rome, Italy, 2007; p. 131. [Google Scholar]
- Takekawa, J.Y.; Newman, S.H.; Xiao, X.; Prosser, D.J.; Spragens, K.A; Palm, E.C.; Yan, B.; Li, T.; Lei, F.; Zhao, D.; et al. Migration of waterfowl in the East Asian flyway and spatial relationship to HPAI H5N1 outbreaks. Avian Dis. 2010, 54, 466–476. [Google Scholar] [CrossRef]
- U.S. Geological Survey. Moderate-resolution Imaging Spectroradiometer (MODIS)/Terra Land Cover Classification. Available online: http://lpdaac.usgs.gov/ (accessed on 11 June 2010).
- Horne, J.; Garton, E.; Krone, S.; Lewis, J. Analyzing animal movements using Brownian bridges. Ecology 2007, 88, 2354–2363. [Google Scholar] [CrossRef]
- Horne, J.S.; Garton, E.O. Animal Space Use 1.3. College of Natural Resources; University of Idaho: Moscow, Idaho, USA, 2009. [Google Scholar]
- Horne, J.S.; Garton, E. Likelihood cross-validation versus least squares cross-validation for choosing the smoothing parameter in Kernel Home-Range analysis. J. Wildlife Manage. 2006, 70, 641–648. [Google Scholar] [CrossRef]
- Agresti, A. A Survey of exact inference for contingency tables. Stat. Sci. 1992, 7, 131–153. [Google Scholar] [CrossRef]
- Fisher, R.A. Statistical Methods for Research Workers; Oliver and Boyd: Edinburgh, UK, 1954. [Google Scholar]
- Manly, B.F.J.; McDonald, L.L.; Thomas, D.L.; McDonald, T.L.; Erickson, W.P. Resource Selection by Animals: Statistical Design and Analysis for Field Studies; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model selection and multimodel inference. In Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Burnham, K.P., Anderson, D.R., Eds.; Springer: New York, NY, USA, 2002; pp. 49–79. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer-Verlag: New York, NY, USA, 1998; pp. 1–515. [Google Scholar]
- Anderson, D.R.; Burnham, K.P.; Thompson, W.L. Null hypothesis testing: Problems, prevalence, and an alternative. J. Wildlife Manage. 2000, 64, 912–923. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Chapman & Hall: New York, NY, USA, 1983. [Google Scholar]
- Therneau, T.M.; Atkinson, E.J. An Introduction to Recursive Partitioning Using the RPART Routines; Technical Report 61; Section of Biostatistics, Mayo Clinic: Rochester, MN, USA, 1997; pp. 1–52. [Google Scholar]
- R: A Language and Environment for Statistical Computing; R Development Core Team: Vienna, Austria, 2010.
- Si, Y.; Skidmore, A.K.; Wang, T.; de Boer, W.F.; Debba, P.; Toxopeus, A.G.; Li, L.; Prins, H.H.T. Spatio-temporal dynamics of global H5N1 outbreaks match bird migration patterns. Geospatial Health 2009, 4, 65–78. [Google Scholar]
- Chen, H.; Deng, G.; Li, Z.; Tian, G.; Li, Y.; Jiao, P.; Zhang, L.; Liu, Z.; Webster, R.; Yu, K. The evolution of H5N1 influenza viruses in ducks in southern China. Proc. Natl. Acad. Sci. USA 2004, 101, 10452–10457. [Google Scholar]
- Kwon, Y.; Thomas, C.; Swayne, D. Variability in pathobiology of South Korean H5N1 high-pathogenicity avian influenza virus infection for 5 species of migratory waterfowl. Vet. Pathol. 2010, 47, 495–506. [Google Scholar] [CrossRef]
- Biswas, P.K.; Christensen, J.P.; Ahmed, S.S.U.; Barua, H.; Das, A.; Rahman, M.H.; Giasuddin, M.; Hannan, A.S.M.A.; Habib, M.A.; Ahad, A.; et al. Avian influenza outbreaks in chickens, Bangladesh. Emerg. Infect. Dis. 2008, 14, 1909–1912. [Google Scholar] [CrossRef]
- Gilbert, M.; Sokha, C.; Joyner, P.H.; Thomson, R.L.; Poole, C. Characterizing the trade of wild birds for merit release in Phnom Penh, Cambodia and associated risks to health and ecology. Biol. Conserv. 2012, 153, 10–16. [Google Scholar] [CrossRef]
- Quan, R.C.; Xianji, W.; Yang, X.; Peng, G.H.; Huang, T.F. Habitat use by wintering ruddy shelduck at Lashihai Lake. Waterbird Soc. 2001, 24, 402–406. [Google Scholar]
- Hill, N.J.; Takekawa, J.Y.; Cardona, C.J.; Meixell, B.W.; Ackerman, J.T.; Runstadler, J.A.; Boyce, W.M. Cross-seasonal patterns of avian influenza virus in breeding and wintering migratory birds: A flyway perspective. Vector-Borne Zoonot. 2012, 12, 243–253. [Google Scholar] [CrossRef]
