Estimation and Uncertainty of the Mixing Effects on Scots Pine—European Beech Productivity from National Forest Inventories Data

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
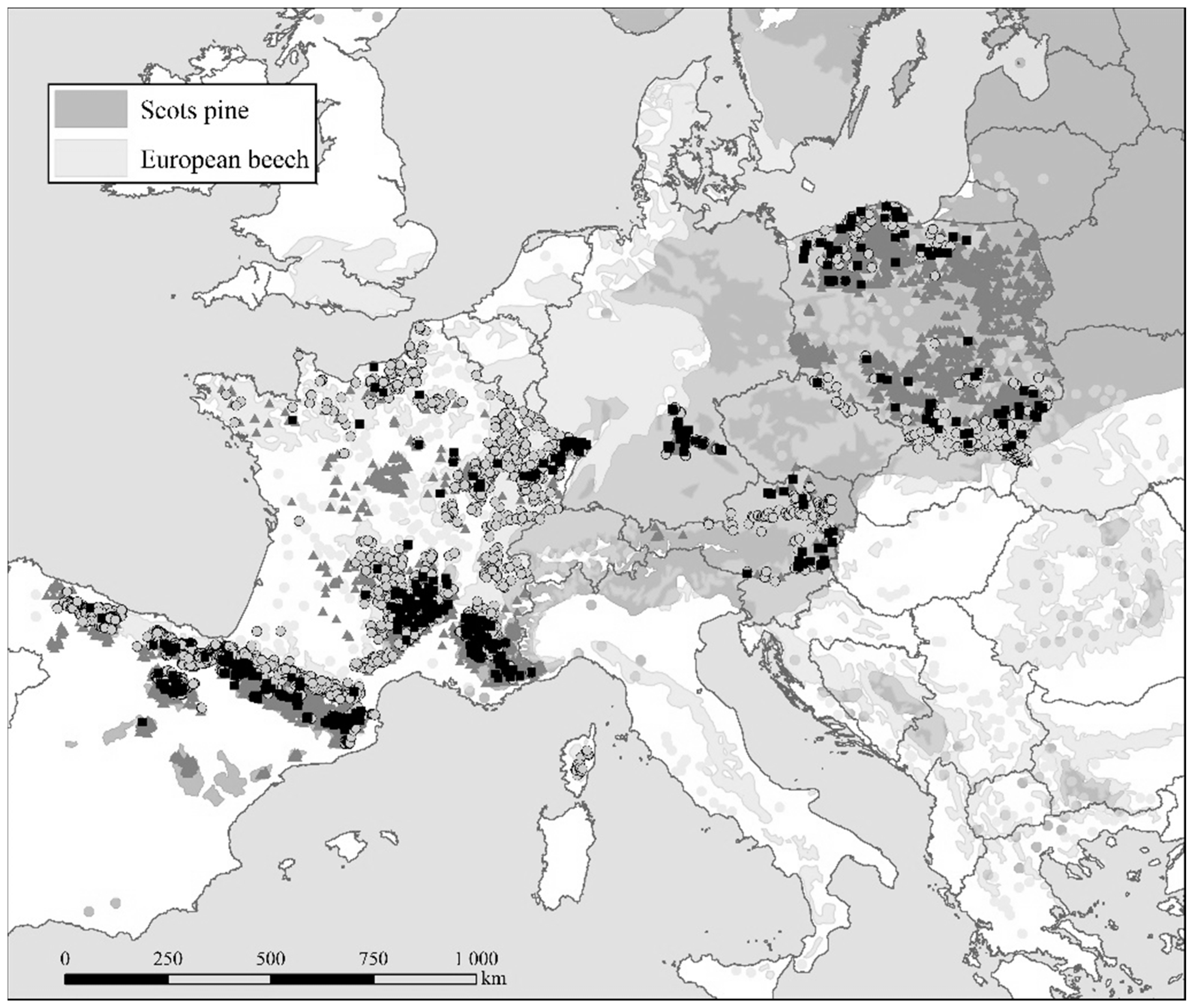
2.1. Data
2.2. Basal Area Growth Efficiency Models
2.2.1. Relative Density and Species Proportions by Area
2.2.2. Model Fitting
2.2.3. Uncertainty Assessment
3. Results
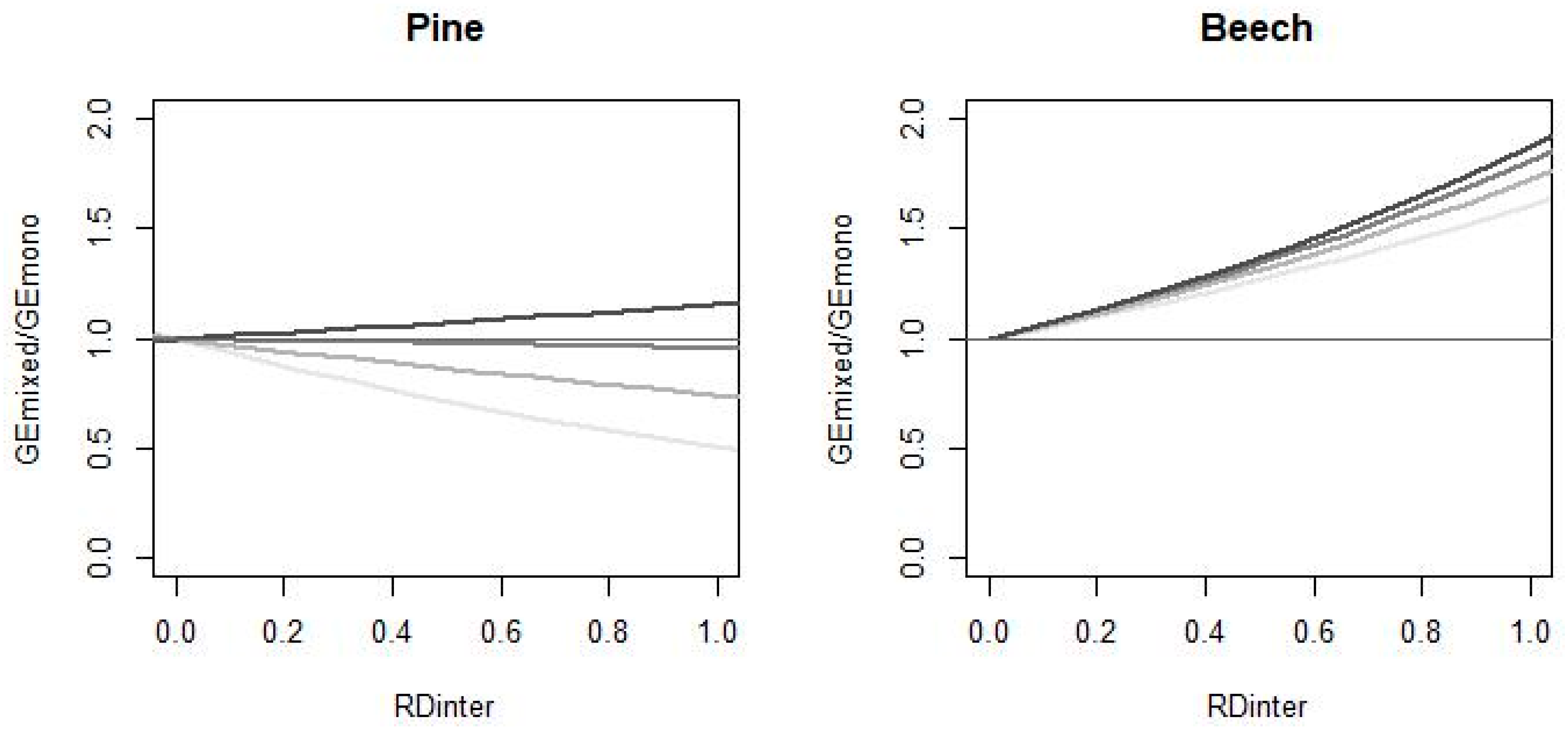
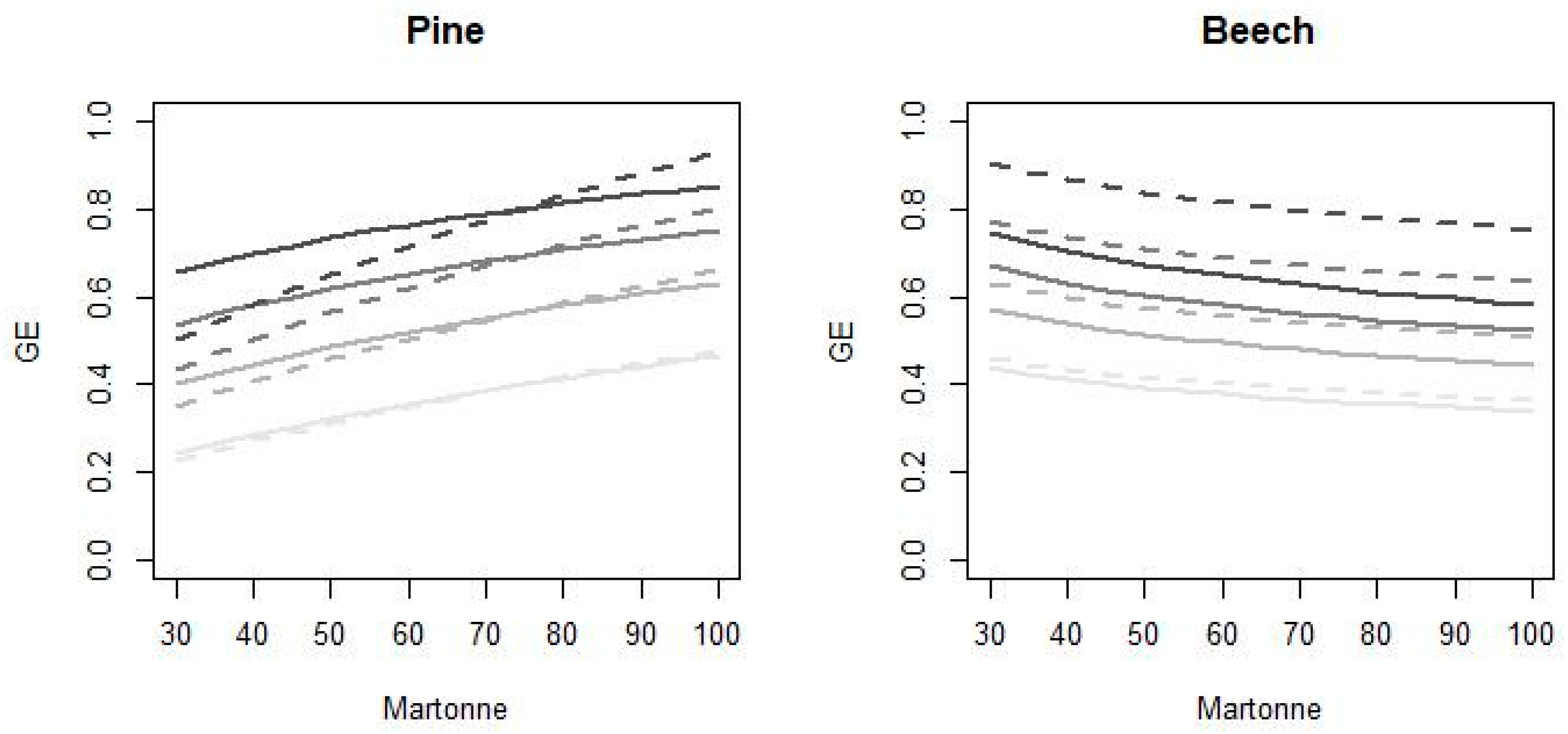
3.1. Mean Mixing Effects on Basal Area Growth
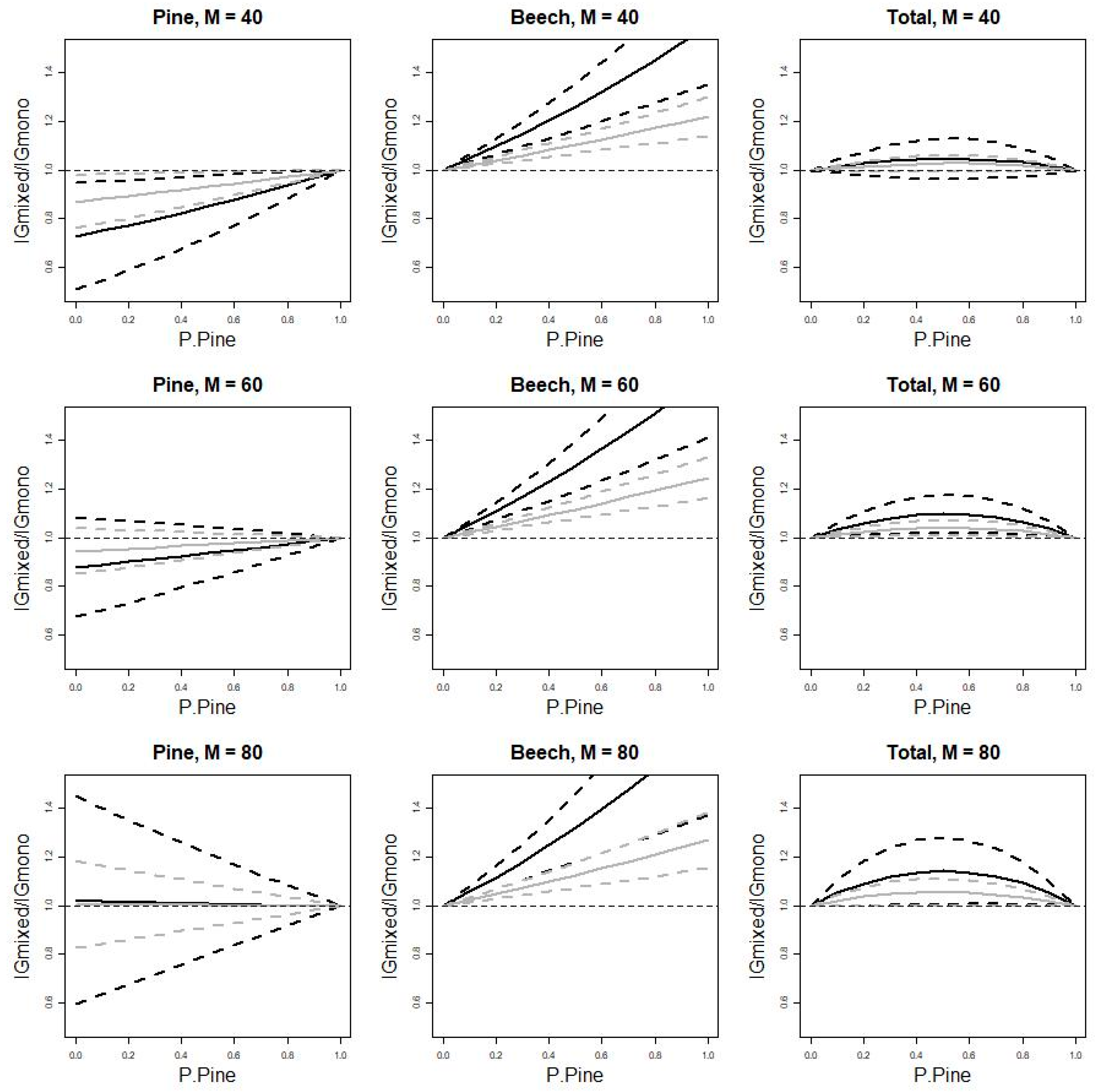
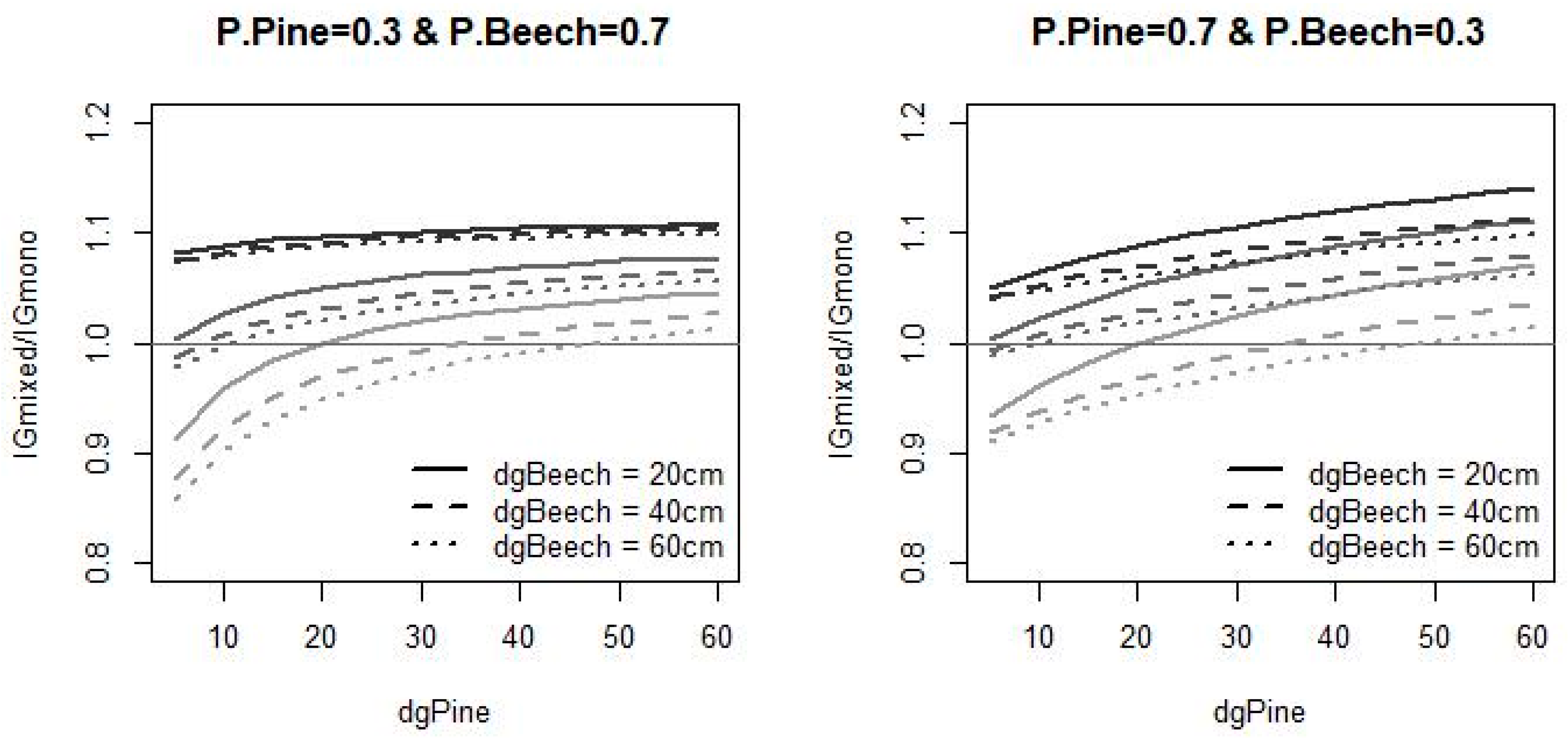
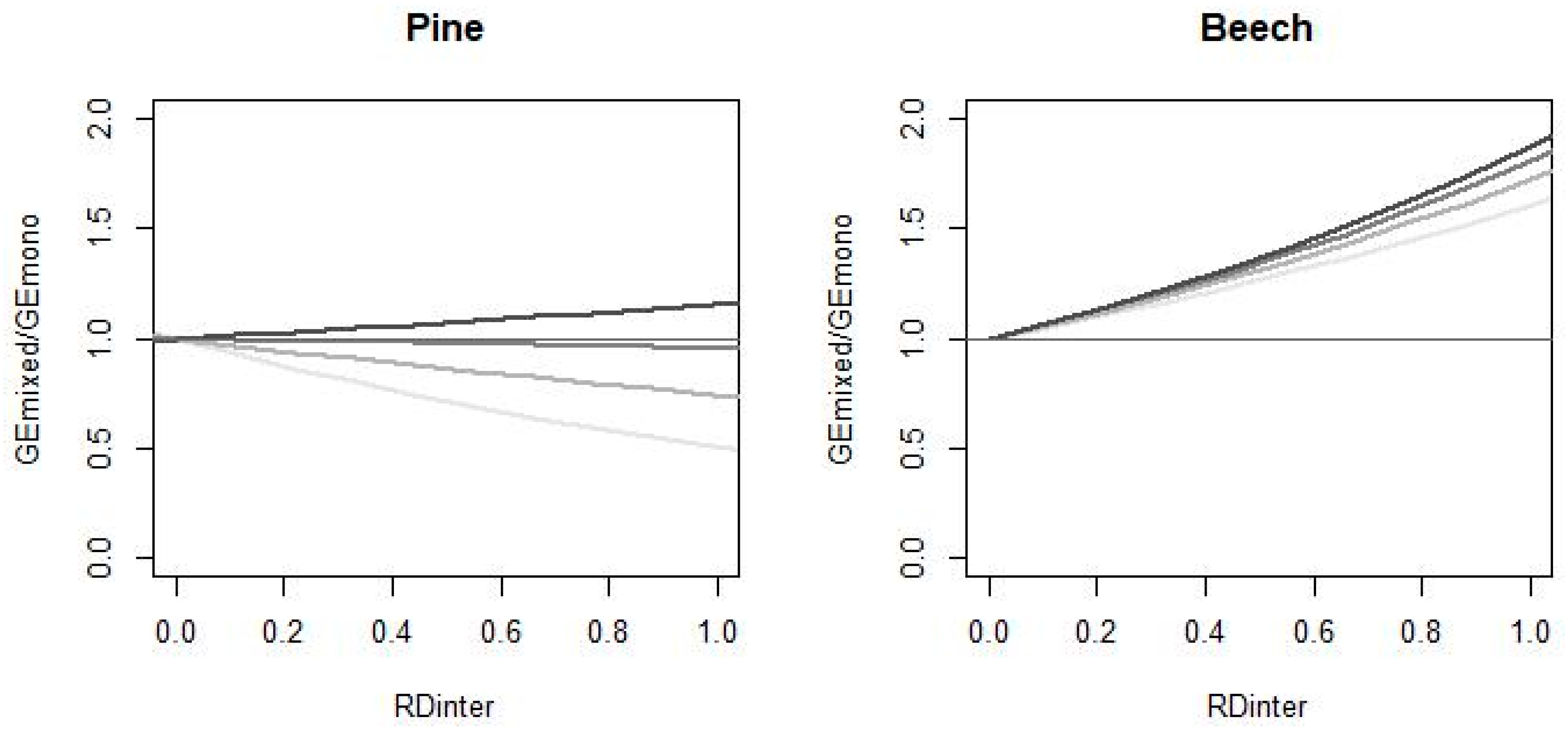
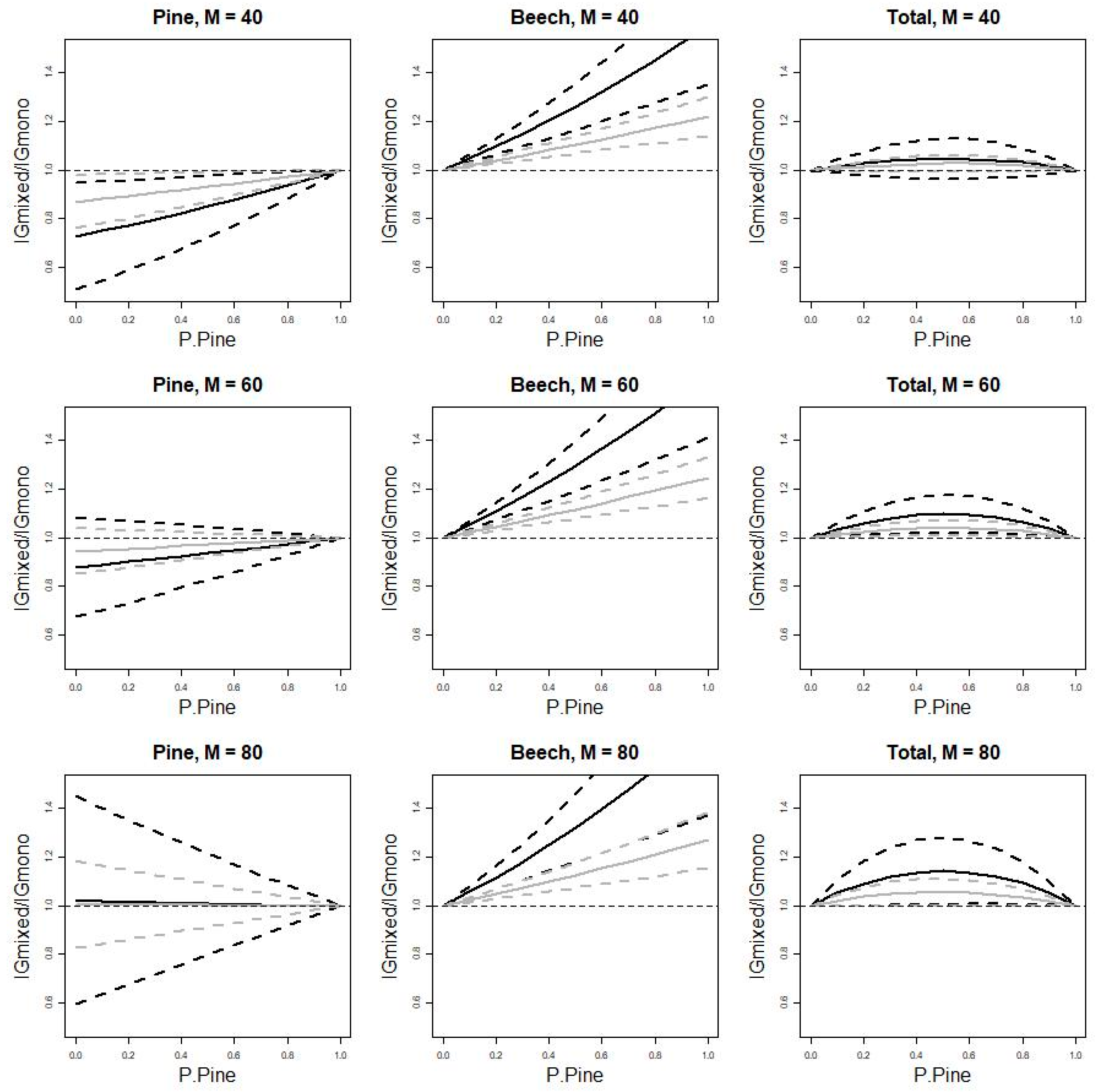
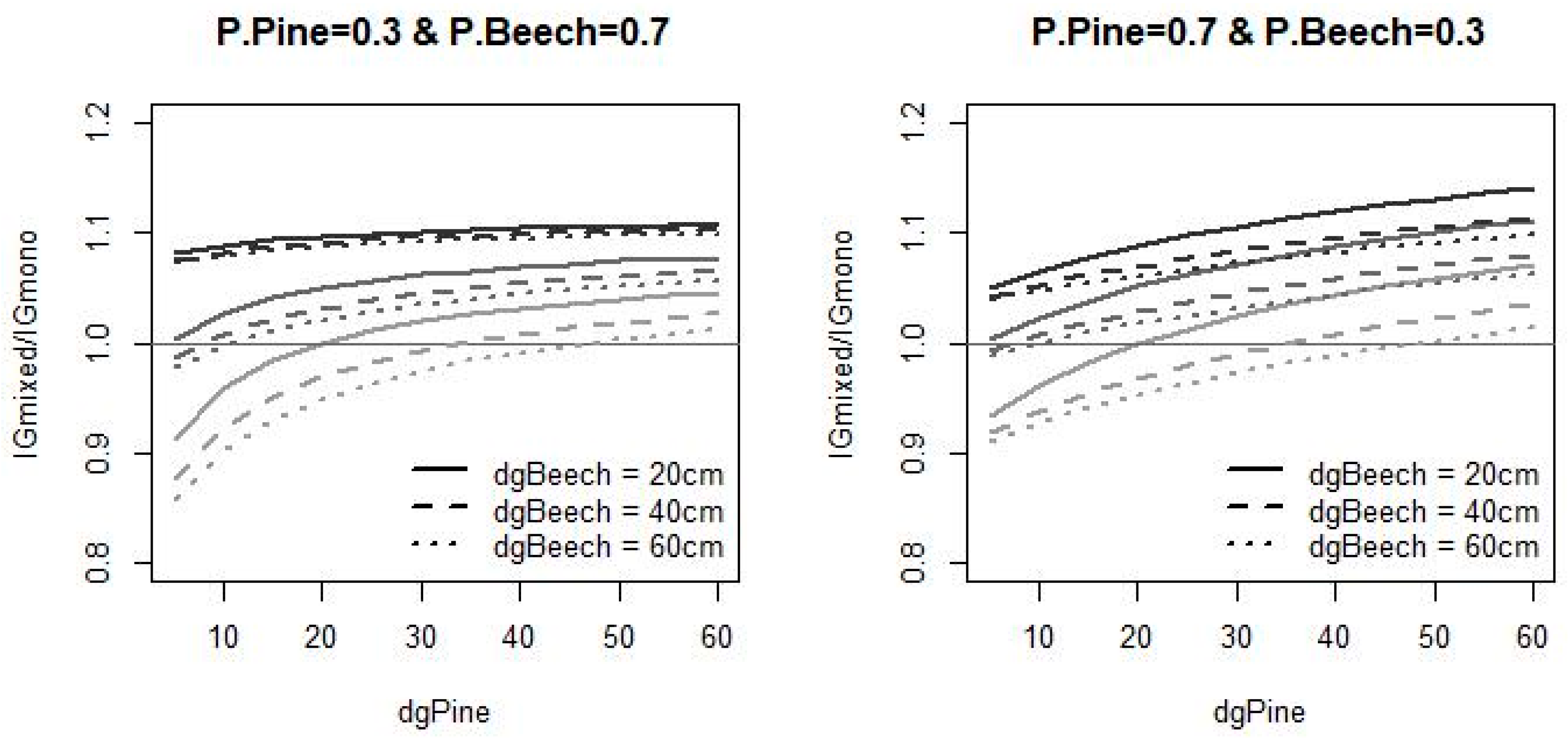
3.2. Uncertainty of Mixing Effects
4. Discussion
4.1. Species Growth Efficiency According to the General Model
4.2. Total Stand Over-Yielding
4.3. Mixing Effect Uncertainty
4.4. Strengths and Limitations of NFI Databases
