Broad-Spectrum Antiviral Activity of an Ankyrin Repeat Protein on Viral Assembly against Chimeric NL4-3 Viruses Carrying Gag/PR Derived from Circulating Strains among Northern Thai Patients

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viral RNA Extraction and Preparation of the Insert DNA Fragment Encoding the Gag/PR Protein
2.3. Preparation of Viral Vector Backbone for Chimeric HIV-1 Construction
2.4. Production and Titration of Chimeric HIV-1
2.5. Production of Pseudotyped Lentiviral Vector to Transfer AnkGAG1D4 or AnkA32D3 Coding Genes into SupT1 Cells
2.6. Production of SupT1 Cells Stably Expressing AnkGAG1D4 or AnkA32D3
2.7. Inhibitory Effects of AnkGAG1D4 Against Gag/PR Chimeric HIV-1
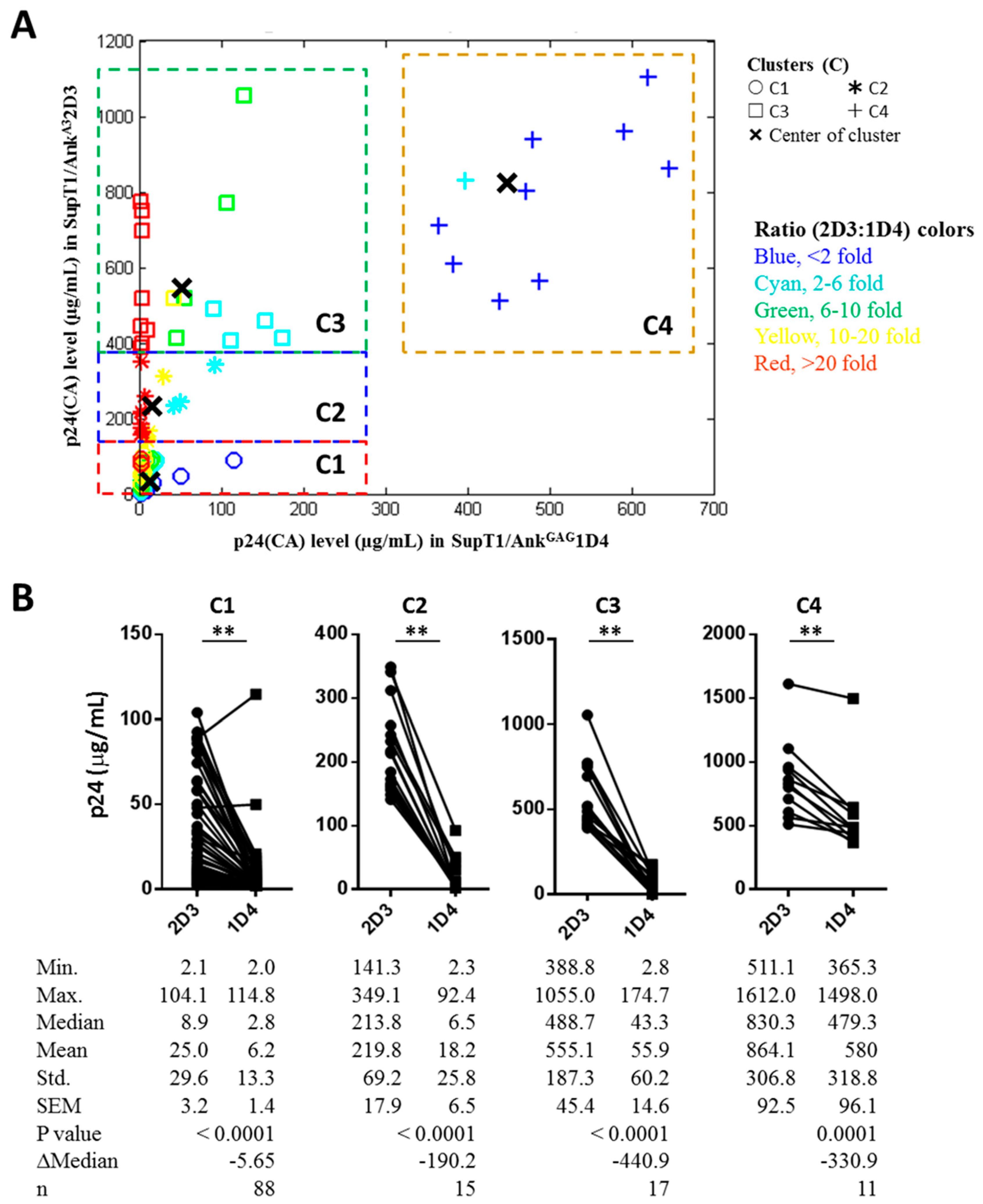
2.8. K-Means Clustering Method
2.9. Analysis of Capsid p24CA DNA Sequences
2.10. Statistical Analysis
2.11. Study Subjects
3. Results
3.1. Establishment of SupT1 Cells Stably Expressing AnkGAG1D4 and AnkA32D3
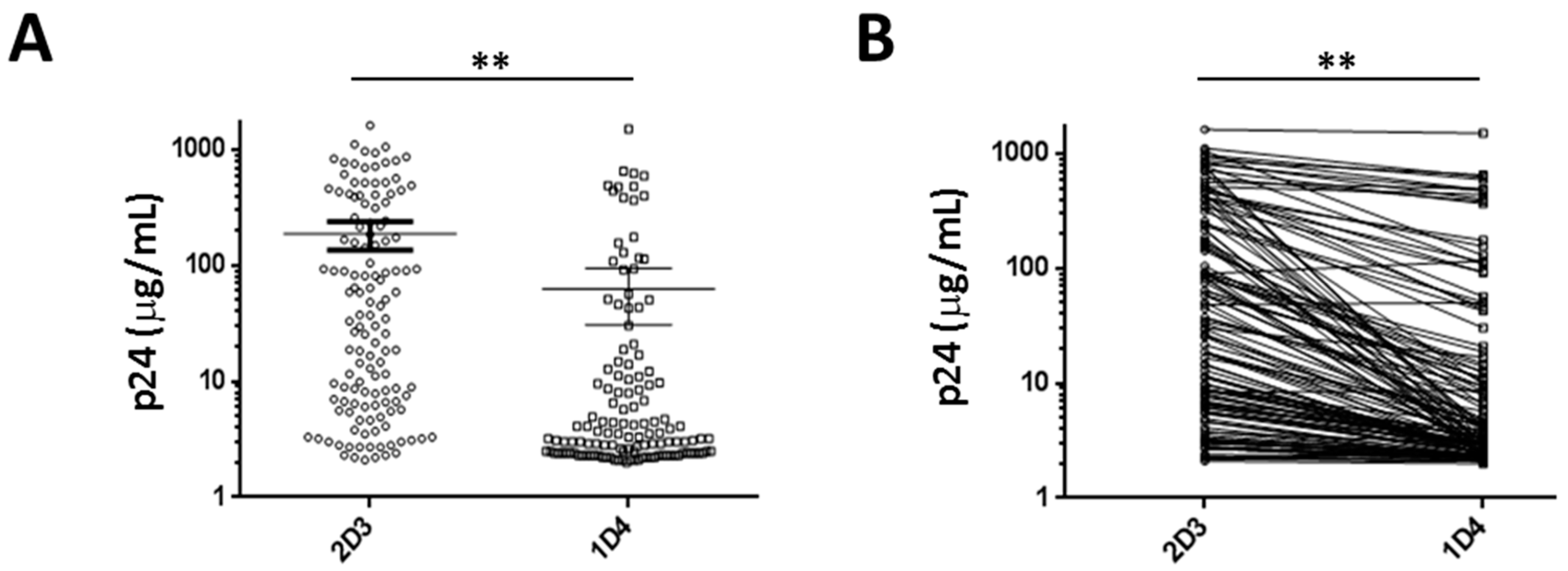
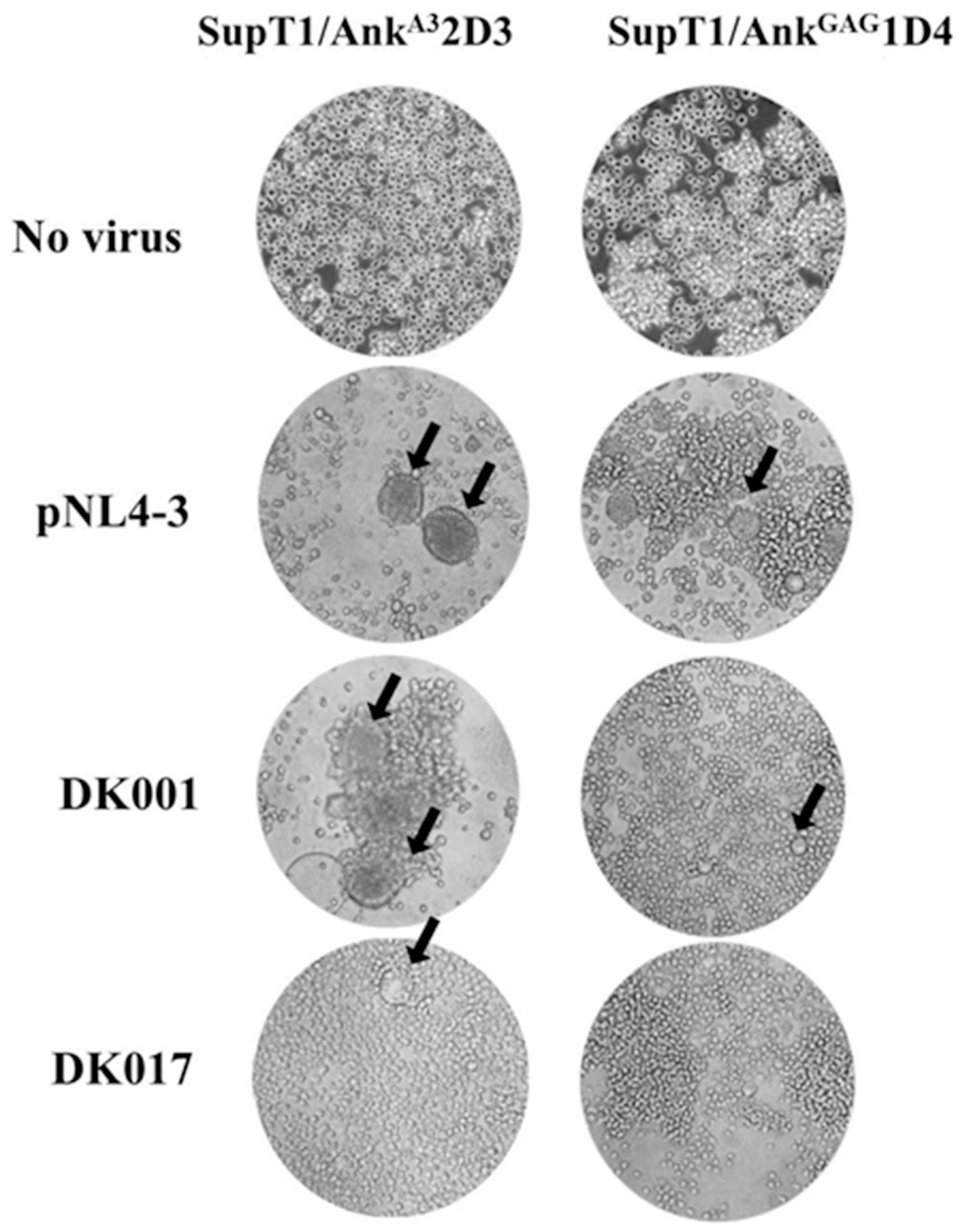
3.2. Effect of AnkGAG1D4 Against Gag/PR Chimeric Viruses
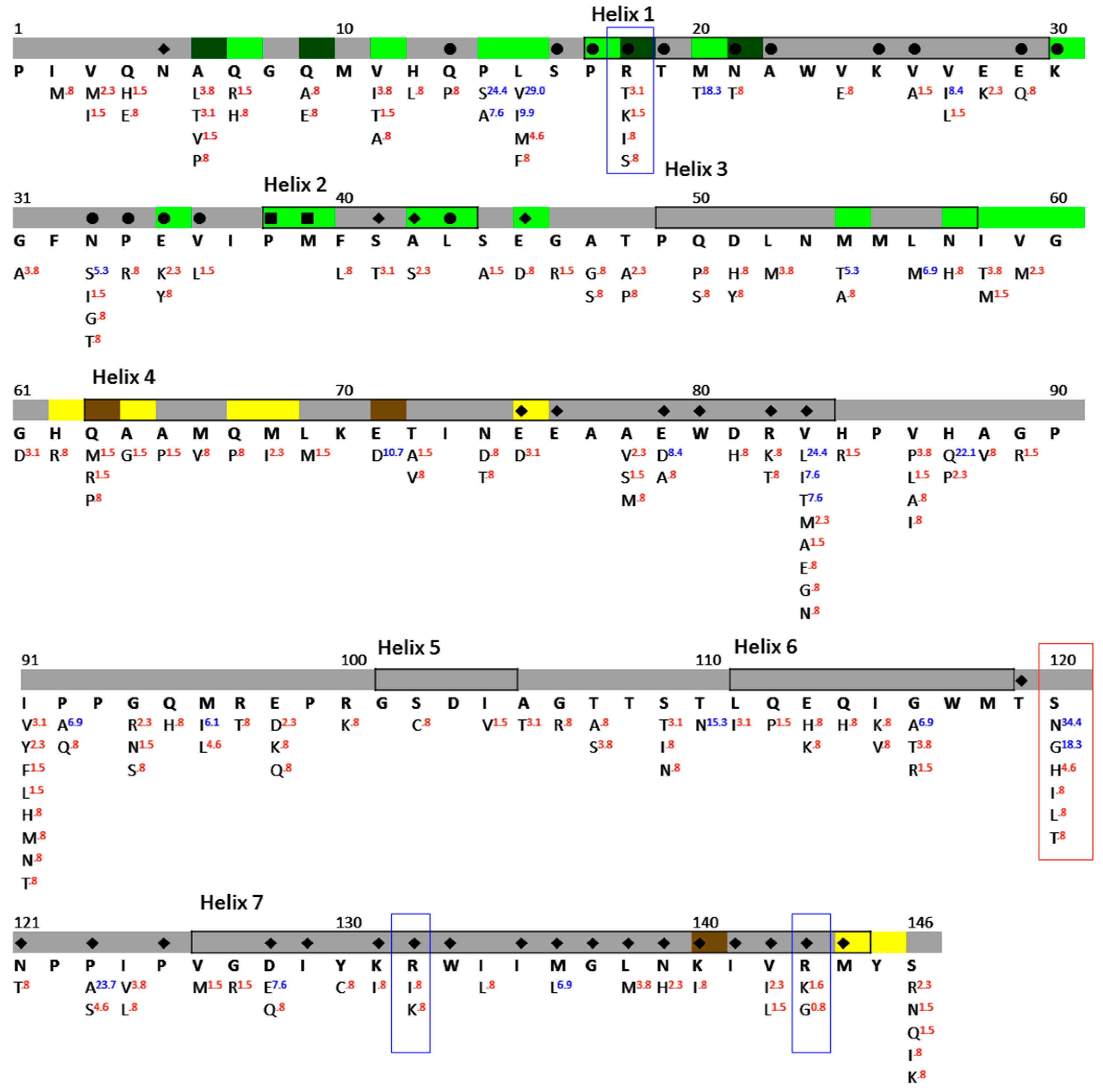
3.3. Sequence Identity of NTDp24CA
3.4. NTDp24CA Amino Acid Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lemke, C.T.; Titolo, S.; von Schwedler, U.; Goudreau, N.; Mercier, J.F.; Wardrop, E.; Faucher, A.M.; Coulombe, R.; Banik, S.S.; Fader, L.; et al. Distinct effects of two HIV-1 capsid assembly inhibitor families that bind the same site within the N-terminal domain of the viral CA protein. J. Virol. 2012, 86, 6643–6655. [Google Scholar] [CrossRef] [PubMed]
- Montessori, V.; Press, N.; Harris, M.; Akagi, L.; Montaner, J.S. Adverse effects of antiretroviral therapy for HIV infection. Can. Med. Assoc. J. 2004, 170, 229–238. [Google Scholar]
