C7a, a Biphosphinic Cyclopalladated Compound, Efficiently Controls the Development of a Patient-Derived Xenograft Model of Adult T Cell Leukemia/Lymphoma

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
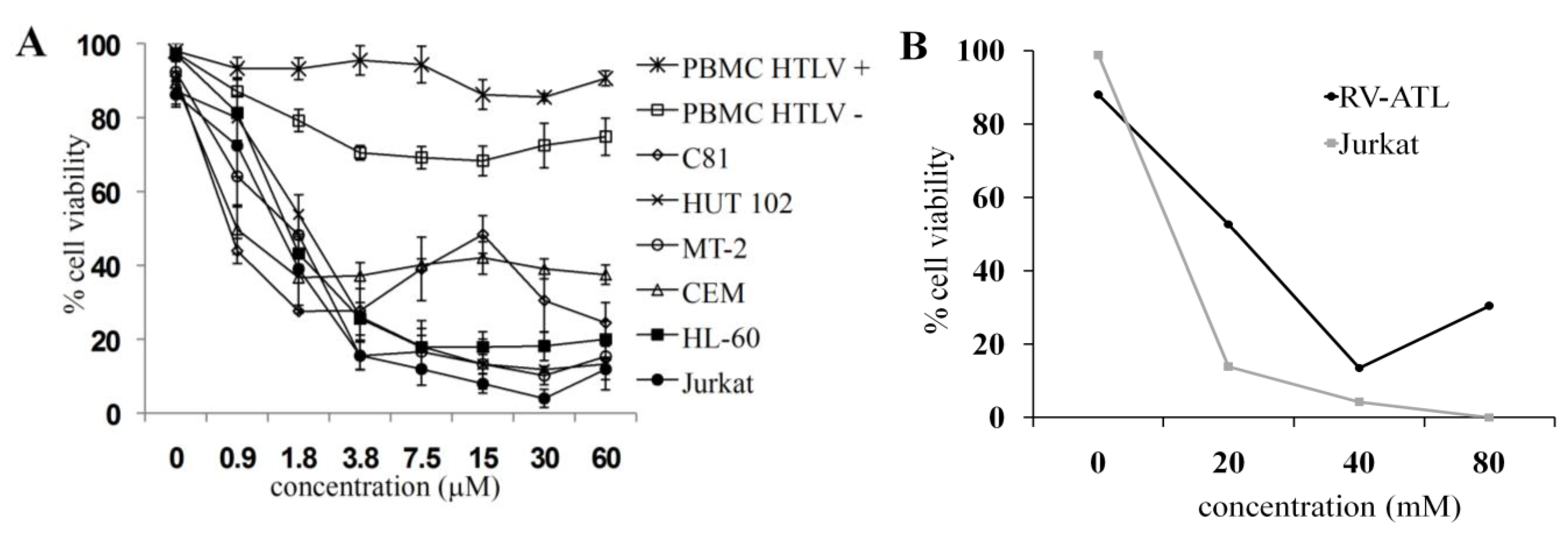
2.1. C7a Is Cytotoxic to Human Leukemia T Cell Lines
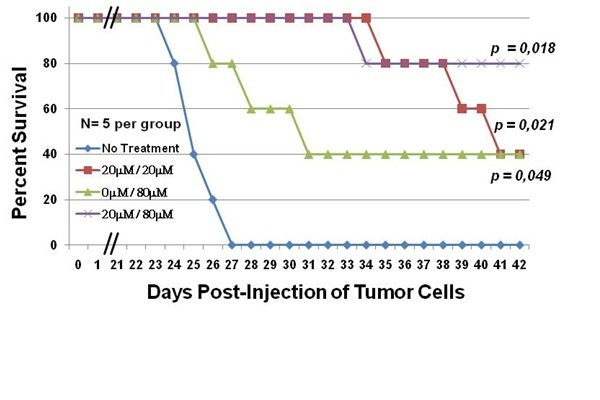
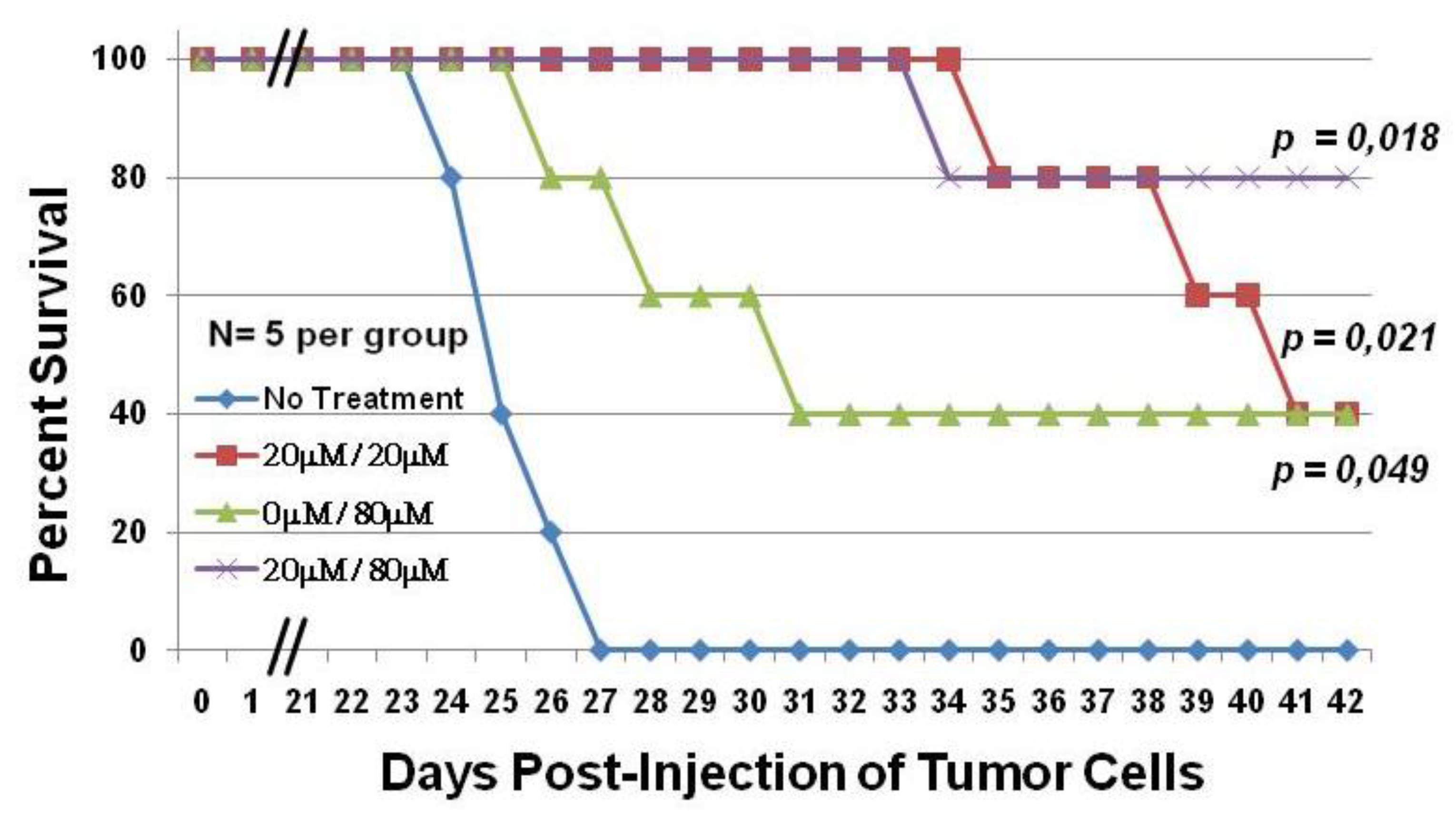
2.2. C7a Is Effective in a Preclinical Model of ATLL
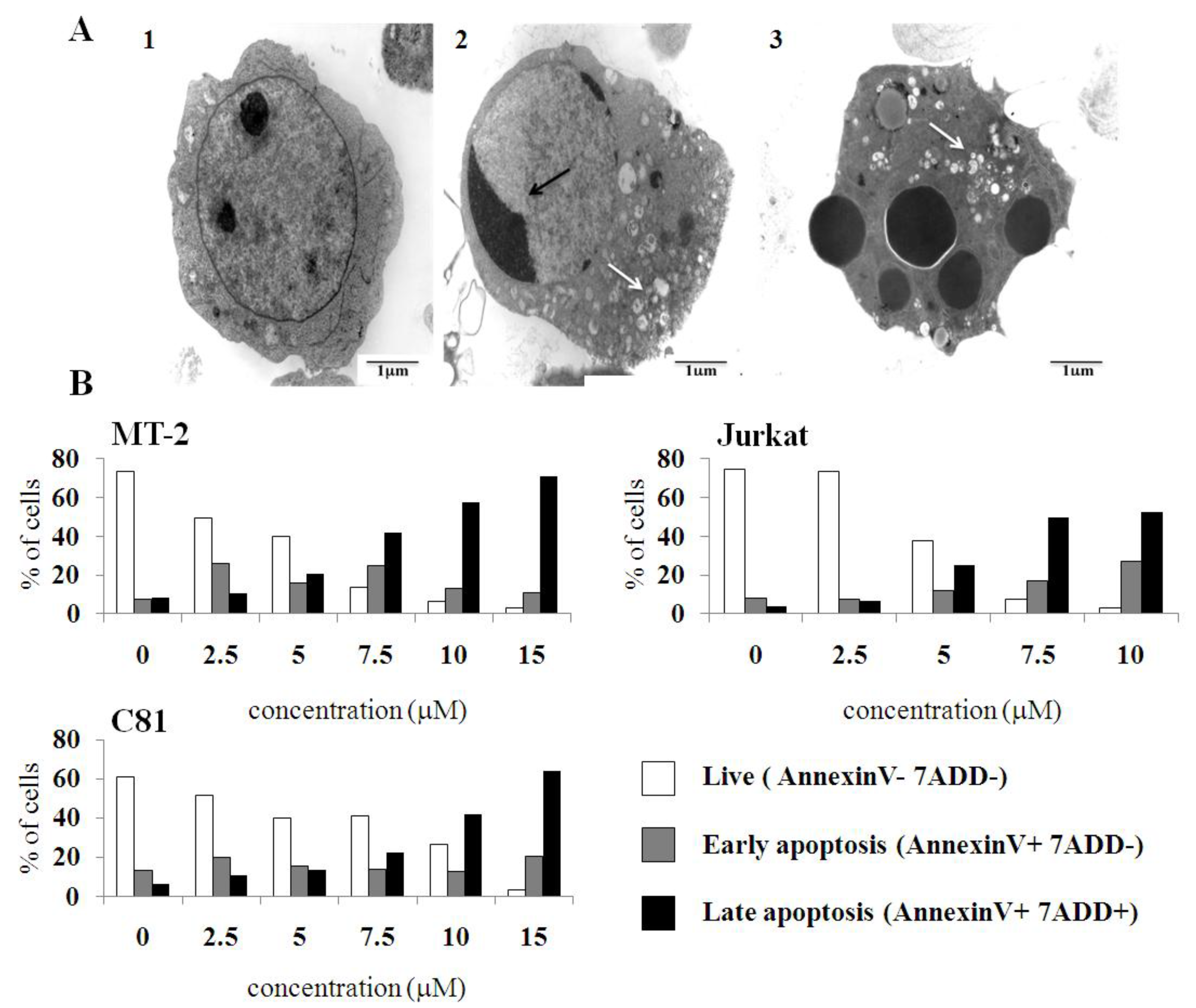
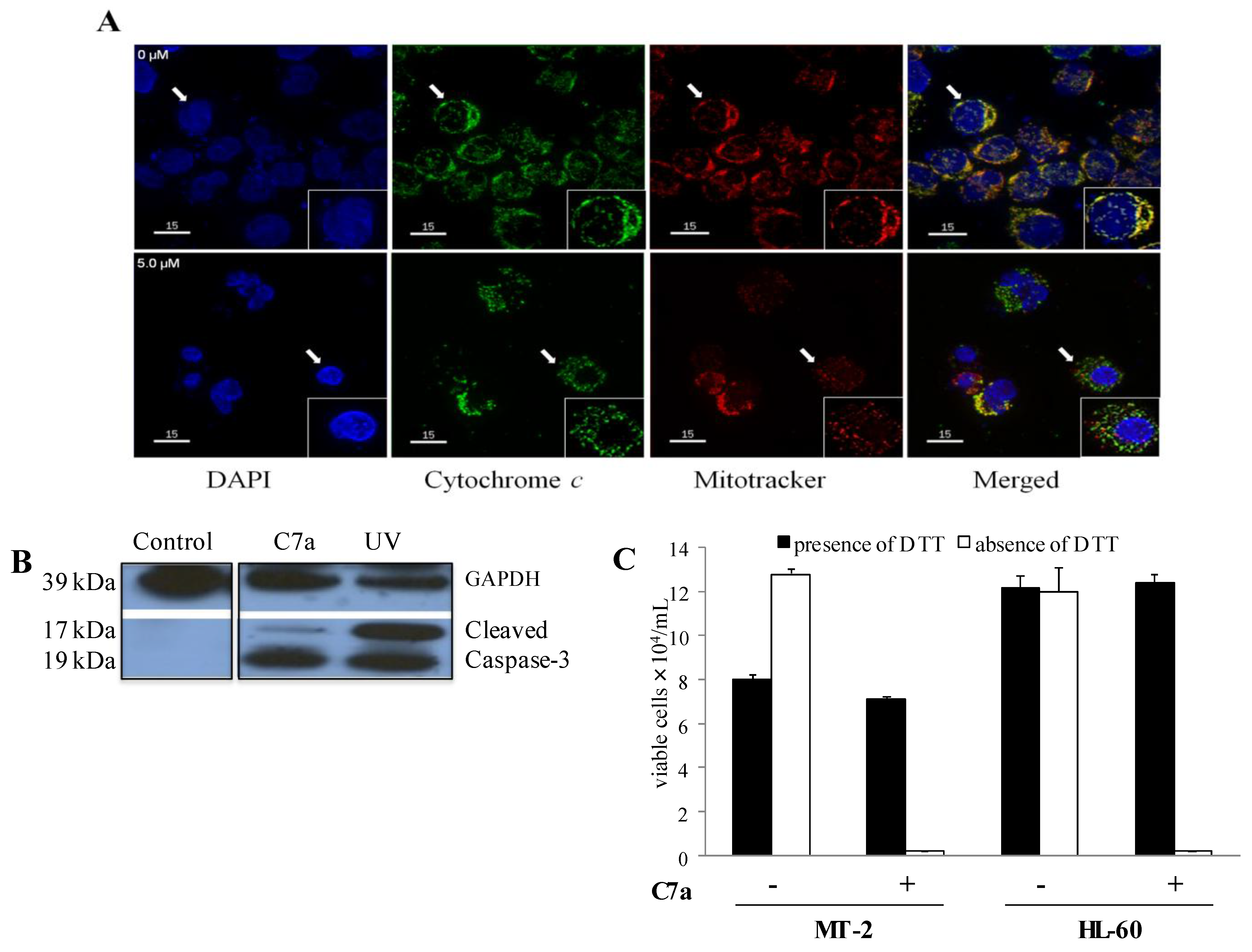
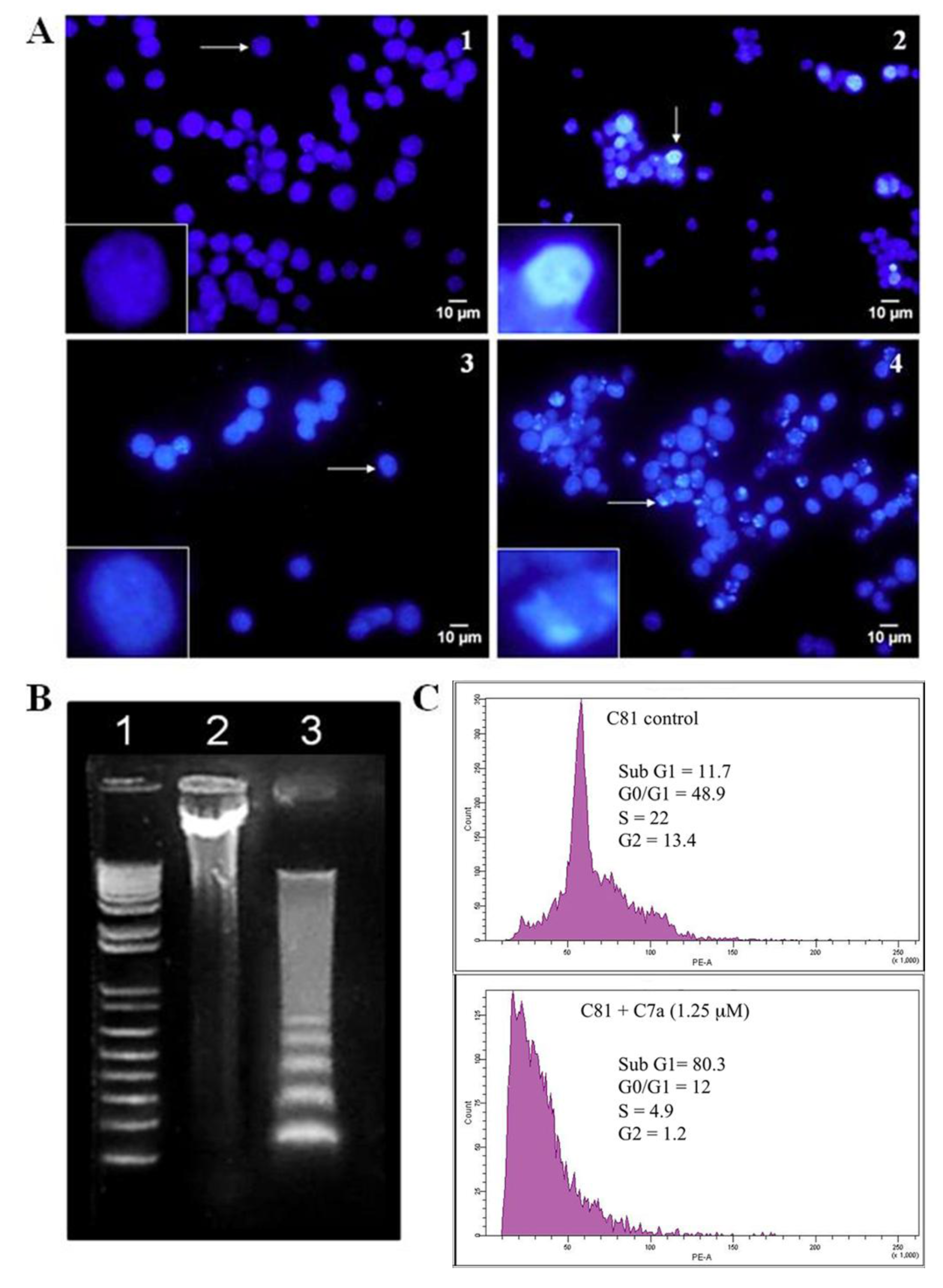
2.3. C7a Induces the Intrinsic Apoptotic Pathway in Human T Cell Leukemia Lines
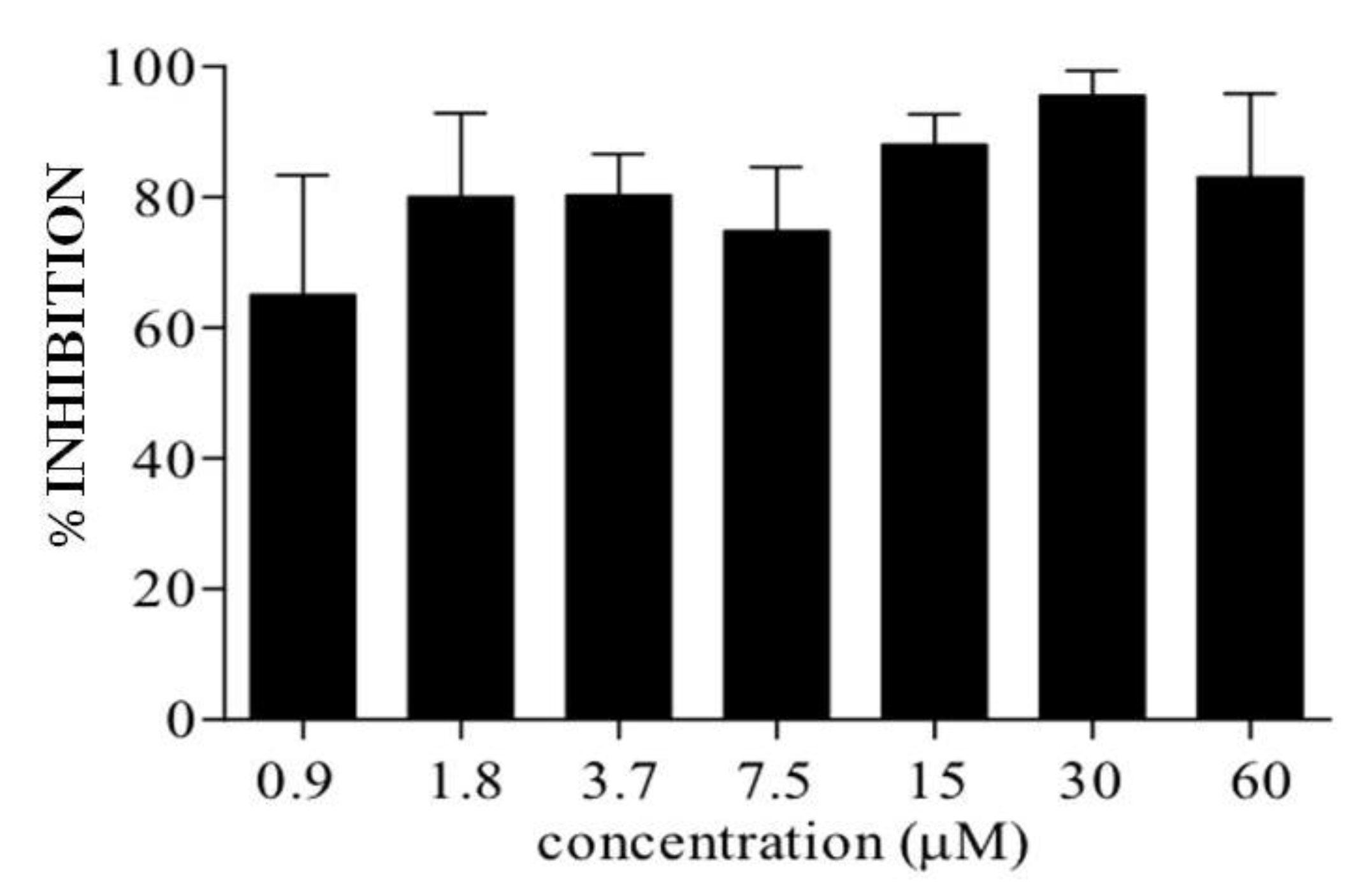
2.4. C7a Treatment Reduces Spontaneous Proliferation of PBMC from HTLV-1-Infected Patients
3. Discussion
4. Experimental Section
4.1. Culture Conditions for Human Leukemia Cell Lineages and PBMC Isolation
4.2. Analysis of Nuclear Alterations
4.3. Flow Cytometry Analysis
5. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Nasr, R.; El Hajj, H.; Kfour, Y.; de Thé, H.; Hermine, O.; Bazarbachi, A. Controversies in targeted therapy of adult T cell leukemia/lymphoma: ON target or off target effects? Viruses 2011, 3. in press. [Google Scholar] [CrossRef] [PubMed]
