Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation


1
Department of Food Science and Human Nutrition, University of Illinois, 905 S. Goodwin Ave., Urbana, IL 61801, USA
2
Carl R. Woese Institute for Genomic Biology, University of Illinois at Urbana-Champaign, 1206 W. Gregory Dr., Urbana, IL 61801, USA
*
Author to whom correspondence should be addressed.
Viruses 2018, 10(6), 281; https://doi.org/10.3390/v10060281
Submission received: 19 February 2018
/
Revised: 2 May 2018
/
Accepted: 17 May 2018
/
Published: 24 May 2018
(This article belongs to the Special Issue Phage Lytic Enzymes and Their Applications)
Abstract
:Contamination of fuel-ethanol fermentations continues to be a significant problem for the corn and sugarcane-based ethanol industries. In particular, members of the Lactobacillaceae family are the primary bacteria of concern. Currently, antibiotics and acid washing are two major means of controlling contaminants. However, antibiotic use could lead to increased antibiotic resistance, and the acid wash step stresses the fermenting yeast and has limited effectiveness. Bacteriophage endolysins such as LysA2 are lytic enzymes with the potential to contribute as antimicrobials to the fuel ethanol industries. Our goal was to evaluate the potential of yeast-derived LysA2 as a means of controlling Lactobacillaceae contamination. LysA2 intracellularly produced by Pichia pastoris showed activity comparable to Escherichia coli produced LysA2. Lactic Acid Bacteria (LAB) with the A4α peptidoglycan chemotype (L-Lys-D-Asp crosslinkage) were the most sensitive to LysA2, though a few from that chemotype were insensitive. Pichia-expressed LysA2, both secreted and intracellularly produced, successfully improved ethanol productivity and yields in glucose (YPD60) and sucrose-based (sugarcane juice) ethanol fermentations in the presence of a LysA2 susceptible LAB contaminant. LysA2 secreting Sacharomyces cerevisiae did not notably improve production in sugarcane juice, but it did control bacterial contamination during fermentation in YPD60. Secretion of LysA2 by the fermenting yeast, or adding it in purified form, are promising alternative tools to control LAB contamination during ethanol fermentation. Endolysins with much broader lytic spectrums than LysA2 could supplement or replace the currently used antibiotics or the acidic wash.
1. Introduction
In the search for renewable sources to replace petroleum-based fuel, ethanol from carbohydrate-rich plant sources continues to outstrip the alternatives in terms of production and efficiency at the desired scales. The United States of America (US) and Brazil combined produced more than 20 billion gallons of fuel ethanol primarily from corn and sugarcane in 2014 [1]. Among the various outstanding issues, bacterial contamination of fuel ethanol fermentation continues to be a challenge to producers [2,3,4,5], and can make the difference between running at a profit or a loss [6]. For example, one analysis determined that a 100 MMgy fuel ethanol plant with a moderate level of contamination can expect an annual revenue loss of $4.5 million at 2016 ethanol prices [7].
Both wild yeasts and bacteria that thrive under the ethanol fermentation conditions preferred by industrial Saccharomyces cerevisiae strains have been identified as sources of contamination. The predominant bacterial contaminants are Gram positive bacteria from the Lactobacillaceae family, including Lactobacillus, Pediococcus, Leuconostoc, Enterococcus, Aerococcus, and Weisella [3,5,6,8]. These bacteria, which produce primarily lactic acid from carbohydrate sources, are often referred to as Lactic Acid Bacteria (LAB), and many have adapted to conditions with comparatively low pHs and oxygen levels, as well as high salt and ethanol concentrations. Consequently, LAB contaminations have been a recurring problem in wine, beer, and spirit fermentations [9,10,11]. Biofuel fermentations are particularly at risk from LAB contamination because the plants that provide the fermentation substrate are the natural habitats of LAB, and these substrates are rarely subjected to bacteriostatic or bactericidal treatment. The predominant genera of LAB contaminants identified in biofuel fermentations is Lactobacillus, regardless of whether the fermentation is from corn [4,8], sugarcane [5,12], or lignocellulose [13]. Treatments to control bacterial contamination of fuel ethanol fermentation must include measures against lactobacilli and other contaminating LAB, in order to be effective. Methods explored to combat contamination by LAB include pretreatment of feedstock, changing process conditions such as solids content and pH, cleaning fermentation equipment, antibiotics, bacteriocins, acid washing of yeast used for sugarcane fermentations, and bacteriophages. Currently, acid treatment of the yeast, cleaning of equipment, and antibiotics are the most commonly used measures in industry. Acid-washing stresses the fermenting yeast and can lead to decreased ethanol production efficiency [14]. US fuel ethanol plants use penicillin and virginiamycin [15,16] while Brazilian plants are more likely to use monensin [17,18] although the efficacy of antibiotic treatments has been challenged [19]. Additional complications from antibiotic use are unacceptably high antibiotic residues in co-products, the dried distillers grain solids from corn ethanol plants [20] or dried deactivated yeasts [2] from sugarcane ethanol plants. Increased development of antibiotic resistance in the contaminating strains has been noted as well [19].
Alternative approaches to control the LAB are needed. Researchers are exploring the efficacy of bacteriophages (phages) [4], and in specific instances, have applied for patents using phages as an antimicrobial strategy [21]. The use of phages is an interesting approach, but has a number of drawbacks, including the specificity of the phage to a particular strain and the numerous adaptation mechanisms bacteria have developed, and continue to develop, to evade phage infection. However, phage lysins, proteins which allow the phages to burst bacterial cells, can be applied externally to lyse Gram positive bacteria [22,23,24]. Some lysins, including LysA2, have been shown to impact multiple different strains of LAB, and could be useful in reducing fuel ethanol contamination [25,26]. Lysins have several potential advantages, including an appropriate lytic spectrum that allows individual lysins to lyse several genera of LAB [26]. In addition, lysins have a reduced potential for resistance development compared to antibiotics [27,28]. Lastly, it is possible that the fermenting yeast could heterologously produce the desired lysin at a lower cost.
LysA2 is an endopeptidase from the Lactobacillus casei phage Ø393 A2 [25]. Lactobacillus casei phage Ø393 A2 was isolated from the whey of a failed fermentation of Gamoneu blue cheese [29]. Previous studies found that LysA2 decreased the optical density, as measured by absorbance readings at 600 nm (OD) of a number of LAB [25,26]. These studies suggest that LysA2 might be effective in inhibiting the unwanted growth of LAB in biofuel fermentations. This study further explores the spectrum and activity of LysA2 produced by the yeasts Pichia pastoris GS115 and Saccharomyces cerevisiae D452-2. To our knowledge, this is the first time that a P. pastoris or S. cerevisiae secreting LysA2, or any other endolysin, has been reported in the literature.
2. Materials and Methods
2.1. Bacterial and Yeast Strains
All bacterial and yeast strains used in this study are listed in Table 1. E. coli were cultured in lysogeny broth (LB) and incubated aerobically at 37 °C. When necessary, LB was supplemented with 100 μg/mL ampicillin. Yeast were cultured in YPD medium (1% yeast extract, 2% peptone and 2% glucose), and incubated aerobically/anaerobically (5% CO2, 5% H2 and 90% N2) at 30°C.
2.2. Construction of a LysA2 Bacterial and Yeast Expression Vectors
Bacterial and yeast expression vectors for LysA2 were constructed based on Invitrogen plasmids (Figure S1). The entire LysA2 open reading frame was codon optimized for E. coli BL21 and P. pastoris GS115. The E. coli optimized LysA2 gene was cloned into a pRSET A expression vector (pRSETA_LysA2). The P. pastoris optimized LysA2 gene was cloned into pPICZA intracellular (pPICZA_LysA2) and pPICZα (pPICZAa_LysA2) secretion expression vectors. The S. cerevisiae D452-2 strain was used for LysA2 secretion and genome integration. The P. pastoris optimized LysA2 gene with α mating factor was PCR amplified from pPICZAa_LysA2, and cloned into pRS423 expression vector containing the GPD promoter and CYC1 terminator for S. cerevisiae secreted expression (pRS423_LysA2). For integration into the S. cerevisiae genome, the plasmid pITy3 (pITY3_LysA2) was employed, as previously described [30].
2.3. Bacterial and Yeast Transformation with the LysA2 Bacterial and Yeast Expression Vectors
E. coli cells were cultured overnight in 6 mL LB and harvested by centrifugation. Cellular pellets were washed four times in 1 mL 300 mM sucrose, and collected by centrifugation. After washing, the cellular pellet was resuspended in 100 μL of 300 mM sucrose, and used for electroporation at 2.5 kV with pRSETA_LysA2. Putative E. coli transformants were selected by ampicillin-containing LB agar plates (100 μg/mL).
P. pastoris cells were cultured in YPD medium until reaching an OD600 of 2.0. Approximately 8 × 108 cells were suspended in 8 mL of P-transformation buffer (100 mM lithium acetate, 10 mM dithiothreitol, 0.6 M sorbitol and 10 mM Tris-HCL, pH 7.5) for 30 min at room temperature. P. pastoris cells were harvested and washed three times in 1.5 mL of ice-cold 1 M sorbitol. The cellular pellet was then re-suspended in 100 μL of ice-cold 1 M sorbitol and used for electroporation (1.5 kV) with pPICZA_LysA2 (intracellular) or pPICZα_LysA2 (secretion). Putative P. pastoris transformants were screened using agar plates containing 2000 μg/mL of Zeocin.
2.4. Protein Expression and Purification
E. coli cells containing pRSETA_LysA2 were cultured overnight at 37 °C in LB with 100 μg/mL ampicillin. Cell cultures were passaged (1%, v/v) into fresh LB and incubated at 37 °C with agitation at 200 rpm until an OD600 of 0.5. To induce protein expression, 0.2 mM IPTG was added, with further incubation for 5 h at 30 °C with agitation at 200 rpm.
P. pastoris cells containing pPICZA_LysA2 or pPICZα_LysA2 were cultured overnight at 30 °C in BMGY-Buffered Glycerol-complex medium, and harvested by centrifugation (4000 rpm at 4 °C). P. pastoris cell pellets were resuspended to an OD600 of 1.0 in BMMY-Buffered Methanol-complex medium. To induce protein expression, methanol was added every 24 h.
