Oral Vaccination with a DNA Vaccine Encoding Capsid Protein of Duck Tembusu Virus Induces Protection Immunity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid, Bacterial Strains, Virus and Ducks
2.2. Construction of DNA Vaccine Plasmids
2.3. Expression of the Capsid Protein from DNA Vaccine in Vitro
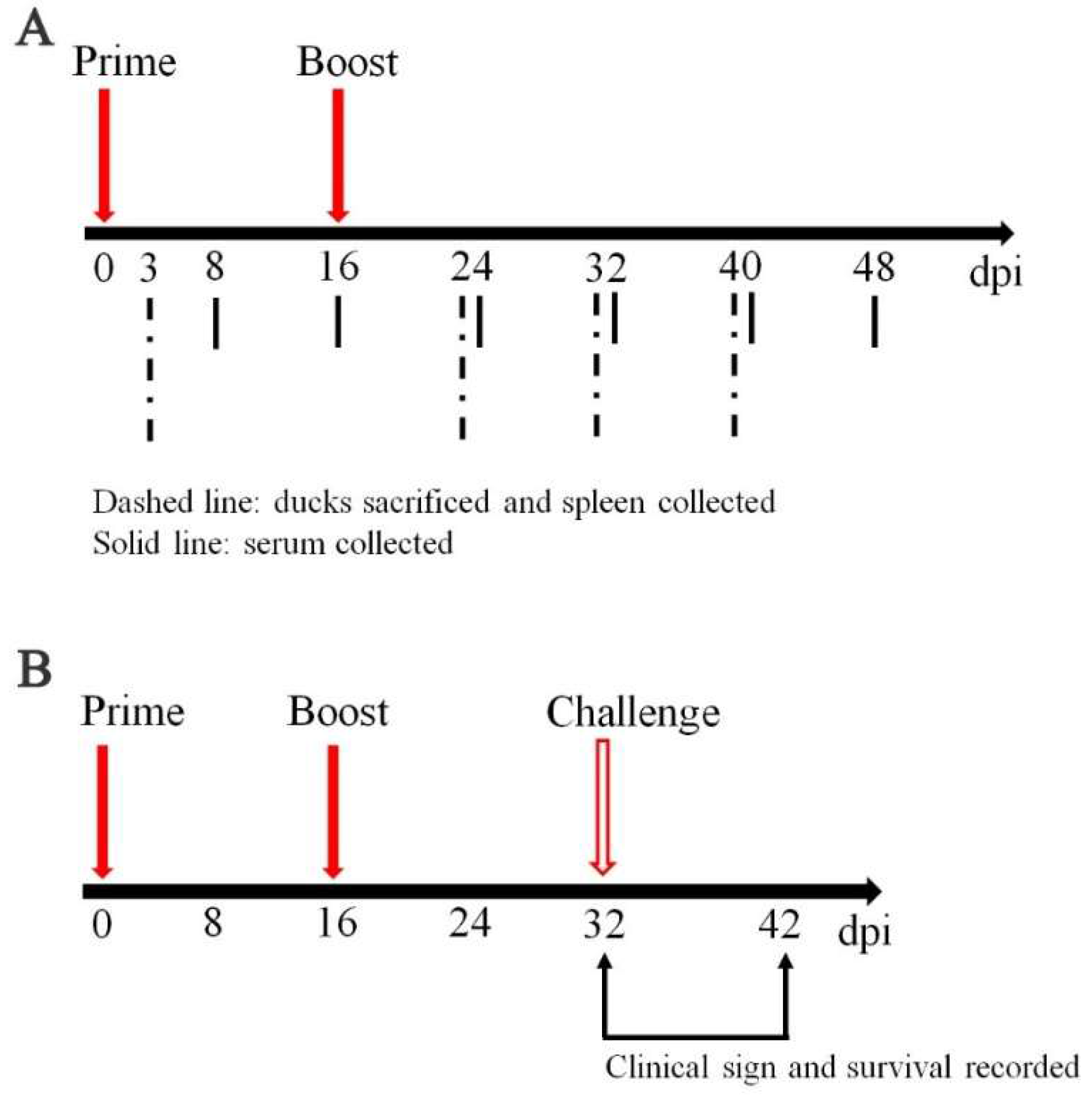
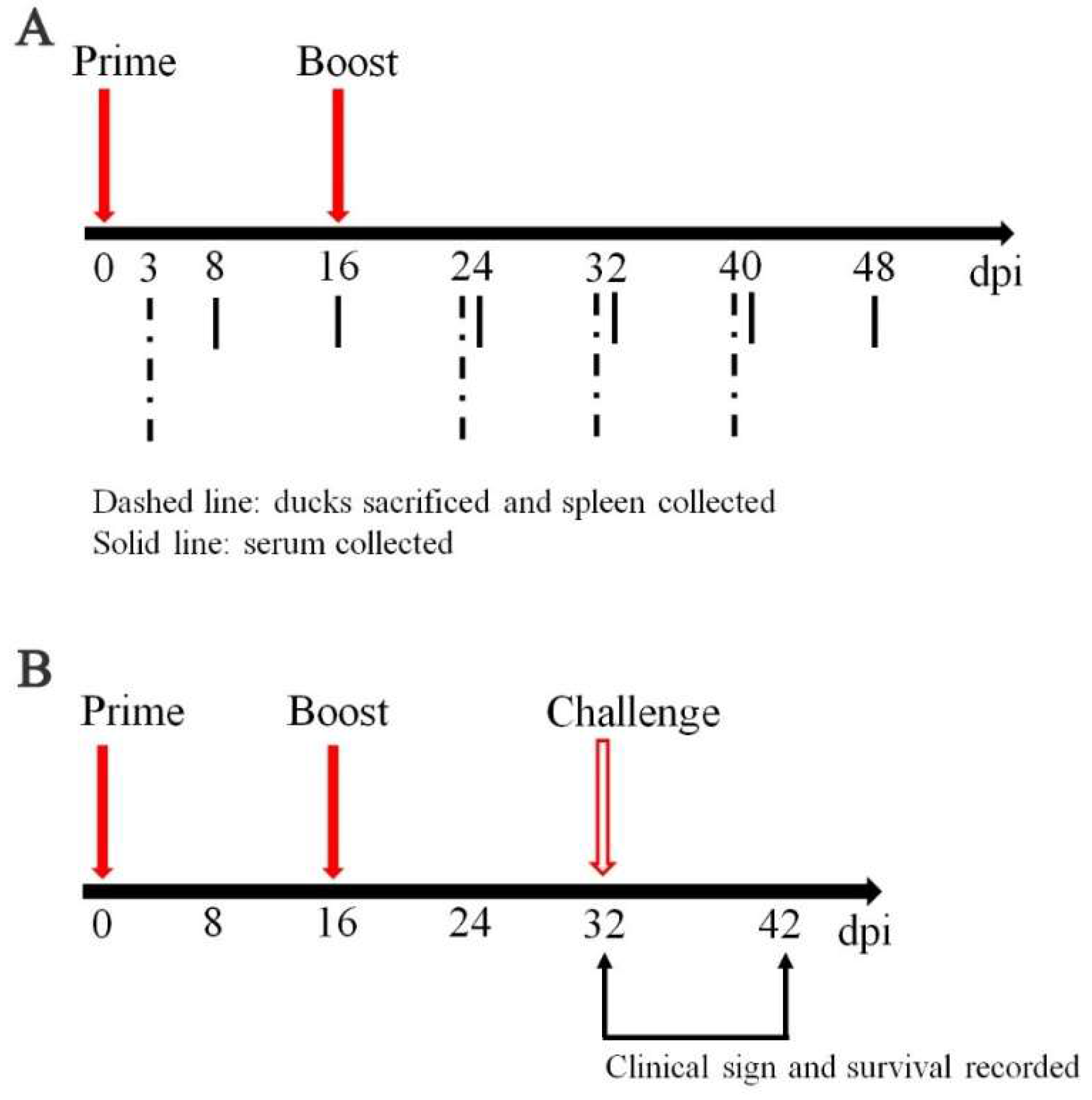
2.4. Sample Collection, Vaccination and Challenge Experiments
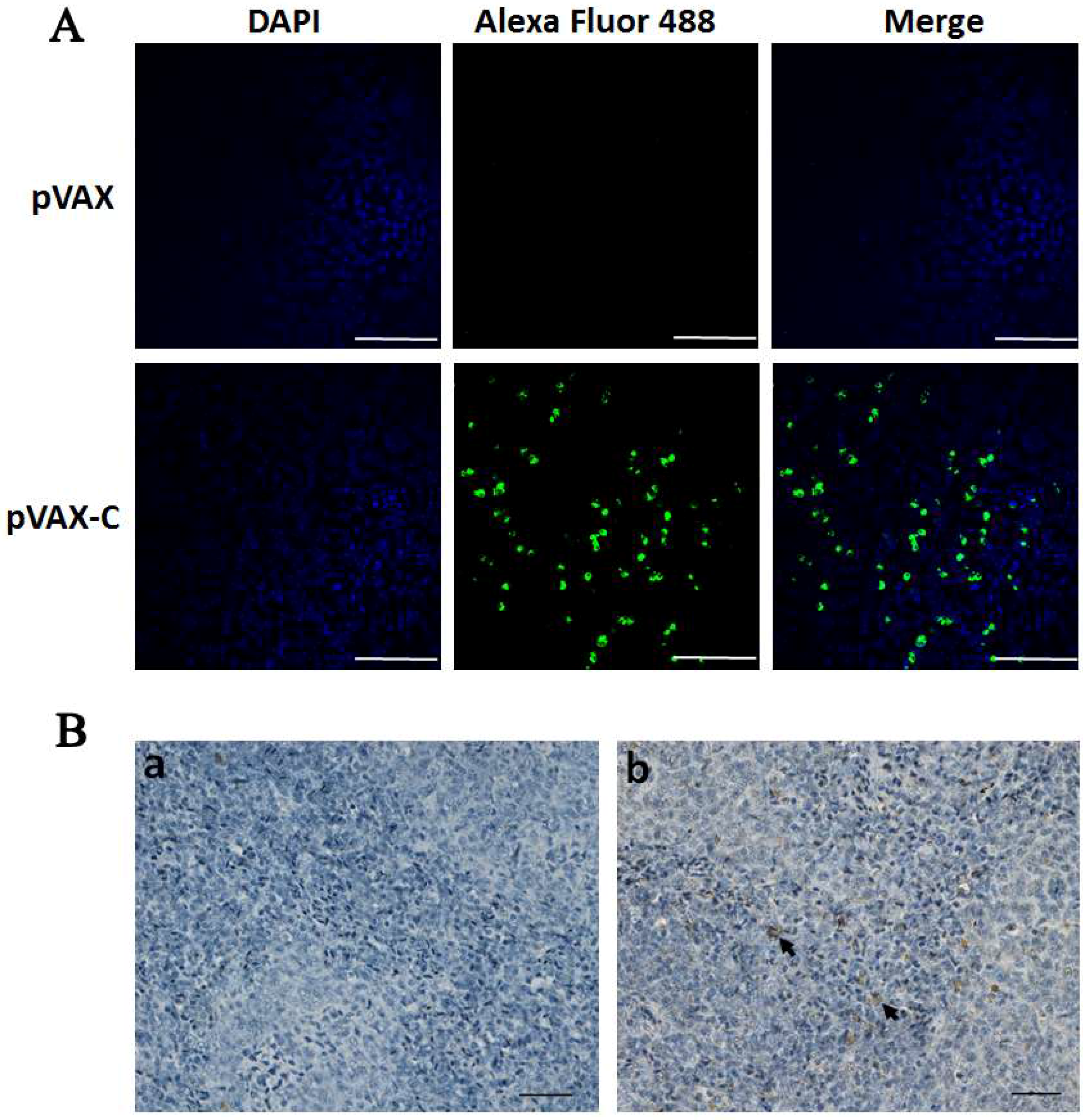
2.5. Expression of the Capsid Protein from DNA Vaccine in Vivo
2.6. Quantitative RT-PCR
2.7. Enzyme-Linked Immunosorbent Assay (dELISA)
2.8. Neutralizing Assay
3. Results
3.1. Expression of the Capsid Protein from the DNA Vaccine in Vitro and in Vivo
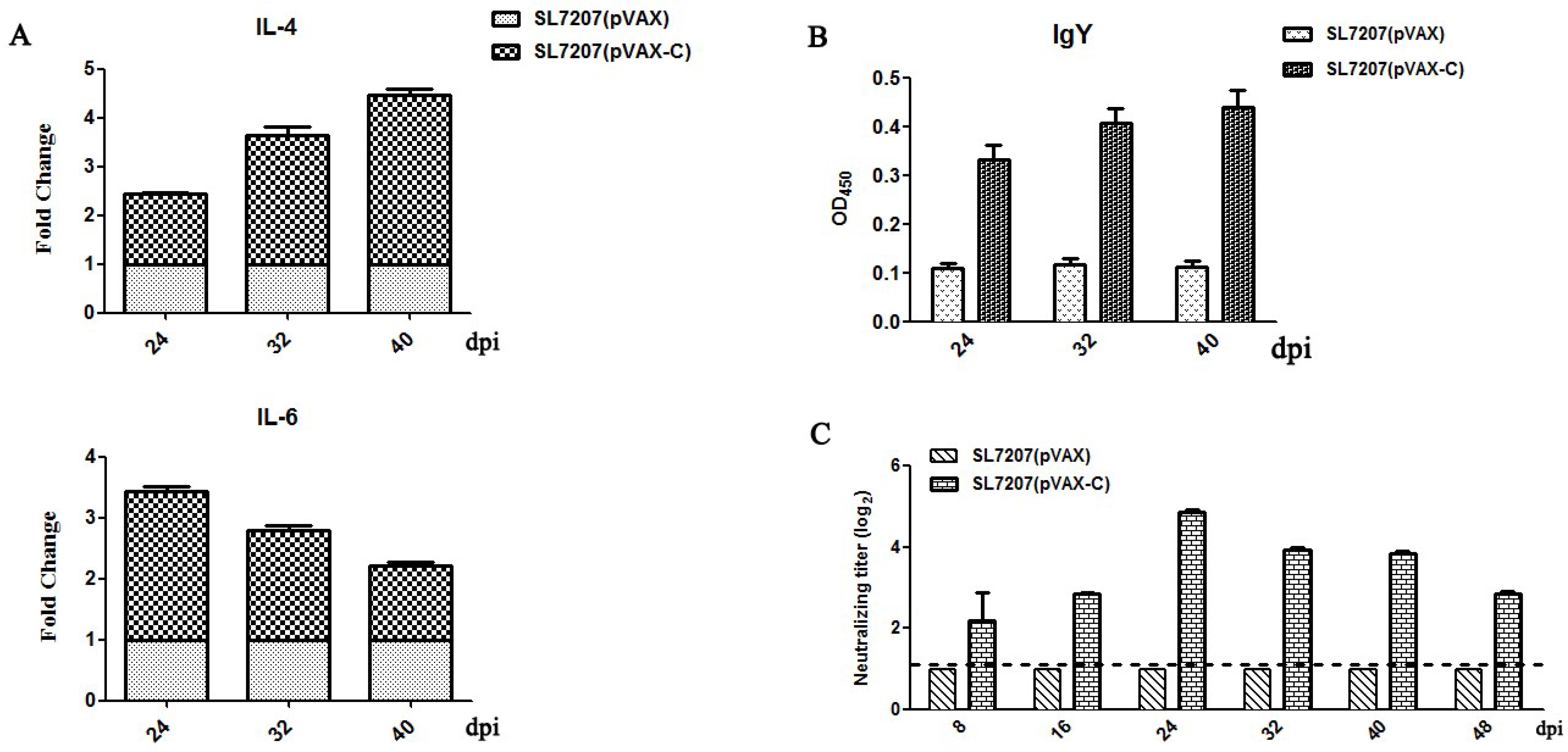
3.2. Cellular and Humoral Immune Responses in Ducks
3.3. Neutralizing Antibodies Responses
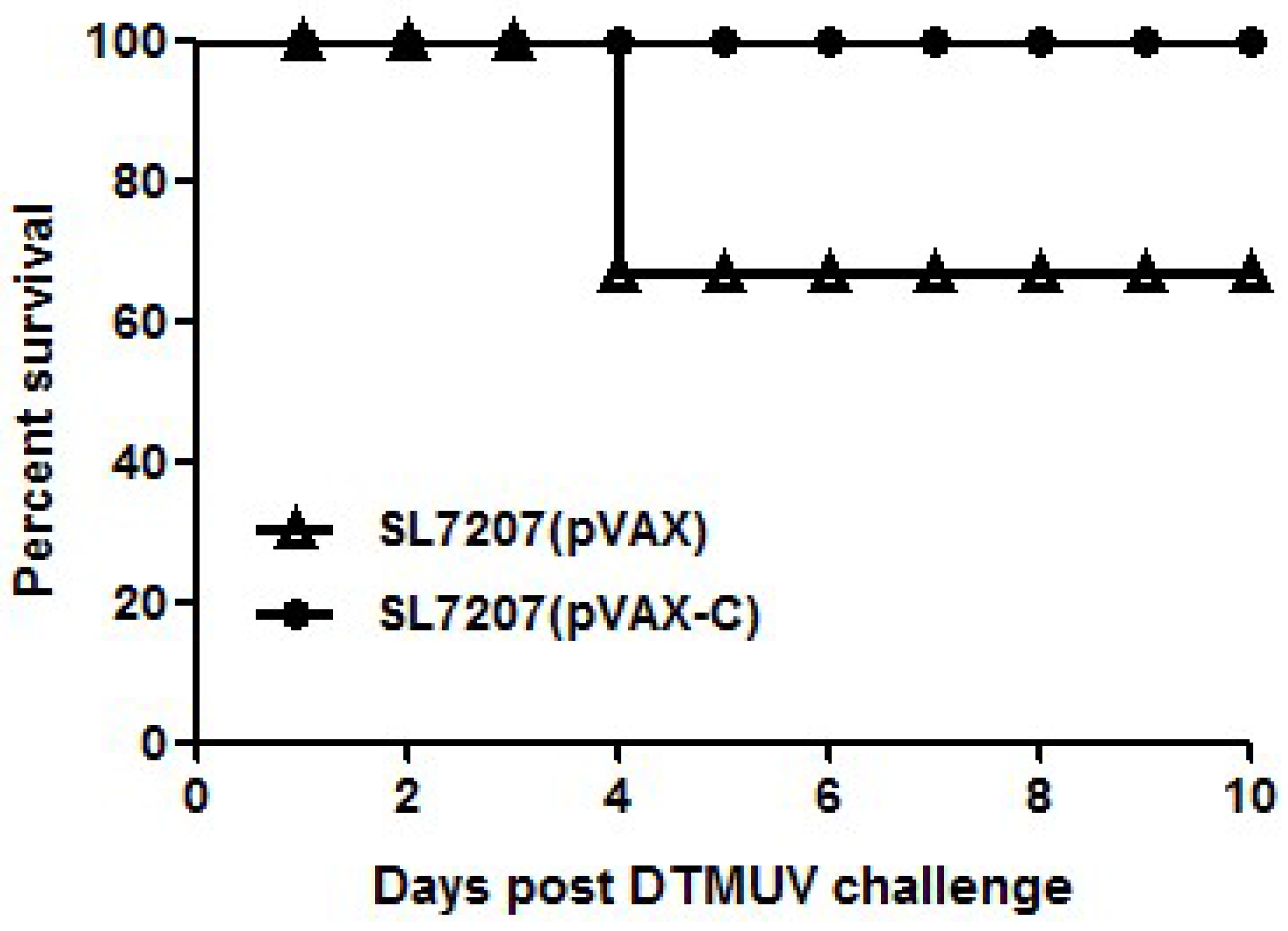
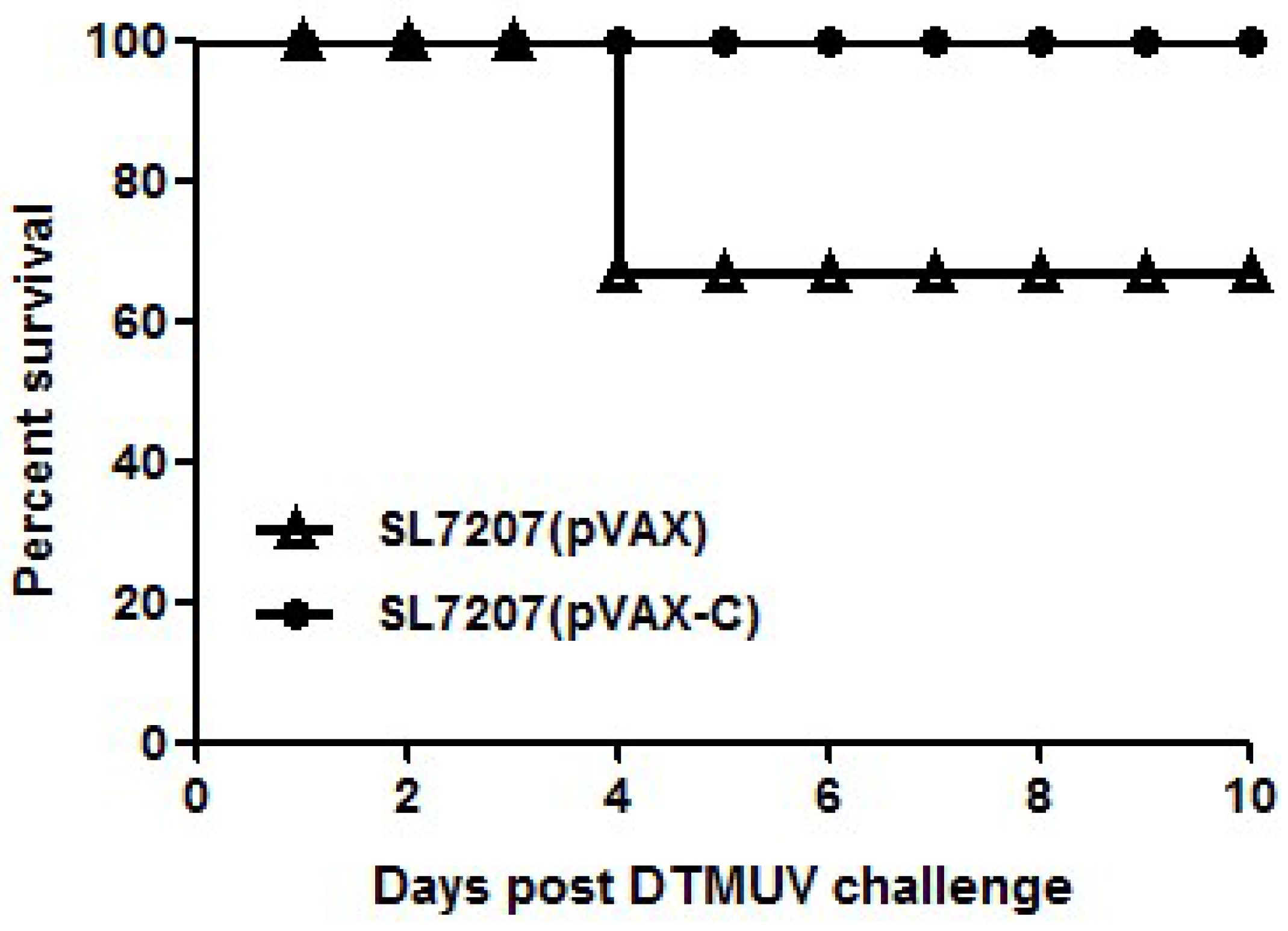
3.4. Protection of Ducklings against DTMUV Challenge
4. Discussion
Acknowledgments
Author contributions
Conflicts of Interest
References
- Thai, K.T.; Nga, T.T.; van Nam, N.; Phuong, H.L.; Giao, P.T.; Hung le, Q.; Binh, T.Q.; van Doornum, G.J.; de Vries, P.J. Incidence of primary dengue virus infections in southern vietnamese children and reactivity against other flaviviruses. Trop. Med. Int. Health 2007, 12, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Qi, J.; Haywood, J.; Shi, Y.; Gao, G.F. Zika virus NS1 structure reveals diversity of electrostatic surfaces among flaviviruses. Nat. Struct. Mol. Biol. 2016, 23, 456–458. [Google Scholar] [CrossRef] [PubMed]