- Feuer, G.; Zack, J.A.; Harrington, W.J., Jr.; Valderama, R.; Rosenblatt, J.D.; Wachsman, W.; Baird, S.M.; Chen, I.S. Establishment of HTLV- I T-cell lymphomas in severe combined immunodeficient mice. Blood 1993, 82, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Feuer, G.; Stewart, S.A.; Baird, S.M.; Lee, F.; Feuer, G.; Chen, I.S. Potential role of natural killer cells in controlling tumorigenesis by HTLVs. J. Virol. 1995, 69, 1328–1333. [Google Scholar] [CrossRef] [PubMed]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells. J. Immunol. 2005, 174, 6477–6489. [Google Scholar] [CrossRef]
- Shultz, L.D.; Ishikawa, F.; Greiner, D.L. Humanized mice in translational biomedical research. Nat. Rev. Immunol. 2007, 7, 118–130. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.A.; Stevanato, A.; Treu-Filho, O.; Netto, A.V.; Mauro, A.E.; Castellano, E.E.; Carlos, I.Z.; Pavan, F.R.; Leite, C.Q. Antimycobacterial and antitumor activities of palladium(II) complexes containing isonicotinamide (isn): X-ray structure of trans-[Pd(N3)2(isn)(2)]. Eur. J. Med. Chem. 2010, 45, 4863–4868. [Google Scholar] [CrossRef] [PubMed]
- Vrzal, R.; Starha, P.; Dvorák, Z.; Trávnícek, Z. Evaluation of in vitro cytotoxicity and hepatotoxicity of platinum(II) and palladium(II) oxalato complexes with adenine derivatives as carrier ligands. J. Inorg. Biochem. 2010, 104, 1130–1132. [Google Scholar] [CrossRef] [PubMed]