- Spreen, W.R.; Margolis, D.A.; Pottage, J.C., Jr. Long-acting injectable antiretrovirals for HIV treatment and prevention. Curr. Opin. HIV AIDS 2013, 8, 565–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolis, D.M.; Koup, R.A.; Ferrari, G. HIV antibodies for treatment of HIV infection. Immunol. Rev. 2017, 275, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.K. Pharmacokinetic and pharmacodynamic considerations for the next generation protein therapeutics. J. Pharmacokinet. Biopharm. 2015, 42, 553–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leader, B.; Baca, Q.J.; Golan, D.E. Protein therapeutics: A summary and pharmacological classification. Nat. Rev. Drug Discov. 2008, 7, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.; Wurch, T.; Bailly, C.; Corvaia, N. Strategies and challenges for the next generation of therapeutic antibodies. Nat. Rev. Immunol. 2010, 10, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Chupradit, K.; Moonmuang, S.; Nangola, S.; Kitidee, K.; Yasamut, U.; Mougel, M.; Tayapiwatana, C. Current Peptide and Protein Candidates Challenging HIV Therapy beyond the Vaccine Era. Viruses 2017, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Stumpp, M.T.; Binz, H.K.; Amstutz, P. DARPins: A new generation of protein therapeutics. Drug Discov. Today 2008, 13, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Simeon, R.; Chen, Z. In vitro-engineered non-antibody protein therapeutics. Protein Cell 2018, 9, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Khamaikawin, W.; Saoin, S.; Nangola, S.; Chupradit, K.; Sakkhachornphop, S.; Hadpech, S.; Onlamoon, N.; Ansari, A.A.; Byrareddy, S.N.; Boulanger, P.; et al. Combined Antiviral Therapy Using Designed Molecular Scaffolds Targeting Two Distinct Viral Functions, HIV-1 Genome Integration and Capsid Assembly. Mol. Ther. Nucleic Acids 2015, 4, e249. [Google Scholar] [CrossRef] [PubMed]
