Isolation of a T7-Like Lytic Pasteurella Bacteriophage vB_PmuP_PHB01 and Its Potential Use in Therapy against Pasteurella multocida Infections

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
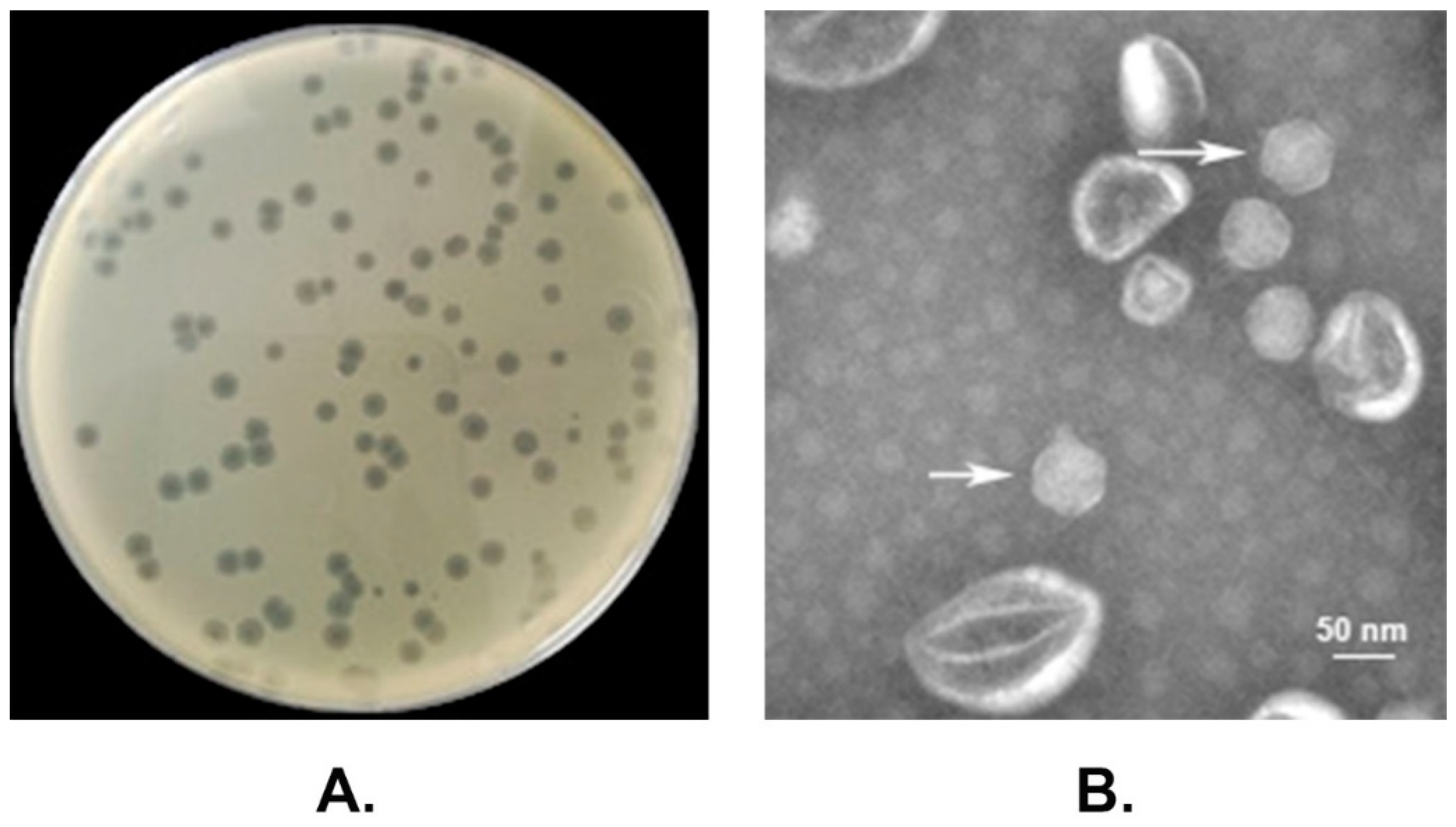
2.1. Phage Isolation
2.2. Electron Microscopy
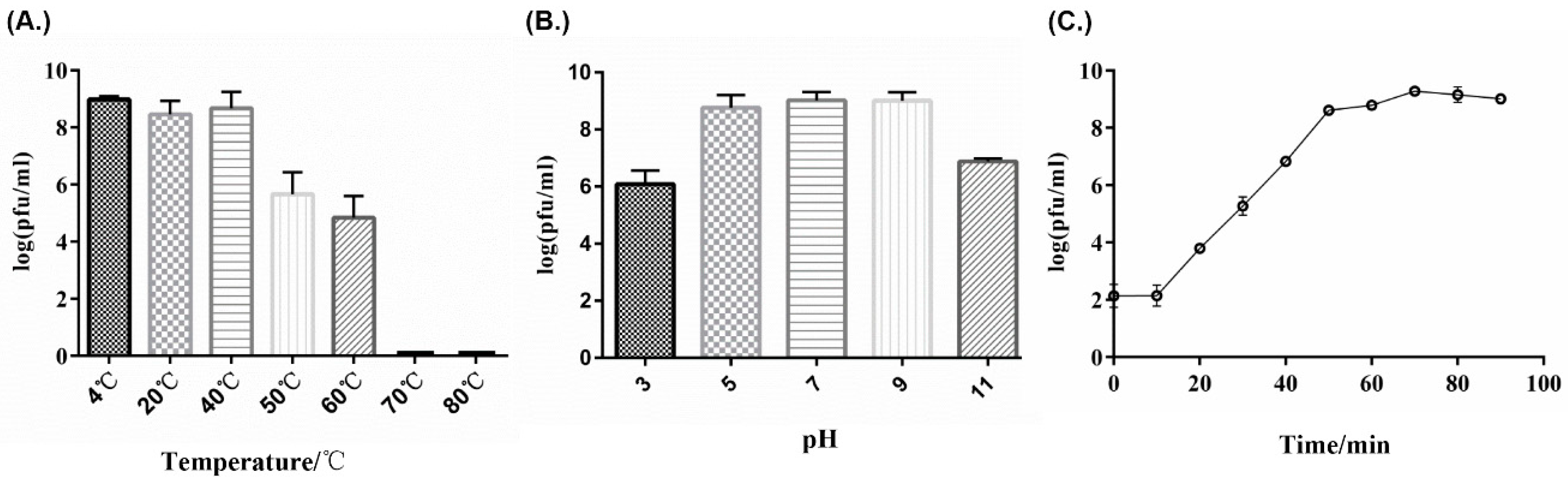
2.3. Thermolability and pH Sensitivity
2.4. One-Step Growth Curve
2.5. Host Range
2.6. DNA Extraction and Analysis of Genome Sequence
2.7. Animals and Ethic Statement
2.8. Safety Test
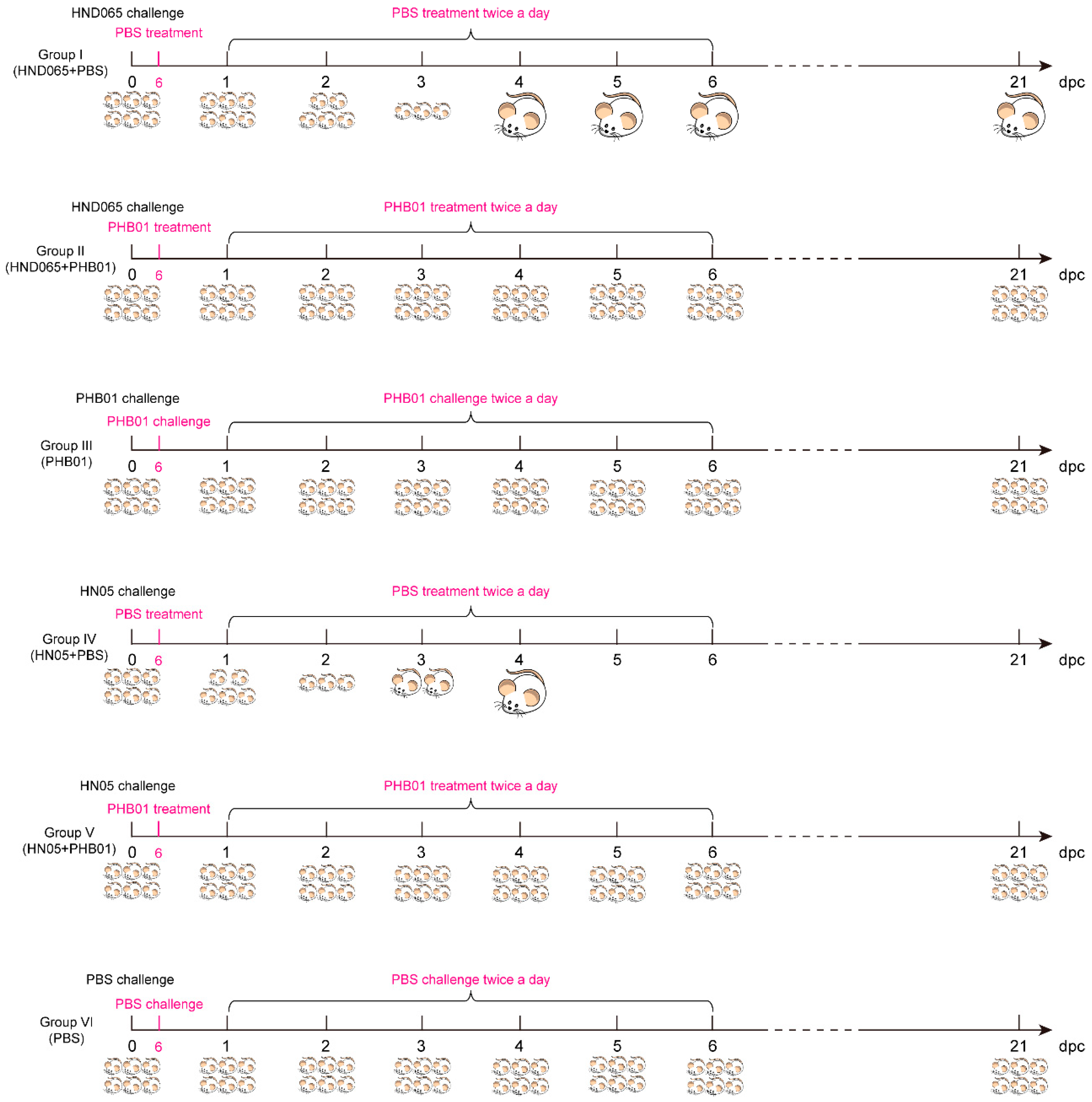
2.9. Mouse Infection and Treatment
3. Results
3.1. Morphological Characteristics
3.2. Life Cycle Parameters
3.3. Host Range of PHB01
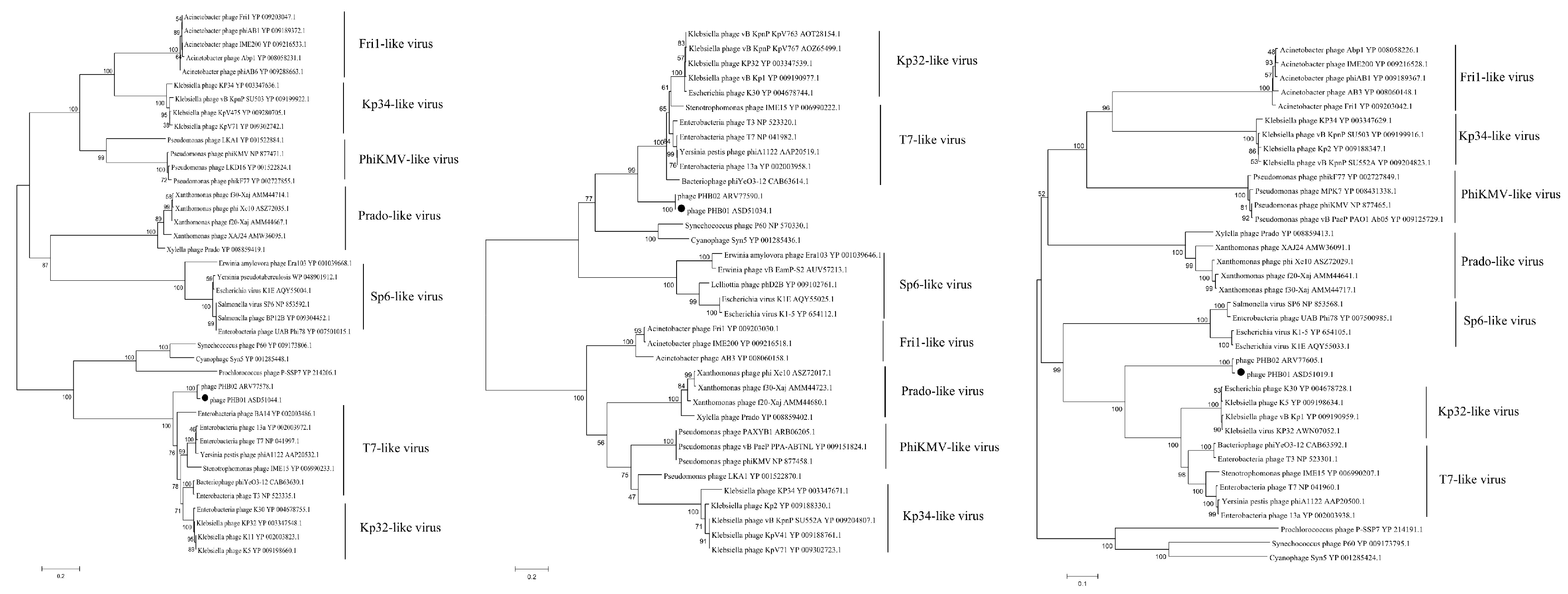
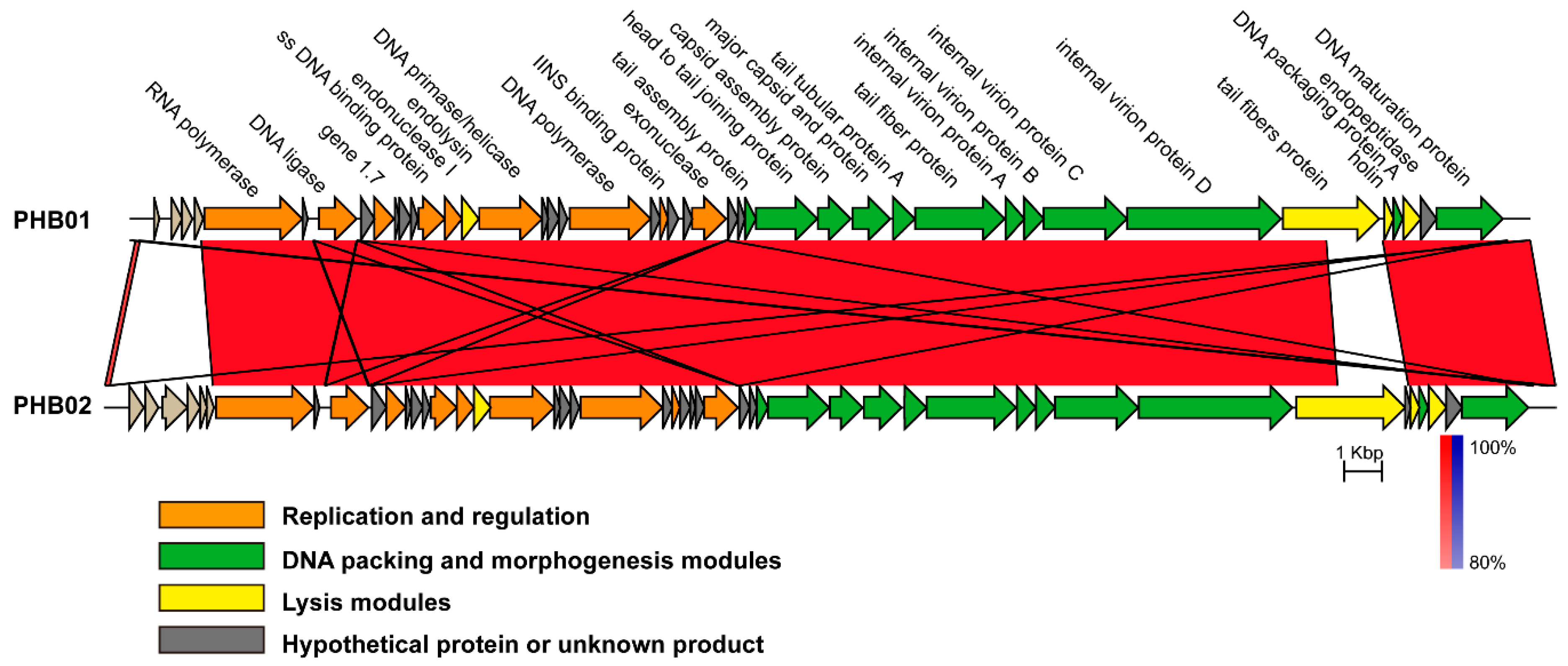
3.4. Genomic Characteristics of PHB01
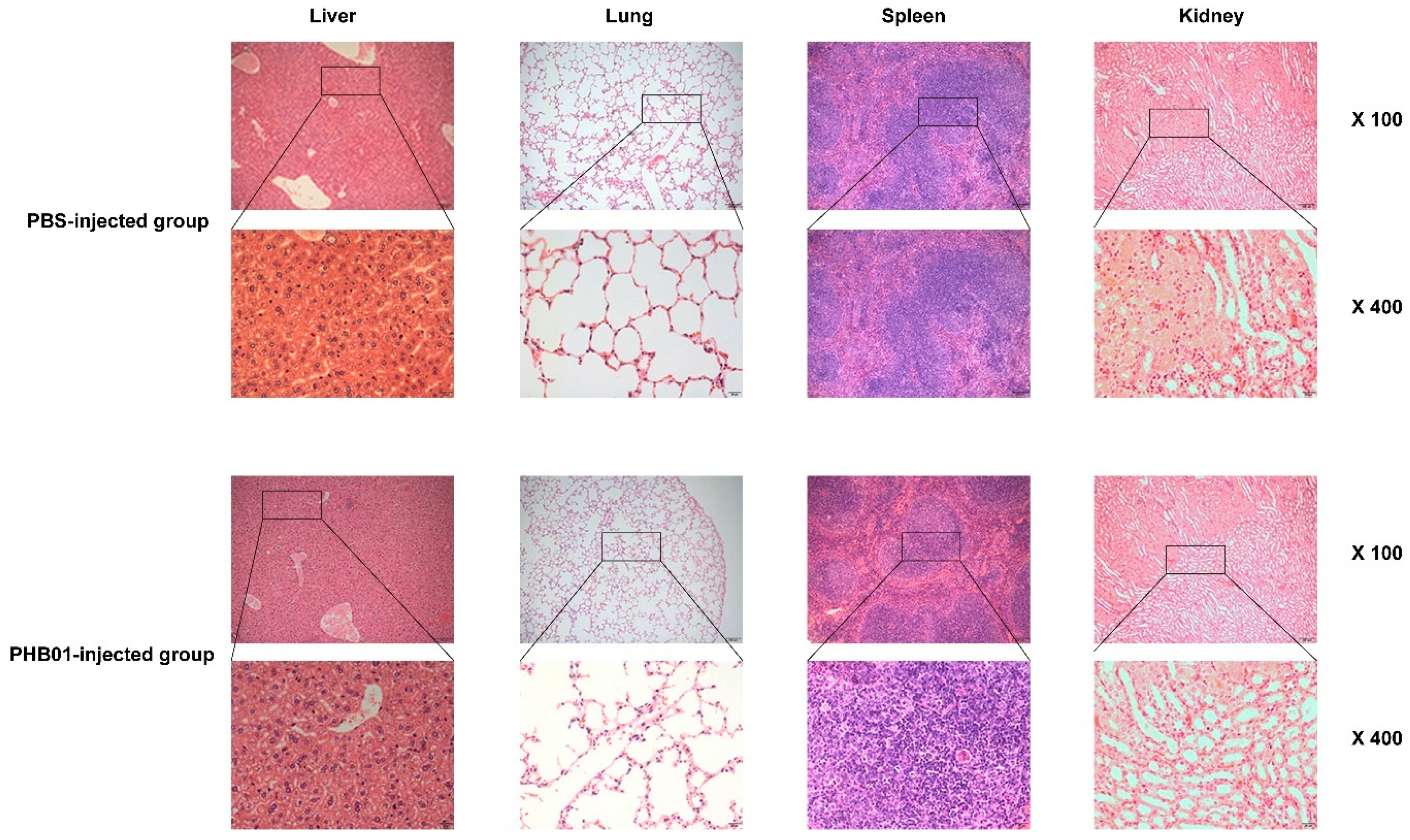
3.5. Acute Toxicity
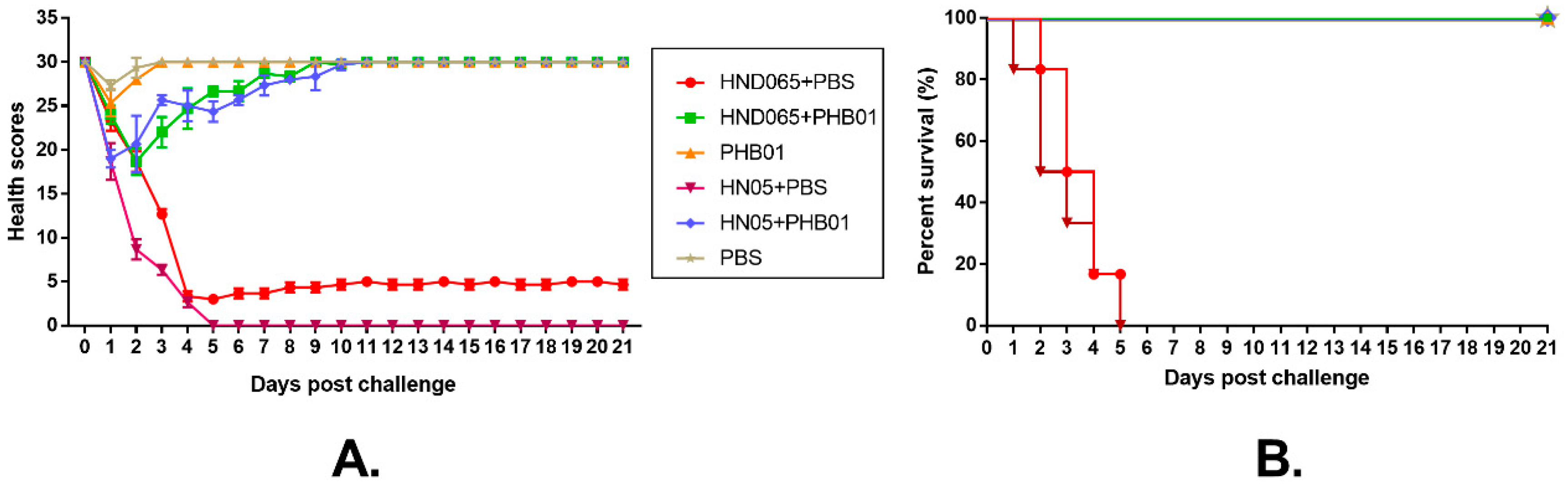
3.6. Therapeutic Effect of the Phage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Carter, G.R. Studies on Pasteurella multocida. I. A hemagglutination test for the identification of serological types. Am. J. Vet. Res. 1955, 16, 481–484. [Google Scholar] [PubMed]
- Wilkie, I.W.; Harper, M.; Boyce, J.D.; Adler, B. Pasteurella multocida: Diseases and pathogenesis. Curr. Top. Microbiol. Immunol. 2012, 361, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Katechakis, N.; Maraki, S.; Dramitinou, I.; Marolachaki, E.; Koutla, C.; Ioannidou, E. An unusual case of Pasteurella multocida bacteremic meningitis. J. Infect. Public Health 2018. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.; Rodriguez-Lucas, C.; Rodicio, M.R.; Vela, A.I.; Fernandez-Garayzabal, J.F.; Leiva, P.S.; Cuesta, F.; Cid, D.; Fernandez, J. Human Pasteurella multocida Infection with Likely Zoonotic Transmission from a Pet Dog, Spain. Emerg. Infect. Dis. 2018, 24, 1145–1146. [Google Scholar] [CrossRef] [PubMed]