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cannell, M.G.R.; Malcolm, D.C.; Robertson, P.A. The Ecology of Mixed-Species Stands of Trees; Blackwell Scientific Publications: Oxford, UK, 1992. [Google Scholar]
- Griess, V.C.; Acevedo, R.; Härtl, F.; Staupendahl, K.; Knoke, T. Does mixing tree species enhance stand resistance against natural hazards? A case study for spruce. For. Ecol. Manag. 2012, 267, 284–296. [Google Scholar] [CrossRef]
- Jactel, H.; Nicoll, B.C.; Branco, M.; Gonzalez-Olabarria, J.R.; Grodzki, W.; Långström, B.; Moreira, F.; Netherer, S.; Orazio, C.; Piou, D. The influences of forest stand management on biotic and abiotic risks of damage. Ann. For. Sci. 2009, 66, 1–18. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snäll, T.; Bagchi, R.; Jonsson, M.; Gustafsson, L.; Kjellander, P.; Ruiz-Jaen, M.C.; Fröberg, M.; Stendahl, J.; Philipson, C.D. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2013, 4, 1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brockerhoff, E.G.; Barbaro, L.; Castagneyrol, B.; Forrester, D.I.; Gardiner, B.; González-Olabarria, J.R.; Lyver, P.O.B.; Meurisse, N.; Oxbrough, A.; Taki, H.; et al. Forest biodiversity, ecosystem functioning and the provision of ecosystem services. Biodiv. Conserv. 2017, 26, 3005–3035. [Google Scholar] [CrossRef] [Green Version]
- Vilà, M.; Vayreda, J.; Comas, L.; Ibáñez, J.J.; Mata, T.; Obón, B. Species richness and wood production: A positive association in mediterranean forests. Ecol. Lett. 2007, 10, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Vayreda, J.; Gracia, C.; Ibáñez, J.J. Does tree diversity increase wood production in pine forests? Oecologia 2003, 135, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Global Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F.; Pretzsch, H. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 8957. [Google Scholar] [CrossRef] [PubMed]
