Recent Updates on Mouse Models for Human Immunodeficiency, Influenza, and Dengue Viral Infections
by
 ,
,
Vinodhini Krishnakumar
1,†, 
Siva Sundara Kumar Durairajan
1,*,†,
Kalichamy Alagarasu
2,
Min Li
3,* and
Aditya Prasad Dash
4 1
Department of Microbiology, School of Life Sciences, Central University of Tamilnadu, Tiruvarur 610 005, India
2
Dengue/Chikungunya Group, ICMR-National Institute of Virology, Pune 411001, India
3
Neuroscience Research Laboratory, Mr. & Mrs. Ko Chi-Ming Centre for Parkinson’s Disease Research, School of Chinese Medicine, Hong Kong Baptist University, Kowloon Tong, Hong Kong, HKSAR, China
4
Central University of Tamilnadu, Tiruvarur 610 005, India
*
Authors to whom correspondence should be addressed.
†
Authors contributed equally.
Viruses 2019, 11(3), 252; https://doi.org/10.3390/v11030252
Submission received: 14 January 2019
/
Revised: 9 February 2019
/
Accepted: 19 February 2019
/
Published: 13 March 2019
(This article belongs to the Special Issue Animal Models for Viral Diseases)
Abstract
:Well-developed mouse models are important for understanding the pathogenesis and progression of immunological response to viral infections in humans. Moreover, to test vaccines, anti-viral drugs and therapeutic agents, mouse models are fundamental for preclinical investigations. Human viruses, however, seldom infect mice due to differences in the cellular receptors used by the viruses for entry, as well as in the innate immune responses in mice and humans. In other words, a species barrier exists when using mouse models for investigating human viral infections. Developing transgenic (Tg) mice models expressing the human genes coding for viral entry receptors and knock-out (KO) mice models devoid of components involved in the innate immune response have, to some extent, overcome this barrier. Humanized mouse models are a third approach, developed by engrafting functional human cells and tissues into immunodeficient mice. They are becoming indispensable for analyzing human viral diseases since they nearly recapitulate the human disease. These mouse models also serve to test the efficacy of vaccines and antiviral agents. This review provides an update on the Tg, KO, and humanized mouse models that are used in studies investigating the pathogenesis of three important human-specific viruses, namely human immunodeficiency (HIV) virus 1, influenza, and dengue.
Keywords:
infectious diseases; human viruses; mouse models; knockout mice; transgenic mice; humanized mice; HIV; influenza; dengue1. Introduction
Infectious diseases caused by human pathogenic viruses remain a huge threat to global health. An estimated 37 million persons are living with human immunodeficiency virus (HIV) infections [1]. More than 90 million new cases of influenza occur every year in children aged less than five years; of these, 20 million are acute lower respiratory infections and one million are severe acute lower respiratory tract infection (ALRI) cases [2]. The mechanisms of how these pathogenic viruses cause human disease differ widely. Using cell culture systems, the effect of these pathogenic viruses on the biology of the cell can be investigated. Organotypic cultures provide an additional opportunity for studying viral spread and tissue remodelling, but offer limited potential for studying host-pathogen interactions [3]. Virologists attempt to understand how the replication of a virus within a host induces an infection process which may be associated with symptoms, disease, or a sub-clinical infection. Animal models are important for investigating the exact in vivo circumstances, and furthermore, are fundamental for developing counter measures against infectious diseases.
Mouse models are the preferred animal models for studying human diseases due to their ready availability, low cost, and convenient husbandry requirements. In addition, species-specific reagents can be used and certain species are amenable to genetic manipulation. With mouse models, the host response to infection can be analyzed in detail and the effectiveness of drugs and vaccines, as well as their safety, can be assessed. However, there are certain drawbacks in the utilization of mouse models for research and in the translation of this animal data to humans. Viruses exhibit high species-specificity and specific tropism, depending on the presence or absence of viral entry receptors, as well as specific innate immune response factors in the mouse cells. Hence, most mouse models show reduced susceptibility to infection by human viruses and do not show disease symptoms. Indeed, with any human virus, investigating immune responses in vivo is arduous, even when utilizing appropriate animal models.
Genetically engineered mouse (GEM)/Transgenic mouse (Tg) models enable researchers to perceive the mechanisms behind viral pathogenesis. Advances in transgenic technology have enabled the analysis of the role of the specific viral proteins found in affected cells and the mechanisms of host susceptibility and disease symptoms. Researchers have developed efficient tools to investigate immune responses to viral infection in vivo. Mice with germline genetic modifications have revealed the mechanisms involved in viral pathogenesis and anti- viral immune responses. Thus, for instance, to analyze virus-specific T-cell responses, Tg mouse models that express T-cell receptor (TCR) transgenes specific to viral proteins have been widely employed. These transgenic mice include mice expressing a transgenic TCR specific to influenza virus hemagglutinin (HA) in the context of the major histocompatibility complex (MHC) class II molecules. These mice, when infected with mouse adapted strains of influenza, show both a CD4+ and CD8+ T cell response, and thus have been used to study the role of CD4+ T cells in the activation of CD8+T cells [4,5]. Furthermore, many human viruses like hepatitis viruses, polioviruses, papillomaviruses, HIV, and measles are not infectious to wild-type mice, but the transgenic mice expressing specific human receptors that aid in the entry of those viruses are susceptible to many of these viruses. These susceptible Tg mice can be used to evaluate the virus’ pathogenesis as in vivo models [6]. Although the transgenic mice are useful in evaluating the involvement of key viral proteins in disease pathogenesis, they cannot recapitulate the entire disease symptom complex as in the host.
Knockout (KO) mice devoid of specific genes have been used to identify and evaluate the cellular and molecular entities involved in the adaptive and innate immune responses that are important in controlling viral infections. These KO mice include: TCR-B KO mice lacking alpha beta T-cells [7]; recombination activation gene (RAG)-1 and RAG-2 KO mice that are devoid of mature B and T cells [8]; KO mice for type I and type II interferons (IFNs) or IFN receptors which lack an antiviral response [9]; KO mice for immune receptors, such as Toll like receptors (TLRs), retinoic acid-inducible gene-I (RIG-I)-like receptors (RLRs) and nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs) [10]; and KO mice for transcription factors, such as Signal Transducer and Activator of Transcription (STAT) molecules [11] and IFN regulatory factors (IRFs) [12] that participate in linking signals from receptors to downstream kinases and transcription factors [10,11].
Humanized mouse models are an advanced approach for evaluating the human immune response to viral pathogens, as well as to unravelling the pathogenic mechanisms of viral diseases. Severely immunodeficient mouse strains are engrafted with human cells and/or tissue xenografts to produce humanized mice (Figure 1). The major breakthrough has been the discovery of mice with mutations in the gene coding for the protein kinase DNA-activated catalytic polypeptide mutation (Prkdc). These mice, called severe combined immunodeficient (SCID) mice, allow the poor engraftment of human fetal tissues, human peripheral blood mononuclear cells (PBMCs), and haematopoetic stem cells (HSCs). Further, mice with mutations in the RAG- 1 or 2 genes (Rag1null or Rag2null, respectively) also have T and B cell deficiency. Backcrossing of mice with the PrkdcSCID mutation (CB17-PrkdcSCID) to mice on the NOD (non-obese diabetic) background resulted in NOD.CB17-PrkdcSCID/J mice, which allowed for the improved engraftment of HSCs, but they developed thymic lymphomas and had a short life span. The production of immunodeficient mice with mutations in the interleukin (IL) 2 receptor common gamma chain (IL2rγnull) allows for higher rates of the engraftment of HSCs due to the absence of natural killer (NK) cells [13]. When IL2rγnull mice were merged with Prkdcsid or SCID mice or with recombination of activating genes (Rag) 1 or 2 null mice (Rag1null or Rag2null), the resulting hybrids lacked adaptive immunity and expressed acute deficiencies of innate immunity, besides lacking murine natural killer (NK) cells and allowing for the engraftment of human cells at high levels [14,15,16]. Adopting this same approach, mice harbouring human hematopoietic cells, myoblasts, neurons, hepatocytes, and different kinds of epidermal derivatives have been raised. Invariably, these humanized mice are susceptible to a spectrum of viruses, and humanized mice can be used to investigate the diseases caused by these viruses (e.g., dengue and HIV-I in mice with a human immune system).
Three strains of immunodeficient IL2rγnull mice, namely NOD Cg-PrkdcidII2rgtm1WjI (NSG), NOD Cg- PrkdcsidII2rγtm1Sug (NOG), and C;129S4-Rag2tm1FvII2rγtm1F1v (popularly denoted as BALB/c-Rag2null, IL2rγnull, or BRGS mice with the NOD SIRPa variant) [14,16,17,18,19], are commonly used today. NOG mice possess a truncated cytoplasmic domain of the gamma chain, but lack the signalling domain, while NSG and BRG mice completely lack the gamma chain. The biological responses of humanized IL2rγnull mice, when engrafted with human tissues, cells, and immune structures, recapitulate those observed in humans better than any previously known models of humanized mice [14,16,17,18,19].
Three common approaches are used to engraft human immune systems into immunodeficient IL2rγnull mice. The first model, denoted as the human peripheral blood leuckocytes (Hu-PBL)-SCID model, is generated by the injection of human PBMCs followed by the engraftment of human CD3+T-cells after the first week. These mice develop a lethal xenogeneic graft-versus-host disease (GVHD) and their life span is short [16,17,18,19]. The second model, referred to as the humanized SCID repopulating cell (Hu-SRC)-SCID model, is created by the intravenous (IV) or intrafemoral injection of precise SRC, i.e., human CD34+ HSCs acquired from bone marrow, fetal liver, cord blood, or peripheral blood. This model (referred to as HSC engrafted) affirms engraftment of the entire human immune system. The third model is the bone marrow/liver/thymus (BLT) model, developed by transplantation of the human fetal liver and thymus below the kidney and IV injection of autologus fetal liver HSCs [20,21]. Similar to the Hu-SRC-SCID model, all lineages of human hematopoietic cells grow. In spite of the development of a healthy mucosal human immune system by BLT mice, the human T cells are educated in an autologous human thymus and are human leuckocyte antigen (HLA) restricted. One of the salient caveats of the BLT model is that, in most laboratories, the mice develop a dissipating GVHD-like syndrome that narrows the window for experimentation [16,17,18,19].
Every model is endowed with specific advantages and restrictions; hence, selecting an appropriate model for a viral disease of interest is of paramount importance. Schematic representation of the generation of transgenic, KO, and humanized mouse models using genetice engineering is depicted in Figure 1 and Figure 2. The most widely used and recently reported KO, Tg, and humanized mouse models of HIV, influenza, and dengue are discussed and presented in Table 1, Table 2 and Table 3.
2. Human Immuno Deficiency Virus
Acquired immunodeficiency syndrome (AIDS) is a spectrum of conditions caused by infections of HIV type 1 (HIV-1) and HIV type 2 (HIV-2). HIV-1 is a major contributor to the global AIDS pandemic. More than 70 million individuals are living with HIV and 940,000 affected individuals died due to AIDS in 2017 [1]. The introduction of antiretroviral therapy (ART) is one of the most significant inventions in modern medicine, providing effective virus suppression and radically enhancing the quality of life and life span of HIV patients; however, the treatment needs to be continued for life. The major barriers posing a stumbling block to developing an effective vaccine and treatment methods are: (i) HIV’s inherent ability to mutate frequently, specifically its external envelope glycoproteins, resulting in the escape of immune variants; (ii) persistence of HIV in resting cells, which leads to incomplete suppression; and (iii) HIV infection in humans induces immune suppression, resulting from the massive and continuous depletion of CD4+ T-cells. Notwithstanding these barriers, encouraging developments in an HIV vaccine have been made, and various promising numbers of vaccines are being developed. So far, animal models are the best approach we have to deciphering the mechanisms of pathogenesis, disease progression, latency, and emergence of drug resistance mutations. Moreover, animal models are important for testing the efficacy of vaccines and anti retroviral drugs [22].
2.1. Knockout Mouse Models of HIV
Recently, retroviral restriction factors have drawn the attention of researchers. HIV-1 infection and replication is restricted by a number of host proteins, known as restriction factors, that interfere with the critical processes involved in the life cycle of retroviruses. Four of these are sterile alpha motif domain-containing protein 1 (SAMHDI) [23,24,25], apolipoprotein B mRNA editing enzyme catalytic subunit 3G (APOBEC3G) [26], viperin [27], and tetherin [28]. These restriction factors inhibit viral replication and the production of new progeny virions by preventing reverse transcription, by inducing viral DNA to mutate, and by promoting TLR 7- and TLR9-mediated synthesis of type I IFN by plasmacytoid dendritic cells (pDCs).