After the appropriate incubation, induced cell cultures were washed twice in binding buffer (300 mM NaCl, 20 mM imidazole, 10% glycerol in 50 mM phosphate buffer, pH 7.5). Cells were broken using a French press cell homogenizer five times, and centrifuged at 20000 ×g for 20 min at 4°C to remove insoluble contents. Soluble lysate was added to a column containing Ni-NTA beads (His60 Ni Superflow Resin, Clontech, Moutain View, CA, USA), and incubated for 3 h at 4°C. The column was washed with 10 column volumes of washing buffer (300 mM NaCl, 40 mM imidazole, 10% glycerol in 50 mM phosphate buffer, pH 7.5). His-tagged LysA2 protein was eluted with elution buffer (300 mM NaCl, 300 mM imidazole, 10% glycerol in 50 mM phosphate buffer, pH 5.5). Purified LysA2 proteins were confirmed by SDS-PAGE using standard procedures. For evaluation of secreted LysA2 from P. pastoris, the culture medium from the induced cells described above was collected by centrifugation. When indicated, the culture medium was concentrated 50 fold using an Amicon ultra centrifugal filter. For evaluation of secreted LysA2 production by S. cerevisiae (both plasmid and chromosomally encoded LysA2 strains), the supernatant of S. cerevisiae cultures at 24 h were taken and concentrated 20 times using an Amicon ultra centrifugal filter, and analyzed by SDS-PAGE (Figure S2). Two target bands were identified and isolated from the protein gel, and analyzed by LC/MS after trypsin treatment. The sequencing results were analyzed using a Mascot distiller and Mascot search engine against the entire NCBI database, confirming that both bands were indeed LysA2.
2.5. Turbidity Reduction Assay
Lactic acid bacteria representative of common biofuel contaminants are listed in Table 2. All LAB were cultured in de Man, Rogosa and Sharpe (MRS) broth and incubated at 37 °C anaerobically overnight. Cells for the turbidy reduction assay were prepared as described by Becker and colleagues [22]. Briefly, LAB overnight cells were centrifuged at 13,000 rpm at 4 °C for 1 h. The supernatant was discarded, and the LAB cellular pellets were stored at −20 °C until use. Prior to the turbidity reduction assay, the LAB cellular pellets were thawed and resuspended in 50 nM phosphate buffer, pH 5.5 to an OD ≈ 2. Purified LysA2 produced by P. pastoris was mixed with the phosphate buffered LAB suspensions at 12.5, 25, 50, 100, 1000 nM in a final volume of 100 μL. The reduction in turbidity at 595 nm was measured using a microplate reader (Multiskan Ascent, Thermo Fisher, Waltham, MA, USA).
Lytic activity of the culture supernatant containing secreted LysA2 from P. pastoris was measured, as described above, for the purified LysA2 proteins. When needed, culture supernatant was concentrated using an Amicon ultra centrifugal filter.
2.6. Fermentation with Simulated Contamination
Mock fermentations were conducted with YPD containing 60 g/L glucose (YPD60) or clarified sugarcane juice. Clarification of sugarcane juice was completed via centrifugation to remove solid contents and filtration with a 0.4 nm bottle-top filter. Clarified sugarcane juice was diluted with purified water to ~6% total sugars content, and supplemented with 0.6 g/L yeast extract. The addition of yeast extract served to replace dead yeast that is typically present in industrial fermentations in Brazil [33,34,35].
S. cerevisiae cells were cultured in synthetic complete glucose broth anaerobically at 30 °C for 24 h, to stabilize the expression vectors. Meanwhile, L. fermentum ATCC 9338 and L. plantarum ATCC 14917 were cultured in MRS broth anaerobically at 30 °C for 24 h. S. cerevisiae and L. fermentum ATCC 9338 or L. plantarum ATCC 14917 were co-inoculated from their separate cultures at an OD600nm 0.5 and 0.05, respectively for YPD60 and OD600nm 5.0 and 0.5, respectively for sugarcane juice. All mock fermentations were performed in a total of 20 mL, and incubated anaerobically at 30 °C, with agitation at 100 rpm for 36 h. At designated intervals, the OD at 600 nm was measured, and 1 mL samples were collected for high-performance liquid chromatography (HPLC) analysis, as previously described [36].
3. Results
3.1. Comparison of Lytic Activity of LysA2 Expressed by Bacteria and Yeast
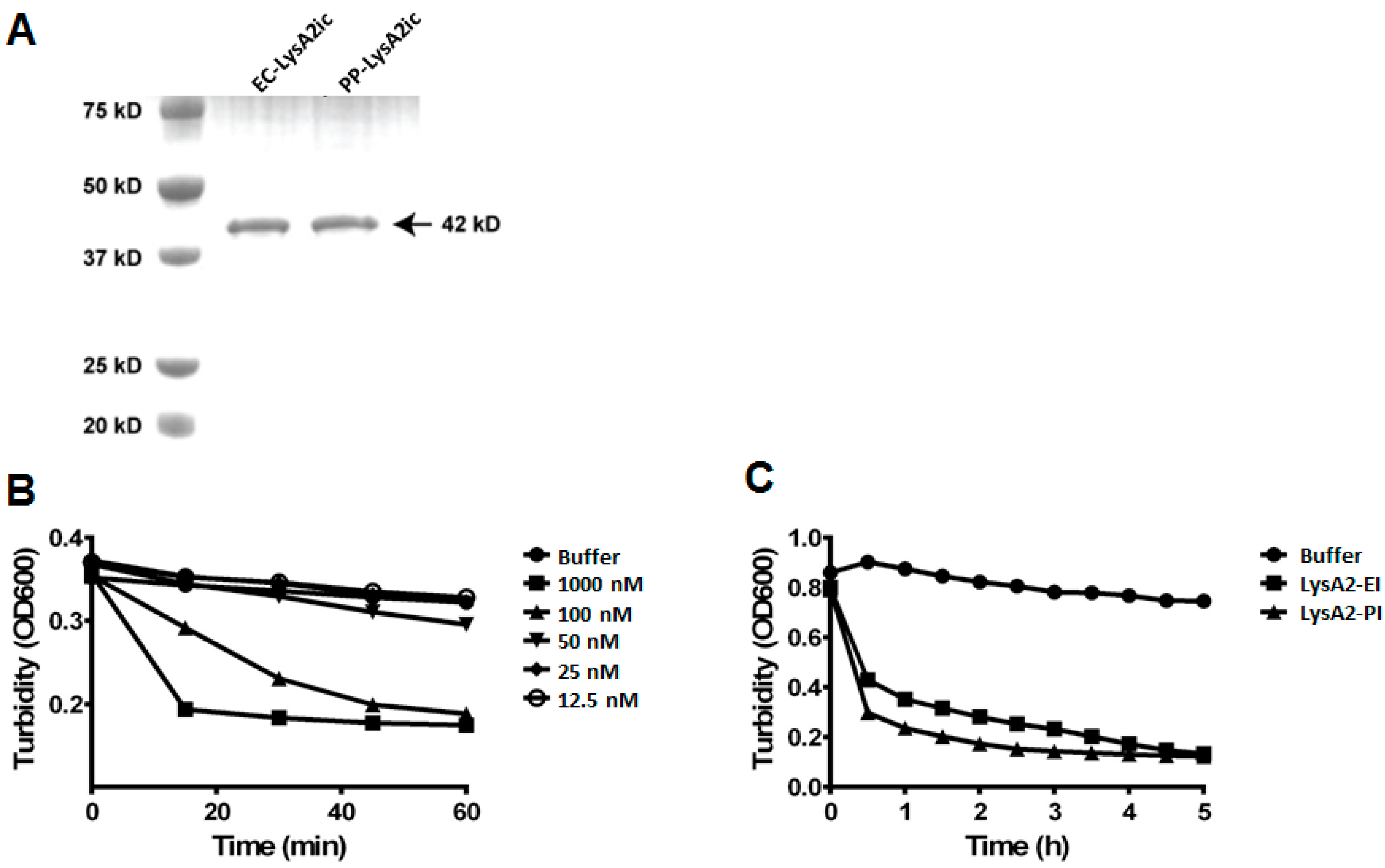
A previous study was unable to get S. cerevisiae to secrete LysA2 [23]. Consequently, we wanted to explore LysA2 expression, both intracellular and secreted, in P. pastoris, which is a eukaryotic model system for heterologous expression of proteins. To compare the activity of LysA2 produced by yeast with the activity of LysA2 from the Escherichia coli BL21 (DE3) expression system used by Ribelles et al. [25], Pichia pastoris GS115 was employed. Intracellularly expressed His6-tagged LysA2 proteins from P. pastoris and E. coli were purified. SDS-PAGE analysis revealed both LysA2 protein bands were the predicted size, approximately 42 kDa (Figure 1A), with no apparent glycosylation of the yeast expressed protein.
To measure the lytic activity of LysA2, the turbidity reduction of a suspension of L. casei was measured after application of the enzyme. Intracellularly produced LysA2 from P. pastoris showed dose-dependent lytic activity, and required concentrations greater than 50 nM to be detected (Figure 1B). The addition of LysA2 from E. coli and P. pastoris produced a comparable turbidity reduction of L. casei cells over five-hour span at the same dose (Figure 1C). This result indicates that LysA2 can be functionally expressed intracellularly in yeast, and the activity of this P. pastoris produced LysA2 is equivalent to that of the E. coli produced LysA2.
3.2. Lytic Activity of Intracellularly Produced LysA2 from P. pastoris on Common Biofuel Contaminants
Thirteen LAB strains, identified as biofuel contaminants in the literature, including six genera with the majority from Lactobacillus, were selected to assess the lytic spectrum of the P. pastoris intracellularly produced LysA2. LysA2 showed the strongest lytic activity against L. casei. Compared to L. casei, L. paracasei (++++), L. delbrueckii (+++), L. fermentum (++), L. rhamnosus (++), E. faecium (++), E. gallinarum (++) and L. lactis (+) showed at least some sensitivity to LysA2 (Table 2). Aerococcus viridans, L. brevis, L. plantarum, Pediococcus acidilacti and P. damnosus were not susceptible to LysA2. LysA2 produced by E. coli had a similar lytic activity and spectrum compared to LysA2 from P. pastoris (data not shown). Interestingly, three out of five of the non-susceptible strains have the same peptidoglycan chemo-type as L. casei (A4α, L-Lys-D-Asp).