- Caires, A.C. Recent advances involving palladium (II) complexes for cancer. Anticancer Agents Med. Chem. 2007, 7, 484–491. [Google Scholar] [CrossRef]
- Rodrigues, E.G.; Silva, L.S.; Fausto, D.M.; Hayashi, M.S.; Dreher, S.; Santos, E.L.; Pesquero, J.B.; Travassos, L.R.; Caires, A.C. Cyclopalladated compounds as chemotherapeutic agents: Antitumoral activity against a murine melanoma cell line. Int. J. Canc. 2003, 107, 489–504. [Google Scholar] [CrossRef]
- Serrano, F.A.; Matsuo, A.L.; Monteforte, P.; Bechara, A.; Smaili, S.S.; Santana, D.P.; Rodrigues, T.; Pereira, F.V.; Silva, L.S.; Machado, J., Jr.; et al. A cyclopalladated complex interacts with mitochondrial membrane thiol-groups and induces the apoptotic intrinsic pathway in murine and cisplatin-resistant human tumor cells. BMC Cancer 2011. submitted for publication. [Google Scholar] [CrossRef]
- Hebeler-Barbosa, F.; Rodrigues, E.G.; Puccia, R.; Caires, A.C.; Travassos, L.R. Gene therapy against murine melanoma B16F10-Nex2 using IL-13Rα2-Fc chimera and interleukin 12 in association with a ciclopalladated drug. Trans. Oncol. 