- Pak, S.; Valencia, D.; Decker, J.; Valencia, V.; Askaroglu, Y. Pasteurella multocida pneumonia in an immunocompetent patient: Case report and systematic review of literature. Lung India 2018, 35, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Maleb, A.; Elmalki, J.; Bouayadi, O.; Ben Lahlou, Y.; Frikh, M.; Abdeljaouad, N.; Lemnouer, A.; Yacoubi, H.; Elouennass, M. Serious phlegmonous lesion of the hand following an injury by vegetal thorn: Never forget Pasteurella multocida! Trauma Case Rep. 2018, 13, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Kim, S.H.; Yoo, G.; Hwang, G.Y.; Uh, Y.; Yoon, K.J. First Case of Pasteurella multocida Pneumonic Bacteremia in Korea. Ann. Lab. Med. 2018, 38, 490–491. [Google Scholar] [CrossRef]
- Wilson, B.A.; Ho, M. Pasteurella multocida: From zoonosis to cellular microbiology. Clin. Microbiol. Rev. 2013, 26, 631–655. [Google Scholar] [CrossRef]
- Peng, Z.; Liang, W.; Wang, F.; Xu, Z.; Xie, Z.; Lian, Z.; Hua, L.; Zhou, R.; Chen, H.; Wu, B. Genetic and Phylogenetic Characteristics of Pasteurella multocida Isolates From Different Host Species. Front. Microbiol. 2018, 9, 1408. [Google Scholar] [CrossRef]
- Kubatzky, K.F. Pasteurella multocida and immune cells. Curr. Top. Microbiol. Immunol. 2012, 361, 53–72. [Google Scholar] [CrossRef]
- Srinivasiah, S.; Bhavsar, J.; Thapar, K.; Liles, M.; Schoenfeld, T.; Wommack, K.E. Phages across the biosphere: Contrasts of viruses in soil and aquatic environments. Res. Microbiol. 