- Villordo, S.M.; Carballeda, J.M.; Filomatori, C.V.; Gamarnik, A.V. RNA structure duplications and flavivirus host adaptation. Trends Microbiol. 2016, 24, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus genome organization, expression, and replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Zhao, Y.; Zhang, X.; Xu, D.; Dai, X.; Teng, Q.; Yan, L.; Zhou, J.; Ji, X.; Zhang, S.; et al. An infectious disease of ducks caused by a newly emerged Tembusu virus strain in mainland China. Virology 2011, 417, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, S.; Mahalingam, S.; Wang, M.; Cheng, A. An updated review of avian-origin Tembusu virus: A newly emerging avian flavivirus. J. Gen. Virol. 2017, 98, 2413–2420. [Google Scholar] [CrossRef] [PubMed]
- Yun, T.; Zhang, D.; Ma, X.; Cao, Z.; Chen, L.; Ni, Z.; Ye, W.; Yu, B.; Hua, J.; Zhang, Y.; et al. Complete genome sequence of a novel flavivirus, duck Tembusu virus, isolated from ducks and geese in China. J. Virol. 2012, 86, 3406–3407. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Diao, Y.; Yu, C.; Gao, X.; Ju, X.; Xue, C.; Liu, X.; Ge, P.; Qu, J.; Zhang, D. Characterization of a Tembusu virus isolated from naturally infected house sparrows (passer domesticus) in northern China. Transbound. Emerg. Dis. 2013, 60, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Diao, Y.; Gao, X.; Yu, C.; Chen, L.; Zhang, D. Analysis of the complete genome of Tembusu virus, a flavivirus isolated from ducks in China. Transbound. Emerg. Dis. 2012, 59, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Gao, X.; Diao, Y.; Feng, Q.; Chen, H.; Liu, X.; Ge, P.; Yu, C. Tembusu virus in human, China. Transbound. Emerg. Dis. 2013, 60, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gao, X.; Xiao, Y.; Liu, S.; Peng, S.; Li, X.; Shi, Y.; Zhang, Y.; Yu, L.; Wu, X.; et al. Development of a live attenuated vaccine candidate against duck Tembusu viral disease. Virology 2014, 