2008, 1, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Santana, D.P.; Faria, P.A.; Paredes-Gamero, E.J.; Caires, A.C.; Nantes, I.L.; Rodrigues, T. Palladacycles catalyse the oxidation of criticals thiols of the mitochondrial membrane proteins and lead to mitochondrial permeabilization and cytocrome c release associated with apoptosis. Biochem. J. 2009, 417, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.M.; Nicot, C. HTLV-I and Apoptosis: Role in cellular transformation and recent advances in therapeutic approaches. Apoptosis 2008, 13, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Hermine, O.; Bouscary, D.; Gessain, A.; Turlure, P.; Leblond, V.; Franck, N.; Buzyn-Veil, A.; Rio, B.; Macintyre, E.; Dreyfus, F.; Bazarbachi, A. Treatment of ATLL with zidovudine and interferon-α. N. Engl. J. Med. 1995, 332, 1749–1751. [Google Scholar] [CrossRef] [PubMed]
- Foss, F.M.; Waldmann, T.A. Interleukin-2 receptor-directed therapies for cutaneous lymphomas. Hematol. Oncol. Clin. North Am. 2003, 17, 1449–1458. [Google Scholar] [CrossRef]
- Hishizawa, M.; Kanda, J.; Utsunomiya, A.; Taniguchi, S.; Eto, T.; Moriuchi, T.; Tanosaki, R.; Kawano, F.; Miyazaki, Y.; Masuda, M.; et al. Transplantation of allogeneic hematopoietic stem cells for ATL: A nationwide retrospective study. Blood 2010, 116, 1369–1376. [Google Scholar] [CrossRef]