3.3. Secreted LysA2 and Its Lytic Activity
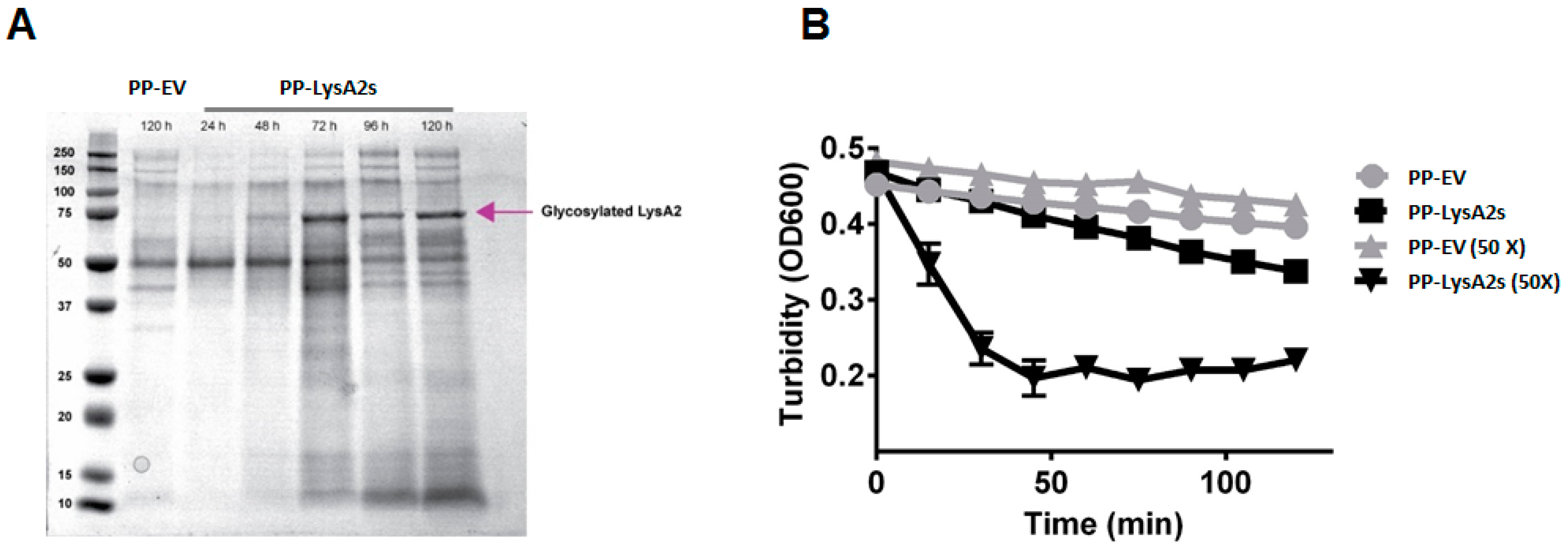
To investigate the lytic activity of LysA2 secreted from yeast, a P. pastoris secreting LysA2 strain was constructed. A protein band of approximately 70 kDa was observed in the concentrated culture medium after 1% methanol induction, and a subsequent 48 h of growth (Figure 2A). There was no comparable band from the concentrated culture medium from P. pastoris containing the empty vector. This putative LysA2 band had a higher molecular weight than the expected protein size of 42 kDa, and could not be purified using a Ni-NTA column. To ensure the putative secreted LysA2 protein was effective at lysing target bacteria, post induction (72 h) culture medium from the LysA2 secreting P. pastoris or from the empty vector P. pastoris was applied to L. casei cells. Turbidity of the L. casei cell suspension only decreased after application of the culture medium from LysA2 expressing P. pastoris (Figure 2B). However, a higher concentration of LysA2 from the culture medium (Figure 2B) was necessary to achieve activity comparable to the 100 nM (4.22 μg/mL) of purified LysA2 that was intracellularly produced by P. pastoris (Figure 1B), indicating that the secretion level of LysA2 is low, but that the protein is active.
Further analysis of the activity of the yeast expressed LysA2 entailed evaluating the impact of LysA2, both intracellular and secreted, on ethanol production by S. cerevisiae in a simulated contamination. Frequently identified LAB contaminants L. fermentum (LF) or L. plantarum (LP) were co-cultured with S. cerevisiae in YPD60 (Figure 3). Without contamination (SC-EV in Figure 3), S. cerevisiae consumed all glucose (60 g/L) in YPD60, and produced ethanol (28 g/L) within 24 h. LF contamination resulted in a 25% reduction in ethanol titer (SC-EV + LF in Figure 3). Adding purified, intracellularly-produced LysA2 (100 μg/mL) completely restored ethanol titer (SC-EV + LysA2-PI + LF in Figure 3). P. pastoris culture media containing secreted LysA2 slightly improved ethanol titer, and the 50-fold concentrated culture media improved control of LF (SC-EV + LysA2-PS + LF and SC-EV + LysA2-PS(50×) + LF in Figure 3); however, the improvement was not as substantial as with 100 µg/mL of intracellularly produced LysA2. The culture media from the empty vector P. pastoris provided no such benefit (SC-EV + PP-EV + LF in Figure 3), even when concentrated (SC-EV + PP-EV(50×) + LF in Figure 3). Since LP is insensitive to LysA2, the addition of purified, intracellularly-produced LysA2 was not effective at improving the reduced ethanol titer caused by LP (SC-EV + LP and SC-EV + LysA2-PI + LP in Figure 3). Based on this indication of lytic activity, the decision was made to attempt to express secreted LysA2 in S. cerevisiae despite the inability of a previous group to get a functional expression of LysA2 by S. cerevisiae [23].
3.4. S. cerevisiae Secreting LysA2 Restores Ethanol Yield in Mock Contamination of Fermentation
Unlike secreted LysA2 from P. pastoris, the putative LysA2 band secreted by S. cerevisiae was approximately 52 kDa (expected size with signal peptide) and another approximately 37 kDa band was observed. After several failed attempts to purify secreted LysA2, the two target bands were isolated from the protein gel and analyzed by LC/MS, following trypsin digestion. The smaller size LysA2 (37 kDa) could be a truncated LysA2 which has been cut by extracellular peptidase. Interestingly, glycosylation of LysA2 was not observed in S. cerevisiae.
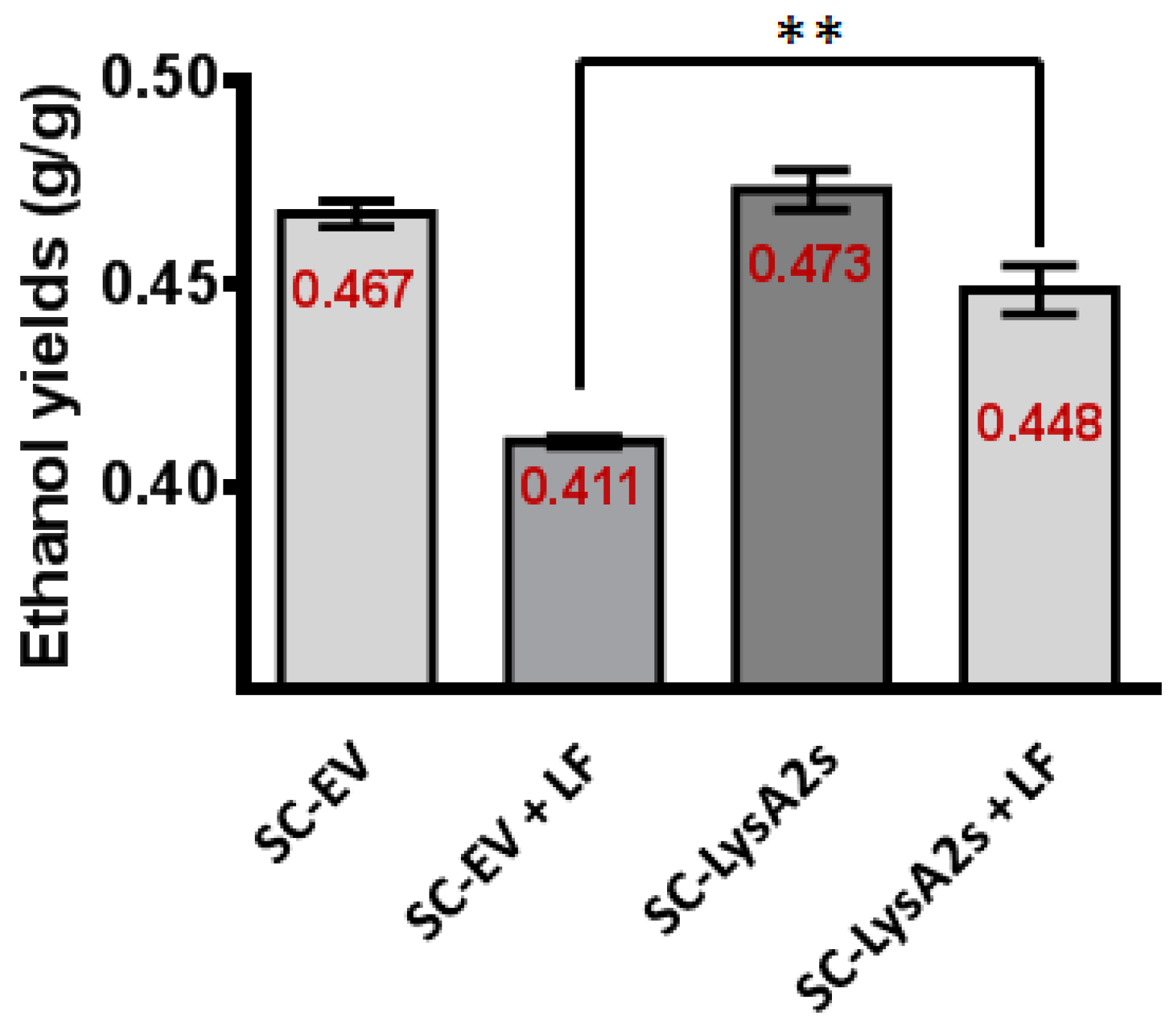
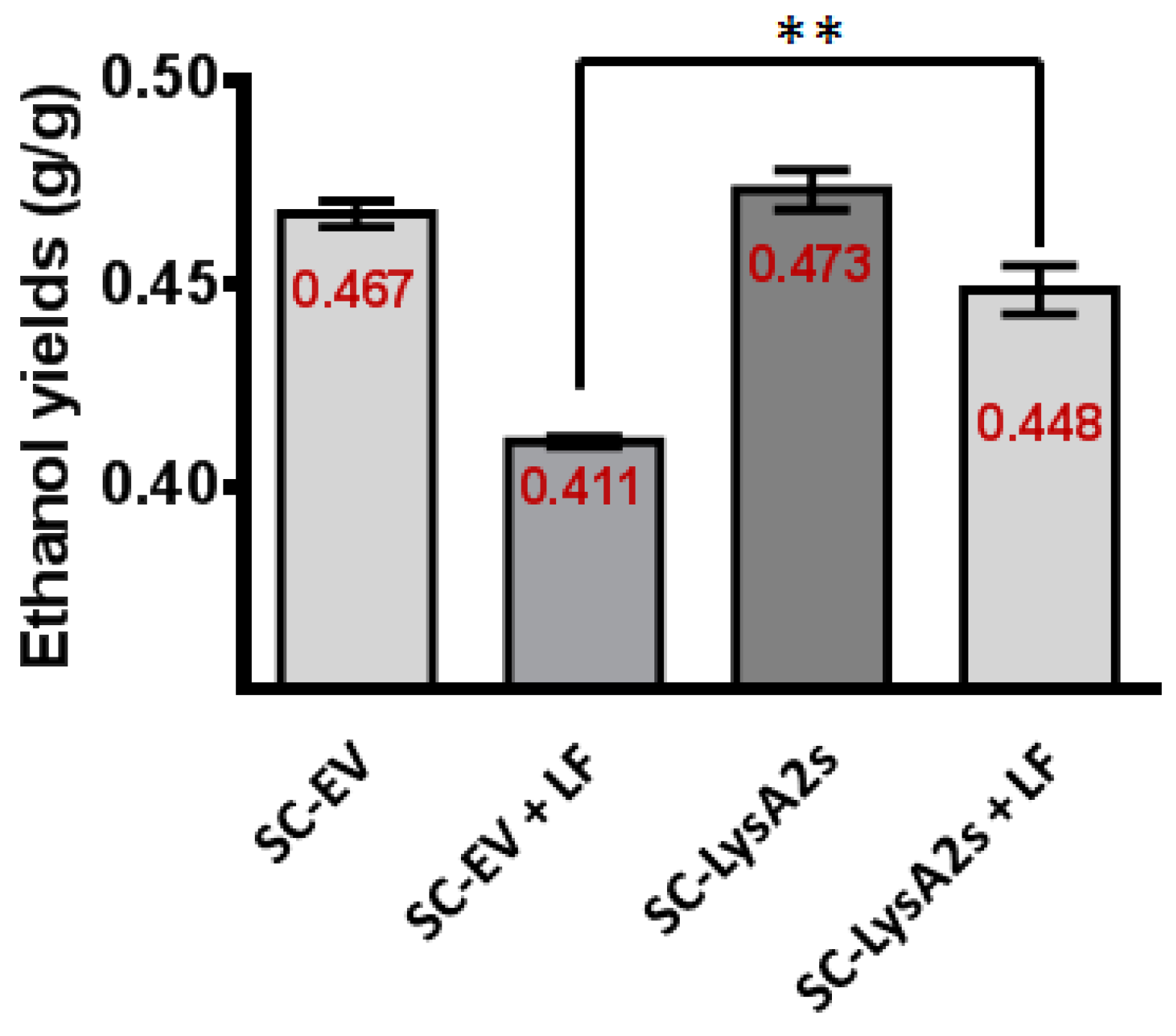
S. cerevisiae containing the LysA2 secreting plasmid (SC-LysA2s), and the same strain with the vector lacking the LysA2 gene (SC-EV), were employed to test whether or not secreted LysA2 directly from S. cerevisiae can prevent contamination (Table 3). The fermentation profiles of both S. cerevisiae strains were similar when LF was not added. In the presence of LF, The LysA2 secreting S. cerevisiae strain fermented glucose and produced almost 29% more ethanol than the S. cerevisiae strain with the empty vector. It is likely that the S. cerevisiae strain with the empty vector failed to finish the fermentation due to inhibition by LF. With LF, the lactic acid concentrations in the culture medium were higher than the culture medium with the LysA2 secreting S. cerevisiae. As expected, the LysA2 secreting S. cerevisiae consumed glucose more rapidly, and increased ethanol yield by approximately 9%, while ethanol yield of the empty vector S. cerevisiae strain decreased in the presence of LF (Figure 4).
3.5. Mock Contamination with Sugarcane Juice
Next, we tested the ability of LysA2 secreting S. cerevisiae to prevent the impact of LF contamination in sugarcane juice (Table 4). In the absence of LF contamination, all strains had similar fermentation profiles. LF contamination reduced ethanol yield with the S. cerevisiae empty vector strain (SC-EV) by approximately 25% (0.48 to 0.31 g ethanol/g sugar) at the end of fermentation (48 h). The S. cerevisiae secreting LysA2 strain showed no improvement. Adding pulses of LysA2 (intracellularly produced by P. pastoris) during ethanol fermentation by S. cerevisiae increased ethanol yield by about 29%, compared to the fermentation with no added or secreted LysA2 (0.31 to 0.4 g ethanol/g sugar), and reduced lactic acid production. Integrating the LysA2 expression cassette into the S. cerevisiae genome increased the ethanol yield slightly in sugarcane juice, but not as much as the treatment with purified LysA2 (Table 4).