450–451, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Z.; Zhang, Q.; Sun, M.; Li, S.; Su, W.; Hu, X.; He, W.; Su, J. Efficacy assessment of an inactivated Tembusu virus vaccine candidate in ducks. Res. Vet. Sci. 2017, 110, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Liu, Y.; Cheng, J.; Liu, Y.; Fan, W.; Cheng, Z.; Niu, X.; Liu, J. Liposomes containing recombinant e protein vaccine against duck Tembusu virus in ducks. Vaccine 2016, 34, 2157–2163. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Liu, L.; Li, X.F.; Ye, Q.; Deng, Y.Q.; Qin, E.D.; Qin, C.F. In vitro and in vivo characterization of chimeric duck Tembusu virus based on Japanese encephalitis live vaccine strain SA14-14-2. J. Gen. Virol. 2016, 97, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Freire, J.M.; Santos, N.C.; Veiga, A.S.; Da Poian, A.T.; Castanho, M.A. Rethinking the capsid proteins of enveloped viruses: Multifunctionality from genome packaging to genome transfection. FEBS J. 2015, 282, 2267–2278. [Google Scholar] [CrossRef] [PubMed]
- Konishi, E.; Pincus, S.; Fonseca, B.A.; Shope, R.E.; Paoletti, E.; Mason, P.W. Comparison of protective immunity elicited by recombinant vaccinia viruses that synthesize e or NS1 of Japanese encephalitis virus. Virology 1991, 185, 401–410. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Li, X.F.; Jiang, T.; Deng, Y.Q.; Zhao, H.; Wang, H.J.; Ye, Q.; Zhu, S.Y.; Qiu, Y.; Zhou, X.; et al. Novel cis-acting element within the capsid-coding region enhances flavivirus viral-RNA replication by regulating genome cyclization. J. Virol. 2013, 87, 6804–6818. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.K.; Ng, M.L. Dephosphorylation of west nile virus capsid protein enhances the processes of nucleocapsid assembly. Microbes Infect. 