2008, 159, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Can, K.; Aksu, U.; Yenen, O.S. Investigation of PhiKZ phage therapy against Pseudomonas aeruginosa in mouse pneumonia model. Turk. J. Med. Sci. 2018, 48, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Akusobi, C.; Chan, B.K.; Williams, E.; Wertz, J.E.; Turner, P.E. Parallel Evolution of Host-Attachment Proteins in Phage PP01 Populations Adapting to Escherichia coli O157:H7. Pharmaceuticals 2018, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Euler, C.W.; Delaune, A.; Fischetti, V.A. Using a Novel Lysin To Help Control Clostridium difficile Infections. Antimicrob. Agents Chemother. 2015, 59, 7447–7457. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Wang, X.; Wang, L.; Li, Z.; Che, J.; Wang, L.; Li, X.; Cao, Z.; Zhang, J.; Jin, L.; et al. Evaluation of the efficacy of a bacteriophage in the treatment of pneumonia induced by multidrug resistance Klebsiella pneumoniae in mice. Biomed. Res. Int. 2015, 2015, 752930. [Google Scholar] [CrossRef]
- Kirchner, C.; Eisenstark, A. Lysogeny in Pasteurella multocida. Am. J. Vet. Res. 1956, 17, 547–548. [Google Scholar] [PubMed]
- Chen, Y.; Sun, E.; Song, J.; Yang, L.; Wu, B. Complete Genome Sequence of a Novel T7-Like Bacteriophage from a Pasteurella multocida Capsular Type A Isolate. Curr. Microbiol. 2018, 75, 574–579. [Google Scholar] [CrossRef]
- Qureshi, S.; Saxena, H.M.; Imam, N.; Kashoo, Z.; Sharief Banday, M.; Alam, A.; Malik, M.Z.; Ishrat, R.; Bhat, B. Isolation and genome analysis of a lytic Pasteurella multocida Bacteriophage PMP-GAD-IND. Lett. Appl. Microbiol. 2018, 67, 244–253. [Google Scholar] [CrossRef]
- Campoy, S.; Aranda, J.; Alvarez, G.; Barbe, J.; Llagostera, M. Isolation and sequencing of a temperate transducing phage for Pasteurella multocida. Appl. Environ. Microbiol. 2006, 72, 3154–3160. [Google Scholar] [CrossRef]