4. Discussion
In this study, our overall goal was to demonstrate that an endolysin secreting S. cerevisiae could be a useful tool for the bioethanol industry. Specifically, ethanol yield reduction mediated by bacterial contamination with engineered S. cerevisiae secreting the endolysin LysA2 during ethanol fermentation was alleviated. To date, bacteria secreting endolysins have been successfully engineered [37,38], and patent applications claim successful secretion by P. pastoris [39]; however, secretion by S. cerevisiae in ethanol fermentation has not been described.
4.1. Bacteria and Yeast-Based LysA2 Showed the Same Lytic Activity and Spectrum
Initial milestones critical to accomplishing this goal included: (1) endolysin produced intracellularly by yeast with activity comparable to bacterially produced endolysin, and (2) secretion of active endolysin by yeast. We employed P. pastoris, a yeast known for efficient protein expression and secretion [40], as a model system to confirm the lytic activity of yeast expressed LysA2. Isolated LysA2 from P. pastoris had similar lytic activity and lytic spectrum as those of LysA2 from E. coli (Figure 1 and Table 2).
In turbidity reduction assays, LysA2 from P. pastoris showed the highest lytic activity against the strains with the A4α peptidoglycan chemotype [41]. This is consistent with previous characterizations of LysA2 as an endopeptidase that hydrolyzes the bond between the terminal D-alanine and the D-aspartate, linking it to the neighboring peptide chain [25]. Interestingly, not all of strains with that cross linkage were equally sensitive (Table 2). One potential cause for this is interference from S-layer proteins, which are common in many L. brevis strains [42], but not confirmed in L. casei [43]. In addition, the cell wall binding domain may also be an important consideration to strain sensitivity [44,45,46]. As with most endolysins, a strain having the targeted linkage is not sufficient for sensitivity to LysA2 [24].
4.2. Secreted Lysa2 from P. Pastoris Successfully Restores Ethanol Productivity during Bacterial Contamination
A drawback of using yeast for secreting recombinant proteins is the prospect that the proteins will be glycosylated, which can result in impairment or loss of functionality. To investigate whether glycosylation would occur, and if it impacted LysA2 activity, we constructed a P. pastoris strain that secretes LysA2. After induction and suitable time for growth, protein expression in culture media was detectable, but the band was 70 kDa, i.e., much larger than the expected value, 42 kDa. Purification using the His6 tag of LysA2-PS failed, despite numerous attempts. The larger size and failure to bind to the resin is likely due to glycosylation of LysA2 at N or O linkages [47].
Although purification failed, we confirmed that LysA2 is secreted by the lytic activity of the culture supernatant and the lack of such activity in the culture supernatant of the P. pastoris empty vector strain. Concentration of the LysA2 containing supernatant resulted in increased lytic activity against L. casei (Figure 2), which is consistent with the dose response we saw with the purified, intracellularly produced LysA2 (Figure 1). Adding purified LysA2 and concentrated supernatant containing LysA2 improved ethanol yield during fermentations contaminated with L. fermentum. This species has been shown to reduce ethanol production in previous studies [3,11,48], and surveys of biofuel contaminants frequently identify L. fermentum [2,4,5]. These results are consistent with previous studies that found that uncontrolled LAB contamination decreases ethanol yield [3,13,48,49].
4.3. S. cerevisiae Secreting LysA2 is Effective Against Bacterial Contamination in YPD Medium
Compared with P. pastoris strains, S. cerevisiae strains may hyper-glycosylate proteins, and add terminal mannoses in a bond which may be allergenic [50]. In addition, increasing secretion yield from S. cerevisiae typically requires complex optimization steps [51]. Despite these challenges, because S. cerevisiae is the most commonly used species for bioethanol production, we wanted to study the question of whether S. cerevisiae D452-2 with the LysA2 secreting plasmid could mitigate the impact of LAB contamination. In simulated contaminations in YPD60, the LysA2 secreting S. cerevisiae strain provided higher ethanol productivity and yield, compared to the S. cerevisiae empty vector strain when challenged with L. fermentum (Table 4 and Figure 4). This result was consistent with our experiment controlling contaminants with P. pastoris produced LysA2 (Figure 3). The ethanol productivity and yield data demonstrate that, in our system, secreted LysA2 from S. cerevisiae could increase ethanol productivity by inhibiting bacterial growth. However, even with LysA2 secretion, the contamination reduced ethanol yield compared to the uncontaminated cultures suggests that improvements in LysA2 secretion could be beneficial.