- Kelty, M.J. Comparative productivity of monocultures and mixed-species stands. In The Ecology and Silviculture of Mixed-Species Forests; Kelty, M.J., Larson, B.C., Oliver, C.D., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1992; Volume 40, pp. 125–141. [Google Scholar]
- Brown, A. Functioning of Mixed-Species Stands at Gisburn, NW England; Blackwell Scientific Publications: London, UK, 1992; pp. 125–150. [Google Scholar]
- Amoroso, M.M.; Turnblom, E.C. Comparing productivity of pure and mixed douglas-fir and western hemlock plantations in the pacific northwest. Can. J. For. Res. 2006, 36, 1484–1496. [Google Scholar] [CrossRef]
- Del Río, M.; Sterba, H. Comparing volume growth in pure and mixed stands of Pinus sylvestris and Quercus pyrenaica. Ann. For. Sci. 2009, 66, 502. [Google Scholar]
- Toïgo, M.; Pérot, T.; Courbaud, B.; Castagneyrol, B.; Gégout, J.C.; Longuetaud, F.; Jactel, H.; Vallet, P. Difference in shade tolerance drives the mixture effect on Oak productivity. J. Ecol. 2018, 106, 1073–1082. [Google Scholar] [CrossRef]
- Del Río, M.; Condés, S.; Sterba, H. Productividad en Masas mixtas vs. Masas puras: Influencia de la Espesura en la Interacción Entre Especies; Congreso Forestal Español, Sociedad Española de Ciencias Forestales: Pontevedra, Spain, 2013. [Google Scholar]
- Bauhus, J.; Van Winden, A.P.; Nicotra, A.B. Aboveground interactions and productivity in mixed-species plantations of acacia mearnsii and eucalyptus globulus. Can. J. For. Res. 2004, 34, 686–694. [Google Scholar] [CrossRef]
- Binkley, D.; Greene, S. Production in Mixtures of Conifers and Red Alder: The Importance of Site Fertility and Stand Age; Ballard, R., Gessel, S., Eds.; International Union of Forestry Research Organizations Symposium on Forest Site and Continuous Productivity; US Department of Agriculture and Forest Service: Portland, OR, USA, 1983; pp. 112–117.