- Che, X.F.; Zheng, C.L.; Owatari, S.; Mutoh, M.; Gotanda, T.; Jeung, H.C.; Furukawa, T.; Ikeda, R.; Yamamoto, M.; Haraguchi, M.; et al. Overexpression of survivin in primary ATL cells and sodium arsenite induces apoptosis by down-regulating survivin expression. Blood 2006, 107, 4880–4887. [Google Scholar] [CrossRef]
- Dewan, M.Z.; Uchihara, J.N.; Terashima, K.; Honda, M.; Sata, T.; Ito, M.; Fujii, N.; Uozumi, K.; Tsukasaki, K.; Tomonaga, M.; et al. Efficient intervention of growth and infiltration of primary adult T-cell leukemia cells by an HIV protease inhibitor, ritonavir. Blood 2006, 107, 716–724. [Google Scholar] [CrossRef]
- Hermine, O.; Dombret, H.; Poupon, J.; Arnulf, B.; Lefrere, F.; Rousselot, P.; Damaj, G.; Delarue, R.; Fermand, JP.; Brouet, JC.; et al. Phase II trial of arsenic trioxide and alpha interferon in patients with relapsed/refractory adult T-cell leukemia/lymphoma. Hematol. J. 2004, 5, 130–134. [Google Scholar] [CrossRef]
- Horie, R.; Watanabe, T.; Umezawa, K. Blocking NF-kappaB as a potential strategy to treat adult T-cell leukemia/lymphoma. Drug News Perspect. 2006, 19, 201–209. [Google Scholar] [CrossRef]
- Tomita, M.; Kawakami, H.; Uchihara, J.N.; Okudaira, T.; Masuda, M.; Matsuda, T.; Tanaka, Y.; Ohshiro, K.; Mori, N. Inhibition of constitutively active Jak-Stat pathway suppresses cell growth of HTLV-1-infected T-cell lines and primary adult