4.4. S. cerevisiae Secreting LysA2 Fails to Protect Against Bacterial Contamination in Sugarcane Juice
To test the impact of LysA2 on the inhibition of bacterial growth in media similar to bioethanol production in Brazil, contaminations using diluted sugarcane juice as the fermentation substrate were performed. Sugarcane juice, a critical feedstock for the ethanol fermentation in Brazil [2,52], contains primarily sucrose, along with lesser amounts of other reducing sugars and organic material and minerals which vary between batches [53]. Unfortunately, in sugarcane juice, secreting LysA2 by S. cerevisiae did not provide a notable improvement of ethanol yield when challenged with LF contamination. We subsequently moved the LysA2 expression cassette into the S. cerevisiae genome, to determine if plasmid instability was an issue. The genome integrated LysA2 strain did show a slight increase of ethanol production and yield. However, adding pulses of 50 μg/mL of intracellularly produced LysA2 from P. pastoris improved ethanol production, confirming that LysA2 is functional in sugarcane juice. S. cerevisiae D452-2 grew very poorly in sugarcane juice (48 h to consume 60 g sucrose; Table 4), especially compared to its growth in YPD (24 h to consume 62 g glucose; Table 3). This indicates that the S. cerevisiae secretion process for LysA2 is not enough to ameliorate bacterial contamination; hence, improvement of the expression level is required. Multicopy integration of the LysA2 gene into the S. cerevisiae genome [2,54] could result in improved contaminant control in sucrose fermentations. Alternatively, using an inducible promoter, instead of expressing the protein constitutively (which may place undue stress on the host), may improve contamination control with the secreted LysA2. Regardless, we recommend that future work focus on improving the lytic spectrum of LysA2, and/or on identifying new endolysins with a suitable lytic spectrum, so that more of the problematic LABs are controlled by this treatment.
5. Conclusions
Both P. pastoris and S. cerevisiae are capable of producing the endolysin LysA2 in its active form. The intracellularly produced LysA2 from P. pastoris showed activity comparable to bacterially produced LysA2. LAB with the A4α peptidoglycan chemotype (L-Lys-D-Asp crosslinkage) were the most sensitive to LysA2, though a few from that chemotype were insensitive. In our experiments, purified Pichia-expressed LysA2 successfully improved ethanol productivity and yields in glucose (YPD60) and sucrose-based (sugarcane juice) ethanol fermentation in the presence of a susceptible LAB contaminant. LysA2 secreting S. cerevisiae was also able to control bacterial contamination during fermentation in YPD60, but not in sugar cane juice. Secretion of LysA2 by the fermenting yeast, or adding it in purified form, is a promising alternative tool for controlling LAB contamination during ethanol fermentation. In an optimized process, both options could supplement or replace the currently used antibiotics, or the acidic washing step which inhibits yeast productivity [35,52,55].
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4915/10/6/281/s1, Figure S1: Constructed vector maps for LysA2 expression in this study. Figure S2: The LysA2 secretion by integrated LysA2 gene into S. cerevisiae.
Author Contributions
J.-S.K. created the overall objectives and design with the advice of M.J.M. and Y.-S.J. Research and optimization of experiments was done by J.-S.K. and M.A.D. J.-S.K. performed the experiments, consolidated and analyzed the data. J.-S.K. and M.A.D. wrote the manuscript with critical review provided by M.J.M. All authors have read and approve this draft of the manuscript.
Acknowledgments
This project was funded by a grant from the Energy Biosciences Institute. We thank Christine Liu, Josh Warren and Eun-Joong Oh for technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Renewable Fuels Association (RFA). Going Global—2015 Ethanol Industry Outlook; RFA: Washington, DC, USA, 2015; p. 4. [Google Scholar]
- Amorim, H.V.; Lopes, M.L.; de Castro Oliveira, J.V.; Buckeridge, M.S.; Goldman, G.H. Scientific challenges of bioethanol production in Brazil. Appl. Microbiol. Biotechnol. 2011, 91, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, K.M.; Liu, S.; Leathers, T.D.; Worthington, R.E.; Rich, J.O. Modeling bacterial contamination of fuel ethanol fermentation. Biotechnol. Bioeng. 2009, 103, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Bischoff, K.M.; Gill, J.J.; Mire-Criscione, M.D.; Berry, J.D.; Young, R.; Summer, E.J. Bacteriophage application restores ethanol fermentation characteristics disrupted by Lactobacillus fermentum. Biotechnol. Biofuels 2015, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Lucena, B.T.L.; dos Santos, B.M.; Moreira, J.L.S.; Moreira, A.P.B.; Nunes, A.C.; Azevedo, V.; Miyoshi, A.; Thompson, F.L.; de Morais, M. Diversity of lactic acid bacteria of the bioethanol process. BMC Microbiol. 2010, 10, 298. [Google Scholar] [CrossRef] [PubMed]
- Jacques, K.A.; Lyons, T.P.; Kelsall, D.R. The Alcohol Textbook: A Reference for the Beverage, Fuel and Industrial Alcohol Industries; Nottingham University Press: Nottingham, UK, 2003. [Google Scholar]
- Scott, Lewis. Options Expand for Effective Bacterial Control in Ethanol Production. Available online: http://ethanolproducer.com/articles/13871/options-expand-for-effective-bacterial-control-in-ethanol-production (accessed on 10 November 2016).
- Skinner, K.A.; Leathers, T.D. Bacterial contaminants of fuel ethanol production. J. Ind. Microbiol. Biotechnol. 2004, 31, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Guzzo, F.; Cappello, M.S.; Azzolini, M.; Tosi, E.; Zapparoli, G. The inhibitory effects of wine phenolics on lysozyme activity against lactic acid bacteria. Int. J. Food Microbiol. 2011, 148, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.; O’Sullivan, O.; Mills, S.; McAuliffe, O.; Ross, R.P.; Neve, H.; Coffey, A. Genome sequence of the phage clP1, which infects the beer spoilage bacterium Pediococcus damnosus. Gene 2012, 504, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.L.; Pettersson, B.; Priest, F.G. Characterization of lactobacilli from Scotch malt whisky distilleries and description of Lactobacillus ferintoshensis sp. nov., a new species isolated from malt whisky fermentations. Microbiology 2001, 147, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.Y.A.; Souto, B.M.; Tupinambá, D.D.; Bergmann, J.C.; Kyaw, C.M.; Kruger, R.H.; Barreto, C.C.; Quirino, B.F. Microbial diversity in sugarcane ethanol production in a Brazilian distillery using a culture-independent method. J. Ind. Microbiol. Biotechnol. 2015, 42, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Schell, D.J.; Dowe, N.; Ibsen, K.N.; Riley, C.J.; Ruth, M.F.; Lumpkin, R.E. Contaminant occurrence, identification and control in a pilot-scale corn fiber to ethanol conversion process. Bioresour. Technol. 2007, 98, 2942–2948. [Google Scholar] [CrossRef] [PubMed]
- Basso, T.O.; de Kok, S.; Dario, M.; do Espirito-Santo, J.C.A.; Müller, G.; Schlölg, P.S.; Silva, C.P.; Tonso, A.; Daran, J.-M.; Gombert, A.K.; et al. Engineering topology and kinetics of sucrose metabolism in Saccharomyces cerevisiae for improved ethanol yield. Metab. Eng. 2011, 13, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, K.M.; Skinner-Nemec, K.A.; Leathers, T.D. Antimicrobial susceptibility of Lactobacillus species isolated from commercial ethanol plants. J. Ind. Microbiol. Biotechnol. 2007, 34, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.O.; Leathers, T.D.; Nunnally, M.S.; Bischoff, K.M. Rapid evaluation of the antibiotic susceptibility of fuel ethanol contaminant biofilms. Bioresour. Technol. 2011, 102, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Oliva Neto, P.D.; Lima, F.A.D.; Silva, K.C.D.; Silva, D.F.D.; Carvalho, A.F.A.; Santos, C.D. Chemical inhibition of the contaminant Lactobacillus fermentum from distilleries producing fuel bioethanol. Braz. Arch. Biol. Technol. 2014, 57, 441–447. [Google Scholar] [CrossRef]
- Stroppa, C.T.; Andrietta, M.G.S.; Andrietta, S.R.; Steckelberg, C.; Serra, G. Use of Penicillin and Monensin to Control Bacterial Contamination of Brazilian Alcohol Fermentations. Int. Sugar J. 2000, 102, 78–82. [Google Scholar]
- Murphree, C.A.; Heist, E.P.; Moe, L.A. Antibiotic Resistance Among Cultured Bacterial Isolates from Bioethanol Fermentation Facilities Across the United States. Curr. Microbiol. 2014, 69, 277–285. [Google Scholar] [CrossRef] [PubMed]
- De Alwis, H.; Heller, D.N. Multiclass, multiresidue method for the detection of antibiotic residues in distillers grains by liquid chromatography and ion trap tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 3076–3084. [Google Scholar] [CrossRef] [PubMed]