- Waskiewicz, J.; Kenefic, L.; Weiskittel, A.; Seymour, R. Species mixture effects in northern red oak–eastern white pine stands in maine, USA. For. Ecol. Manag. 2013, 298, 71–81. [Google Scholar] [CrossRef]
- Condés, S.; del Río, M.; Sterba, H. Mixing effect on volume growth of Fagus sylvatica and Pinus sylvestris is modulated by stand density. For. Ecol. Manag. 2013, 292, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Condés, S.; del Río, M.; Goudiaby, V.; den Ouden, J.; Mohren, G.M.; Schelhaas, M.-J.; de Waal, R.; Sterck, F.J. Species and soil effects on over-yielding of tree species mixtures in The Netherlands. For. Ecol. Manag. 2018, 409, 105–118. [Google Scholar] [CrossRef]
- Toïgo, M.; Vallet, P.; Perot, T.; Bontemps, J.D.; Piedallu, C.; Courbaud, B. Over-yielding in mixed forests decreases with site productivity. J. Ecol. 2015, 103, 502–512. [Google Scholar] [CrossRef]
- Jactel, H.; Gritti, E.; Drössler, L.; Forrester, D.; Mason, W.; Morin, X.; Pretzsch, H.; Castagneyrol, B. Positive biodiversity–productivity relationships in forests: Climate matters. Biol. Lett. 2018, 14, 20170747. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Houpert, L.; Rohner, B.; Forrester, D.I.; Mina, M.; Huber, M.O. Mixing effects in Norway spruce—European beech stands are modulated by site quality, stand age and moisture availability. Forests 2018, 9, 83. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 1–12. [Google Scholar] [CrossRef]
- Pretzsch, H.; Bielak, K.; Block, J.; Bruchwald, A.; Dieler, J.; Ehrhart, H.-P.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zasada, M. Productivity of mixed versus pure stands of oak (Quercus petraea (Matt.) Liebl. and Quercus robur L.) and European beech (Fagus sylvatica Ll.) along an ecological gradient. Eur. J. For. Res. 2013, 132, 1–18. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I.; Bauhus, J. Mixed-Species Forests; Springer: Berlin, Germany, 2017. [Google Scholar]
- Bielak, K.; Dudzińska, M.; Pretzsch, H. Mixed stands of Scots pine (Pinus sylvestris L.) and Norway spruce (Picea abies (L.) Karst) can be more productive than monocultures. Evidence from over 100 years of observation of long-term experiments. For. Syst. 2014, 23, 573–589. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Pretzsch, H.; Forrester, D.I.; Rötzer, T. Representation of species mixing in forest growth models. A review and perspective. Ecol. Model. 2015, 313, 276–292. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H.; del Río, M.; Ammer, C.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; et al. Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through europe. Eur. J. For. Res. 2015, 134, 927–947. [Google Scholar] [CrossRef] [Green Version]
- Mina, M.; Huber, M.O.; Forrester, D.I.; Thürig, E.; Rohner, B. Multiple factors modulate tree growth complementarity in central European mixed forests. J. Ecol. 2017, 106, 1106–1119. [Google Scholar] [CrossRef]
- Vallet, P. Silver fir stand productivity is enhanced when mixed with norway spruce: Evidence based on large-scale inventory data and a generic modelling approach. J. Veg. Sci. 2011, 22, 932–942. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Heym, M.; Drössler, L.; Condés, S.; Corona, P.; Bravo, F.; Pretzsch, H.; Bravo-Oviedo, A.; del Río, M. Data platforms for mixed forest research: Contributions from the eumixfor network. In Dynamics, Silviculture and Management of Mixed Forests; Bravo-Oviedo, A., Pretzsch, H., del Río, M., Eds.; Springer: Berlin, Germany, 2018. [Google Scholar]
- Vilà, M.; Carrillo-Gavilán, A.; Vayreda, J.; Bugmann, H.; Fridman, J.; Grodzki, W.; Haase, J.; Kunstler, G.; Schelhaas, M.; Trasobares, A. Disentangling biodiversity and climatic determinants of wood production. PLoS ONE 2013, 8, e53530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, D.I.; Ammer, C.; Annighöfer, P.J.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; del Río, M.; Drössler, L. Predicting the spatial and temporal dynamics of species interactions in Fagus sylvatica and Pinus sylvestris forests across Europe. For. Ecol. Manag. 2017, 405, 112–133. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P. Tree species mixing can increase maximum stand density. Can. J. For. Res. 2016, 46, 1179–1193. [Google Scholar] [CrossRef] [Green Version]
- Condés, S.; Vallet, P.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; Ducey, M.J.; Pach, M.; Pretzsch, H.; Sterba, H.; Vayreda, J. Climate influences on the maximum size-density relationship in Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) stands. For. Ecol. Manag. 2017, 385, 295–307. [Google Scholar] [CrossRef]
- De Río, M.; Pretzsch, H.; Alberdi, I.; Bielak, K.; Bravo, F.; Brunner, A.; Condés, S.; Ducey, M.J.; Fonseca, T.; von Lüpke, N.; et al. Characterization of the structure, dynamics, and productivity of mixed-species stands: Review and perspectives. Eur. J. For. Res. 2016, 135, 23–49. [Google Scholar] [CrossRef]
- Sterba, H.; del Río, M.; Brunner, A.; Condés, S. Effect of species proportion definition on the evaluation of growth in pure vs. mixed stands. For. Syst. 2014, 23, 547–559. [Google Scholar] [CrossRef]