Mostly, these restriction factors are conserved and orthologous in mice. KO mouse models (Table 1) help researchers to validate in vitro results obtained from cultured cells and to gain a better understanding of the biology of restriction factors. In vivo studies of mouse models provide greater insight into the mechanisms by which the proteins act to restrict viral infection and replication. Wittmann et al. [23] and Bloch et al. [29] showed that SAMHD1 in the KO mouse model blocks retroviral infection and that endogenous mouse SAMHD1 restricts not only HIV-1, but also MLV reporter virus infection at the level of reverse transcription in primary myeloid cells. Intracellular dNTP levels and SAMHD1 phosphorylation in different T-cell types are the two most important factors affecting the antiviral activity of murine SAMHD1. They also reported that the antiviral restriction in a mouse by SAMHD1 is mechanically similar to that in humans in terms of dNTP hydrolase activity and cell-dependent phosphorylation. This study provided evidence that the SAMHD1 KO mouse is a valuable tool for studying the replication of different viruses, including retroviruses, retro elements, and DNA viruses [23,29].
2.2. Transgenic Mouse Models of HIV
To overcome the barrier to HIV entry and infection in mouse cells, Browning et al. [30] generated human CD4 and CCR5 double Tg mice (hu CD4/CCR5 Tg mice). Though human CD4 and a chemokine receptor CCR5 helped HIV-1 to enter mouse cells, establishment of further in vivo infection and replication in these human CD4/CCR5 Tg mice was not observed due to other cellular blocks [31]. Tg rats with human CD4 and CCR5 also supported HIV infection; however, as in double Tg mice, spread of the virus was blocked [32]. Transient trans complementation of human Tat interacting protein Cyclin T1 elevated HIV gene expression, suggesting novel ways of enhancing HIV infection and replication in rat models [32]. Multiple research groups have constructed different Tg mice that express the whole or partial HIV-1 genome. They include Tg mice expressing the HIV-1 genome with modified long terminal repeats (LTR), resulting in cataracts, weeping eyes, and wasting in Tg mice [33]; 3′ half of the HIV genome, causing severe nephropathy [34]; and expression of HIV-1 Tat or Nef proteins, leading to epidermal hyperplasia [35]. Hanna et al [36] constructed Tg mice by expressing entire HIV coding sequences in T-cells and cells of the macrophage/dendritic cell lineage under the control of a human CD4 promoter flanked by an enhancer of mouse CD4. Tg mice expressing defective provirus with gag and pol genes deleted or expressing individual HIV-1 genes have been shown to develop various pathologies resembling those in humans [37]. In vivo, a negative factor (Nef) was observed to promote viral replication and pathogenicity. Tg mice expressing entire coding sequences of HIV-1 (CD4/HIVWT) or HIV-1 Nef alone (CD4C/HIVNef) under the control of a CD4 promoter in HIV-1 target cells showed human AIDS-like symptoms [38]. Inducible HIV-1 type Nef Tg mice (Table 2) were generated using a tetracycline-inducible system to prevent the developmental defects due to the continuous expression of Nef in CD4C/HIVnef Tg mice. The Nef gene was induced in (CD4C/rtTA X TRE/HIVNef) or (CD4C/rtTA2S-M2 X TRE/HIVNef) double-Tg mice upon doxycycline (DOX) treatment, and these mice developed disease similar to that seen in constitutively Nef-expressing
CD4C/HIVNef Tg mice. In the absence of lymphopenia, CD4+ T-cell activation was observed only in Nef-expressing T-cells, but not in T-cells that do not express Nef. This model helped in understanding the functions of Nef [39]. A Tg mouse that expresses gp120 of HIV-1 in the astrocytes developed neuropathological features similar to those observed in AIDS patients with neurological symptoms and can be utilized in neuro AIDS research [40]. The same mouse model expressing gp120 of HIV-1 has been utilized to demonstrate the pathological role of CCR5 in HIV-associated brain injury. Genetic ablation of CCR5 inhibits microgilial activation (hallmark of brain pathology) in a Tg mouse model expressing envelope glycoprotein 120 (gp120) that uses CXCR4 as a receptor to induce HIV-associated brain injury. A pharmacological blockade of CCR5 and the acute phase protein lipocalin prevent neurotoxicity in CCR5-deficient mice expressing gp120 of HIV-1 [41]. Different research groups have used Tg mice extensively for studies related to HIV-associated nephropathy. To list a few, Tg26 and its variants, as well as CD4/HIV Tg mice and variants, have been used [42]. Tg26 is a transgenic mouse which expresses seven of the nine HIV proteins under the control of a viral LTR promoter. Since the DNA expressed is replication deficient, it randomly integrates into the host genome, and transcription of these genes takes place. Tg26 is used to study the long-term effect of HIV proteins on the host. Tg mouse models which express the entire genome or selected genes of HIV have given productive results and information on pathogenesis [42].
2.3. Humanized Mouse Models of HIV
Humanized mice have provided the opportunity to study HIV entry and infection, virology, latency, and disease progression. They are good and convenient models for testing prophylactic drugs and anti-HIV antibodies. HIV infection, replication, and pathogenesis can be studied in vivo using Hu-PBL-SCID mice. Human disease pathogenesis can be studied using HSCs-engrafted and BLT models. BLT models can be infected with either CCR5 or CXCR4-tropic HIV strains to study HIV mutations and how they escape from the CD8+ T-cell response [43,44,45,46]. NOD-SCID BLT humanized mice were used to study cell-to-cell transmission and retroviral spread in vivo; their results provided new insights and novel approaches for blocking the spread of viruses [47]. NOD-SCID BLT models have been helpful in understanding the utility of ART in decreasing the transmission of HIV from infected individuals to their partners. Studies of HIV-infected NSG-BLT mice revealed that ART significantly suppresses HIV in cervico-vaginal secretions and restores the CD4+ and CD8+ T-cell numbers [48]. A rapid rebound of viremia was also demonstrated when ART was discontinued in NSG-BLT mice [48]. Despite the many other uses of BLT-humanized mice, the development of GVHD in these mice limited their use for HIV cure studies until the development of C57BL/6 Rag2−/−γc −/−CD47−/− triple knockout (TKO)-BLT mice. Lavender et al. [49] reported that, in TKO-BLT mice, HIV-1 latency can be maintained over unlimited periods when the mice are on ART, and quick viral recovery occurs following therapy removal. Compared to other BLT models, TKO-BLT mice offer a sufficient time to investigate the effects of extended periods of ART (15–18 weeks) on the latent reservoir and it delays in recrudescence in HIV-1 cure studies [49,50].
Advancement in the technology and understanding of the usage of animal models has enabled researchers to explore the establishment of stable, integrated, and non-productive states of HIV infection in individual cells, termed HIV latency; this latency has been the primary barrier to the development of an HIV cure, and is the most studied aspect of the virus. Brooks et al. [51] used SCID-hu thy/lv mice to demonstrate HIV latency by injecting HIV into human thymic organoids that develop in these animals. Following the above model and strategy, different studies have reported reconstituting many types of tissues in different humanized mice (human thymus and liver mice (Thy/Liv mice); human CD34+ HSCs (huCD34 mice), or both BLT mice). NOD/ SCID (NS), NOD/SCID IL2rgc−/− (NSG), or NOD/Rag1−/− IL2rgc −/− (NRG)). A humanized mice model reconstituted with different tissues and organs appears to be an ideal model to study the effect of ART on systemic HIV infection [52,53,54,55,56]. Upon ART treatment, huCD34 and BLT humanized mice (Table 3) tissues reconstituted with human hematopoietic lineages resulted in the repression of viral replication, reduction in plasma viral RNA (vRNA) below the detection limit, and maintainence or preservation of the CD4+ T-cell counts [45,46,47,48,49,50,51,52,53,54,55,56,57]. However, ART could not absolutely remove HIV infection, and the presence of latent viruses has been demonstrated in huCD34-NSG humanized mouse models [46,57]. The newer generation of T-cell-only mice (TOM) or myeloid-only mice (MOM), developed by reconstituting specific hematopoietic cell lineages, allows investigators to study the individual contribution of T-cells or macrophages to HIV persistence in vivo. NSG-hu thy/liv mice were generated by implanting human fetal thymus and liver tissues under the kidney capsule. They are scientifically reconstituted with T-cells alone and not with any other lymphoid lineages like monocytes/macrophages, B cells, or DCs. These mice were susceptible to HIV-1 infection and did not develop any signs of GVHD. Following combination ART, HIV latent reservoirs and resting CD4+ T-cells were observed [58,59]. This clearly indicates that TOM humanized models can be used to study HIV latency. In addition to studies with these models, there are reports suggesting that macrophages can serve as HIV-1 reservoirs [58,60]. ART treatment of HIV-infected MOM mice suggested that the half-life of the infected macrophages from MOM mice is shorter than that of infected T-cells in BLT mice [60]. The seemingly unlimited ability to transplant human immune cells into immunodeficient mice has led to the development of a wide array of techniques and assays. One prominent example is the in vivo viral outgrowth assay that quantifies the latent HIV reservoir. This assay not only validates the results of in vitro latent viral detection, but also quantifies the viral content with a greater sensitivity [61,62]. Improvements with humanized mice are endless. Satheesan et al. [63] used a human HSC-engrafted NSG humanized model (hu-NSG) to investigate and demonstrate HIV latency in vivo. They demonstrated the ability of HIV-infected human cells from HIV-infected hu–NSG mice on combinatorial antiretroviral therapy (cART) to act as a latent HIV reservoir. This model is an attractive alternative to humanized BLT mice and SCID-hu-thy/liv mice, which require the surgical manipulation and reconstitution of immune cells and tissues models to study HIV latency and latent T-cell reservoirs [63]. Thus, mice models have immensely contributed to the improved understanding of HIV pathogenesis, persistence, prophylactic and therapeutic intervention; they could be a vital tool in the eradication of HIV.
3. Influenza Virus
Influenza A pandemics have caused considerable disease and deaths. While influenza A and B cause only acute febrile illness in a majority of infected people, people at high risk, including pregnant women, children below the age of five years, elderly people aged more than 65 years, and persons with chronic medical conditions, can develop severe disease which may be lethal [64]. Influenza infections may promote and intensify numerous conditions like chronic obstructive pulmonary disease (COPD) [65] and, asthma [66]; They can increase the risk of a cerebro-vascular accident and myocardial infarction [67], and fetal loss in pregnant women [68]. Though antiviral therapies, drugs, and vaccines are available, influenza viruses still threaten humans and animals all over the world. The diversity of the virus enables it to have multiple hosts, while antigenic drift and shift play major roles in the wide occurrence of epidemics and pandemics [69,70,71]. Mouse models have played a significant role in delineating the pathogenesis of influenza and in the development of vaccines and therapeutics against the different types of the virus. Laboratory inbred strains of mice, such as BALB/c, C57BL/6, and DBA/2j, can be infected with mouse adapted strains of influenza. DBA/2j is highly susceptible to non-adapted influenza strains [72,73,74]. The most common symptoms exhibited by the infected mice include hypothermia, anorexia, and weight loss. However, depending on the dose and the strains of the influenza virus used, the infection might be lethal. The most commonly used mouse adapted strains of influenza include H1N1 influenza A/Puerto Rico/8/34 (PR8) or H1N1 influenza A/WSN/1933 (WSN). However, certain strains, such as the 1918 H1N1 pandemic influenza A and the 2009 H1N1 pandemic influenza A, do not require adaptation to infect mice [72,73]. The lack of suitable small-animal models for studies of influenza pathogenesis and for the development of vaccines and antivirals is one of the most serious obstacles to progress in research.