- Summer, E.J.; Liu, M. Application of Bacteriophages for the Control of Unwanted Bacteria in Biofuel Production Mediated by Bacterial Reactive Agents. U.S. Patents US13,423,010, 2012. [Google Scholar]
- Becker, S.C.; Foster-Frey, J.; Donovan, D.M. The phage K lytic enzyme LysK and lysostaphin act synergistically to kill MRSA. FEMS Microbiol. Lett. 2008, 287, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, P.A.; Roach, D.R.; Donovan, D.M.; Hughes, S.R.; Bischoff, K.M. Saccharomyces cerevisiae expressing bacteriophage endolysins reduce Lactobacillus contamination during fermentation. Biotechnol. Biofuels 2014, 7, 104. [Google Scholar] [CrossRef]
- Van Tassell, M.L.; Angela Daum, M.; Kim, J.-S.; Miller, M.J. Creative lysins: Listeria and the engineering of antimicrobial enzymes. Curr. Opin. Biotechnol. 2016, 37, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Ribelles, P.; Rodríguez, I.; Suárez, J.E. LysA2, the Lactobacillus casei bacteriophage A2 lysin is an endopeptidase active on a wide spectrum of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2012, 94, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Khatibi, P.A.; Bischoff, K.M.; Hughes, S.R.; Donovan, D.M. Bacteriophage-encoded lytic enzymes control growth of contaminating Lactobacillus found in fuel ethanol fermentations. Biotechnol. Biofuels 2013, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar] [PubMed]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; de los Reyes-Gavilan, C.G.; Caso, J.L.; Suarez, J.E. Characterization of o393-A2, a bacteriophage that infects Lactobacillus casei. Microbiology 1994, 140, 2585–2590. [Google Scholar] [CrossRef]
- Parekh, R.N.; Shaw, M.R.; Wittrup, K.D. An Integrating Vector for Tunable, High Copy, Stable Integration into the Dispersed Ty δ Sites of Saccharomyces cerevisiae. Biotechnol. Prog. 1996, 12, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Schiestl, R.H. Frozen competent yeast cells that can be transformed with high efficiency using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.S.; Alper, H.; Yang, Y.T.; Stephanopoulos, G. Improvement of Xylose Uptake and Ethanol Production in Recombinant Saccharomyces cerevisiae through an Inverse Metabolic Engineering Approach. Appl. Environ. Microbiol. 2005, 71, 8249–8256. [Google Scholar] [CrossRef] [PubMed]
- Della-Bianca, B.E.; Gombert, A.K. Stress tolerance and growth physiology of yeast strains from the Brazilian fuel ethanol industry. Antonie Leeuwenhoek 2013, 104, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Gombert, A.K.; van Maris, A.J.A. Improving conversion yield of fermentable sugars into fuel ethanol in 1st generation yeast-based production processes. Curr. Opin. Biotechnol. 2015, 33, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.L.; Duarte, W.F.; Freire, A.L.; Dias, D.R.; Eleutherio, E.C.A.; Schwan, R.F. Evaluation of stress tolerance and fermentative behavior of indigenous Saccharomyces cerevisiae. Braz. J. Microbiol. 2013, 44, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Turchi, B.; Mok, K.C.; Taga, M.E.; Miller, M.J. HM2-phage resistant solventogenic Clostridium saccharoperbutylacetonicum N1-4 shows increased exopolysaccharide production. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Gaeng, S.; Scherer, S.; Neve, H.; Loessner, M.J. Gene cloning and expression and secretion of Listeria monocytogenes bacteriophage-lytic enzymes in Lactococcus lactis. Appl. Environ. Microbiol. 2000, 66, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, T.; Lo Curto, R.; Minniti, E.; Narbad, A.; Mayer, M.J. Application of Lactobacillus johnsonii expressing phage endolysin for control of Clostridium perfringens. Lett. Appl. Microbiol. 2014, 59, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Lavigne, R. Volckaert G Antimicrobial Agents. U.S. Patents US13061053, 2009. [Google Scholar]
- Damasceno, L.M.; Huang, C., Jr.; Batt, C.A. Protein secretion in Pichia pastoris and advances in protein production. Appl. Microbiol. Biotechnol. 2012, 93, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Schleifer, K.H.; Kandler, O. Peptidoglycan types of bacterial cell walls and their taxonomic implications. Bacteriol. Rev. 1972, 36, 407–477. [Google Scholar] [PubMed]
- Sleytr, U. Bacterial S-layers. Trends Microbiol. 1999, 7, 253–260. [Google Scholar] [CrossRef]
- Hynönen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.C.; Swift, S.; Korobova, O.; Schischkova, N.; Kopylov, P.; Donovan, D.M.; Abaev, I. Lytic activity of the staphylolytic Twort phage endolysin CHAP domain is enhanced by the SH3b cell wall binding domain. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Eugster, M.R.; Haug, M.C.; Huwiler, S.G.; Loessner, M.J. The cell wall binding domain of Listeria bacteriophage endolysin PlyP35 recognizes terminal GlcNAc residues in cell wall teichoic acid: GlcNAc is the binding ligand for Listeria phage endolysin PlyP35. Mol. Microbiol. 2011, 81, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Dorado, I.; Campillo, N.E.; Monterroso, B.; Hesek, D.; Lee, M.; Páez, J.A.; García, P.; Martínez-Ripoll, M.; García, J.L.; Mobashery, S.; et al. Elucidation of the Molecular Recognition of Bacterial Cell Wall by Modular Pneumococcal Phage Endolysin CPL-1. J. Biol. Chem. 2007, 282, 24990–24999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cereghino, G.P.L.; Cereghino, J.L.; Ilgen, C.; Cregg, J.M. Production of recombinant proteins in fermenter cultures of the yeast Pichia pastoris. Curr. Opin. Biotechnol. 2002, 13, 329–332. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Hynes, S.H.; Thomas, K.C.; Ingledew, W.M. Effects of lactobacilli on yeast-catalyzed ethanol fermentations. Appl. Environ. Microbiol. 1997, 63, 4158–4163. [Google Scholar] [PubMed]
- De Oliva-Neto, P.; Yokoya, F. Evaluation of bacterial contamination in a fed-batch alcoholic fermentation process. World J. Microbiol. Biotechnol. 1994, 10, 697–699. [Google Scholar] [CrossRef] [PubMed]
- Gellissen, G.; Strasser, A.W.M.; Suckow, M. Key and Criteria to the Selection of An Expression Platform. In Production of Recombinant Proteins; Gellissen, G., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2005; pp. 1–5. [Google Scholar]
- Idiris, A.; Tohda, H.; Kumagai, H.; Takegawa, K. Engineering of protein secretion in yeast: Strategies and impact on protein production. Appl. Microbiol. Biotechnol. 2010, 86, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Cavalheiro, A.; Monteiro, G. Solving ethanol production problems with genetically modified yeast strains. Braz. J. Microbiol. 2013, 44, 665–671. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.B.; de Menezes, J.A.S.; de Souza, R.D.F.R.; Dutra, E.D.; de Morais, M.A. Mineral composition of the sugarcane juice and its influence on the ethanol fermentation. Appl. Biochem. Biotechnol. 2015, 175, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Argueso, J.L.; Carazzolle, M.F.; Mieczkowski, P.A.; Duarte, F.M.; Netto, O.V.C.; Missawa, S.K.; Galzerani, F.; Costa, G.G.L.; Vidal, R.O.; Noronha, M.F.; et al. Genome structure of a Saccharomyces cerevisiae strain widely used in bioethanol production. Genome Res. 2009, 19, 2258–2270. [Google Scholar] [CrossRef] [PubMed]
- Basso, L.C.; de Amorim, H.V.; de Oliveira, A.J.; Lopes, M.L. Yeast selection for fuel ethanol production in Brazil. FEMS Yeast Res. 2008, 8, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Purification and lytic activity of intracellularly expressed LysA2 from P. pastoris and E. coli. Intracellularly produced LysA2 from E. coli (LysA2-EI) and P. pastoris (LysA2-PI) were purified and confirmed by protein size and lytic activity. SDS-PAGE analysis shows LysA2-EI and LysA2-PI are approximately the expected size of 42 kDa (A). Lytic activity of LysA2-PI measured by turbidity reduction with L. casei. Lytic activity increased proportionally with increased protein concentration (B). Lytic activity of LysA2-PI (100 nM, 4.22 μg/mL) was compared with LysA2-EI (100 nM, 4.22 μg/mL) using turbidity reduction with L. casei (C).
Figure 1.
Purification and lytic activity of intracellularly expressed LysA2 from P. pastoris and E. coli. Intracellularly produced LysA2 from E. coli (LysA2-EI) and P. pastoris (LysA2-PI) were purified and confirmed by protein size and lytic activity. SDS-PAGE analysis shows LysA2-EI and LysA2-PI are approximately the expected size of 42 kDa (A). Lytic activity of LysA2-PI measured by turbidity reduction with L. casei. Lytic activity increased proportionally with increased protein concentration (B). Lytic activity of LysA2-PI (100 nM, 4.22 μg/mL) was compared with LysA2-EI (100 nM, 4.22 μg/mL) using turbidity reduction with L. casei (C).
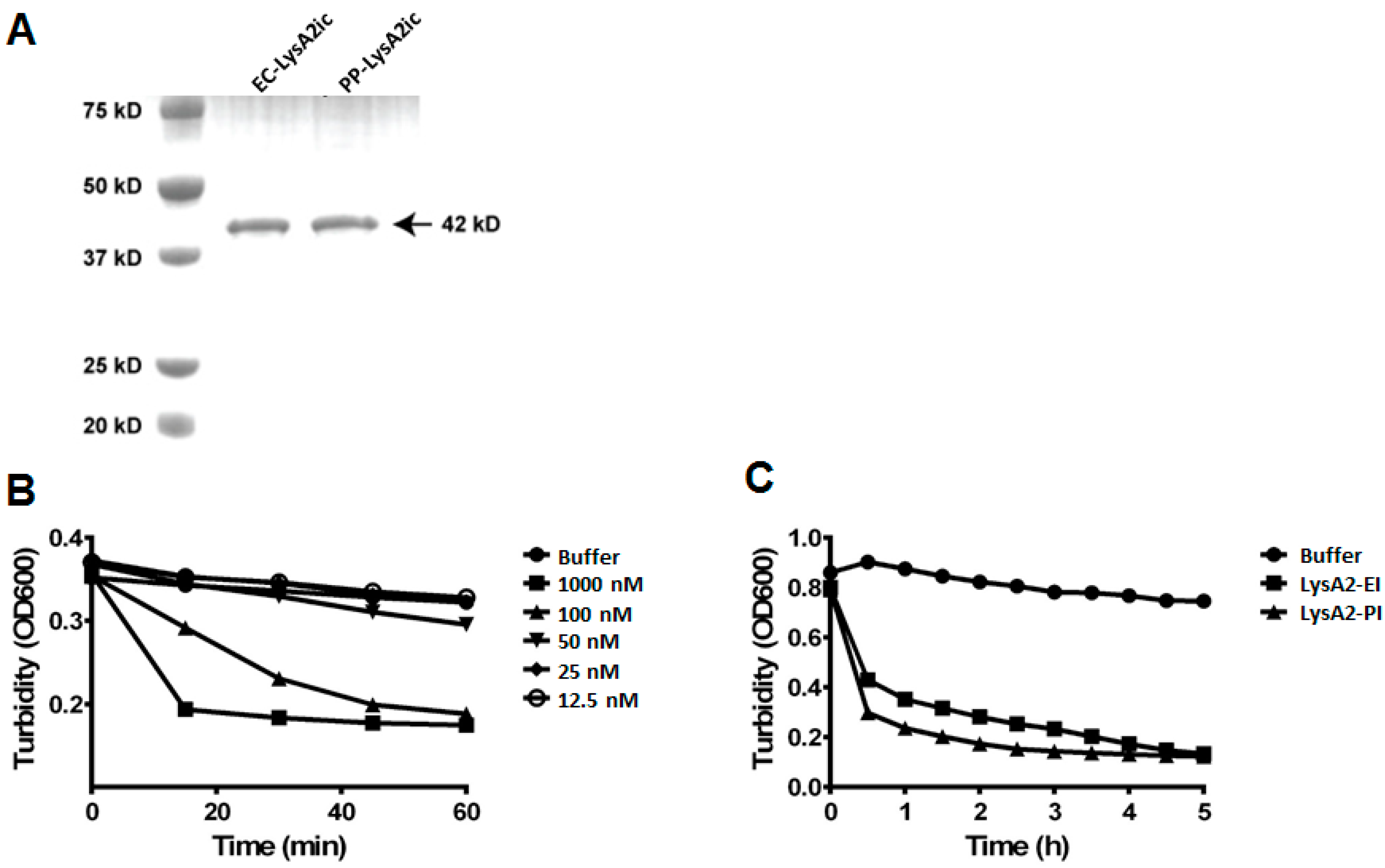
Figure 2.