- Dirnberger, G.F.; Sterba, H. A comparison of different methods to estimate species proportions by area in mixed stands. For. Syst. 2014, 23, 534–546. [Google Scholar] [CrossRef]
- Dirnberger, G.; Sterba, H.; Condés, S.; Ammer, C.; Annighöfer, P.; Avdagić, A.; Bielak, K.; Brazaitis, G.; Coll, L.; Heym, M. Species proportions by area in mixtures of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.). Eur. J. For. Res. 2017, 136, 171–183. [Google Scholar] [CrossRef]
- Kangas, A.S. Methods for assessing uncertainty of growth and yield predictions. Can. J. For. Res. 1999, 29, 1357–1364. [Google Scholar] [CrossRef]
- Fortin, M.; Robert, N.; Manso, R. Uncertainty assessment of large-scale forest growth predictions based on a transition-matrix model in catalonia. Ann. For. Sci. 2016, 73, 871–883. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Westfall, J.A. Effects of uncertainty in model predictions of individual tree volume on large area volume estimates. For. Sci. 2014, 60, 34–42. [Google Scholar] [CrossRef]
- Brus, D.; Hengeveld, G.; Walvoort, D.; Goedhart, P.; Heidema, A.; Nabuurs, G.; Gunia, K. Statistical mapping of tree species over Europe. Eur. J. For. Res. 2012, 131, 145–157. [Google Scholar] [CrossRef]
- Vospernik, S.; Nothdurft, A. Can trees at high elevations compensate for growth reductions at low elevations due to climate warming? Can. J. For. Res. 2018, 48, 650–662. [Google Scholar] [CrossRef] [Green Version]
- Martonne, E. Une nouvelle fonction climatologique: L’indice d’aridite (a new climatological function: The aridity index). La Météorologie 1926, 2, 449–458. [Google Scholar]
- EUFORGEN. Distribution Map of European Beech (Fagus sylvatica L.) and Scots pine (Pinus sylvestris L.). Available online: http://www.euforgen.org (accessed on 4 August 2018).
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Core Team (2017) Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-131. Available online: http://CRAN. R-project. org/package = nlme (accessed on 4 August 2018).
- Maj, A. Lmmfit: Goodness-of-Fit-Measures for Linear Mixed Models with One-Level-Grouping. R package version 2011, 1. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5426279/ (accessed on 4 August 2018).
- Riofrío, J.; del Río, M.; Bravo, F. Mixing effects on growth efficiency in mixed pine forests. For. Int. J. For. Res. 2017, 90, 381–392. [Google Scholar] [CrossRef]
- Vilà, M.; Inchausti, P.; Vayreda, J.; Barrantes, O.; Gracia, C.; Ibáñez, J.; Mata, T. Confounding factors in the observational productivity-diversity relationship in forests. In Forest Diversity and Function; Springer: Berlin/Heidelberg, Germany, 2005; pp. 65–86. [Google Scholar]
- Pretzsch, H. Stand density and growth of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.): Evidence from long-term experimental plots. Eur. J. For. Res. 2005, 124, 193–205. [Google Scholar] [CrossRef]
- Vanclay, J.K.; Henry, N.B. Assessing site productivity of indigenous cypress pine forest in southern Queensland. Commn. For. Rev. 1988, 67, 53. [Google Scholar]
- Huang, S.; Titus, S.J. An index of site productivity for uneven-aged or mixed-species stands. Can. J. For. Res. 1993, 23, 558–562. [Google Scholar] [CrossRef]
- Meier, E.S.; Edwards, T.C., Jr.; Kienast, F.; Dobbertin, M.; Zimmermann, N.E. Cooccurrence patterns of trees along macroclimatic gradients and their potential influence on the present and future distribution of Fagus sylvatica L. J. Biogeogr. 2011, 38, 371–382. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Smith, K.T.; Čufar, K. Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Meteorol. 2013, 180, 142–151. [Google Scholar] [CrossRef]
- Assmann, E. The principles of yield study. In Studies in the Organic Production, Structure, Increment and Yield of Forest Stands; Oxford Pergamon Press: Oxford, UK, 1970. [Google Scholar]
- Forrester, D.I.; Pretzsch, H. Tamm review: On the strength of evidence when comparing ecosystem functions of mixtures with monocultures. For. Ecol. Manag. 2015, 356, 41–53. [Google Scholar] [CrossRef]
- Garber, S.M.; Maguire, D.A. Stand productivity and development in two mixed-species spacing trials in the central Oregon cascades. For. Sci. 2004, 50, 92–105. [Google Scholar]
- Heym, M.; Ruíz-Peinado, R.; del Río, M.; Bielak, K.; Forrester, D.; Dirnberger, G.; Barbeito, I.; Brazaitis, G.; Ruškytė, I.; Coll, L.; et al. Data from: Eumixfor empirical forest mensuration and ring width data from pure and mixed stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) through Europe. Dryad Digit. Repos. 2017, 10, 5061. [Google Scholar]
- Heym, M.; Ruíz-Peinado, R.; Del Río, M.; Bielak, K.; Forrester, D.I.; Dirnberger, G.; Barbeito, I.; Brazaitis, G.; Ruškytkė, I.; Coll, L.; et al. Eumixfor empirical forest mensuration and ring width data from pure and mixed stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) through europe. Ann. For. Sci. 2017, 74, 63. [Google Scholar] [CrossRef]
- Condés, S.; McRoberts, R.E. Updating national forest inventory estimates of growing stock volume using hybrid inference. For. Ecol. Manag. 2017, 400, 48–57. [Google Scholar] [CrossRef]