- Nangola, S.; Urvoas, A.; Valerio-Lepiniec, M.; Khamaikawin, W.; Sakkhachornphop, S.; Hong, S.S.; Boulanger, P.; Minard, P.; Tayapiwatana, C. Antiviral activity of recombinant ankyrin targeted to the capsid domain of HIV-1 Gag polyprotein. Retrovirology 2012, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reichen, C.; Hansen, S.; Pluckthun, A. Modular peptide binding: From a comparison of natural binders to designed armadillo repeat proteins. J. Struct. Biol. 2014, 185, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, M.; Skerra, A. Engineered protein scaffolds as next-generation antibody therapeutics. Curr. Opin. Chem. Biol. 2009, 13, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Guellouz, A.; Valerio-Lepiniec, M.; Urvoas, A.; Chevrel, A.; Graille, M.; Fourati-Kammoun, Z.; Desmadril, M.; van Tilbeurgh, H.; Minard, P. Selection of specific protein binders for pre-defined targets from an optimized library of artificial helicoidal repeat proteins (alphaRep). PLoS ONE 2013, 8, e71512. [Google Scholar] [CrossRef] [PubMed]
- Weidle, U.H.; Auer, J.; Brinkmann, U.; Georges, G.; Tiefenthaler, G. The emerging role of new protein scaffold-based agents for treatment of cancer. Cancer Genom. Proteom. 2013, 10, 155–168. [Google Scholar]
- Smithwick, E.; Stewart, M.W. Designed Ankyrin Repeat Proteins: A Look at their Evolving Use in Medicine with a Focus on the Treatment of Chorioretinal Vascular Disorders. Antiinflamm. Antiallergy Agents Med. Chem. 2017, 16, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Campochiaro, P.A.; Channa, R.; Berger, B.B.; Heier, J.S.; Brown, D.M.; Fiedler, U.; Hepp, J.; Stumpp, M.T. Treatment of diabetic macular edema with a designed ankyrin repeat protein that binds vascular endothelial growth factor: A phase I/II study. Am. J. Ophthalmol. 2013, 155, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Souied, E.H.; Devin, F.; Mauget-Faysse, M.; Kolar, P.; Wolf-Schnurrbusch, U.; Framme, C.; Gaucher, D.; Querques, G.; Stumpp, M.T.; Wolf, S.; et al. Treatment of exudative age-related macular degeneration with a designed ankyrin repeat protein that binds vascular endothelial growth factor: A phase I/II study. Am. J. Ophthalmol. 2014, 158, 724–732.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langs-Barlow, A.; Paintsil, E. Impact of human immunodeficiency virus type-1 sequence diversity on antiretroviral therapy outcomes. Viruses 2014, 6, 3855–3872. [Google Scholar] [CrossRef] [PubMed]