- Fillol-Salom, A.; Martinez-Rubio, R.; Abdulrahman, R.F.; Chen, J.; Davies, R.; Penades, J.R. Phage-inducible chromosomal islands are ubiquitous within the bacterial universe. ISME J. 2018. [Google Scholar] [CrossRef]
- Chang, H.C.; Chen, C.R.; Lin, J.W.; Shen, G.H.; Chang, K.M.; Tseng, Y.H.; Weng, S.F. Isolation and characterization of novel giant Stenotrophomonas maltophilia phage phiSMA5. Appl. Environ. Microbiol. 2005, 71, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Chibani-Chennoufi, S.; Sidoti, J.; Bruttin, A.; Kutter, E.; Sarker, S.; Brussow, H. In vitro and in vivo bacteriolytic activities of Escherichia coli phages: Implications for phage therapy. Antimicrob. Agents Chemother. 2004, 48, 2558–2569. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Xu, J.; Yao, H.; Lu, C.; Zhang, W. Isolation, genome sequencing and functional analysis of two T7-like coliphages of avian pathogenic Escherichia coli. Gene 2016, 582, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Ul Haq, I.; Chaudhry, W.N.; Andleeb, S.; Qadri, I. Isolation and partial characterization of a virulent bacteriophage IHQ1 specific for Aeromonas punctata from stream water. Microb. Ecol. 2012, 63, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cai, L.; Ma, R.; Xu, Y.; Tong, Y.; Huang, Y.; Jiao, N.; Zhang, R. A novel roseosiphophage isolated from the oligotrophic South China Sea. Viruses 2017, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.C.; Hair, B.B.; Wienclaw, T.M.; Murdock, M.H.; Hatch, J.B.; Trent, A.T.; White, T.D.; Haskell, K.J.; Berges, B.K. Isolation and Host Range of Bacteriophage with Lytic Activity against Methicillin-Resistant Staphylococcus aureus and Potential Use as a Fomite Decontaminant. PLoS ONE 2015, 10, e0131714. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, 1–6. [Google Scholar] [CrossRef]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Li, Y.; Li, S.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
- Li, S.; Fan, H.; An, X.; Fan, H.; Jiang, H.; Chen, Y.; Tong, Y. Scrutinizing virus genome termini by high-through put sequencing. PLoS ONE 2014, 9, e85806. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Li, D.; Li, X.; Hu, L.; Cheng, M.; Xia, F.; Gong, P.; Wang, B.; Ge, J.; Zhang, H.; et al. LysGH15 kills Staphylococcus aureus without being affected by the humoral immune response or inducing inflammation. Sci. Rep. 2016, 6, 29344. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, E.; Yang, L.; Song, J.; Wu, B. Therapeutic Application of Bacteriophage PHB02 and its Putative Depolymerase Against Pasteurella multocida Capsular Type A in Mice. Front. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Le, S.; Tan, Y.; Zhu, J.; Li, M.; Rao, X.; Zou, L.; Li, S.; Wang, J.; Jin, X.; et al. Genomic and proteomic analyses of the terminally redundant genome of the Pseudomonas aeruginosa phage PaP1: Establishment of genus PaP1-like phages. PLoS ONE 2013, 8, e62933. [Google Scholar] [CrossRef]
- Ackermann, H.W.; Karaivanov, L. Morphology of Pasteurella multocida bacteriophages. Can. J. Microbiol. 1984, 30, 1141–1148. [Google Scholar] [CrossRef]
- Kolaskar, A.S.; Tongaonkar, P.C. A semi-empirical method for prediction of antigenic determinants on protein antigens. FEBS Lett. 1990, 276, 172–174. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Mao, J.; Xie, J. Bacteriophage polysaccharide depolymerases and biomedical applications. BioDrugs Clin. Immunother. Biopharma. Gene Ther. 2014, 28, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Battisti, A.J.; Bowman, V.D.; Stummeyer, K.; Muhlenhoff, M.; Gerardy-Schahn, R.; Scholl, D.; Molineux, I.J. The structures of bacteriophages K1E and K1-5 explain processive degradation of polysaccharide capsules and evolution of new host specificities. J. Mol. Biol. 2007, 371, 836–849. [Google Scholar] [CrossRef] [PubMed]
- Leiman, P.G.; Molineux, I.J. Evolution of a new enzyme activity from the same motif fold. Mol. Microbiol. 2008, 69, 287–290. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Koc, C.; Kuhner, P.; Stierhof, Y.D.; Krismer, B.; Enright, M.C.; Penades, J.R.; Wolz, C.; Stehle, T.; Cambillau, C.; et al. An essential role for the baseplate protein Gp45 in phage adsorption to Staphylococcus aureus. Sci. Rep. 2016, 6, 26455. [Google Scholar] [CrossRef] [PubMed]
- Latka, A.; Maciejewska, B.; Majkowska-Skrobek, G.; Briers, Y.; Drulis-Kawa, Z. Bacteriophage-encoded virion-associated enzymes to overcome the carbohydrate barriers during the infection process. Appl. Microbiol. Biotechnol. 2017, 101, 3103–3119. [Google Scholar] [CrossRef] [PubMed]
- Boyce, J.D.; Chung, J.Y.; Adler, B. Pasteurella multocida capsule: Composition, function and genetics. J. Biotechnol. 2000, 83, 153–160. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Strain (Genotype) | Isolated Locations | Efficiency of Plating (EOP) |
|---|---|---|---|
| 1 | Pasteurella multocida D strain HND065 | Henan, China | 1 |
| 2 | Pasteurella multocida D | Guangdong, China | <0.01 |
| 3 | Pasteurella multocida D | Hubei, China | 1.1 |
| 4 | Pasteurella multocida D | Anhui, China | 0.66 |
| 5 | Pasteurella multocida D | Henan, China | 1.33 |