3.1. Knockout Mouse Models of Influenza A
KO mice models have been widely used in understanding the two most important aspects of influenza virus, namely: (i) how the early inflammatory response to influenza virus leads to the progression of lung diseases; and (ii) how reassortment and parental strains differ in virulence. IL-1R1−/− mice infected with influenza virus were used to demonstrate the role of IL-1α and β in the pathogenesis of influenza. The study revealed that IL-1 contributes to the recruitment of CD4+ T cells to the site of infection and enhances IgM production, but also causes acute pulmonary inflammation [75]. Studies on IL-18−/− mice revealed the importance of IL-18 in restricting influenza virus replication in lungs by augmenting the NK cell response [75,76]. Since IL-1 and IL-18 are produced by macrophages through the caspase-1 pathway, Thomas and his group in 2009 investigated the role of cryopyrin, an NLR, in inducing caspase-1 using cryopyrin−/− and caspase 1−/− mice. Cryopyrin and caspase-deficient mice succumbed to infection, and their demise was attributed to the reduction in pro inflammatory cytokines which play a central role in innate immunity and in moderating lung pathology in influenza pneumonia [77]. Mice models knocked out for multiple genes can be used for studying the combined effects of different molecules on the influenza pathogenesis. TKO mice (Table 1) deficient in the three signalling receptors, namely tumor necrosis factor (TNF)-R1, TNF-R2, and IL-1-RI, were used to determine the combined contribution of the TNF and IL-1 inflammatory response to H5N1 influenza A infection. Triple mutant C57BL/6J x 129S background mice [IL1-RI single cytokine receptor knock-out mice (IL1-R KO), C57BL/ 6J x 129Sv -Il1r1tm1Roml/J), and single cytokine receptor TNFR1 KO mice (TNF-R KO, C57BL/6J x 129Sv-Tnfrsf1atm1Imx Tnfrsf1btm1Imx/J)] infected with H5N1 influenza A were observed to have longer survival, less morbidity, reduced lung inflammation, and a diminished cytokine response compared to wild-type mice. These results strengthen the suggested hypothesis that TNF and IL-1 contribute to the pathogenesis of H5N1 influenza A virus [78]. The anti inflammatory role of fatty acid binding protein 5 (FABP5) in H1N1 influenza A virus infection has been demonstrated using FABP5−/− mice. Infection of C57BL/6J background mice with H1N1 influenza A virus from which the FABP5 (FABP5−/−) gene had been deleted revealed that FABP5 deficiency augments excessive oxidative damage, lipid peroxidation, and inflammation [79].
The emergence of new pandemic viruses is mainly due to genetic mutations and reassortments. A reassortant seasonal H3N2 influenza A virus containing the PA gene of highly pathogenic avian influenza H5N1 influenza A caused severe pneumonia in Casp 1−/− mice [80]. Knockout mice models have been utilized to discern differences in the pathogenicity between reassortment and parental strains of influenza virus differences that may not be easily observed in wild-type mice.
3.2. Transgenic Mouse Models of Influenza A
Highly pathogenic avian H5 and H7 influenza A viruses continue to cross the species barrier and infect humans, causing diseases. Hence, they pose a constant threat and give rise to devastating pandemics [81,82]. Tg mice are powerful tools to identify the antiviral power of different human proteins that act as effective species barriers and also to identify the changes required in these avian viruses to overcome the species barriers. The IFN-regulated MX1 gene plays a major role in providing an effective innate immune response to control the influenza virus [83]. Tg mice expressing the entire human MX locus have been used to demonstrate that those Tg mice (Table 2) exhibit a higher resistance to H5 and H7 influenza A viruses, but are susceptible to H1N1 and H3N2 influenza A viruses. Moreover, it was also shown that engineered avian H7N7 influenza virus with signature mutations found in human viruses was able to overcome the resistance conferred by the Tg mice [84].
With the help of Tg mice, researchers can implement new strategies against the influenza virus. Wang et al. [85] came up with a novel anti-influenza strategy involving short-hairpin RNA (ShRNA) to disrupt the activity of hemagglutinin (HA) of influenza A virus, thereby inhibiting the virus. They developed a Tg mouse expressing an shRNA that specifically targets the conserved sequence of the HA of influenza A virus. The Tg mice displayed a constant ability to reduce influenza A virus infection and replication, suggesting the utility of an shRNA-based approach to prevent and control influenza virus infections in animals [85].
Tg mice expressing the MHC class I allele, HLA-A*02:01, were used to study the CD8+ T cell response to a heat inactivated H7N3 influneza A vaccine and to a modified vaccinia Ankara vectored vaccine expressing influenza virus epitopes. The studies reported that CD8+ T cells specific to immunodominant and subdominant epitopes were elicited, indicating the potential of the vaccine candidates [86,87]. Tg mice expressing the MHC class II allele, HLA-DR3, were used to evaluate the effect of an epitope-based vaccine candidate expressing cross conserved H1N1 influneza A CD4+ T cell epitopes and it was found that the viral load was reduced when the vaccinated HLA-DR3 expressing mice were challenged with pandemic 2009 H1N1 influenza A [88]. Therefore, Tg mice expressing HLA class I and class II alleles can be utilized in evaluating the CD8+ and CD4+ T cell responses to different influenza A vaccine candidates.
3.3. Humanized Mouse Models of Influenza A
Humanized mice models permit the testing of novel therapeutic approaches to control influenza, the identification of critical epitopes for vaccine development, the preclinical analysis of the human immune response against influenza vaccines, and the evaluation of vaccine cytotoxicity. Direct manipulation or boosting of the host immune system is being considered as an alternative therapeutic strategy, which will help the individuals to protect themselves against influenza virus [89]. An advanced strategy of augmenting the innate and adaptive immune response to infectious disease is to expand the γδ-T cells using phosphoantigens [90,91,92]. In huPBMC sreconstituted immunodeficient Rag2−/−γc−/− mice (Table 3) infected with H1N1 influenza A and H5N1 influneza A, phosphoantigen selectively activated and expanded Vδ2-T cells and controlled H1N1 infection, suggesting a new therapeutic approach [93].
Among the available vaccines, those that induce memory antibody responses with virus-neutralizing activity are considered the best vaccines against influenza virus infection. Due to the sequence variations in the envelope antigens HA and neuraminidase (NA), multiple subtypes of influenza A virus have evolved and complicated the effective vaccine development. Antigenic drift in the envelope proteins enables H1N1 influneza A and H3N1 influneza A viruses to escape from HA-binding antibodies. As a result, for adults who usually have exposure to seasonal influenza viruses, an annual single dose of the vaccine, boosting the humoral immune response, is sufficient. For emerging influenza viruses, a single dose of a vaccine is sufficient to induce protective immune responses due to the presence of cross-reactive pre-existing CD4+ T cells [94]. However, vaccines against H7N9 are less immunogenic [95]. Identification and modification of critical residues in the T cell epitope using immunoinformatics, followed by testing in a humanized (NOD/SCID/Jak3−/− (NOJ) mouse, revealed improvements in the HA-binding IgG response, suggesting the utility of a humanized mice model to recapitulate the cross-reactive memory response and subsequent response to the vaccine [96]. Humanized mice can also be used to generate monoclonal antibodies. Humanized mice HLA-A2. HLA-DR4. Rag1KO. IL2rγc KO. NOD, (DRAGA), which lack a murine immune system and express a functional human immune system, were used to develop cross-reactive human anti-influenza monoclonal antibodies (hu-mAb) to study influenza infection and investigate the efficacy of anti-influenza antibody-based therapeutics for human use. The hu mAB that targeted the HA protein of H1N1 was able to clear influenza infection in DRAGA mice [97].
The efficacy of vaccines developed using different strategies, including antiviral DNA vaccines, can be tested in humanized mice models. Ivanova et al. [98] generated anti-influenza IgG antibodies and influenza-specific cytotoxic T lymphocyte (CTL) activity by constructing a chimeric scFv-IP DNA molecule and administering it directly in experimental humanized NOD-SCID gamma mice. The pTriEx-sc22-IPJun/sc22-IP3-Fos chimeric DNA molecule efficiently binds human monocytes and evokes a strong humoral and CTL response in humanized mice [98]. Sasaki et al. [99] succeeded in establishing a humanized mouse model to demonstrate the evaluation of influenza vaccine safety based on the expression of bio marker genes (ZBP1, MX2, PSMB9, TAP2, CXCL11, CXCL9, TRAFD1, and PSME1) in lungs of the mice model. NOGs engrafted with human PBMCs were used to test NOD/Shi-SCID IL2rγnull mice in the short- and long-term. The results revealed that a short-term reconstitution model of NOD/Shi-SCID IL2rγnull is the most suitable model for biomarker gene-based safety evaluation of vaccines. Human CD14+ cells, pDCs, CD4+ and CD8+ T cells, and B cells were retained in the lungs in the short-term model, while human CD14+ cells and pDCs were not detected in the lungs of the long-term model. Moreover, increased levels of human cytokines and chemokines and the expression of human biomarker genes were observed in response to the toxicity of reference vaccines. The observed results suggest that a humanized model can be used for evaluating vaccine safety at the initial level in human peripheral blood mononuclear cells in vivo [99].
4. Dengue Virus (DENV)
Dengue is a vector-borne viral illness in humans, and it is endemic in more than 100 countries (Available online: http://www.who.int/denguecontrol/epidemiology/en/). Half of the world’s population is at risk for infection. Dengue virus (DENV) belongs to the family Flaviviridae and is closely related to hepatitis C, West Nile, Zika, [100] and Japanese encephalitis viruses. These family virus outbreaks are being reported throughout the world (Available online: http://www.who.int/csr/disease/epidemic-focus/flavivirus-epidemics/en/), since they co-circulate and cause similar symptoms, but produce different outcomes. Though several screening and control measures have been developed to detect and differentiate them at an early stage of infection [101], this family of viruses, especially dengue, still poses a great threat to public health. Four different serotypes of dengue have been reported to cause illness in humans (DENV-1-4). They occur concurrently in different parts of the world, and they are the most common viral infection in tropical and sub-tropical countries [102]. Availability of an appropriate small animal model for DENV infection has been, and remains, a challenge.
4.1. Knockout Mouse Models of DENV
At the beginning of the 20th century, researchers struggled to develop an animal model for dengue fever/dengue hemorrhagic fever (DF/DHF) [103]. None of them (small animal models, murine models, and hamsters) have shown any signs of dengue disease. Non-human primate models poorly developed any clinical disease and hence were not a suitable model for DF/DHF. This situation led to the development of different varieties of mouse models. Through cross breeding of mice lacking type I IFN receptors (A129) and those lacking IFN-γ (G129), mice that lacked both the type I and II IFN receptors, AG129 mice (Table 1), were developed in the 1990’s and these mice have become a mainstay of dengue research [104]. Johnson et al. [105] first tested the utility of AG129 mice in DENV vaccine trials. Their study reported that AG129 mice infected with mouse-adapted DENV-2 succumbed to death regardless of age, while immunized mice survived the virus challenge, and survival times increased following passive transfer of the anti-DENV polyclonal antibody. These successful results suggested the utility of AG129 in testing vaccines, as well as antivirals [105]. AG129 mice are now being used extensively in dengue vaccine and antiviral studies [106,107]. C57BL/6 mice lacking only type I IFN receptors succumb to death, even when infected with low doses of a lethal strain of DENV-2 (D220), which was generated by alternatively passaging the virus between C6/36 cells and AG129 mice. C57BL/6 mice lacking only type I IFN receptors in combination with DENV-2 (D220) are being considered as an improved model to study DENV infection and disease and for testing vaccines and antivirals, even under antibody-enhanced infection conditions [108]. Studies on mice with the individual deletion of IFNs (type I & II receptors-IFN-α/β and -γ) and STAT genes have highlighted the role of the STAT-1 and STAT-2 protein in DENV biology [109,110]. Mice lacking either STAT-1 or STAT-2 infected with DENV possess a higher level of viral RNA compared to wild-type mice, but they survive. However, in mice knocked out for both STAT-1 and STAT-2, early death was observed. This demonstrated the role of the STAT2-dependent pathway in mediating transcription of the interferon-stimulated genes against DENV in the absence of STAT-1 [111]. Mice lacking IFN response factors (IRF)-3 and -7 have been shown to be infected with DENV by the bite of infectious mosquitoes; infected mice were also able to transmit the virus to mosquitoes. Hence, this model is being used to understand the transmission dynamics of DENV [112]. Triple KO mice lacking IRF-3, -5, and -7, as well as quadruplet KO mice lacking IRF-1, -3, -5, and -7, have been used to identify the role of IRF-1 in inducing IFN-γ, as well type I IFN, responses against DENV [113]. Moreover, quadruplet KO mice-based experiments have revealed the role of this alternative pathway involving IRF-1 in resistance to DENV [113]. The pathogenic role of CCR5, a chemokine receptor, in supporting viral infection and replication in murine macrophages has been demonstrated in CCR5 KO mice [114]. Even though the use of KO mice has encountered disadvantages, it has contributed enormously to demonstrating the involvement of several cellular signalling pathways in DENV infection.