- Tovanabutra, S.; Kijak, G.H.; Beyrer, C.; Gammon-Richardson, C.; Sakkhachornphop, S.; Vongchak, T.; Jittiwutikarn, J.; Razak, M.H.; Sanders-Buell, E.; Robb, M.L.; et al. Identification of CRF34_01B, a second circulating recombinant form unrelated to and more complex than CRF15_01B, among injecting drug users in northern Thailand. AIDS Res. Hum. Retrovir. 2007, 23, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Tovanabutra, S.; Watanaveeradej, V.; Viputtikul, K.; De Souza, M.; Razak, M.H.; Suriyanon, V.; Jittiwutikarn, J.; Sriplienchan, S.; Nitayaphan, S.; Benenson, M.W.; et al. A new circulating recombinant form, CRF15_01B, reinforces the linkage between IDU and heterosexual epidemics in Thailand. AIDS Res. Hum. Retrovir. 2003, 19, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Viputtijul, K.; de Souza, M.; Trichavaroj, R.; Carr, J.K.; Tovanabutra, S.; McCutchan, F.E.; Sriplienchan, S.; Buapunth, P.; Chuenchitra, C.; McNeil, J.G.; et al. Heterosexually acquired CRF01_AE/B recombinant HIV type 1 found in Thailand. AIDS Res. Hum. Retrovir. 2002, 18, 1235–1237. [Google Scholar] [CrossRef] [PubMed]
- Phanuphak, N.; Lo, Y.R.; Shao, Y.; Solomon, S.S.; O’Connell, R.J.; Tovanabutra, S.; Chang, D.; Kim, J.H.; Excler, J.L. HIV Epidemic in Asia: Implications for HIV Vaccine and Other Prevention Trials. AIDS Res. Hum. Retrovir. 2015, 31, 1060–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praditwongwan, W.; Chuankhayan, P.; Saoin, S.; Wisitponchai, T.; Lee, V.S.; Nangola, S.; Hong, S.S.; Minard, P.; Boulanger, P.; Chen, C.-J. Crystal structure of an antiviral ankyrin targeting the HIV-1 capsid and molecular modeling of the ankyrin-capsid complex. J. Comput. Aided Mol. Des. 2014, 28, 869–884. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Brockman, M.A.; Brumme, Z.L.; Brumme, C.J.; Pereyra, F.; Trocha, A.; Block, B.L.; Schneidewind, A.; Allen, T.M.; Heckerman, D.; et al. HLA-associated alterations in replication capacity of chimeric NL4-3 viruses carrying gag-protease from elite controllers of human immunodeficiency virus type 1. J. Virol. 2009, 83, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Seber, G.A.F. Multivariate Observations; Wiley: New York, NY, USA, 1984. [Google Scholar]
- Spath, H. The Cluster Dissection and Analysis Theory FORTRAN Programs Examples; Prentice-Hall, Inc.: Upper Saddle River, NJ, USA, 1985. [Google Scholar]
- Castro-Nallar, E.; Pérez-Losada, M.; Burton, G.F.; Crandall, K.A. The evolution of HIV: Inferences using phylogenetics. Mol. Phylogenet. Evol. 2012, 62, 777–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clutter, D.S.; Jordan, M.R.; Bertagnolio, S.; Shafer, R.W. HIV-1 drug resistance and resistance testing. Infect. Genet. Evol. 2016, 46, 292–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, W.S.; Pickford, C.; Irving, S.L.; Brown, D.G.; Anderson, M.; Bazin, R.; Cao, J.; Ciaramella, G.; Isaacson, J.; Jackson, L. HIV capsid is a tractable target for small molecule therapeutic intervention. PLoS Pathog. 2010, 6, e1001220. [Google Scholar] [CrossRef] [PubMed]
- Goudreau, N.; Lemke, C.T.; Faucher, A.-M.; Grand-Maître, C.; Goulet, S.; Lacoste, J.-E.; Rancourt, J.; Malenfant, E.; Mercier, J.-F.O.; Titolo, S. Novel inhibitor binding site discovery on HIV-1 capsid N-terminal domain by NMR and X-ray crystallography. ACS Chem. Biol. 2013, 8, 1074–1082. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Loeliger, E.; Kinde, I.; Kyere, S.; Mayo, K.; Barklis, E.; Sun, Y.; Huang, M.; Summers, M.F. Antiviral inhibition of the HIV-1 capsid protein. J. Mol. Biol. 2003, 327, 1013–1020. [Google Scholar] [CrossRef]