T-cell leukemia cells. Retrovirology 2006, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, B.; Niewiesk, S.; Lairmore, M.D. Mouse models of HTLV type-1-associated adult T-cell leukemia/lymphoma. Vet. Pathol. 2010, 47, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Mahieux, R.; Pise-Masison, C.; Gessain, A.; Brady, J.N.; Olivier, R.; Perret, E.; Misteli, T.; Nicot, C. Arsenic trioxide induces apoptosis in HTLV type 1-and type 2-infected cells by a caspase-3-dependent mechanism involving Bcl-2 cleavage. Blood 2001, 98, 3762–379. [Google Scholar] [CrossRef]
- Mahieux, R.; Hermine, O. In vivo and in vitro treatment of HTLV-1 and HTLV-2 infected cells with arsenic trioxide and interferon-alpha. Leuk. Lymphoma 2005, 46, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Lin-Shiau, S.Y.; Lin, J.K. Involvement of reactive oxygen species and caspase 3 activation in arsenite-induced apoptosis. J. Cell. Physiol. 1998, 177, 324–333. [Google Scholar] [CrossRef]
- El Hajj, H.; El-Sabban, M.; Hasegawa, H.; Zaatari, G.; Ablain, J.; Saab, S.T.; Janin, A.; Mahfouz, R.; Nasr, R.; Kfoury, Y.; et al. Therapy-induced selective loss of leukemia-initiating activity in murine adult T cell leukemia. J. Exp. Med. 2010, 207, 2785–2792. [Google Scholar] [CrossRef]