4.2. Transgenic Mouse Models of DENV
Transgenic mice over-expressing human tumor necrosis factor (TNF)-α have been utilized to demonstrate the role of TNF-α in the development of DENV encephalitis-like symptoms and neurotoxicity. Moreover, anti TNF-α antibodies reduced dengue encephalitis and mortality, suggesting anti TNF-α-based therapeutics for dengue encephalitis [115]. Since mice lacking the IFN-α/β receptor are susceptible to DENV infection and demonstrate T cell responses against DENV infection, studying T cell responses in the context of human MHC molecules might help identify T cell epitopes that are recognized by human T cells [110,116,117,118]. IFNα/βR−/− Tg mice (Table 2) expressing human MHC alleles such as HLA, A*0201, A*0101, A*1101, B*0701, and DRB1*0101 have been used to determine the T cell response against DENV. About 42 T cell epitopes have been identified, and most are also recognized by PBMCs from humans. Therefore, Tg mice expressing human MHC alleles might help in identifying T cell epitopes relevant to vaccine design [110].
4.3. Humanized Mouse Models of DENV
Human cell responses to DENV infection can be studied better in humanized mouse models since these models provide the opportunity to measure the contribution of host factors in an in vivo system. NOD/SCID mice transplanted with human cord blood hematopoietic progenitor (CD34+) cells and infected with DENV mimicking natural infection conditions have been used to demonstrate the clinical symptoms of dengue with a fever, rash, and thrombocytopenia [118]. Antibody responses mimicking primary dengue infection have been demonstrated in RAG2−/−γc−/− mice that were xenografted with human CD34+ hematopoietic stem cells [119]. A humanized NOD-SCID IL2rγ null mice (Table 3) model has been used to demonstrate differences in the virulence of different DENV-2 genotype strains. Southeast Asian strains produced the highest viremia, followed by Indian strains and Sylvatic strains [120]. An improved humoral response, HLA-A*02-restricted T cell response, and associated IFN-γ production were observed in BLT-NSG mice, but not in NSG mice. Therefore, BLT-NSB mice are an attractive platform for assessing human immune responses to the DENV vaccine [121,122]. However, humanized mice models predominantly develop IgM in response to DENV infection; only a few developed an IgG response because of inefficient class switching, and hence the kinetics of the antibody response was slow [119,123]. Research efforts are expected to lead to the development of ideal models that will show both IgM and IgG responses for studying dengue pathogenesis [124]. The serum metabolomic profile of DENV-2 infected humanized mice has also been utilized to identify the prognostic markers to predict severe dengue. The results yielded a profile similar to those reported in human longitudinal studies [125]. Human immune system (HIS) BLT-NOD/SCID mice have also been demonstrated to be useful in preclinical testing of the efficacy of antiviral drugs against dengue [126]. The following future aspects can be aimed at with the observed results: (i) to test the safety of the DENV vaccine in the recipient without pathology [120,127]; and (ii) to test the immunogenicity of the DENV vaccine [126,128,129].
5. Conclusions
Indeed, it is not easy to establish a small animal model for research purposes. An ideal animal model for human viral disease should closely recapitulate the spectrum of clinical symptoms and pathogenesis seen during the course of human infection. Moreover, a perfect animal model should meet other criteria, viz., maximum permissibility to the pathogens, nil resistance to the pathogens, subservience to the same human pathogenic strain, and having the route of infection similar to the human condition. Fulfilling the “Animal rule” of the US Food and Drug Administration [130] is essential for approval and optimally characterized animal models are critical. This rule suits conditions where vaccines and therapeutics cannot be tested safely or ethically on humans; in these cases, approval is possible only after preclinical tests are conducted on animal models. Research in many fields in virology is focussing on standardized models that meet the institutional requirements to evaluate the efficacy of vaccines and therapeutics. In this manuscript, we have summarized available mouse models, such as knockout mice, transgenic mice, and humanized mice models, for HIV-1, influenza, and dengue viruses, and discussed their applications in research. The characters of an ideal animal model cannot always be fulfilled. Since viruses frequently change, mice models need to be changed accordingly, otherwise the natural course of infection will not be accurately mirrored in the animal model. Humanized mice, (CD34+-HSC transplanted immunodeficient mice and BLT mice, NSG, NRG, NOD-SCID) are great models, allowing researchers to recapitulate the key aspects of viral pathogenesis in humans by recreating functional human immune systems in mice. Humanized mouse models are serving as important research tools to experimentally characterize human viral diseases and immune responses. Though humanized mice have advantages over other mouse models, the limitations include: (i) Mouse-to-mouse species variation during the development of a humanized mouse is frequently observed, leading to difficulties in interpreting the results at the end of the experiment; (ii) development of the molecules and cells is not complete in mouse–human chimeric models, and this incompleteness may interrupt the interactions between the cells; (iii) the mechanism of viral pathogenesis is also differently regulated in humanized mice, for example, with regard to B-cell maturation, antibody production, and the coagulation cascade, which are not regulated in the same way [44,130,131,132,133]; (iv) drawbacks include the substantial cost and limited supply and handling of animals in comparison to other models. Despite these limitations, recent developments involving transgenic humanized mice expressing different human genes regulating cellular development are also being attempted [16,18]. Moreover, in traditional KO and Tg mice models, the resulting phenotype due to the knocked out gene or transgenes is expressed during embryonic development itself, and disease onset in these mice models might occur earlier than in humans. To tackle this issue, researchers have developed mice models that can be made to express transgenes when needed using the Cre/lox site specific recombination system. This method provides the opportunity to control gene expression in time and space. These mouse strains express the Cre recombinase enzyme under the control of a promoter that can be induced by drugs such as tetracycline or doxycycline. Then, Cre can activate the expression of transgenes that are flanked by Lox P sites [133,134,135]. Such conditional KO and Tg mice are available for studying cancer and need to be developed for studying viral diseases. With recent advances in genomics, complete information about the mouse genome will serve as an important resource for developing new mice models that will help to completely understand the disease pathogenesis mechanisms and might lead to the discovery of novel therapeutic interventions and prophylactic approaches.
Funding
This work was supported by grants of HMR 14150811, HMRF/ 15163481 and GRF/HKBU 12101417 from Hong Kong Government, and also partly by grants from RC-IRCs/17-18/03 from Hong Kong Baptist University.
Acknowledgments
We thank Martha Dahlen for her critical review, English editing and revision.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Global HIV and AIDS Statistics-2018 Fact Sheet. Available online: http://www.unaids.org/en/resources/fact-sheet (accessed on 28 August 2018).
- Nair, H.; Brooks, W.A.; Katz, M.; Roca, A.; Berkley, J.A.; Madhi, S.A.; Simmerman, J.M.; Gordon, A.; Sato, M.; Howie, S.; et al. Global burden of respiratory infections due to seasonal influenza in young children: A systematic review and meta-analysis. Lancet 2011, 378, 1917–1930. [Google Scholar] [CrossRef]
- Barrila, J.; Radtke, A.L.; Crabbé, A.; Sarker, S.F.; Herbst-Kralovetz, M.M.; Ott, C.M.; Nickerson, C.A. Organotypic 3D cell culture models: Using the rotating wall vessel to study host-pathogen interactions. Nat. Rev. Microbiol. 2010, 8, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Bot, A.; Casares, S.; Bot, S.; Von Boehmer, H.; Bona, C. Cellular mechanisms involved in protection against influenza virus infection in transgenic mice expressing a TCR receptor specific for class II hemagglutinin peptide in CD4+ and CD8+ T cells. J. Immunol. 1998, 160, 4500–4507. [Google Scholar] [PubMed]
- Kirberg, J.; Baron, A.; Jakob, S.; Rolink, A.; Karjalainen, K.; Von Boehmer, H. Thymic selection of CD8+ single positive cells with a class II major histocompatibility complex-restricted receptor. J. Exp. Med. 1994, 180, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rall, G.F.; Lawrence, D.M.; Patterson, C.E. The application of transgenic and knockout mouse technology for the study of viral pathogenesis. Virology 2000, 271, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Mombaerts, P.; Iacomini, J.; Johnson, R.S.; Herrup, K.; Tonegawa, S.; Papaioannou, V.E. RAG-1-deficient mice have no mature B and T lymphocytes. Cell 1992, 68, 869–877. [Google Scholar] [CrossRef]
- Shinkai, Y.; Rathbun, G.; Lam, K.P.; Oltz, E.M.; Stewart, V.; Mendelsohn, M.; Charron, J.; Datta, M.; Young, F.; Stall, A.M.; et al. RAG-2-deficient mice lack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell 1992, 68, 855–867. [Google Scholar] [CrossRef]
- Hwang, S.Y.; Hertzog, P.J.; Holland, K.A.; Sumarsono, S.H.; Tymms, M.J.; Hamilton, J.A.; Whitty, G.; Bertoncello, I.; Kola, I. A null mutation in the gene encoding a type I interferon receptor component eliminates antiproliferative and antiviral responses to interferons alpha and beta and alters macrophage responses. Proc. Natl. Acad. Sci. USA 1995, 92, 11284–11288. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [Green Version]
- Meraz, M.A.; White, J.M.; Sheehan, K.C.; Bach, E.A.; Rodig, S.J.; Dighe, A.S.; Kaplan, D.H.; Riley, J.K.; Greenlund, A.C.; Campbell, D.; et al. Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell 1996, 84, 431–442. [Google Scholar] [CrossRef]
- Matsuyama, T.; Kimura, T.; Kitagawa, M.; Pfeffer, K.; Kawakami, T.; Watanabe, N.; Kundig, T.M.; Amakawa, R.; Kishihara, K.; Wakeham, A.; et al. Targeted disruption of IRF-1 or IRF-2 results in abnormal type I IFN gene induction and aberrant lymphocyte development. Cell 1993, 75, 83–97. [Google Scholar] [CrossRef]
- Kennedy, M.K.; Glaccum, M.; Brown, S.N.; Butz, E.A.; Viney, J.L.; Embers, M.; Matsuki, N.; Charrier, K.; Sedger, L.; Willis, C.R.; et al. Reversible defects in natural killer and memory CD8 T cell lineages in interleukin 15-deficient mice. J. Exp. Med. 2000, 91, 771–780. [Google Scholar] [CrossRef]
- Theocharides, A.P.; Rongvaux, A.; Fritsch, K.; Flavell, R.A.; Manz, M.G. Humanized hemato-lymphoid system mice. Haematologica 2016, 101, 5–19. [Google Scholar] [CrossRef]
- Ishikawa, F.; Yasukawa, M.; Lyons, B.; Yoshida, S.; Miyamoto, T.; Yoshimoto, G.; Watanabe, T.; Akashi, K.; Shultz, L.D.; Harada, M. Development of functional human blood and immune systems in NOD/SCID/IL2 receptor γ chainnull mice. Blood 2005, 106, 1565–1573. [Google Scholar] [CrossRef]
- Lai, F.; Chen, Q. Humanized Mouse Models for the Study of Infection and Pathogenesis of Human Viruses. Viruses 2018, 10, 643. [Google Scholar] [CrossRef]
- Brehm, M.A.; Bortell, R.; Verma, M.; Shultz, L.D.; Greiner, D.L. Humanized mice in translational immunology. Transl. Immunol. Mech. Pharmacol. Approaches 2016, 285–326. [Google Scholar]