2011, 13, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Urbanowski, M.D.; Ilkow, C.S.; Hobman, T.C. Modulation of signaling pathways by RNA virus capsid proteins. Cell. Signal. 2008, 20, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Jia, R.; Huang, J.; Shu, B.; Zhu, D.; Liu, Q.; Gao, X.; Lin, M.; Yin, Z.; Wang, M.; et al. Attenuated salmonella typhimurium delivering DNA vaccine encoding duck enteritis virus UL24 induced systemic and mucosal immune responses and conferred good protection against challenge. Vet. Res. 2012, 43, 56. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cheng, A.; Wang, M.; Zhu, D.; Luo, Q.; Liu, F.; Chen, X. Immunohistochemical detection and localization of new type gosling viral enteritis virus in paraformaldehyde-fixed paraffin-embedded tissue. Vet. Immunol. Immunopathol. 2009, 130, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, Y.; Li, R.; Liu, J.; Zhang, J.; Cai, Y.; Liu, S.; Chai, T.; Wei, L. Immune responses of ducks infected with duck Tembusu virus. Front. Microbiol. 2015, 6, 425. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Liu, J.; Jiang, Y.; Zhao, Y.; Li, Q.; Wu, L.; He, X.; Chen, H. The vaccine efficacy of recombinant duck enteritis virus expressing secreted E with or without PrM proteins of duck tembusu virus. Vaccine 2014, 32, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.; McArdle, S.; Gale, M.J., Jr.; Geller, D.A.; Tenoever, B.; Hiscott, J.; Gretch, D.R.; Polyak, S.J. Effects of the hepatitis c virus core protein on innate cellular defense pathways. J. Interferon Cytokine Res. 2004, 24, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Lazo, L.; Hermida, L.; Zulueta, A.; Sanchez, J.; Lopez, C.; Silva, R.; Guillen, G.; Guzman, M.G. A recombinant capsid protein from dengue-2 induces protection in mice against homologous virus. Vaccine 2007, 25, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.B.; Zhang, J.P.; Cheng, L.F.; Zhang, G.W.; Li, Y.; Li, Z.C.; Lu, Z.H.; Zhang, Z.X.; Lu, Y.C.; Zheng, L.H.; et al. Hantavirus Gc induces long-term immune protection via lamp-targeting DNA vaccine strategy. Antivir. Res. 2017, 150, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Tomusange, K.; Wijesundara, D.; Gummow, J.; Wesselingh, S.; Suhrbier, A.; Gowans, E.J.; Grubor-Bauk, B. Mucosal vaccination with a live recombinant rhinovirus followed by intradermal DNA administration elicits potent and protective HIV-specific immune responses. Sci. Rep. 2016, 6, 36658. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cao, S.; Huang, X.; Liu, J.; Tang, Y.; Wen, X. Intragastric administration of attenuated salmonella typhimurium harbouring transmissible gastroenteritis virus (TGEV) DNA vaccine induced specific antibody production. Vaccine 2009, 27, 5035–5040. [Google Scholar] [CrossRef] [PubMed]
- Roland, K.L.; Brenneman, K.E. Salmonella as a vaccine delivery vehicle. Expert Rev. Vaccines 2013, 12, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Flo, J.; Tisminetzky, S.; Baralle, F. Oral transgene vaccination mediated by attenuated salmonellae is an effective method to prevent herpes simplex virus-2 induced disease in mice. Vaccine 2001, 19, 1772–1782. [Google Scholar] [CrossRef]
- Zhang, D.; Huang, X.; Zhang, X.; Cao, S.; Wen, X.; Wen, Y.; Wu, R.; Liang, E. Construction of an oral vaccine for transmissible gastroenteritis virus based on the TGEV N gene expressed in an attenuated salmonella typhimurium vector. J. Virol. Methods 2016, 227, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, J.; Shaozhou, W.; Bai, X.; Zhang, Q.; Hua, R.; Liu, J.H.; Liu, M.; Zhang, Y. Epitope identification and application for diagnosis of duck Tembusu virus infections in ducks. Viruses 2016, 8, 306. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.H.; Chiang, C.C.; Ng, M.L. Immunization of flavivirus West Nile recombinant envelope domain III protein induced specific immune response and protection against West Nile virus infection. J. Immunol. 2007, 178, 2699–2705. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, S.J.; Zeng, W.; Kurane, I.; Ennis, F.A. Identification of two epitopes on the dengue 4 virus capsid protein recognized by a serotype-specific and a panel of serotype-cross-reactive human CD4+ cytotoxic t-lymphocyte clones. J. Virol. 1996, 70, 141–147. [Google Scholar] [PubMed]
- Roohvand, F.; Aghasadeghi, M.R.; Sadat, S.M.; Budkowska, A.; Khabiri, A.R. HCV core protein immunization with montanide/CpG elicits strong Th1/Th2 and long-lived CTL responses. Biochem. Biophys. Res. Commun. 2007, 354, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Duenas-Carrera, S.; Alvarez-Lajonchere, L.; Alvarez-Obregon, J.C.; Herrera, A.; Lorenzo, L.J.; Pichardo, D.; Morales, J. A truncated variant of the hepatitis c virus core induces a slow but potent immune response in mice following DNA immunization. Vaccine 2000, 19, 992–997. [Google Scholar] [CrossRef]
- Kimura, A.; Kishimoto, T. IL-6: Regulator of Treg/Th17 balance. Eur. J. Immunol. 2010, 40, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.A.; Scheller, J.; Rose-John, S. Therapeutic strategies for the clinical blockade of IL-6/gp130 signaling. J. Clin. Investig. 2011, 121, 3375–3383. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T. Interleukin 6 and its receptor: Ten years later. Int. Rev. Immunol. 1998, 16, 249–284. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, S.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Liu, F.; Yang, Q.; Wu, Y.; Sun, K.; et al. Antigen distribution of TMUV and GPV are coincident with the expression profiles of cd8α-positive cells and goose ifngamma. Sci. Rep. 2016, 6, 25545. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.; Pfeffer, K. Impaired germinal centre formation and humoral immune response in the absence of cd28 and interleukin-4. Immunology 2002, 106, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; Ko, S.Y.; Morabito, K.M.; Yang, E.S.; Pelc, R.S.; DeMaso, C.R.; Castilho, L.R.; Abbink, P.; Boyd, M.; Nityanandam, R.; et al. Rapid development of a DNA vaccine for Zika virus. Science 2016, 354, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, G. Current status and future perspectives of DNA vaccine delivery by attenuated intracellular bacteria. Arch. Immunol. Ther. Exp. 2000, 48, 177–182. [Google Scholar]
- Martin, M.; Michalek, S.M.; Katz, J. Role of innate immune factors in the adjuvant activity of monophosphoryl lipid A. Infect. Immun. 2003, 71, 2498–2507. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.R.; Wen, L.; Huang, H.B.; Wang, J.X.; Huang, X.Y.; Peng, K.M.; Liu, H.Z. Lipopolysaccharide stimulation upregulated toll-like receptor 4 expression in chicken cerebellum. Vet. Immunol. Immunopathol. 2015, 166, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Amberg, S.M.; Rice, C.M. Mutagenesis of the NS2B-NS3-mediated cleavage site in the flavivirus capsid protein demonstrates a requirement for coordinated processing. J. Virol. 1999, 73, 8083–8094. [Google Scholar] [PubMed]
- VanBlargan, L.A.; Davis, K.A.; Dowd, K.A.; Akey, D.L.; Smith, J.L.; Pierson, T.C. Context-dependent cleavage of the capsid protein by the West Nile virus protease modulates the efficiency of virus assembly. J. Virol. 2015, 89, 8632–8642. [Google Scholar] [CrossRef] [PubMed]