- Corona, P.; Fattorini, L.; Franceschi, S.; Scrinzi, G.; Torresan, C. Estimation of standing wood volume in forest compartments by exploiting airborne laser scanning information: Model-based, design-based, and hybrid perspectives. Can. J. For. Res. 2014, 44, 1303–1311. [Google Scholar] [CrossRef]
- McRoberts, R.E.; Chen, Q.; Domke, G.M.; Ståhl, G.; Saarela, S.; Westfall, J.A. Hybrid estimators for mean aboveground carbon per unit area. For. Ecol. Manag. 2016, 378, 44–56. [Google Scholar] [CrossRef]
- Porté, A.; Bartelink, H. Modelling mixed forest growth: A review of models for forest management. Ecol. Model. 2002, 150, 141–188. [Google Scholar] [CrossRef]
- Vidal, C.; Alberdi, I.; Redmond, J.; Vestman, M.; Lanz, A.; Schadauer, K. The role of European national forest inventories for international forestry reporting. Ann. For. Sci. 2016, 73, 793–806. [Google Scholar] [CrossRef]
- Tomppo, E.; Gschwantner, T.; Lawrence, M.; McRoberts, R.E. National Forest Inventories: Pathways for Common Reporting; Springer: Berlin, Germany, 2010; p. 601. [Google Scholar]
- Álvarez-González, J.; Cañellas, I.; Alberdi, I.; Gadow, K.; Ruiz-González, A. National forest inventory and forest observational studies in spain: Applications to forest modeling. For. Ecol. Manag. 2013, 316, 54–64. [Google Scholar] [CrossRef]
- McCullagh, A.; Black, K.; Nieuwenhuis, M. Evaluation of tree and stand-level growth models using national forest inventory data. Eur. J. For. Res. 2017, 136, 1–8. [Google Scholar] [CrossRef]
- Dănescu, A.; Albrecht, A.T.; Bauhus, J. Structural diversity promotes productivity of mixed, uneven-aged forests in southwestern Germany. Oecologia 2016, 182, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Silva Pedro, M.; Rammer, W.; Seidl, R. Disentangling the effects of compositional and structural diversity on forest productivity. J. Veg. Sci. 2017, 28, 649–658. [Google Scholar] [CrossRef]
- Zeller, L.; Liang, J.; Pretzsch, H. Tree species richness enhances stand productivity while stand structure can have opposite effects, based on forest inventory data from Germany and the United States of America. For. Ecosyst. 2018, 5, 4. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pine | Beech | Total | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | dg | Ho | RD | IG | N | dg | Ho | RD | IG | N | dg | Ho | RD | IG | M | |||
| Pine | 8109 plots | Mean | 757 | 23.5 | 18.3 | 0.55 | 0.65 | 794 | 22.9 | 18.3 | 0.56 | 0.67 | 45.2 | |||||
| Sd | 584 | 8.2 | 7.8 | 0.25 | 0.41 | 595 | 7.8 | 7.8 | 0.25 | 0.42 | 11.9 | |||||||
| Min | 28 | 6.8 | 2.8 | 0.10 | 0.01 | 28 | 6.8 | 2.8 | 0.10 | 0.01 | 30.0 | |||||||
| Max | 5612 | 67.1 | 42.1 | 1.87 | 3.48 | 5612 | 61.9 | 42.1 | 1.87 | 3.59 | 99.8 | |||||||
| Mixtures | 540 plots | Mean | 301 | 29.7 | 19.6 | 0.32 | 0.32 | 489 | 23.1 | 17.3 | 0.25 | 0.33 | 838 | 23.6 | 20.4 | 0.58 | 0.67 | 53.6 |
| Sd | 259 | 9.5 | 8.5 | 0.22 | 0.29 | 466 | 14.8 | 6.6 | 0.17 | 0.22 | 551 | 8.3 | 8.2 | 0.25 | 0.36 | 14.9 | ||
| Min | 14 | 5.3 | 2.3 | 0.02 | 0.01 | 5 | 7.2 | 4.6 | 0.02 | 0.00 | 75 | 7.4 | 6.1 | 0.11 | 0.07 | 30.5 | ||
| Max | 2108 | 57.9 | 43.9 | 1.09 | 2.21 | 2865 | 114.8 | 39.3 | 1.02 | 1.19 | 3701 | 56.6 | 43.9 | 1.46 | 3.02 | 99.0 | ||
| Beech | 2460 plots | Mean | 675 | 29.8 | 21.1 | 0.53 | 0.55 | 698 | 29.2 | 21.1 | 0.54 | 0.57 | 64.7 | |||||
| Sd | 679 | 14.9 | 6.9 | 0.25 | 0.32 | 688 | 14.4 | 6.9 | 0.25 | 0.33 | 17.9 | |||||||
| Min | 10 | 5.5 | 5.6 | 0.10 | 0.01 | 10 | 5.5 | 5.6 | 0.10 | 0.01 | 30.0 | |||||||
| Max | 8524 | 155.7 | 47.5 | 1.93 | 2.71 | 8524 | 155.7 | 47.5 | 2.03 | 2.98 | 99.9 | |||||||
| Pine | Beech | |||
|---|---|---|---|---|
| Value | Standard Error | Value | Standard Error | |
| (Intercept) | 0.8392 | 0.2127 | 0.9262 | 0.2010 |
| log(Ho) | 0.2712 | 0.0284 | 0.4887 | 0.0389 |
| log(dgsp) | −0.8089 | 0.0242 | −0.6279 | 0.0250 |
| log(RD) | 1.4873 | 0.1657 | 0.3879 | 0.0216 |
| log(M) | 0.1671 | 0.0470 | −0.2069 | 0.0390 |
| log(M)·log(RD) | −0.2285 | 0.0431 | ns | |
| RDinter | −3.2002 | 1.1075 | ns | |
| log(M)·RDinter | 0.7424 | 0.2811 | 0.1390 | 0.0172 |
| R2 marginal | 0.3711 | 0.3588 | ||
| R2 conditional | 0.4946 | 0.4002 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Condés, S.; Sterba, H.; Aguirre, A.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; Pach, M.; Pretzsch, H.; Vallet, P.; Del Río, M. Estimation and Uncertainty of the Mixing Effects on Scots Pine—European Beech Productivity from National Forest Inventories Data. Forests 2018, 9, 518. https://doi.org/10.3390/f9090518
Condés S, Sterba H, Aguirre A, Bielak K, Bravo-Oviedo A, Coll L, Pach M, Pretzsch H, Vallet P, Del Río M. Estimation and Uncertainty of the Mixing Effects on Scots Pine—European Beech Productivity from National Forest Inventories Data. Forests. 2018; 9(9):518. https://doi.org/10.3390/f9090518
Chicago/Turabian StyleCondés, Sonia, Hubert Sterba, Ana Aguirre, Kamil Bielak, Andrés Bravo-Oviedo, Lluís Coll, Maciej Pach, Hans Pretzsch, Patrick Vallet, and Miren Del Río. 2018. "Estimation and Uncertainty of the Mixing Effects on Scots Pine—European Beech Productivity from National Forest Inventories Data" Forests 9, no. 9: 518. https://doi.org/10.3390/f9090518