| 6 | Pasteurella multocida D | Hubei, China | 1 |
| 7 | Pasteurella multocida D | Hubei, China | 0.5 |
| 8 | Pasteurella multocida D | Hubei, China | 0.06 |
| 9 | Pasteurella multocida D | Guangdong, China | <0.01 |
| 10 | Pasteurella multocida D | Hubei, China | <0.01 |
| 11 | Pasteurella multocida D | Hubei, China | 0.84 |
| 12 | Pasteurella multocida D | Anhui, China | 0.83 |
| 13 | Pasteurella multocida D | Fujian, China | <0.001 |
| 14 | Pasteurella multocida D | Guangdong, China | 0.25 |
| 15 | Pasteurella multocida D | Hubei, China | 1.5 |
| 16 | Pasteurella multocida D | Shanxi, China | 0.58 |
| 17 | Pasteurella multocida D | Shanghai, China | 0.5 |
| 18 | Pasteurella multocida D | Shanxi, China | 0.75 |
| 19 | Pasteurella multocida D | Hubei, China | 0.5 |
| 20 | Pasteurella multocida D | Hubei, China | 0.66 |
| 21 | Pasteurella multocida D | Hubei, China | <0.001 |
| 22 | Pasteurella multocida D | Guangdong, China | <0.001 |
| 23 | Pasteurella multocida D | Guangdong, China | - |
| 24 | Pasteurella multocida D | Anhui, China | - |
| 25 | Pasteurella multocida D | Shanxi, China | - |
| 26 | Pasteurella multocida D | Guangdong, China | - |
| 27 | Pasteurella multocida D | Unknown | - |
| 28 | Pasteurella multocida D | Unknown | - |
| 29 | Pasteurella multocida D | Unknown | - |
| 30 | Pasteurella multocida D | Unknown | - |
| 31 | Pasteurella multocida D | Unknown | - |
| 32 | Pasteurella multocida D | Unknown | - |
| 33 | Pasteurella multocida D | Unknown | - |
| 34 | Pasteurella multocida D strain HN06 | Hainan, China | - |
| 35 | Pasteurella multocida D | Hubei, China | - |
| 36 | Pasteurella multocida D | Guangdong, China | - |
| 37 | Pasteurella multocida D | Shanxi, China | - |