- Bhuvanakantham, R.; Cheong, Y.K.; Ng, M.L. West Nile virus capsid protein interaction with importin and HDM2 protein is regulated by protein kinase C-mediated phosphorylation. Microbes Infect. 2010, 12, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Bhuvanakantham, R.; Ng, M.L. West Nile virus and dengue virus capsid protein negates the antiviral activity of human Sec3 protein through the proteasome pathway. Cell. Microbiol. 2013, 15, 1688–1706. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Length of PCR Products (bp) | GenBank Number |
|---|---|---|---|
| C (Fw) | TACAGAATTCACTATGGCATCTAACAAAAAACCAGGAAGACCC | 360 | JX196334.1 |
| C (Rev) | TACACTCGAGCTACCCAGCAACTATCGGGAGTAACATA | ||
| Il-4 (Fw) | TCTATCAGAGAAAGACAACAC | 157 | XM_013104023.1 |
| Il-4 (Rev) | GGTGACTATTTCTTTCAAGT | ||
| Il-6 (Fw) | AAGTTGAGTCGCTGTGCT | 120 | JQ728554.1 |
| Il-6 (Rev) | GCTTTGTGAGGAGGGATT | ||
| GAPDH (Fw) | CAAGGCTGAGAATGGGAAAC | 171 | GU564233.1 |
| GAPDH (Rev) | CTGCCCACTTGATGTTGC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Shen, H.; Jia, R.; Wang, M.; Chen, S.; Zhu, D.; Liu, M.; Zhao, X.; Yang, Q.; Wu, Y.; et al. Oral Vaccination with a DNA Vaccine Encoding Capsid Protein of Duck Tembusu Virus Induces Protection Immunity. Viruses 2018, 10, 180. https://doi.org/10.3390/v10040180
Huang J, Shen H, Jia R, Wang M, Chen S, Zhu D, Liu M, Zhao X, Yang Q, Wu Y, et al. Oral Vaccination with a DNA Vaccine Encoding Capsid Protein of Duck Tembusu Virus Induces Protection Immunity. Viruses. 2018; 10(4):180. https://doi.org/10.3390/v10040180
Chicago/Turabian StyleHuang, Juan, Haoyue Shen, Renyong Jia, Mingshu Wang, Shun Chen, Dekang Zhu, Mafeng Liu, Xinxin Zhao, Qiao Yang, Ying Wu, and et al. 2018. "Oral Vaccination with a DNA Vaccine Encoding Capsid Protein of Duck Tembusu Virus Induces Protection Immunity" Viruses 10, no. 4: 180. https://doi.org/10.3390/v10040180