- Shultz, L.D.; Brehm, M.A.; Garcia-Martinez, J.V.; Greiner, D.L. Humanized mice for immune system investigation: Progress, promise and challenges. Nat. Rev. Immunol. 2012, 12, 786–798. [Google Scholar] [CrossRef]
- Walsh, N.; Kenney, L.; Jangalwe, S.; Aryee, K.E.; Greiner, D.L.; Brehm, M.A.; Shultz, L.D. Humanized mouse models of clinical disease. Annu. Rev. Pathol. 2017, 12, 187–215. [Google Scholar] [CrossRef]
- King, M.A.; Covassin, L.; Brehm, M.A.; Racki, W.; Pearson, T.; Leif, J.; Laning, J.; Fodor, W.; Foreman, O.; Burzenski, L.; et al. Hu-PBL-NOD-SCID IL2rgnull mouse model of xenogeneic graft-versus-host-like disease and the role of host MHC. Clin. Exp. Immunol. 2009, 157, 104–118. [Google Scholar] [CrossRef]
- Melkus, M.W.; Estes, J.D.; Padgett-Thomas, A.; Gatlin, J.; Denton, P.W.; Othieno, F.A.; Wege, A.K.; Haase, A.T.; Garcia, J.V. Humanized mice mount specific adaptive and innate immune responses to EBV and TSST-1. Nat. Med. 2006, 12, 1316–1322. [Google Scholar] [CrossRef]
- Hatziioannou, T.; Evans, D.T. Animal models for HIV/AIDS research. Nat. Rev. Microbiol. 2012, 10, 852–867. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, S.; Behrendt, R.; Eissmann, K.; Volkmann, B.; Thomas, D.; Ebert, T.; Cribier, A.; Benkirane, M.; Hornung, V.; Bouzas, N.F.; et al. Phosphorylation of murine SAMHD1 regulates its antiretroviral activity. Retrovirology 2015, 12, 103. [Google Scholar] [CrossRef]
- Rehwinkel, J.; Maelfait, J.; Bridgeman, A.; Rigby, R.; Hayward, B.; Liberatore, R.A.; Bieniasz, P.D.; Towers, G.J.; Moita, L.F.; Crow, Y.J.; et al. SAMHD1-dependent retroviral control and escape in mice. EMBO J. 2013, 32, 2454–2462. [Google Scholar] [CrossRef] [Green Version]
- Behrendt, R.; Schumann, T.; Gerbaulet, A.; Nguyen, L.A.; Schubert, N.; Alexopoulou, D.; Berka, U.; Lienenklaus, S.; Peschke, K.; Gibbert, K.; et al. Mouse SAMHD1 has antiretroviral activity and suppresses a spontaneous cell-intrinsic antiviral response. Cell Rep. 2013, 4, 689–696. [Google Scholar] [CrossRef]
- Okeoma, C.M.; Lovsin, N.; Peterlin, B.M.; Ross, S.R. APOBEC3 inhibits mouse mammary tumour virus replication in vivo. Nature 2007, 445, 927. [Google Scholar] [CrossRef]
- Saitoh, T.; Satoh, T.; Yamamoto, N.; Uematsu, S.; Takeuchi, O.; Kawai, T.; Akira, S. Antiviral protein Viperin promotes Toll-like receptor 7-and Toll-like receptor 9-mediated type I interferon production in plasmacytoid dendritic cells. Immunity 2011, 34, 352–363. [Google Scholar] [CrossRef]
- Swiecki, M.; Wang, Y.; Gilfillan, S.; Lenschow, D.J.; Colonna, M. Cutting edge: Paradoxical roles of BST2/tetherin in promoting type I IFN response and viral infection. J. Immunol. 2012, 1103145. [Google Scholar] [CrossRef]
- Bloch, N.; Glasker, S.; Sitaram, P.; Hofmann, H.; Shepard, C.N.; Schultz, M.L.; Kim, B.; Landau, N.R. A highly active isoform of lentivirus restriction factor SAMHD1 in mouse. J. Biol. Chem. 2017, 292, 1068–1080. [Google Scholar] [CrossRef]
- Browning, J.; Horner, J.W.; Pettoello-Mantovani, M.; Raker, C.; Yurasov, S.; DePinho, R.A.; Goldstein, H. Mice transgenic for human CD4 and CCR5 are susceptible to HIV infection. Proc. Natl. Acad. Sci. USA 1997, 94, 14637–14641. [Google Scholar] [CrossRef] [Green Version]
- Seay, K.; Qi, X.; Zheng, J.H.; Zhang, C.; Chen, K.; Dutta, M.; Deneroff, K.; Ochsenbauer, C.; Kappes, J.C.; Littman, D.R.; et al. Mice transgenic for CD4-specific human CD4, CCR5 and cyclin T1 expression: A new model for investigating HIV-1 transmission and treatment efficacy. PLoS ONE 2013, 8, 63537. [Google Scholar] [CrossRef]
- Keppler, O.T.; Welte, F.J.; Ngo, T.A.; Chin, P.S.; Patton, K.S.; Tsou, C.L.; Abbey, N.W.; Sharkey, M.E.; Grant, R.M.; You, Y.; et al. Progress toward a human CD4/CCR5 transgenic rat model for de novo infection by human immunodeficiency virus type 1. J. Exp. Med. 2002, 195, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.M.; Abramczuk, J.W.; Pezen, D.S.; Rutledge, R.; Belcher, J.H.; Hakim, F.; Shearer, G.; Lamperth, L.; Travis, W.; Fredrickson, T. Development of disease and virus recovery in transgenic mice containing HIV proviral DNA. Science 1988, 242, 1665–1670. [Google Scholar] [CrossRef] [PubMed]
- Dickie, P.; Gazzinelli, R.; Chang, L.J. Models of HIV type 1 proviral gene expression in wild-type HIV and MLV/HIV transgenic mice. AIDS Res. Hum. Retrovir. 1996, 12, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Dickie, P.; Ramsdell, F.; Notkins, A.L.; Venkatesan, S. Spontaneous and inducible epidermal hyperplasia in transgenic mice expressing HIV-1 Nef. Virology 1993, 197, 431–438. [Google Scholar] [CrossRef]
- Hanna, Z.; Kay, D.G.; Cool, M.; Jothy, S.; Rebai, N.; Jolicoeur, P. Transgenic mice expressing human immunodeficiency virus type 1 in immune cells develop a severe AIDS-like disease. J. Virol. 1998, 72, 121–132. [Google Scholar] [PubMed]
- Rahim, M.M.A.; Chrobak, P.; Hu, C.; Hanna, Z.; Jolicoeur, P. Adult AIDS-like disease in a novel inducible human immunodeficiency virus type 1 Nef transgenic mouse model: CD4+ T-cell activation is Nef dependent and can occur in the absence of lymphophenia. J. Virol. 2009, 83, 11830–11846. [Google Scholar] [CrossRef] [PubMed]
- Hanna, Z.; Weng, X.; Kay, D.G.; Poudrier, J.; Lowell, C.; Jolicoeur, P. The pathogenicity of human immunodeficiency virus (HIV) type 1 Nef in CD4C/HIV transgenic mice is abolished by mutation of its SH3-binding domain, and disease development is delayed in the absence of Hck. J. Virol. 2001, 75, 9378–9392. [Google Scholar] [CrossRef] [PubMed]
- Hanna, Z.; Priceputu, E.; Kay, D.G.; Poudrier, J.; Chrobak, P.; Jolicoeur, P. In vivo mutational analysis of the N-terminal region of HIV-1 Nef reveals critical motifs for the development of an AIDS-like disease in CD4C/HIV transgenic mice. Virology 2004, 327, 273–286. [Google Scholar] [CrossRef]
- Thaney, V.E.; Sanchez, A.B.; Fields, J.A.; Minassian, A.; Young, J.W.; Maung, R.; Kaul, M. Transgenic mice expressing HIV-1 envelope protein gp120 in the brain as an animal model in neuroAIDS research. J. Neurovirol. 2018, 1, 1–2. [Google Scholar] [CrossRef]
- Maung, R.; Hoefer, M.M.; Sanchez, A.B.; Sejbuk, N.E.; Medders, K.E.; Desai, M.K.; Catalan, I.C.; Dowling, C.C.; De Rozieres, C.M.; Garden, G.A.; et al. CCR5 knockout prevents neuronal injury and behavioral impairment induced in a transgenic mouse model by a CXCR4-using HIV-1 glycoprotein 120. J. Immunol. 2014, 1302915. [Google Scholar] [CrossRef]
- Putatunda, R.; Zhang, Y.; Li, F.; Yang, X.F.; Barbe, M.F.; Hu, W. Adult neurogenic deficits in HIV-1 Tg26 transgenic mice. J. Neuroinflamm. 2018, 1, 287. [Google Scholar] [CrossRef]
- Denton, P.W.; Sogaard, O.S.; Tolstrup, M. Using animal models to overcome temporal, spatial and combinatorial challenges in HIV persistence research. J. Transl. Med. 2016, 14, 44. [Google Scholar] [CrossRef]
- Akkina, R. New generation humanized mice for virus research: Comparative aspects and future prospects. Virology 2013, 435, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, S.K.; Archin, N.M.; Cheema, M.; Dahl, N.P.; Garcia, J.V.; Margolis, D.M. Latent HIV-1 infection of resting CD4+ T cells in the humanized Rag2−/− γc−/− mouse. J. Virol. 2012, 86, 114–120. [Google Scholar] [CrossRef]
- Marsden, M.D.; Kovochich, M.; Suree, N.; Shimizu, S.; Mehta, R.; Cortado, R.; Bristol, G.; An, D.S.; Zack, J.A. HIV latency in the humanized BLT mouse. J. Virol. 2012, 86, 339–347. [Google Scholar] [CrossRef]
- Honeycutt, J.B.; Sheridan, P.A.; Matsushima, G.K.; Garcia, J.V. Humanized mouse models for HIV-1 infection of the CNS. J. Neurovirol. 2015, 21, 301–309. [Google Scholar] [CrossRef]
- Olesen, R.; Swanson, M.D.; Kovarova, M.; Nochi, T.; Chateau, M.; Honeycutt, J.B.; Long, J.M.; Denton, P.W.; Hudgens, M.G.; Richardson, A.; et al. ART influences HIV persistence in the female reproductive tract and cervicovaginal secretions. J. Clin. Investig. 2016, 126, 892–904. [Google Scholar] [CrossRef] [Green Version]
- Lavender, K.J.; Pace, C.; Sutter, K.; Messer, R.J.; Pouncey, D.L.; Cummins, N.W.; Natesampillai, S.; Zheng, J.; Goldsmith, J.; Widera, M.; et al. An advanced BLT-humanized mouse model for extended HIV-1 cure studies. Aids 2018, 32, 1–10. [Google Scholar] [CrossRef]
- Cohen, M.S.; Chen, Y.Q.; McCauley, M.; Gamble, T.; Hosseinipour, M.C.; Kumarasamy, N.; Hakim, J.G.; Kumwenda, J.; Grinsztejn, B.; Pilotto, J.H.; et al. Prevention of HIV-1 infection with early antiretroviral therapy. N. Engl. J. Med. 2011, 365, 493–505. [Google Scholar] [CrossRef]
- Brooks, D.G.; Kitchen, S.G.; Kitchen, C.M.; Scripture-Adams, D.D.; Zack, J.A. Generation of HIV latency during thymopoiesis. Nat. Med. 2001, 7, 459–464. [Google Scholar] [CrossRef]
- Brehm, M.A.; Wiles, M.V.; Greiner, D.L.; Shultz, L.D. Generation of improved humanized mouse models for human infectious diseases. J. Immunol. 2014, 410, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Karpel, M.E.; Boutwell, C.L.; Allen, T.M. BLT humanized mice as a small animal model of HIV infection. Curr. Opin. Virol. 2015, 13, 75–80. [Google Scholar] [CrossRef]
- Akkina, R.; Allam, A.; Balazs, A.B.; Blankson, J.N.; Burnett, J.C.; Casares, S.; Garcia, J.V.; Hasenkrug, K.J.; Kashanchi, F.; Kitchen, S.G.; et al. Improvements and limitations of humanized mouse models for HIV research: NIH/NIAID “meet the experts” 2015 workshop summary. AIDS Res. Hum. Retrovir. 2016, 32, 109–119. [Google Scholar] [CrossRef]
- Horwitz, J.A.; Halper-Stromberg, A.; Mouquet, H.; Gitlin, A.D.; Tretiakova, A.; Eisenreich, T.R.; Malbec, M.; Gravemann, S.; Billerbeck, E.; Dorner, M.; et al. HIV-1 suppression and durable control by combining single broadly neutralizing antibodies and antiretroviral drugs in humanized mice. Proc. Natl. Acad. Sci. USA 2013, 110, 16538–16543. [Google Scholar] [CrossRef] [Green Version]
- Kovochich, M.; Marsden, M.D.; Zack, J.A. Activation of latent HIV using drug-loaded nanoparticles. PLoS ONE 2011, 6, e18270. [Google Scholar] [CrossRef]
- Denton, P.W.; Olesen, R.; Choudhary, S.K.; Archin, N.M.; Wahl, A.; Swanson, M.D.; Chateau, M.; Nochi, T.; Krisko, J.F.; Spagnuolo, R.A.; et al. Generation of HIV latency in humanized BLT mice. J. Virol. 2012, 86, 630–634. [Google Scholar] [CrossRef]
- Arainga, M.; Edagwa, B.; Mosley, R.L.; Poluektova, L.Y.; Gorantla, S.; Gendelman, H.E. A mature macrophage is a principal HIV-1 cellular reservoir in humanized mice after treatment with long acting antiretroviral therapy. Retrovirology 2017, 14, 17. [Google Scholar] [CrossRef]
- Honeycutt, J.B.; Wahl, A.; Archin, N.; Choudhary, S.; Margolis, D.; Garcia, J.V. HIV-1 infection, response to treatment and establishment of viral latency in a novel humanized T cell-only mouse (TOM) model. Retrovirology 2013, 10, 121. [Google Scholar] [CrossRef]
- Honeycutt, J.B.; Thayer, W.O.; Baker, C.E.; Ribeiro, R.M.; Lada, S.M.; Cao, Y.; Cleary, R.A.; Hudgens, M.G.; Richman, D.D.; Garcia, J.V. HIV persistence in tissue macrophages of humanized myeloid-only mice during antiretroviral therapy. Nat. Med. 2017, 23, 638–643. [Google Scholar] [CrossRef]
- Metcalf Pate, K.A.; Pohlmeyer, C.W.; Walker-Sperling, V.E.; Foote, J.B.; Najarro, K.M.; Cryer, C.G.; Salgado, M.; Gama, L.; Engle, E.L.; Shirk, E.N.; et al. A murine viral outgrowth assay to detect residual HIV type 1 in patients with undetectable viral loads. J. Infect. Dis. 2015, 212, 1387–1396. [Google Scholar] [CrossRef]
- Yuan, Z.; Kang, G.; Lu, W.; Li, Q. Reactivation of HIV-1 proviruses in immune-compromised mice engrafted with human VOA- negative CD4+ T cells. J. Virus Erad. 2017, 3, 61–65. [Google Scholar]
- Satheesan, S.; Li, H.; Burnett, J.C.; Takahashi, M.; Li, S.; Wu, S.X.; Synold, T.W.; Rossi, J.J.; Zhou, J. HIV replication and latency in a humanized NSG mouse model during suppressive oral combinational ART. J. Virol. 2018, JVI-02118. [Google Scholar]
- WHO: Influenza (Seasonal). Available online: https://www.who.int/en/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 12 October 2018).