Activity of supernatant containing secreted LysA2 from P. pastoris. Secreted LysA2 from P. pastoris (PP-LysA2s) was confirmed by SDS-PAGE analysis of the supernatant. The LysA2 in the supernatant increased with increased incubation time. The molecular weight (~72 kDa) was larger than the expected size (42 kDa) which could be due to glycosylation (A). Lytic activity was confirmed by turbidity reduction with L. casei treated with concentrated (50×) and unconcentrated culture supernatant from the P. pastoris secreting LysA2 strain (PP-LysA2s) compared to the culture supernatant from the P. pastoris with the empty vector strain (PP-EV) (B).
Figure 2.
Activity of supernatant containing secreted LysA2 from P. pastoris. Secreted LysA2 from P. pastoris (PP-LysA2s) was confirmed by SDS-PAGE analysis of the supernatant. The LysA2 in the supernatant increased with increased incubation time. The molecular weight (~72 kDa) was larger than the expected size (42 kDa) which could be due to glycosylation (A). Lytic activity was confirmed by turbidity reduction with L. casei treated with concentrated (50×) and unconcentrated culture supernatant from the P. pastoris secreting LysA2 strain (PP-LysA2s) compared to the culture supernatant from the P. pastoris with the empty vector strain (PP-EV) (B).
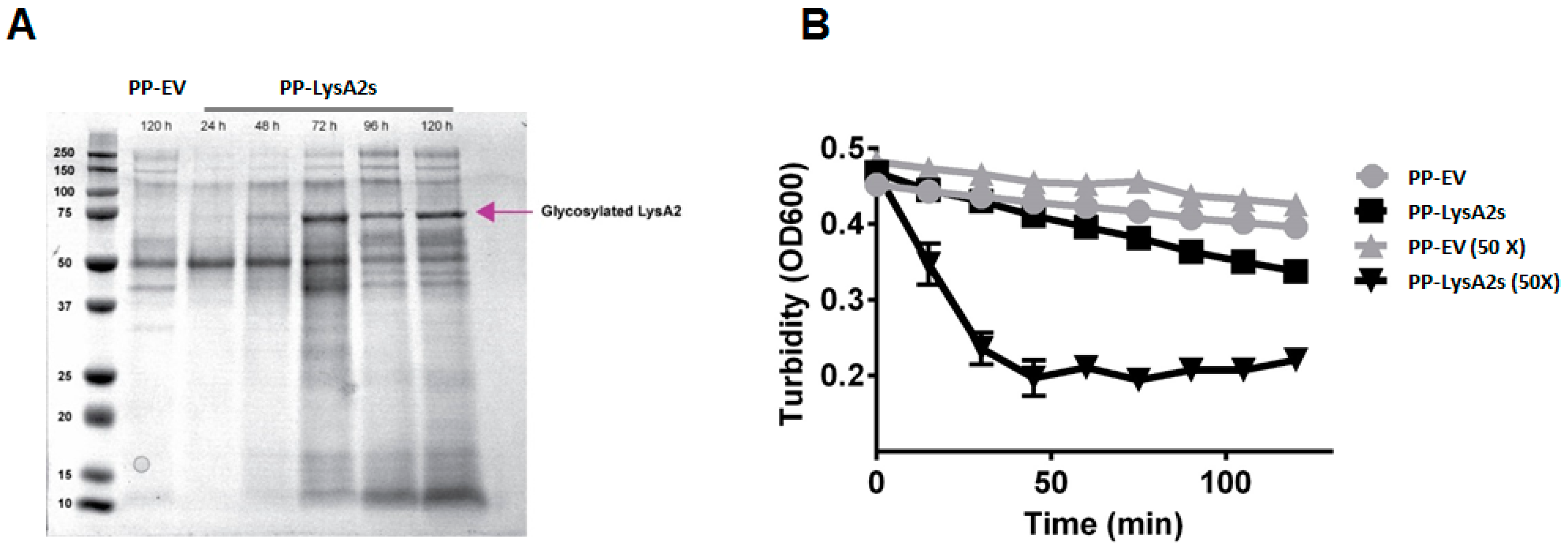
Figure 3.
Activity of secreted LysA2 from P. pastoris compared to purified intracellularly produced LysA2 from P. pastoris in YPD60 fermentations by S. cerevisiae D-452 with an empty vector (SC-EV). In the chart above, lane 1 contains SC-EV only, lanes 2–7 are SC-EV and L. fermentum (LF) and lanes 8–9 contains SC-EV and L. plantarum (LP). The conditions are: (1) no treatment, no contamination; (2) no treatment; (3) purified intracellular LysA2 (LysA2-PI; 100 nM); (4) Supernatant from LysA2 secreting P. pastoris (LysA2-PS); (5) Concentrated (50×) supernatant from LysA2 secreting P. pastoris (LysA2-PS); (6) Supernatant from P. pastoris with empty secretion vector (PS-EV); (7) Concentrated supernatant from P. pastoris with empty secretion vector (PS-EV); and (8) no treatment; (9) purified intracellular LysA2 (LysA2-PI; 100 nM). In a fermentation by S. cerevisiae empty vector (SC-EV) in YPD60, adding the contaminants L. fermentum and L. plantarum decreased ethanol compared to the uncontaminated control. Treatment with purified LysA2 produced intracellularly (LysA2-PI) and concentrated or unconcentrated supernatant from P. pastoris secreting LysA2 (LysA2-PS) restored ethanol production in contaminations with L. fermentum but not with L. plantarum. p values (** p < 0.01) were determined using t- test.
Figure 3.
Activity of secreted LysA2 from P. pastoris compared to purified intracellularly produced LysA2 from P. pastoris in YPD60 fermentations by S. cerevisiae D-452 with an empty vector (SC-EV). In the chart above, lane 1 contains SC-EV only, lanes 2–7 are SC-EV and L. fermentum (LF) and lanes 8–9 contains SC-EV and L. plantarum (LP). The conditions are: (1) no treatment, no contamination; (2) no treatment; (3) purified intracellular LysA2 (LysA2-PI; 100 nM); (4) Supernatant from LysA2 secreting P. pastoris (LysA2-PS); (5) Concentrated (50×) supernatant from LysA2 secreting P. pastoris (LysA2-PS); (6) Supernatant from P. pastoris with empty secretion vector (PS-EV); (7) Concentrated supernatant from P. pastoris with empty secretion vector (PS-EV); and (8) no treatment; (9) purified intracellular LysA2 (LysA2-PI; 100 nM). In a fermentation by S. cerevisiae empty vector (SC-EV) in YPD60, adding the contaminants L. fermentum and L. plantarum decreased ethanol compared to the uncontaminated control. Treatment with purified LysA2 produced intracellularly (LysA2-PI) and concentrated or unconcentrated supernatant from P. pastoris secreting LysA2 (LysA2-PS) restored ethanol production in contaminations with L. fermentum but not with L. plantarum. p values (** p < 0.01) were determined using t- test.
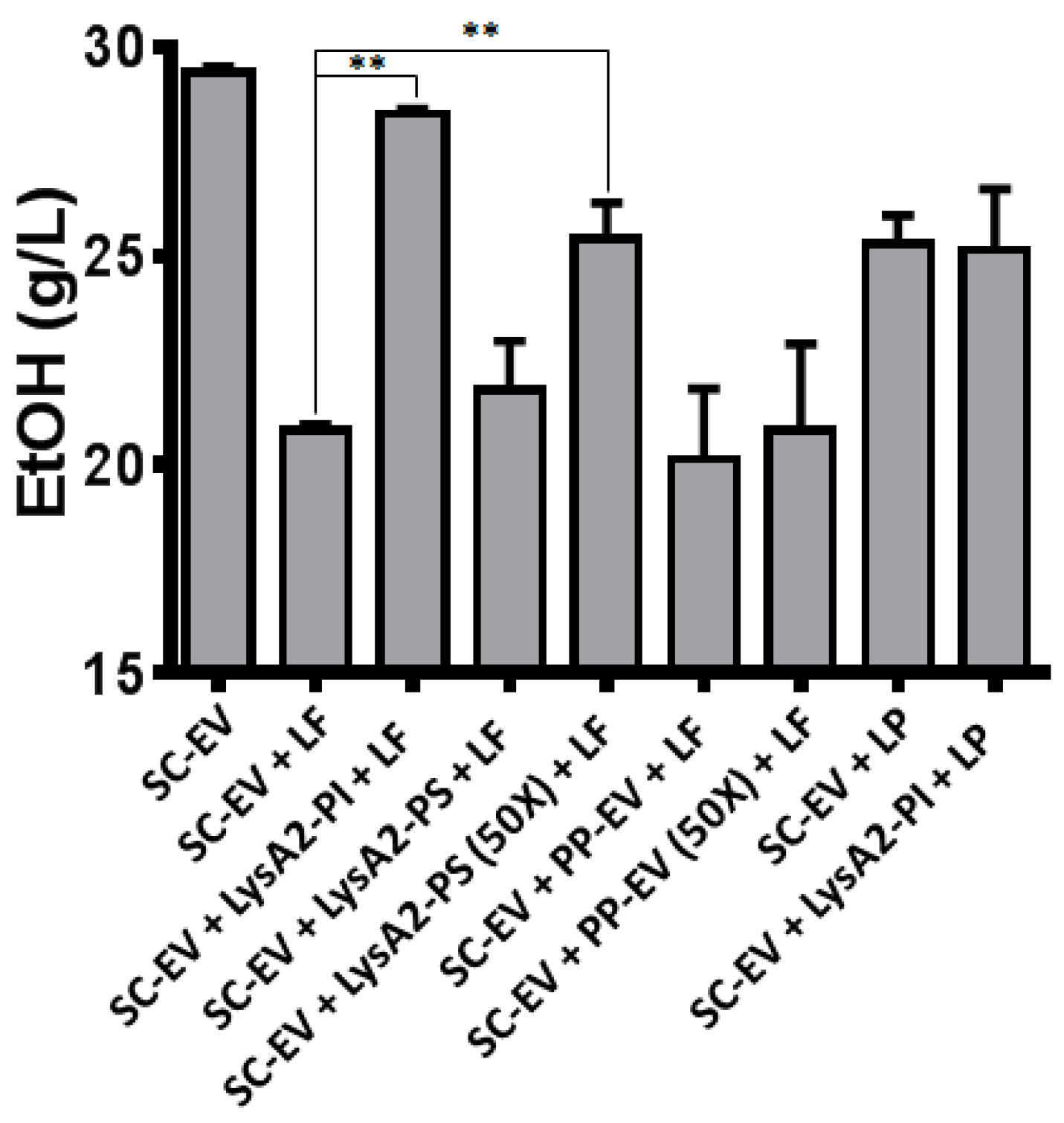
Figure 4.