- Sticht, J.; Humbert, M.; Findlow, S.; Bodem, J.; Müller, B.; Dietrich, U.; Werner, J.; Kräusslich, H.-G. A peptide inhibitor of HIV-1 assembly in vitro. Nat. Struct. Mol. Biol. 2005, 12, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Tedbury, P.R.; Freed, E.O. HIV-1 gag: An emerging target for antiretroviral therapy. Curr. Top. Microbiol. Immunol. 2015, 389, 171–201. [Google Scholar] [PubMed]
- Ganser-Pornillos, B.K.; Yeager, M.; Sundquist, W.I. The structural biology of HIV assembly. Curr. Opin. Struct. Biol. 2008, 18, 203–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, N.M.; Lever, A.M. HIV Gag polyprotein: Processing and early viral particle assembly. Trends Microbiol. 2013, 21, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Perilla, J.R.; Yufenyuy, E.L.; Meng, X.; Chen, B.; Ning, J.; Ahn, J.; Gronenborn, A.M.; Schulten, K.; Aiken, C. Mature HIV-1 capsid structure by cryo-electron microscopy and all-atom molecular dynamics. Nature 2013, 497, 643–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Verheyen, J.; Rhee, S.-Y.; Voet, A.; Vandamme, A.-M.; Theys, K. Functional conservation of HIV-1 Gag: Implications for rational drug design. Retrovirology 2013, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- Rihn, S.J.; Wilson, S.J.; Loman, N.J.; Alim, M.; Bakker, S.E.; Bhella, D.; Gifford, R.J.; Rixon, F.J.; Bieniasz, P.D. Extreme genetic fragility of the HIV-1 capsid. PLoS Pathog. 2013, 9, e1003461. [Google Scholar] [CrossRef] [PubMed]
- Nityanandam, R.; Serra-Moreno, R. BCA2/Rabring7 targets HIV-1 Gag for lysosomal degradation in a tetherin-independent manner. PLoS Pathog. 2014, 10, e1004151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goverdhana, S.; Puntel, M.; Xiong, W.; Zirger, J.M.; Barcia, C.; Curtin, J.F.; Soffer, E.B.; Mondkar, S.; King, G.D.; Hu, J.; et al. Regulatable gene expression systems for gene therapy applications: Progress and future challenges. Mol. Ther. 2005, 12, 189–211. [Google Scholar] [CrossRef] [PubMed]