| 38 | Pasteurella multocida A | Beijing, China | - |
| 39 | Pasteurella multocida A | Hubei, China | - |
| 40 | Pasteurella multocida A | Hubei, China | - |
| 41 | Pasteurella multocida A | Hunan | - |
| 42 | Pasteurella multocida A | Sichuan | - |
| 43 | Pasteurella multocida A | Sichuan | - |
| 44 | Pasteurella multocida A | Guangdong | - |
| 45 | Pasteurella multocida A | Guangdong | - |
| 46 | Pasteurella multocida A strain HB01 | Hubei | - |
| 47 | Pasteurella multocida A strain HB03 | Hubei | - |
| 48 | Pasteurella multocida F strain HN07 | Henan | - |
| 49 | Escherichia coli DH5α | - | |
| 50 | Salmonella enterica serovar typhimurium ATCC14028 | - | |
| 51 | Salmonella enterica serovar choleraesuis | - | |
| 52 | Bordetella bronchiseptica | - | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Guo, G.; Sun, E.; Song, J.; Yang, L.; Zhu, L.; Liang, W.; Hua, L.; Peng, Z.; Tang, X.; et al. Isolation of a T7-Like Lytic Pasteurella Bacteriophage vB_PmuP_PHB01 and Its Potential Use in Therapy against Pasteurella multocida Infections. Viruses 2019, 11, 86. https://doi.org/10.3390/v11010086
Chen Y, Guo G, Sun E, Song J, Yang L, Zhu L, Liang W, Hua L, Peng Z, Tang X, et al. Isolation of a T7-Like Lytic Pasteurella Bacteriophage vB_PmuP_PHB01 and Its Potential Use in Therapy against Pasteurella multocida Infections. Viruses. 2019; 11(1):86. https://doi.org/10.3390/v11010086
Chicago/Turabian StyleChen, Yibao, Guanghao Guo, Erchao Sun, Jiaoyang Song, Lan Yang, Lili Zhu, Wan Liang, Lin Hua, Zhong Peng, Xibiao Tang, and et al. 2019. "Isolation of a T7-Like Lytic Pasteurella Bacteriophage vB_PmuP_PHB01 and Its Potential Use in Therapy against Pasteurella multocida Infections" Viruses 11, no. 1: 86. https://doi.org/10.3390/v11010086