Impact of LysA2 secretion by S. cerevisiae on ethanol yield. In YPD60, the ethanol yield of S. cerevisiae secreting LysA2 (SC-LysA2s) showed 5% increase compared with S. cerevisiae with the empty vector (SC-EV) when L. fermentum (LF) is present. This result is significantly different. ** < 0.01.
Figure 4.
Impact of LysA2 secretion by S. cerevisiae on ethanol yield. In YPD60, the ethanol yield of S. cerevisiae secreting LysA2 (SC-LysA2s) showed 5% increase compared with S. cerevisiae with the empty vector (SC-EV) when L. fermentum (LF) is present. This result is significantly different. ** < 0.01.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bacterial and yeast host strains used in this study.
| Strains | Description |
|---|---|
| Escherichia coli | BL21 (DE3) |
| Escherichia coli | BL21 (DE3) containing pRSETA_LysA2 |
| Pichia pastoris | GS115 |
| Pichia pastoris | GS115 containing pPICZAa |
| Pichia pastoris | GS115 containing pPICZA_LysA2 |
| Pichia pastoris | GS115 containing pPICZAa_LysA2 |
| Saccharomyces cerevisiae | D452-2 |
| Saccharomyces cerevisiae | D452-2 containing pRS423_LysA2 |
| Saccharomyces cerevisiae | D452-2 containing pITY3_LysA2 1 |
| Saccharomyces cerevisiae | D452-2 empty vector |
1 Chromosomal insertion.
Table 2.
Lytic activity of LysA2 produced intracellularly by P. pastoris against 13 lactic acid bacteria representative of common biofuel contaminants.
Table 2.
Lytic activity of LysA2 produced intracellularly by P. pastoris against 13 lactic acid bacteria representative of common biofuel contaminants.
| Bacterial Contaminants | ATCC # | Peptidoglycan Chemotype | Relative Activity 1 |
|---|---|---|---|
| Aerococcus viridans | 11563 | A1α direct | − |
| Lactobacillus plantarum | 14917 | A1γ mesoDpm | − |
| Lactobacillus casei | 393 | A4α L-Lys-D-Asp | Positive control 2 |
| Enterococcus faecium | 6057 | A4α L-Lys-D-Asp | ++ |
| Enterococcus gallinarum | 49573 | A4α L-Lys-D-Asp | ++ |
| Lactobacillus brevis | 14869 | A4α L-Lys-D-Asp | − |
| Lactobacillus delbrueckii | 9649 | A4α L-Lys-D-Asp | +++ |
| Lactobacillus paracasei | 25598 | A4α L-Lys-D-Asp | ++++ |
| Lactobacillus rhamnosus | 53103 | A4α L-Lys-D-Asp | + |
| Lactococcus lactis | 19257 | A4α L-Lys-D-Asp | + |
| Pediococcus acidilactici | NA | A4α L-Lys-D-Asp | − |
| Pediococcus damnosus | 29358 | A4α L-Lys-D-Asp | − |
| Lactobacillus fermentum | 9338 | A4β L-Orn-D-Asp | ++ |
1 Relative activity measured by turbidity reduction in the presence of 100 nM LysA2 after 1 h incubation as described in the methods section; (−) = 0–10%, (+) = 11–25%, (++) = 26–50%, (+++) = 51–75%, (++++) = 76–100%. 2 L. casei used as the positive control with all data relative to the turbidity reduction of this strain. NA = not available.
Table 3.
Fermentation profile of SC-EV 1 and SC-LysA2s 2 in simulated contamination.
| Contaminants | Yeast Strain | n | Glucose Consumption (g/L ± SEM) | Lactic Acid (g/L ± SEM) | Ethanol (g/L ± SEM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 6 h | 12 h | 24 h | 0 h | 6 h | 12 h | 24 h | 0 h | 6 h | 12 h | 24 h | |||
| None | SC-EV 1 | 3 | 0.0 ± 0.0 | 2.6 ± 0.3 | 13.7 ± 0.3 | 62 ± 0.4 | ND | ND | ND | ND | 0.0 ± 0.0 | 0.6 ± 0.1 | 6.1 ± 0.1 | 26.8 ± 1.8 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 3.2 ± 0.0 | 15.1 ± 0.5 | 62 ± 0.0 | ND | ND | ND | ND | 0.0 ± 0.0 | 0.8 ± 0.3 | 8.0 ± 0.2 | 27.3 ± 0.2 | |
| L. fermentum | SC-EV 1 | 3 | 0.0 ± 0.0 | 4.7 ± 0.0 | 17.1 ± 0.3 | 50.6 ± 0.2 | 0.0 ± 0.0 | 0.7 ± 0.0 | 1.8 ± 0.0 | 3.4 ± 0.0 | 0.0 ± 0.0 | 0.8 ± 0.0 | 6.2 ± 0.3 | 20.8 ± 0.1 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 5.1 ± 0.8 | 19.3 ± 0.4 | 62.0 ± 0.0 | 0.0 ± 0.0 | 0.6 ± 0.0 | 1.2 ± 0.0 | 2.5 ± 0.0 | 0.0 ± 0.0 | 1.1 ± 0.3 | 7.1 ± 0.2 | 26.8 ± 0.2 | |
1S. cerevisiae with empty vector; 2 S. cerevisiae that contains LysA2 secretion vector; ND = not determined.
Table 4.
Fermentation profile of SC-EV 1, SC-LysA2s 2, SC-LysA2s* 3 and purified LysA2 4 with L. fermentum in sugarcane.
Table 4.
Fermentation profile of SC-EV 1, SC-LysA2s 2, SC-LysA2s* 3 and purified LysA2 4 with L. fermentum in sugarcane.
| Contaminant | Yeast Strain | n | Ethanol Production (g/L ± SEM) | Lactic Acid (g/L ± SEM) | Ethanol yield (g/g ± SEM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 12 h | 24 h | 48 h | 0 h | 12 h | 24 h | 48 h | 0 h | 12 h | 24 h | 48 h | |||
| None | SC-EV 1 | 3 | 0.0 ± 0.0 | 6.8 ± 0.5 | 15.5 ± 0.5 | 28.4 ± 0.1 | ND | ND | ND | ND | 0 | 0.46 | 0.48 | 0.48 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 6.9 ± 0.1 | 15.4 ± 0.2 | 27.8 ± 0.0 | ND | ND | ND | ND | 0 | 0.45 | 0.48 | 0.48 | |
| SC-LysA2s* 3 | 3 | 0.0 ± 0.0 | 6.9 ± 0.1 | 16.4 ± 0.3 | 28.8 ± 0.2 | ND | ND | ND | ND | 0 | 0.45 | 0.48 | 0.48 | |
| SC-EV/LysA2-PI 4 | 3 | 0.0 ± 0.0 | 6.7 ± 0.3 | 16.2 ± 0.2 | 27.8 ± 0.2 | ND | ND | ND | ND | 0 | 0.46 | 0.48 | 0.48 | |
| L. fermentum | SC-EV 1 | 3 | 0.0 ± 0.0 | 6.4 ± 0.1 | 13.4 ± 0.4 | 18.6 ± 0.4 | 0.0 ± 0.0 | 1.1 ± 0.0 | 2.8 ± 0.0 | 3.5 ± 0.0 | 0 | 0.25 | 0.27 | 0.31 |
| SC-LysA2s 2 | 3 | 0.0 ± 0.0 | 6.6 ± 0.2 | 13.2 ± 0.2 | 18.4 ± 0.0 | 0.0 ± 0.0 | 1.3 ± 0.0 | 2.6 ± 0.0 | 3.4 ± 0.0 | 0 | 0.24 | 0.27 | 0.32 | |
| SC-LysA2s* 3 | 3 | 0.0 ± 0.0 | 6.2 ± 0.2 | 15.5 ± 0.2 | 20.3 ± 0.0 | 0.0 ± 0.0 | 0.6 ± 0.0 | 2.2 ± 0.0 | 3.1 ± 0.0 | 0 | 0.24 | 0.30 | 0.35 | |
| SC-EV/LysA2-PI 4 | 3 | 0.0 ± 0.0 | 6.0 ± 0.2 | 18.7 ± 0.2 | 23.6 ± 0.3 | 0.0 ± 0.0 | 0.4 ± 0.0 | 1.2 ± 0.0 | 2.2 ± 0.0 | 0 | 0.41 | 0.39 | 0.4 | |
1S. cerevisiae with empty vector; 2 S. cerevisiae that contains LysA2 secretion vector; 3 S. cerevisiae that has the LysA2 secretion vector integrated into the yeast genome; 4 S. cerevisiae with empty vector supplemented with purified, intracellularly produced LysA2 from P. pastoris (LysA2-PI); ND = not determined.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, J.-S.; Daum, M.A.; Jin, Y.-S.; Miller, M.J. Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation. Viruses 2018, 10, 281. https://doi.org/10.3390/v10060281
AMA Style
Kim J-S, Daum MA, Jin Y-S, Miller MJ. Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation. Viruses. 2018; 10(6):281. https://doi.org/10.3390/v10060281
Chicago/Turabian StyleKim, Jun-Seob, M. Angela Daum, Yong-Su Jin, and Michael J. Miller. 2018. "Yeast Derived LysA2 Can Control Bacterial Contamination in Ethanol Fermentation" Viruses 10, no. 6: 281. https://doi.org/10.3390/v10060281
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.