- Moonmuang, S.; Saoin, S.; Chupradit, K.; Sakkhachornphop, S.; Israsena, N.; Rungsiwiwut, R.; Tayapiwatana, C. Modulated expression of the HIV-1 2LTR zinc finger efficiently interferes with the HIV integration process. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.C.; Abraha, A.; Collins, K.R.; Marozsan, A.J.; Baird, H.; Quinones-Mateu, M.E.; Penn-Nicholson, A.; Murray, M.; Richard, N.; Lobritz, M.; et al. Comparing the ex vivo fitness of CCR5-tropic human immunodeficiency virus type 1 isolates of subtypes B and C. J. Virol. 2003, 77, 1021–1038. [Google Scholar] [CrossRef] [PubMed]
- Marozsan, A.J.; Moore, D.M.; Lobritz, M.A.; Fraundorf, E.; Abraha, A.; Reeves, J.D.; Arts, E.J. Differences in the fitness of two diverse wild-type human immunodeficiency virus type 1 isolates are related to the efficiency of cell binding and entry. J. Virol. 2005, 79, 7121–7134. [Google Scholar] [CrossRef] [PubMed]
- Rangel, H.R.; Weber, J.; Chakraborty, B.; Gutierrez, A.; Marotta, M.L.; Mirza, M.; Kiser, P.; Martinez, M.A.; Este, J.A.; Quinones-Mateu, M.E. Role of the human immunodeficiency virus type 1 envelope gene in viral fitness. J. Virol. 2003, 77, 9069–9073. [Google Scholar] [CrossRef] [PubMed]
- Ganser-Pornillos, B.K.; Von Schwedler, U.K.; Stray, K.M.; Aiken, C.; Sundquist, W.I. Assembly properties of the human immunodeficiency virus type 1 CA protein. J. Virol. 2004, 78, 2545–2552. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | AA Position | Sequence Identity (%) | Individual | p24CA Level (µg/mL) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ccAA | IS#1 | IS#2 | IS#3 | IS#4 | IS#5 | AnkA32D3 | AnkGAG1D4 | |||
| IS#1 | - | 100 | 100 | 99.3 | 99.3 | 97.9 | 97.3 | 015F | 3.5 | 2.3 |
| UU53 | 937.5 | 479.3 | ||||||||
| IS#2 | L15V | 99.3 | 99.3 | 100 | 98.6 | 98.6 | 97.9 | UU51 | 92.4 | 14.8 |
| UU23 | 511.1 | 440.1 | ||||||||
| IS#3 | S120N | 99.3 | 99.3 | 98.6 | 100 | 97.2 | 97.3 | 004F | 58.6 | 4.9 |
| 025M | 2.3 | 2.1 | ||||||||
| 041F | 6.2 | 2.2 | ||||||||
| 044M | 18.7 | 2.9 | ||||||||
| 046F | 563.9 | 487.4 | ||||||||
| 049M | 3.3 | 2.4 | ||||||||