- Oliveira, C.R.; Barbosa, C.M.; Nascimento, F.D.; Lanetzki, C.S.; Meneghin, M.B.; Pereira, F.E.; Paredes-Gamero, E.J.; Ferreira, A.T.; Rodrigues, T.; Queiroz, M.L.; et al. Pre-clinical antitumour evaluation of Biphosphinic Palladacycle Complex in human leukaemia cells. Chem. Biol. Interact. 2009, 177, 181–189. [Google Scholar] [CrossRef]
- Popovic, M.; Flomenberg, N.; Volkman, D.J.; Mann, D.; Fauci, A.; Dupont, B.; Gallo, R.C. Alteration of T-cell functions by infection with HTLV-I or HTLV-II. Science 1984, 226, 459–462. [Google Scholar] [CrossRef]
- Gisselbrecht, C.; Gaulard, P.; Lepage, E.; Coiffier, B.; Brière, J.; Haioun, C.; Cazals-Hatem, D.; Bosly, A.; Xerri, L.; Tilly, H.; et al. Prognostic significance of T-cell phenotype in aggressive non-Hodgkin’s lymphomas: Groupe d’Etudes des Lymphomes de l’Adulte (GELA). Blood 1998, 92, 76–82. [Google Scholar]
- Shu, S.T.; Nadella, M.V.; Dirksen, W.P.; Fernandez, S.A.; Thudi, N.K.; Werbeck, J.L.; Lairmore, M.D.; Rosol, T.J. A novel bioluminescent mouse model and effective therapy for adult T-cell leukemia/lymphoma. Canc. Res. 2007, 67, 11859–11866. [Google Scholar] [CrossRef]
- Gatza, M.L.; Dayaram, T.; Marriott, S.J. Ubiquitination of HTLV-I Tax in response to DNA damage regulates nuclear complex formation and nuclear export. Retrovirology 2007, 4, 95. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2011 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimaraes-Correa, A.B.; Crawford, L.B.; Figueiredo, C.R.; Gimenes, K.P.; Pinto, L.A.; Grassi, M.F.R.; Feuer, G.; Travassos, L.R.; Caires, A.C.F.; Rodrigues, E.G.; et al. C7a, a Biphosphinic Cyclopalladated Compound, Efficiently Controls the Development of a Patient-Derived Xenograft Model of Adult T Cell Leukemia/Lymphoma. Viruses 2011, 3, 1041-1058. https://doi.org/10.3390/v3071041
Guimaraes-Correa AB, Crawford LB, Figueiredo CR, Gimenes KP, Pinto LA, Grassi MFR, Feuer G, Travassos LR, Caires ACF, Rodrigues EG, et al. C7a, a Biphosphinic Cyclopalladated Compound, Efficiently Controls the Development of a Patient-Derived Xenograft Model of Adult T Cell Leukemia/Lymphoma. Viruses. 2011; 3(7):1041-1058. https://doi.org/10.3390/v3071041
Chicago/Turabian StyleGuimaraes-Correa, Ana B., Lindsey B. Crawford, Carlos R. Figueiredo, Karina P. Gimenes, Lorena A. Pinto, Maria Fernanda Rios Grassi, Gerold Feuer, Luiz R. Travassos, Antonio C.F. Caires, Elaine G. Rodrigues, and et al. 2011. "C7a, a Biphosphinic Cyclopalladated Compound, Efficiently Controls the Development of a Patient-Derived Xenograft Model of Adult T Cell Leukemia/Lymphoma" Viruses 3, no. 7: 1041-1058. https://doi.org/10.3390/v3071041