- Mallia, P.; Johnston, S.L. Influenza infection and COPD. Int. J. Chron Obstruct. Pulmon. Dis. 2007, 2, 55. [Google Scholar] [CrossRef]
- Kim, M.H.; Song, W.J.; Yang, M.S.; Lee, S.H.; Kwon, J.W.; Kim, S.H.; Kang, H.R.; Park, H.W.; Cho, Y.J.; Cho, S.H.; et al. Clinical course of asthma patients with H1N1 influenza infection and oseltamivir. Minerva Med. 2018, 109, 7–14. [Google Scholar]
- Barnes, M.; Heywood, A.E.; Mahimbo, A.; Rahman, B.; Newall, A.T.; Macintyre, C.R. Acute myocardial infarction and influenza: A meta-analysis of case–control studies. Heart 2015, 101, 1738–1747. [Google Scholar] [CrossRef]
- Stanwell-Smith, R.; Parker, A.M.; Chakraverty, P.; Soltanpoor, N.; Simpson, C.N. Possible association of influenza A with fetal loss: Investigation of a cluster of spontaneous abortions and stillbirths. Commun. Dis. Rep. CDR Rev. 1994, 4, 28–32. [Google Scholar]
- Palese, P. Influenza: Old and new threats. Nat. Med. 2004, 10, S82. [Google Scholar] [CrossRef]
- Horimoto, T.; Kawaoka, Y. Pandemic threat posed by avian influenza A viruses. Clin. Microbiol. Rev. 2001, 14, 129–149. [Google Scholar] [CrossRef]
- Guarnaccia, T.; Carolan, L.A.; Maurer-Stroh, S.; Lee, R.T.; Job, E.; Reading, P.C.; Petrie, S.; McCaw, J.M.; McVernon, J.; Hurt, A.C.; et al. Antigenic drift of the pandemic 2009 A (H1N1) influenza virus in A ferret model. PLoS Pathog. 2013, 9, 1003354. [Google Scholar] [CrossRef]
- Margine, I.; Krammer, F. Animal models for influenza viruses: Implications for universal vaccine development. Pathogens 2014, 3, 845–874. [Google Scholar] [CrossRef]
- Blazejewska, P.; Koscinski, L.; Viegas, N.; Anhlan, D.; Ludwig, S.; Schughart, K. Pathogenicity of different PR8 influenza A virus variants in mice is determined by both viral and host factors. Virology 2011, 412, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, B.; Błażejewska, P.; Heßmann, M.; Bruder, D.; Geffers, R.; Mauel, S.; Gruber, A.D.; Schughart, K. Host genetic background strongly influences the response to influenza a virus infections. PLoS ONE 2009, 4, e4857. [Google Scholar] [CrossRef]
- Schmitz, N.; Kurrer, M.; Bachmann, M.F.; Kopf, M. Interleukin-1 is responsible for acute lung immunopathology but increases survival of respiratory influenza virus infection. J. Virol. 2005, 79, 6441–6448. [Google Scholar] [CrossRef]
- Liu, B.; Mori, I.; Hossain, M.J.; Dong, L.; Takeda, K.; Kimura, Y. Interleukin-18 improves the early defence system against influenza virus infection by augmenting natural killer cell-mediated cytotoxicity. J. Gen. Virol. 2004, 85, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.G.; Dash, P.; Aldridge, J.R., Jr.; Ellebedy, A.H.; Reynolds, C.; Funk, A.J.; Martin, W.J.; Lamkanfi, M.; Webby, R.J.; Boyd, K.L.; et al. NLRP3 (NALP3/CIAS1/Cryopyrin) mediates key innate and healing responses to influenza A virus via the regulation of caspase-1. Immunity. 2009, 30, 566. [Google Scholar] [CrossRef]
- Perrone, L.A.; Szretter, K.J.; Katz, J.M.; Mizgerd, J.P.; Tumpey, T.M. Mice lacking both TNF and IL-1 receptors exhibit reduced lung inflammation and delay in onset of death following infection with a highly virulent H5N1 virus. J. Infect. Dis. 2010, 202, 1161–1170. [Google Scholar] [CrossRef]
- Gally, F.; Kosmider, B.; Weaver, M.R.; Pate, K.M.; Hartshorn, K.L.; Oberley-Deegan, R.E. FABP5 deficiency enhances susceptibility to H1N1 influenza A virus-induced lung inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, 64–72. [Google Scholar] [CrossRef]
- Huang, C.H.; Chen, C.J.; Yen, C.T.; Yu, C.P.; Huang, P.N.; Kuo, R.L.; Lin, S.J.; Chang, C.K.; Shih, S.R. Caspase-1 deficient mice are more susceptible to influenza A virus infection with PA variation. J. Infect. Dis. 2013, 208, 1898–1905. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe, S.; Maher, E.A.; Neumann, G.; Kawaoka, Y. Pandemic potential of avian influenza A (H7N9) viruses. Trends Microbiol. 2014, 22, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G.; Kawaoka, Y. Transmission of influenza A viruses. Virology 2015, 479, 234–246. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Deeg, C.M.; Hassan, E.; Mutz, P.; Rheinemann, L.; Gotz, V.; Magar, L.; Schilling, M.; Kallfass, C.; Nurnberger, C.; Soubies, S.; et al. In vivo evasion of MxA by avian influenza viruses requires human signature in the viral nucleoprotein. J. Exp. Med. 2017, 214, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, C.; Yang, Z.; Chi, X.; Zhang, J.; Chen, J.L. Targeted disruption of influenza A virus hemagglutinin in genetically modified mice reduces viral replication and improves disease outcome. Sci. Rep. 2016, 6, 23746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Mario, G.; Garulli, B.; Sciaraffia, E.; Facchini, M.; Donatelli, I.; Castrucci, M.R. A heat-inactivated H7N3 vaccine induces cross-reactive cellular immunity in HLA-A2. 1 transgenic mice. Virol. J. 2016, 13, 56. [Google Scholar] [CrossRef] [PubMed]
- Di Mario, G.; Sciaraffia, E.; Facchini, M.; Gubinelli, F.; Soprana, E.; Panigada, M.; Bernasconi, V.; Garulli, B.; Siccardi, A.; Donatelli, I.; et al. Protective immunity against influenza in HLA-A2 transgenic mice by modified vaccinia virus Ankara vectored vaccines containing internal influenza proteins. Pathog. Glob. Health 2017, 111, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moise, L.; Tassone, R.; Latimer, H.; Terry, F.; Levitz, L.; Haran, J.P.; Ross, T.M.; Boyle, C.; Martin, W.D.; De Groot, A.S. Immunization with cross-conserved H1N1 influenza CD4+ T-cell epitopes lowers viral burden in HLA DR3 transgenic mice. Hum. Vaccin Immunother. 2013, 9, 2060–2068. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.M.; Poon, L.L.; Guan, Y. Emergence of a novel swine-origin influenza A virus (S-OIV) H1N1 virus in humans. J. Clin. Virol. 2009, 45, 169–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konigshofer, Y.; Chien, Y.H. γδ T cells—Innate immune lymphocytes. Curr. Opin. Immunol. 2006, 18, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Eberl, M.; Moser, B. Monocytes and γδ T cells: Close encounters in microbial infection. Trends Immunol. 2009, 30, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Bonneville, M.; O’brien, R.L.; Born, W.K. γδ T cell effector functions: A blend of innate programming and acquired plasticity. Nat. Rev. Immunol. 2010, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Tu, W.W.; Lau, Y.L.; Peiris, J.S. Use of Humanized mice to study antiviral activity of human γδ. Hong Kong Med. J. 2014, 20, 4–6. [Google Scholar] [PubMed]
- Richards, K.A.; Topham, D.; Chaves, F.A.; Sant, A.J. Cutting edge: CD4 T cells generated from encounter with seasonal influenza viruses and vaccines have broad protein specificity and can directly recognize naturally generated epitopes derived from the live pandemic H1N1 virus. J. Immunol. 2010, 1001395. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Bernstein, D.I.; Winokur, P.; Rupp, R.; Anderson, E.; Rouphael, N.; Dickey, M.; Stapleton, J.T.; Edupuganti, S.; Spearman, P.; et al. Serological responses to an avian influenza A/H7N9 vaccine mixed at the point-of-use with MF59 adjuvant: A randomized clinical trial. JAMA 2014, 312, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Nithichanon, A.; Nobusawa, E.; Moise, L.; Martin, W.D.; Yamamoto, N.; Terahara, K.; Hagiwara, H.; Odagiri, T.; Tashiro, M.; et al. Hhumanized mouse model identifies key amino acids for low immunogenicity of H7N9 vaccines. Sci. Rep. 2017, 7, 1283. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, M.; Ballesteros, A.; Qiu, Q.; Pow Sang, L.; Shashikumar, S.; Casares, S.; Brumeanu, T.D. Generation and testing anti-influenza human monoclonal antibodies in a new humanized mouse model (DRAGA: HLA-A2. HLA-DR4. Rag1 KO. IL-2Rγc KO. NOD). Hum. Vaccin Immunother. 2018, 14, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, I.I.; Mihaylova, N.M.; Manoylov, I.K.; Makatsori, D.; Lolov, S.; Nikolova, M.H.; Mamalaki, A.; Prechl, J.; Tchorbanov, A.I. Targeting of Influenza Viral Epitopes to Antigen-Presenting Cells by Genetically Engineered Chimeric Molecules in a Humanized NOD SCID Gamma Transfer Model. Hum. Gene Ther. 2018, 29, 1056–1070. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, E.; Momose, H.; Hiradate, Y.; Furuhata, K.; Mizukami, T.; Hamaguchi, I. Development of a preclinical humanized mouse model to evaluate acute toxicity of an influenza vaccine. Oncotarget 2018, 9, 25751. [Google Scholar] [CrossRef] [PubMed]
- Alagarasu, K.; Bachal, R.V.; Bhagat, A.B.; Shah, P.S.; Dayaraj, C. Elevated levels of vitamin D and deficiency of mannose binding lectin in dengue hemorrhagic fever. Virol. J. 2012, 9, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecilia, D.; Kakade, M.; Alagarasu, K.; Patil, J.; Salunke, A.; Parashar, D.; Shah, P.S. Development of a multiplex real-time RT-PCR assay for simultaneous detection of dengue and chikungunya viruses. Arch. Virol. 2015, 160, 323–327. [Google Scholar] [CrossRef]