| DK056 | 148.8 | 8.4 | ||||||||
| UU06 | 6.4 | 3.1 | ||||||||
| IS#4 | L15V, E79D, V83L | 97.9 | 97.9 | 98.6 | 97.2 | 100 | 96.6 | DK029 | 8.1 | 2.4 |
| DK063 | 64 | 11.1 | ||||||||
| IS#5 | L15V, V83L, G91N, P123S | 97.3 | 96.6 | 97.9 | 97.9 | 97.3 | 100 | DK001 | 25.4 | 2.3 |
| DK002 | 4.1 | 2.2 | ||||||||
| NTDp24CA # | 18 | 132 | 143 |
|---|---|---|---|
| Residue | R | R | R |
| 012M | T | - | - |
| DK064 | T | - | - |
| DK007 | T | - | - |
| DK050 | T | - | - |
| DK051 | K | - | - |
| UU08 | K | - | - |
| UU01 | S | - | - |
| 040F | I | I | - |
| DK032 | - | K | K |
| UU27 | - | - | G |
| UU049 | - | - | K |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakkhachornphop, S.; Hadpech, S.; Wisitponchai, T.; Panto, C.; Kantamala, D.; Utaipat, U.; Praparattanapan, J.; Kotarathitithum, W.; Taejaroenkul, S.; Yasamut, U.; et al. Broad-Spectrum Antiviral Activity of an Ankyrin Repeat Protein on Viral Assembly against Chimeric NL4-3 Viruses Carrying Gag/PR Derived from Circulating Strains among Northern Thai Patients. Viruses 2018, 10, 625. https://doi.org/10.3390/v10110625
Sakkhachornphop S, Hadpech S, Wisitponchai T, Panto C, Kantamala D, Utaipat U, Praparattanapan J, Kotarathitithum W, Taejaroenkul S, Yasamut U, et al. Broad-Spectrum Antiviral Activity of an Ankyrin Repeat Protein on Viral Assembly against Chimeric NL4-3 Viruses Carrying Gag/PR Derived from Circulating Strains among Northern Thai Patients. Viruses. 2018; 10(11):625. https://doi.org/10.3390/v10110625
Chicago/Turabian StyleSakkhachornphop, Supachai, Sudarat Hadpech, Tanchanok Wisitponchai, Chansunee Panto, Doungnapa Kantamala, Utaiwan Utaipat, Jutarat Praparattanapan, Wilai Kotarathitithum, Sineenart Taejaroenkul, Umpa Yasamut, and et al. 2018. "Broad-Spectrum Antiviral Activity of an Ankyrin Repeat Protein on Viral Assembly against Chimeric NL4-3 Viruses Carrying Gag/PR Derived from Circulating Strains among Northern Thai Patients" Viruses 10, no. 11: 625. https://doi.org/10.3390/v10110625