- Takekawa, J.Y.; Heath, S.R.; Douglas, D.C.; Perry, W.M.; Javed, S.; Newman, S.H.; Suwal, R.N.; Rahmani, A.R.; Choudhury, B.C.; Prosser, D.J.; et al. Geographic variation in bar-headed geese Anser Indicus: Connectivity of wintering areas and breeding grounds across a broad front. Wildfowl 2010, 59, 100–123. [Google Scholar]
- Warnock, N. Stopping vs. staging: The difference between a hop and a jump. J. Avian Biol. 2010, 41, 621–626. [Google Scholar] [CrossRef]
- Wang, X.D.; Zhong, X.H.; Liu, S.Z.; Liu, J.G.; Wang, Z.Y.; Li, M.H. Regional assessment of environmental vulnerability in the Tibetan Plateau: Development and application of a new method. J. Arid Environ. 2008, 72, 1929–1939. [Google Scholar] [CrossRef]
- Keating, K.A.; Cherry, S. Modeling utilization distributions in space and time. Ecology 2009, 90, 1971–1980. [Google Scholar] [CrossRef]
- Lebarbenchon, C.; Albespy, F.; Brochet, A.-L.; Grandhomme, V.; Renaud, F.; Fritz, H.; Green, A.J.; Thomas, F.; van der Werf, S.; Aubry, P.; et al. Spread of avian influenza viruses by common teal (Anas crecca) in Europe. PLoS One 2009, 4, e7289. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.D.; Stallknecht, D.E.; Swayne, D.E. Experimental infection of swans and geese with highly pathogenic avian influenza virus (H5N1) of Asian lineage. Emerg. Infect. Dis. 2008, 14, 136–142. [Google Scholar] [CrossRef]
- Hill, N.J.; Takekawa, J.Y.; Cardona, C.J.; Ackerman, J.T.; Schultz, A.K.; Spragens, K.A.; Boyce, W.M. Waterfowl ecology and avian influenza in California: Do host traits inform us about viral occurrence? Avian Dis. 2010, 54, 426–432. [Google Scholar] [CrossRef]
- Kear, J. Ducks, Geese and Swans; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Takekawa, J.Y.; Western Ecological Research Center, U.S. Geological Survey, Niadao, Qinghai Province, China. Personal observation, 2010.
- Muzaffar, S.B.; Takekawa, J.Y.; Prosser, D.J.; Douglas, D.C.; Yan, B.; Xing, Z.; Hou, Y.; Palm, E.C.; Newman, S.H. Seasonal movements and migration of Pallas’s Gulls Larus ichthyaetus from Qinghai Lake, China. Forktail 2008, 24, 100–107. [Google Scholar]
- Yamaguchi, N.; Hupp, J.W.; Higuchi, H.; Flint, P.L.; Pearce, J.M. Satellite-tracking of Northern Pintail Anas acuta during outbreaks of the H5N1 virus in Japan: Implications for virus spread. Ibis 2010, 152, 262–271. [Google Scholar] [CrossRef]
- Melville, D.S.; Shortridge, K.F. Spread of H5N1 avian influenza virus: An ecological conundrum. 2006, 42, 435–437. [Google Scholar]
- Yasué, M.A.Ï.; Feare, C.J.; Bennun, L.; Fiedler, W. The epidemiology of H5N1 avian influenza in wild birds: Why we need better ecological data. BioScience 2006, 56, 923–929. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Takekawa, J.Y.; Prosser, D.J.; Collins, B.M.; Douglas, D.C.; Perry, W.M.; Yan, B.; Ze, L.; Hou, Y.; Lei, F.; Li, T.; et al. Movements of Wild Ruddy Shelducks in the Central Asian Flyway and Their Spatial Relationship to Outbreaks of Highly Pathogenic Avian Influenza H5N1. Viruses 2013, 5, 2129-2152. https://doi.org/10.3390/v5092129
Takekawa JY, Prosser DJ, Collins BM, Douglas DC, Perry WM, Yan B, Ze L, Hou Y, Lei F, Li T, et al. Movements of Wild Ruddy Shelducks in the Central Asian Flyway and Their Spatial Relationship to Outbreaks of Highly Pathogenic Avian Influenza H5N1. Viruses. 2013; 5(9):2129-2152. https://doi.org/10.3390/v5092129
Chicago/Turabian StyleTakekawa, John Y., Diann J. Prosser, Bridget M. Collins, David C. Douglas, William M. Perry, Baoping Yan, Luo Ze, Yuansheng Hou, Fumin Lei, Tianxian Li, and et al. 2013. "Movements of Wild Ruddy Shelducks in the Central Asian Flyway and Their Spatial Relationship to Outbreaks of Highly Pathogenic Avian Influenza H5N1" Viruses 5, no. 9: 2129-2152. https://doi.org/10.3390/v5092129