- Zompi, S.; Harris, E. Animal models of dengue virus infection. Viruses 2012, 4, 62–82. [Google Scholar] [CrossRef] [PubMed]
- Falzarano, D.; Bente, D.A. Animal models for viral hemorrhagic fever. Clin. Microbiol. Infect. 2014. [Google Scholar] [CrossRef]
- Van den Broek, M.F.; Muller, U.; Huang, S.; Aguet, M.; Zinkernagel, R.M. Antiviral defense in mice lacking both alpha/beta and gamma interferon receptors. J. Virol. 1995, 69, 4792–4796. [Google Scholar] [PubMed]
- Johnson, A.J.; Roehrig, J.T. New mouse model for dengue virus vaccine testing. J. Virol. 1999, 73, 783–786. [Google Scholar] [PubMed]
- Sarathy, V.V.; Milligan, G.N.; Bourne, N.; Barrett, A.D. Mouse models of dengue virus infection for vaccine testing. Vaccine 2015, 33, 7051–7060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milligan, G.N.; White, M.; Zavala, D.; Pyles, R.B.; Sarathy, V.V.; Barrett, A.D.; Bourne, N. Spectrum of activity testing for therapeutics against all four dengue virus serotypes in AG129 mouse models: Proof-of-concept studies with the adenosine nucleoside inhibitor NITD-008. Antivir. Res. 2018, 154, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Orozco, S.; Schmid, M.A.; Parameswaran, P.; Lachica, R.; Henn, M.R.; Beatty, R.; Harris, E. Characterization of a model of lethal dengue virus 2 infection in C57BL/6 mice deficient in the alpha/beta interferon receptor. J. Gen. Virol. 2012, 93, 2152–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, S.T.; Prestwood, T.R.; Lada, S.M.; Benedict, C.A.; Shresta, S. Cardif-mediated signaling controls the initial innate response to dengue virus in vivo. J. Virol. 2009, 83, 8276–8281. [Google Scholar] [CrossRef]
- Yauch, L.E.; Zellweger, R.M.; Kotturi, M.F.; Qutubuddin, A.; Sidney, J.; Peters, B.; Prestwood, T.R.; Sette, A.; Shresta, S. A protective role for dengue virus-specific CD8+ T cells. J. Immunol. 2009, 182, 4865–4873. [Google Scholar] [CrossRef]
- Perry, S.T.; Buck, M.D.; Lada, S.M.; Schindler, C.; Shresta, S. STAT2 mediates innate immunity to Dengue virus in the absence of STAT1 via the type I interferon receptor. PLoS Pathog. 2011, 7, 1001297. [Google Scholar] [CrossRef] [PubMed]
- Christofferson, R.C.; McCracken, M.K.; Johnson, A.M.; Chisenhall, D.M.; Mores, C.N. Development of a transmission model for dengue virus. J. Virol. 2013, 10, 127. [Google Scholar] [CrossRef]
- Carlin, A.F.; Plummer, E.M.; Vizcarra, E.A.; Sheets, N.; Joo, Y.; Tang, W.; Day, J.; Greenbaum, J.; Glass, C.K.; Diamond, M.S.; et al. An IRF-3-, IRF-5-, and IRF-7-independent pathway of dengue viral resistance Utilizes IRF-1 to stimulate type I and II interferon responses. Cell Rep. 2017, 21, 1600–1612. [Google Scholar] [CrossRef]
- Marques, R.E.; Guabiraba, R.; Del Sarto, J.L.; Rocha, R.F.; Queiroz, A.L.; Cisalpino, D.; Marques, P.E.; Pacca, C.C.; Fagundes, C.T.; Menezes, G.B.; et al. Dengue virus requires the CC-chemokine receptor CCR5 for replication and infection development. Immunology 2015, 145, 583–596. [Google Scholar] [CrossRef] [Green Version]
- Jhan, M.K.; HuangFu, W.C.; Chen, Y.F.; Kao, J.C.; Tsai, T.T.; Ho, M.R.; Shen, T.J.; Tseng, P.C.; Wang, Y.T.; Lin, C.F. Anti-TNF-α restricts dengue virus-induced neuropathy. J. Leukoc. Biol. 2018, 104, 961–968. [Google Scholar] [CrossRef]
- Weiskopf, D.; Yauch, L.E.; Angelo, M.A.; John, D.V.; Greenbaum, J.A.; Sidney, J.; Kolla, R.V.; De Silva, A.D.; de Silva, A.M.; Grey, H.; et al. Insights into HLA-restricted T cell responses in a novel mouse model of dengue virus infection point toward new implications for vaccine design. J. Immunol. 2011, 187, 4268. [Google Scholar] [CrossRef]
- Yauch, L.E.; Prestwood, T.R.; May, M.M.; Morar, M.M.; Zellweger, R.M.; Peters, B.; Sette, A.; Shresta, S. CD4+ T cells are not required for the induction of dengue virus-specific CD8+ T cell or antibody responses but contribute to protection after vaccination. J. Immunol. 2010, 1001709. [Google Scholar] [CrossRef]
- Bente, D.A.; Melkus, M.W.; Garcia, J.V.; Rico-Hesse, R. Dengue fever in humanized NOD/SCID mice. J. Virol. 2005, 79, 13797–13799. [Google Scholar] [CrossRef]
- Kuruvilla, J.G.; Troyer, R.M.; Devi, S.; Akkina, R. Dengue virus infection and immune response in humanized RAG2−/− γc−/−(RAG-hu) mice. Virology 2007, 369, 143–152. [Google Scholar] [CrossRef]
- Mota, J.; Rico-Hesse, R. Dengue virus tropism in humanized mice recapitulates human dengue fever. PLoS ONE 2011, 6, 20762. [Google Scholar] [CrossRef]
- Jaiswal, S.; Pazoles, P.; Woda, M.; Shultz, L.D.; Greiner, D.L.; Brehm, M.A.; Mathew, A. Enhanced humoral and HLA-A2-restricted dengue virus-specific T-cell responses in humanized BLT NSG mice. Immunology 2012, 136, 334–343. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, S.; Smith, K.; Ramirez, A.; Woda, M.; Pazoles, P.; Shultz, L.D.; Greiner, D.L.; Brehm, M.A.; Mathew, A. Dengue virus infection induces broadly cross-reactive human IgM antibodies that recognize intact virions in humanized BLT-NSG mice. Exp. Biol. Med. 2015, 240, 67–78. [Google Scholar] [CrossRef]
- Akkina, R. Humanized mice for studying human immune responses and generating human monoclonal antibodies. Microbiol. Spectr. 2014, 2, AID-0003-2012. [Google Scholar]
- Jangalwe, S.; Shultz, L.D.; Mathew, A.; Brehm, M.A. Improved B cell development in humanized NOD-SCID IL2Rγnull mice transgenically expressing human stem cell factor, granulocyte-macrophage colony-stimulating factor and interleukin-3. Immun. Inflamm. Dis. 2016, 4, 427–440. [Google Scholar] [CrossRef]
- Cui, L.; Hou, J.; Fang, J.; Lee, Y.H.; Costa, V.V.; Wong, L.H.; Chen, Q.; Ooi, E.E.; Tannenbaum, S.R.; Chen, J.; et al. Serum metabolomics investigation of humanized mouse model with dengue infection. J. Virol. 2017, 91, e00386-17. [Google Scholar] [CrossRef]
- Frias-Staheli, N.; Dorner, M.; Marukian, S.; Billerbeck, E.; Labitt, R.N.; Rice, C.M.; Ploss, A. Utility of humanized BLT mice for analysis of dengue virus infection and antiviral drug testing. J. Virol. 2014, 88, 2205–2218. [Google Scholar] [CrossRef]
- Mota, J.; Rico-Hesse, R. Humanized mice show clinical signs of dengue fever according to infecting virus genotype. J. Virol. 2009, 83, 8638–8645. [Google Scholar] [CrossRef]
- Cox, J.; Mota, J.; Sukupolvi-Petty, S.; Diamond, M.S.; Rico-Hesse, R. Mosquito bite delivery of dengue virus enhances immunogenicity and pathogenesis in humanized mice. J. Virol. 2012, 86, 7637–7649. [Google Scholar] [CrossRef]
- Züst, R.; Dong, H.; Li, X.F.; Chang, D.C.; Zhang, B.; Balakrishnan, T.; Toh, Y.X.; Jiang, T.; Li, S.H.; Deng, Y.Q.; et al. Rational design of a live attenuated dengue vaccine: 2′-o-methyltransferase mutants are highly attenuated and immunogenic in mice and macaques. PLoS Pathog. 2013, 9, e1003521. [Google Scholar] [CrossRef]
- Snoy, P.J. Establishing efficacy of human products using animals: The US food and drug administration’s “animal rule”. Vet. Pathol. 2010, 47, 774–778. [Google Scholar] [CrossRef]
- Mairuhu, A.T.; Mac Gillavry, M.R.; Setiati, T.E.; Soemantri, A.; Ten Cate, H.; Brandjes, D.P.; Van Gorp, E.C. Is clinical outcome of dengue-virus infections influenced by coagulation and fibrinolysis? A critical review of the evidence. Lancet Infect. Dis. 2003, 3, 33. [Google Scholar] [CrossRef]
- Lynch, S.F.; Ludlam, C.A. Plasma microparticles and vascular disorders. Br. J. Haematol. 2007, 137, 36. [Google Scholar] [CrossRef]
- Doyle, A.; McGarry, M.P.; Lee, N.A.; Lee, J.J. The construction of transgenic and gene knockout/knockin mouse models of human disease. Transgenic Res. 2012, 21, 327–349. [Google Scholar] [CrossRef]
- Ostrand-Rosenberg, S. Animal models of tumor immunity, immunotherapy and cancer vaccines. Curr. Opin. Immunol. 2004, 16, 143–150. [Google Scholar] [CrossRef]
- Bouabe, H.; Okkenhaug, K. Gene Targeting in Mice: A Review. Methods Mol. Biol. 2013, 1064, 315–336. [Google Scholar]
Figure 1.
Humanized mouse model represents the engraftment of human cells and tissues into immunodeficient mice. Humanized mouse model representing infection with HIV, FLU, or DENV.
Figure 1.
Humanized mouse model represents the engraftment of human cells and tissues into immunodeficient mice. Humanized mouse model representing infection with HIV, FLU, or DENV.

Figure 2.
Generation of transgenic (Tg) mice and knock out (KO) mice. The embryonic stem cells (ES) with inserted or disturbed specific gene of interest are microinjected into the mouse blastocyst, which allows the development of a new Tg or KO progeny.
Figure 2.
Generation of transgenic (Tg) mice and knock out (KO) mice. The embryonic stem cells (ES) with inserted or disturbed specific gene of interest are microinjected into the mouse blastocyst, which allows the development of a new Tg or KO progeny.


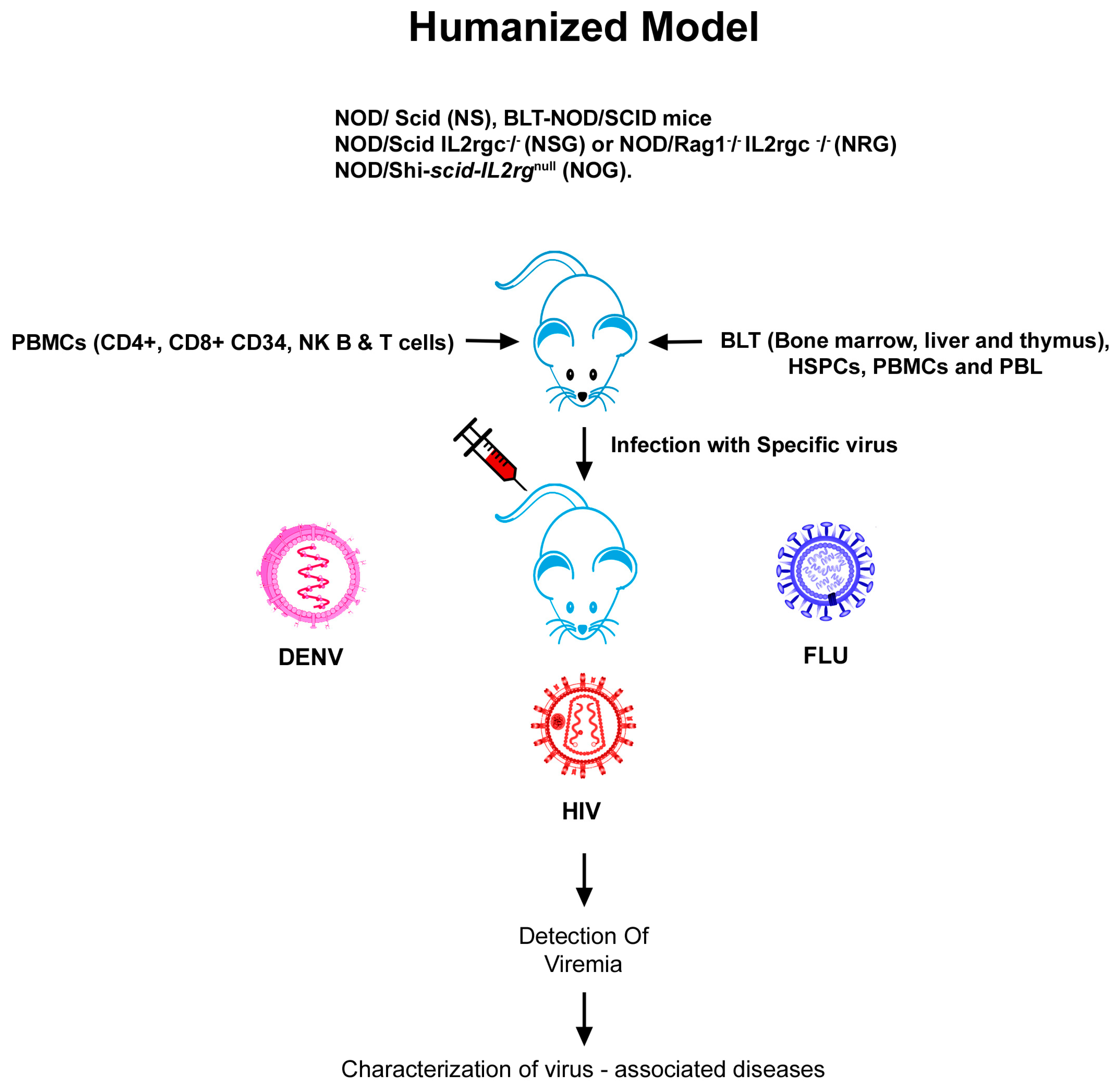{kind=link}
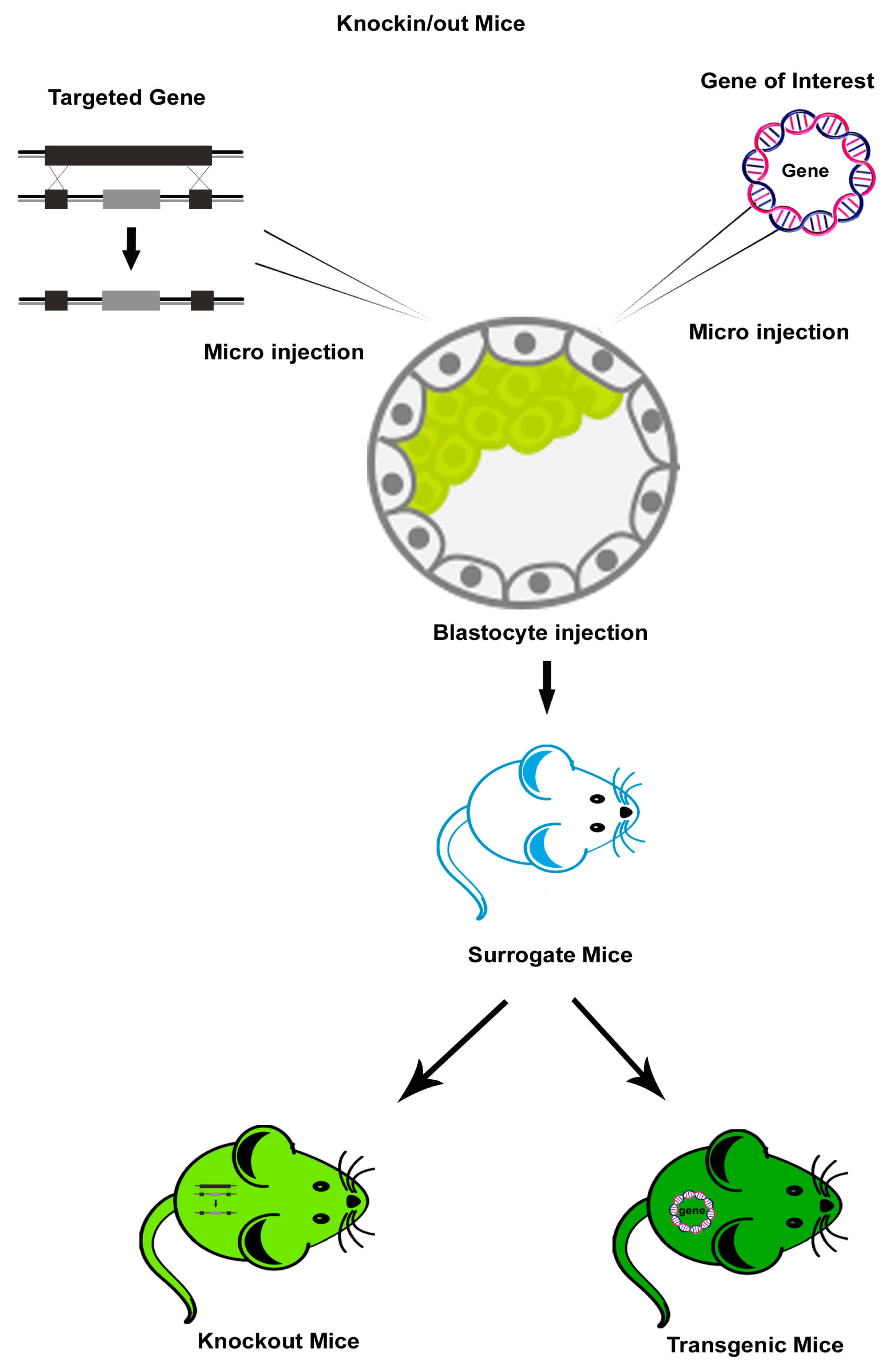{kind=link}
Table 1.
Knockout mouse models for viral infections.
| S.No | Virus | Mouse Model | Name of the Knocked out Gene | Research Application | Reference |
|---|---|---|---|---|---|
| 1. | HIV | Samhd1 KO (samhd1−/−) | Samhd1 deletion. | HIV-1 and HIV-2 entry and pathogenesis | [23,24] |
| 2. | Influenza | C57BL/6J (IL-1R1−/−) | Interleukin receptors α &β. | Pathogenesis of influenza. | [75] |
| C57BL/6J (IL-81−/−) | Interleukin 18 | Pathogenesis of influenza. | [76] | ||
| C57BL/6J (cryp−/− and casp1−/−) | Cryopyrin and caspase deficient. | Innate immunity and moderating lung pathology in influenza pneumonia | [77] | ||
| C57BL/6J (FABP5−/−) | Deletion of FABP5. | Anti inflammatory response against H1N1 influenza A | [79] | ||
| 3. | Dengue | 129Sv(ev) (AG129 KO) | Type I& II- IFN receptors deficient. | Vaccines research and anti-viral drug design. | [105,106,107] |
| STAT1−/− 129/Sv/Ev | Deletion in the DNA binding domain of the STAT1 gene. | STAT1-independent host defense mechanism against viruses | [109,110,111] | ||
| C57BL/6J (Irf3−/−x Irf5−/−x Irf7−/− triple knockout) | Deletion of interfereon regulators factors (IRF) 3, 5 and 7 | Antiviral role of IRF-1 by indcuing IFN responses against DENV infection | [113] |
Table 2.
Transgenic mouse models for viral infections.
| S.No | Virus | Mouse Model | Transgene | Application | Reference |
|---|---|---|---|---|---|
| 1. | HIV | C57BL/6. hu CD4/CCR5 Tg | Human CD4 and chemokine receptor genes | To observe pathological phenotypes of HIV | [31] |
| C57BL/6 × C3HF2. CD4C/rtTA × TRE/HIVNef) or (CD4C/rtTA2S-M2 × TRE/HIVNef) double-Tg mice upon doxycycline (DOX) | HIV Nef gene | Cellular and molecular pathways of Nef in HIV pathogenesis | [37,38] | ||
| C57BL/6 HIVgp120Tg | HIV gp120 | To reveal the role of HIV glycoportein gp120 in binidng to the corecptor CXCR4 in absence of CCR5. CCR5 depletion protects Tg mice against deficits in spatial learning and memory | [41] | ||
| HIV-1 Tg26 transgenic mice | Truncated HIV-1 NL4-3 genome with a 3.1-kb deletion in the Gag and Pol regions | HIV-associated nephropathy | [42] | ||
| 2. | Influenza | B6.SJL Tg Mice | MX1, MX2, FAM3B and TMPRSS2 genes | Zoonotic transmission of influenza A viruses | [83,84] |
| BALB/c. Tg Influenza A HA | Sh-RNAcodes for the knockdown of heamagglutinin | Prevention and control of a viral zoonosis of influenza | [84,85] | ||
| 3. | Dengue | C57BL/6J Tg HLA-A*02:01 and B10. Tg. HLA-DR3 | Geness coding for interspecies hybrid MHC class I molecule of the human HLA-A*0201 allele and the cytoplasmic and transmembrane domains of the mouse H-2Dd class I molecule Genes coding for MHC Class II gene comprising HLA- DR α genomic fragment and a DRB1*030113 | To study the CD8+ T cell response to H7N3 influneza A vaccine. Identification of CD4+T cell epitopes for vaccine development | [85,86] |
Table 3.
Humanized mouse models for viral infections.
| S.No | Virus | Mouse Model | Humanization | Application | Reference |
|---|---|---|---|---|---|
| 1. | HIV | Hu-PBL-SCID mice | SCID mice populated with human peripheral blood leukocytes | HIV infection, replication and pathogenesis | [16] |
| HSCs-BLT mice, NOD-SCID BLT and NSG-BLT. | HSCs-mice engrafted and bone liver/thymus | Human disease pathogenesis, retroviral spread and restored CD4+ and CD8+ T cell numbers on ART treatment | [43,44,45,46,47,48] | ||
| C57BL/6 Rag2−/− γc −/−CD47−/− triple knockout (TKO)-BLT mouse. | Xenotransplantation with human immune system | HIV-latency | [50] | ||
| NOD/ SCID (NS), NOD/SCID IL2rgc−/− (NSG) or NOD/Rag1−/− IL2rgc−/− (NRG) | Reconstitution of different types of human tissues | Treatment of systemic HIV infection with ART and HIV latency | [45,46,47,48] | ||
| 2. | Influenza | C57BL/10SgAiRag2−/−γc−/− mice. | Humanized with huPBMCs | Vaccine based studies and therapeutics for human pathogens | [93] |
| DRAGA mouse; HLA-A2. HLADR4. Rag1KO. IL-2Rgc KO. NOD. | Humanized with functional human immune system | Anti-influenza monoclonal antibodies | [97] | ||
| NOD/Shi-SCID-IL2rγnull (NOG). | Humanized with huPBMCs. | Evaluating vaccine safety | [99] | ||
| 3. | Dengue | RAG2−/−γc−/− mice | Xenografted with human CD34+ hematopoietic stem cells | Antibody responses against DENV | [119] |
| NOD-SCID IL2rγ null | Transplanation of purified cord blood CD34+ cells | To demonstrate differences in the virulence of different DENV-2 strains | [120] | ||
| HIS BLT-NOD/SCID mice | Human immune system | Preclinical testing of antiviral drugs against dengue | [127] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Krishnakumar, V.; Durairajan, S.S.K.; Alagarasu, K.; Li, M.; Dash, A.P. Recent Updates on Mouse Models for Human Immunodeficiency, Influenza, and Dengue Viral Infections. Viruses 2019, 11, 252. https://doi.org/10.3390/v11030252
AMA Style
Krishnakumar V, Durairajan SSK, Alagarasu K, Li M, Dash AP. Recent Updates on Mouse Models for Human Immunodeficiency, Influenza, and Dengue Viral Infections. Viruses. 2019; 11(3):252. https://doi.org/10.3390/v11030252
Chicago/Turabian StyleKrishnakumar, Vinodhini, Siva Sundara Kumar Durairajan, Kalichamy Alagarasu, Min Li, and Aditya Prasad Dash. 2019. "Recent Updates on Mouse Models for Human Immunodeficiency, Influenza, and Dengue Viral Infections" Viruses 11, no. 3: 252. https://doi.org/10.3390/v11030252
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.